lastscan4 (9)
larwy węgorza (Leptocephalus) i kilka gatunków ryb antarktycznych z rodziny Chaenichthyidąe.
KRZYWE DYSOCJACJI TLENU
Właściwością krwi o największym znaczeniu dla transportu tlenu jest odwracalne wiązanie tlenu z cząsteczką hemoglobiny. Wiązanie to można przedstawić w postaci zwyczajnej, odwracalnej reakcji chemicznej:
Hb + 02 *-* HbQ2.
Przy dużym stężeniu tlenu hemoglobina (Hb) łączy się z tlenem, tworzgc oksyhemo-globińęTHbÓ2), i reakcją bicgiue w prawo. Przy małym stężeniu tlen Jest oddawany i reakcja przebiega w lewo. Jeśli stężenie tlenu spad nie do jze r a JTi emo glo bi n a odda cały, niesiony przez siebie, tlen.
Każdy atom żelaza w cząsteczce hemoglobiny wiąże jedną cząsteczkę tlenu, a gdy w dużym stężeniu (czyli ciśnieniu) tlenu wszystkie dostępne miejsca wiązania są zajęte, hemoglobina jest całkowicie nasycona i nie może przyjąć więcej tlenu. Dla danego stężenia tlenu istnieje ściśle określona proporcja ilości hemoglobiny do ok-sjTiemoglobinyj
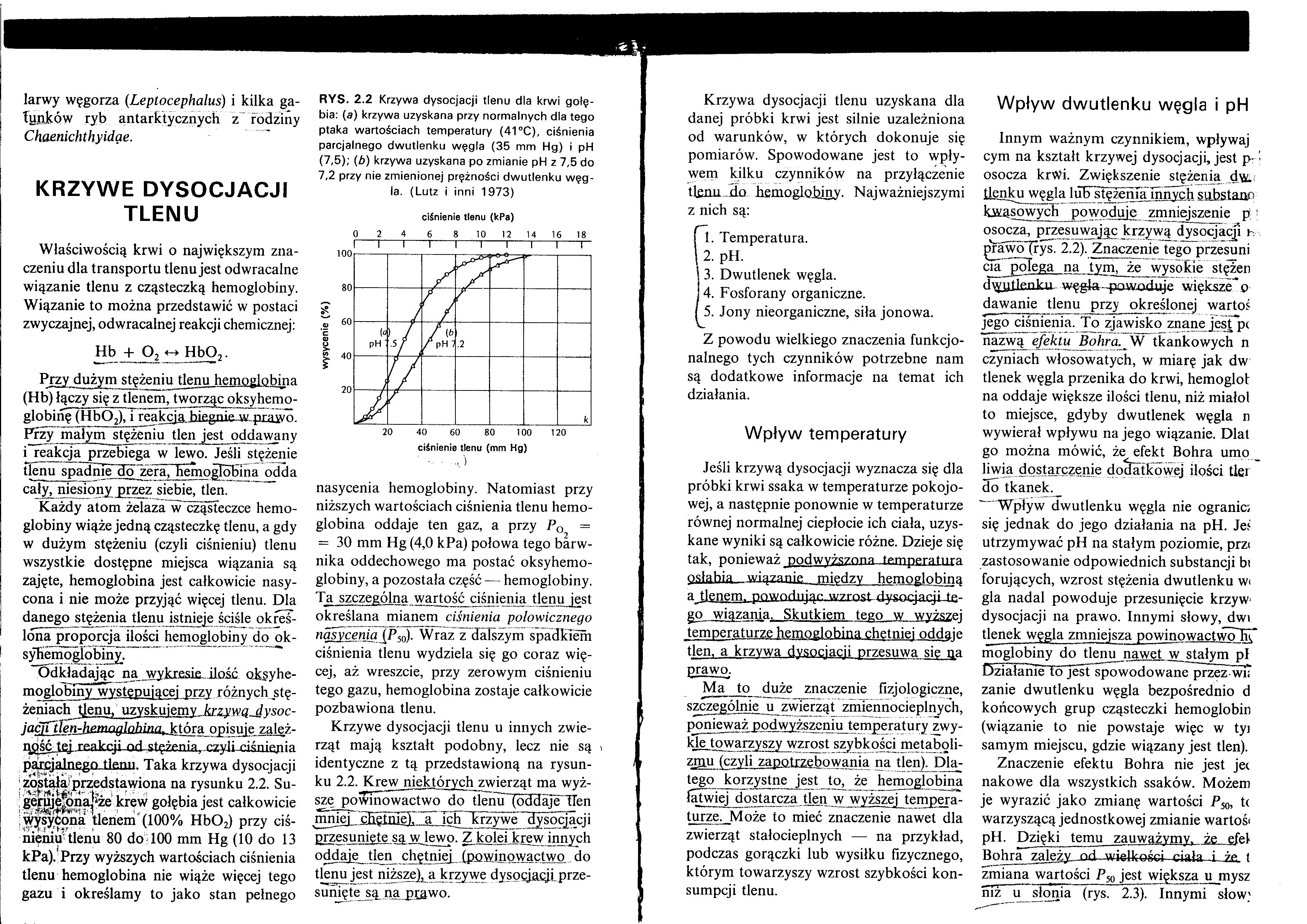
Odkładając na wykresie, ilość oksyhe-moglobiny występującej przy różnych stężeniach tlenu, uzyskujemy krzywa dvsoc-jaćjrUen-henmglnhina, która_opisuje ząleż-gÓŚĆJej reakcji nrl.Łtężenia rr/yli r.iśniepia parcjalnego, tlenu. Taka krzywa dysocjacji została przedstawiona na rysunku 2.2. Su-geruje^ona|‘że krew gołębia jest całkowicie wysycona tlenem (100% Hb02) przy ciśnieniu tlenu 80 do 100 mm Hg (10 do 13 kPa). Przy wyższych wartościach ciśnienia tlenu hemoglobina nie wiąże więcej tego gazu i określamy to jako stan pełnego
RYS. 2.2 Krzywa dysocjacji tlenu dla krwi gołębia: (a) krzywa uzyskana przy normalnych dla tego ptaka wartościach temperatury (41 °C), ciśnienia parcjalnego dwutlenku węgla (35 mm Hg) i pH (7,5); (b) krzywa uzyskana po zmianie pH z 7,5 do 7,2 przy nie zmienionej prężności dwutlenku węgla. (Lutz i inni 1973)
ciśnienie tlenu (kPa)

ciśnienie tlenu (mm Hg)
nasycenia hemoglobiny. Natomiast przy niższych wartościach ciśnienia tlenu hemoglobina oddaje ten gaz, a przy PQ =
