Obraz (2029)

Łańcuchy polinukleotydowe mają dwa końce, 3* i 5’. W zapisach sekwencji nukleołydów w kwasach nukleinowych fragment z wolną grupą OH przy C-5* umieszcza się na początku. Sekwencja jest zatem zapisywana w kierunku 5* -* 3'. Dla uproszczenia wymienia się tylko literowe symbole zasad i pomija pozostałe elementy strukturalne, bo są one takie same dla całego łańcucha. Kolejność nukleołydów jest istotna dla odczytywania kodu genetycznego. Na przykład fragment TTAGCC koduje sekwencję LeuAia w łańcuchu peptydo-wym a odwrotny fragment CCGATT jest kodem dla sekwencji Prolle.
Drugo rzędowa struktura DNA
DNA ma budowę podwójnej helisy. Dwa łańcuchy polinukleotydowe, z których jeden biegnie w kierunku 5*-»3’a drugi w odwrotnym kierunku 3’-*5\ są ułożone równolegle i tworzą helisę, jak dwa równoległe druty śrubowo owinięte na walcu. Łańcuchy polinukleotydowe są połączone ze sobą przez wiązania wodorowe, tworzące się między zasadami wypełniającymi wnętrze helisy. Rys. 222 pokazuje schemat budowy podwójnej nici DNA, bez szczegółów przestrzennych. Celem rysunku jest tylko podkreślenie, że grubość podwójnego łańcucha jest taka sama na całej jego długości.
ł o
5'-pentoza-fosforan-pentoza-fosforan-pentoza-fosforan-pŁ'nto/a-3'
2.0 nm
1.1 nm
___f .
3 -pentoza-fosforan-pentoza-fosforan-pentoza-fosforan-pentoza- 5'
Rys. 22.2. Schemat podwójnego łańcucha DNA
W podwójnej helisie zasady purynowe i pirymidynowe, należące do jednego z łańcuchów polinukJeotydowych. są ustawione dokładnie naprzeciw zasad, należących do drugiego łańcucha. Takie ustawienie umożliwia powstawanie wiązań wodorowych, stabilizujących podwójną helisę. Z jednakowej grubości helisy na całej jej długości wynika, że naprzeciw zasad purynowych muszą leżeć zasady pirymidynowe. Zasady purynowe mają znacznie większe rozmiary niż pirymidynowe, a więc ustawienie zasad purynowych naprzeciw siebie wymaga-loby pogrubienia helisy, gdyby zaś naprzeciw siebie znalazły się dwie zasady pirymidynowe, to musiałoby w tym miejscu wystąpić zwężenie.
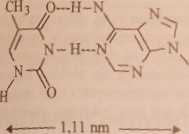
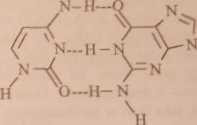
Bliższa analiza wykazuje, że najsilniejsze wiązania wodorowe tworzą się między adeniną (A) i tyminą (T) oraz guaniną (G) i cytozyną (C). A i T oraz G i C nazywamy zasadami komplementarnymi. Tc właśnie zasady leżą naprzeciw siebie wewnątrz podwójnej helisy DNA. Rozmiary układu A-T i G-C (rys. 22.3) dobrze pasują do zmierzonych rentgenograficznie rozmiarów wewnętrznej prze-strzcni w podwójnej helisie.
H
\
/
T - A
1,08 nm ■ ►
C - G
Rys. 22.3. Wiązania wodorowe między komplementarnymi zasadami w DNA
Wymiary par T-G i C-A (pary takie nie występują w kwasach nukleinowych) są w przybliżeniu takie same, jak wymiary par A-T i G-C, ale wiązania wodorowe byłyby znacznie słabsze.
22.5. Kwasy rybonukleinowe (RNA)
Kwasy rybonukleinowe są zbudowane z rybonukleotydów, połączonych wiązaniem fosfodiestrowym. Tak samo jak w DNA, wiązanie to łączy atomy C-3* i C-5’ pentoz. Najłatwiej dostrzegalne różnice między RNA i DNA polegają na tym, że w RNA zamiast tyminy znajduje się uracyl (rozdz. 19.7) a przy atomie C-2’ reszty pentozowej znajduje się grupa OH. Kwasy rybonukleinowe występują w formie jednoniciowej.
W biosyntezie białek uczestniczą trzy rodzaje RNA: informacyjny, transportujący i rybosomalny. Informacja genetyczna, czyli informacja o sekwencji ami-
Wyszukiwarka
Podobne podstrony:
Obraz (2030) 638 nokwasów w białkach, jest zapisana w sekwencji nukleotydów w DNA. W komórkach eukar
Obraz (2030) 638 nokwasów w białkach, jest zapisana w sekwencji nukleotydów w DNA. W komórkach eukar
P4280270 (2) budowa DNA ■ dwa helikalne łańcuchy polinukleotydowe zwijają się
Zdjęcie105 /a Iow w łańcuchu połinuUoaiydo«ym 1 połmuklcGłydiw/e Utrzymywane wtpoinw odry komplement
Skane 4sj łańcuch polinukleotydowy wiązanie 13 — N ^ - glikozydowe Zasada Cukier Kwasy nukleinowe
Tokarki te mają dwa suporty: suport wzdłużny SUP.W (z przodu) do toczenia wzdłużnego, suport poprzec
3 (389) uniemożliwiając swobodny obrót tworzących je atomów. Jeśli atomy mają dwa rodzaje pod stawni
77253 SSL25332 Bacillus ■ Gram+. tlenowe laseczki. komórki są stosunkowo duże i mają kwadratowe końc
więcej podobnych podstron