= 30 mm Hg (4,0 kPa) połowa tego barwnika oddechowego ma postać oksyhemo-globiny, a pozostała część— hemoglobiny. Ta szczególną wartość ciśnienia tlenu jest określana mianem ciśnienia połowicznego nasycenia {J*^n). Wraz z dalszym spadkiem ciśnienia tlenu wydziela się go coraz więcej, aż wreszcie, przy zerowym ciśnieniu tego gazu, hemoglobina zostaje całkowicie pozbawiona tlenu.
Krzywe dysocjacji tlenu u innych zwierząt mają kształt podobny, lecz nie są i identyczne z tą przedstawioną na rysunku 2.2. Krew niektórych zwierząt ma wyższe powinowactwo do tlenu (oddaje TTen mniej ćHetme). a ich Tcrzywe dysocjacji przesunięte są w lewo. Z kolei krew innych oddaje tlen chętniej (powinowactwo do tlenu jest niższejja krzywe dysocjacji prze-suńiete sa na prawo.
zmiana wartości P50 jest większa u mysz ruz u słonia (rys. 2.3). Innymi slow.'
Krzywa dysocjacji tlenu uzyskana dla danej próbki krwi jest silnie uzależniona od warunków, w których dokonuje się pomiarów. Spowodowane jest to wpływem kilku czynników na przyłączenie tlenu do hemoglobiny. Najważniejszymi z nich są:
Cl. Temperatura.
2. pH.
3. Dwutlenek węgla.
4. Fosforany organiczne.
5. Jony nieorganiczne, siła jonowa.
V.
Z powodu wielkiego znaczenia funkcjonalnego tych czynników potrzebne nam są dodatkowe informacje na temat ich działania.
Wpływ temperatury
Jeśli krzywą dysocjacji wyznacza się dla próbki krwi ssaka w temperaturze pokojowej, a następnie ponownie w temperaturze równej normalnej ciepłocie ich ciała, uzyskane wyniki są całkowicie różne. Dzieje się tak, ponieważ podwyższona temperatura osłabia wiązanie miedzy hemoglobina a tlę.nem, powodując wzrnsLjjysnejacji tego wiązania. Skutkiem tego w wyższej temperaturze hemoglobina chetniei oddaie tlen, a krzywa dysociacii przesuwa się na prawo
Ma_to_duże znaczenie fizjologiczne, szczególnie u zwierząt zmiennocieplnych, ponieważ podwyższeniu temperatury zwy-kle towarzyszy wzrost szybkości metabolizmu (czyli zapotrzebowania na tlen). Dla-tego korzystne jest to, że hemoglobina łatwiej dostarcza tlen w wyższej temperaturze. Może to mieć znaczenie nawet dla zwierząt stałocieplnych — na przykład, podczas gorączki lub wysiłku fizycznego, którym towarzyszy wzrost szybkości konsumpcji tlenu.
Wpływ dwutlenku węgla i pH
Innym ważnym czynnikiem, wpływaj cym na kształt krzywej dysocjacji, jest p-' osocza krwi. Zwiększenie stężenia dw. tlenku węgla 1 iTFstężenugmoych substann kwasowych powoduje zmniejszenie p; ; osocza, przesuwając krzywą dysocjacji r ^rawó~(rys. 2.2). Znaczenie tego przesuni cia polega na tym, że wysokie stężeń dwutlenku, węgla- powoduje większe o dawanie tlenu przy określonej wartoś jego ciśnienia. To zjawisko znane jesTpi nazwą efektu Bohra. _ W tkankowych n czyniach włosowatych, w miarę jak dw tlenek węgla przenika do krwi, hemogloł na oddaje większe ilości tlenu, niż miało! to miejsce, gdyby dwutlenek węgla n wywierał wpływu na jego wiązanie. Dlat go można mówić, że efekt Bohra urno _ liwia dostarczenie dodatkowej ilości tler do tkanek.
"' Wpływ dwutlenku węgla nie ogranie; się jednak do jego działania na pH. Jei utrzymywać pH na stałym poziomie, przi zastosowanie odpowiednich substancji bi forujących, wzrost stężenia dwutlenku w< gla nadal powoduje przesunięcie krzyw' dysocjacji na prawo. Innymi słowy, dwi tlenek węgla zmniejsza powinowactwo!^ moglobiny do tlenu nawet w stałym pi Działanie to jeltspowodowanc przez wi; zanie dwutlenku węgla bezpośrednio d końcowych grup cząsteczki hemoglobin (wiązanie to nie powstaje więc w tyi samym miejscu, gdzie wiązany jest tlen).
Znaczenie efektu Bohra nie jest jec nakowe dla wszystkich ssaków. Możem je wyrazić jako zmianę wartości P^, tc warzyszącą jednostkowej zmianie wartośi pH. Dzięki temu zauważymy, że efel
Wyszukiwarka
Podobne podstrony:
skanuj0021 (168) 5. Międzynarodowe aspekty ochrony przyrody172 Kilka gatunków delfinów: butlonosy, b
60 WSZECHŚWIAT lice wyspy Bouveta. Wyłowił on kilka „białych ryb krokodylowych**, a fotografie ich
12 Rozprawy Naukowe i Zawodowe PWSZ w Elblągu, zeszyt 13 (15 gatunków ryb) oraz przedstawiciele rodz
Kierunki działalności biologia ryb; monitoring stada; ocena zasobów ryb Antarktyki; ocena zasobów ry
anatomii i fizjologii lyb. Biologia chronionych gatunków ryb słodkowodnych. Biologia raków. Wędrówki
skanuj0004 (93) Dotychczas stwierdzono w wodach Zatoki Gdańskiej 74 gatunki ryb.Zatoka Gdańska (spis
65 (76) 16. Czy zarybianie wód otwartych obcymi gatunkami ryb takimi jak amur i tołpyga zwiększ
CCF20081123�011 192 Rodzaj: Mklik - Anagasta Do tego rodzaju zaliczono kilka gatunków małych motyli
łowi się lam wiele gatunków ryb, ale le, które Paskowi się podobają wyrzucają dla psów i ptaków, a s
DSC06507 * DciZy wpływ na decyzję o wyborze gatunku ryb do hodowli ma powierzchnia gruntów, jaką
DSCF6363 Czynniki bakteryjne W obtębic zmian trądzikowych znaleźć można kilka gatunków drobnoustrojó
Ptaki [bielik, rybolów, bocian, cietrzew, glusiec (ok. 10 gat.)] • Kilka gatunków
Zrównoważone rybołówstwo Niektóre gatunki ryb są zagrożone wyginięciem lub znajdują się na granicy
Rozpoznajemy gatunki ryb - warsztaty przyrodniczo -ekologiczne Projekt współfinansowany przez Unię
DSC08657 (4) Osoby z grupą krwi AB - typ „ENIGMA DOZWOLONE, baranina mięso Królicze małże i kawior n
Rośliny i zwierzętaWielkiej Raty Koralowej Także różne gatunki ryb zjadają żyjące
więcej podobnych podstron