skanowanie0009
54
hp
54
t
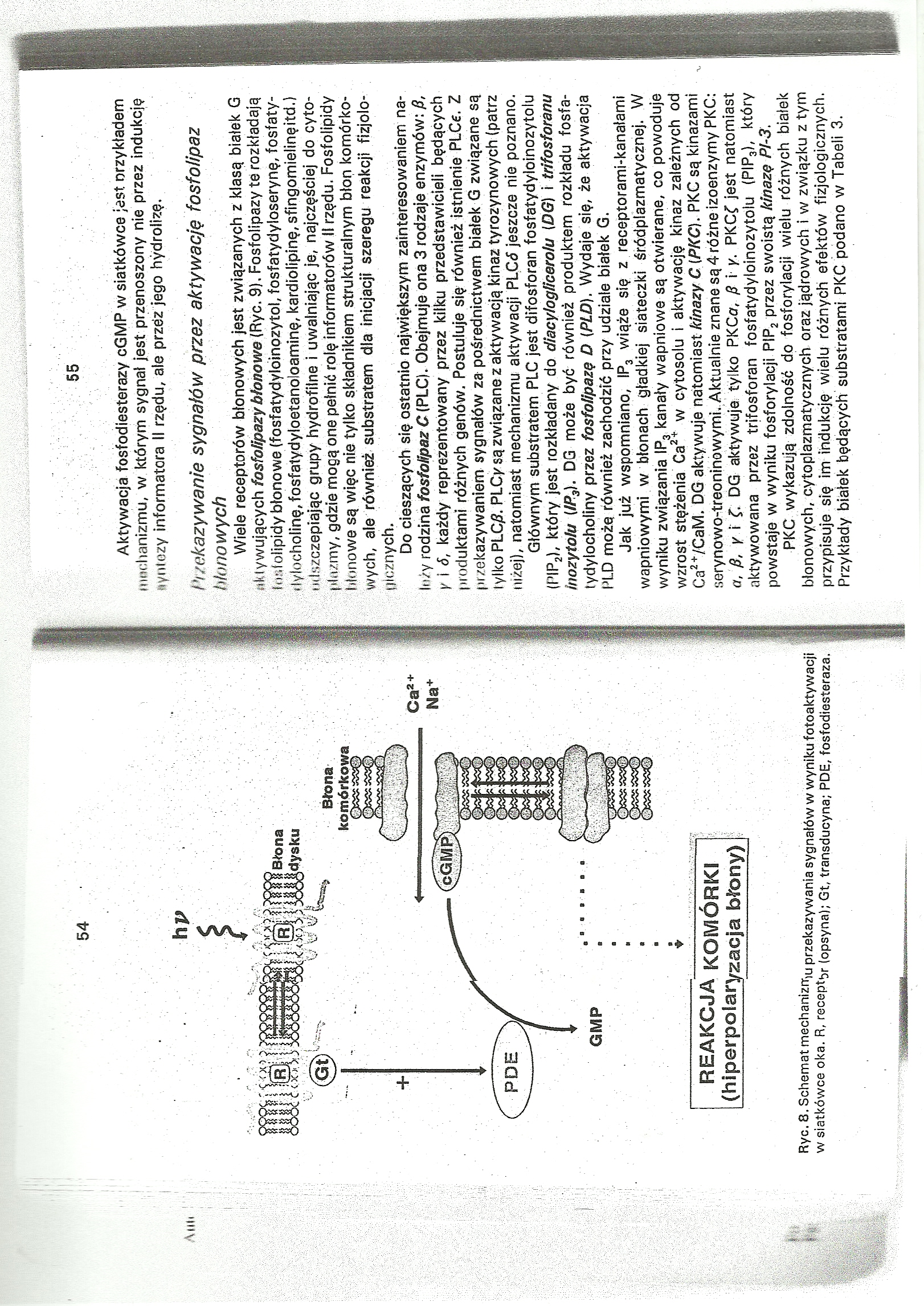
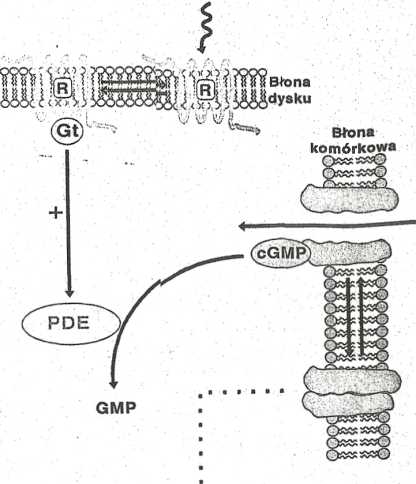
Ryc. 8. Schemat mechaniznr,u przekazywania sygnałów w wyniku fotoaktywacji w siatkówce oka. R, recepty (opsyna); Gt, transducyna; PDE, fosfodiesteraza.
Aktywacja fosfodiesterazy cGMP w siatkówce jest nrzykładem Mmchanizmu, w którym sygnał jest przenoszony nie przez indukcję tyntezy informatora II rzędu, ale przez jego hydrolizę.
Przekazywanie sygnałów przez aktywację fosfolipaz błonowych
Wiele receptorów błonowych jest związanych z klasą białek G Aktywujących fosfołipazybłonowe (Ryc. 9). Fosfolipazy te rozkładają fosfolipidy błonowe (fosfatydylóinozytol, fosfatydyloserynę, fosfaty-tlylocholinę, fosfatydyloetanoloaminę, kardiolipinę, sfingomieiinę itd.) imIszczepiając grupy hydrofilne i uwalniając je, najczęściej do cyto-płązmy, gdzie mogą one pełnić rolę informatorów II rzędu. Fosfolipidy błonowe są więc nie tylko składnikiem strukturalnym błon komórkowych, ale również substratem dla inicjacji szeregu reakcji fizjologicznych.
Do cieszących się ostatnio największym zainteresowaniem nałoży rodzina fosfolipaz C (PLC). Obejmuje ona 3 rodzaje enzymów: fi, y i 6, każdy reprezentowany przez kilku przedstawicieli będących produktami różfiych genów. Postuluje się również istnienie PLĆe. Z przekazywaniem sygnałów za pośrednictwem białek G związane są tylko PLC£. PLCysą związane z aktywacją kinaz tyrozyno wych (patrz niżej), natomiast mechanizmu aktywacji PLGó jeszcze nie poznano.
Głównym substratem PLC jest difosforan fosfatydyloinozytoiu (PIP2), który jest rozkładany do diacylogUcerolu (DG) i trifosforanu Inozytolu (IP3). DG może być również produktem rozkładu fosfa-tydylochpliny przez fosfollpazę D {PLO). Wydaje się, że aktywacja PLD może również zachodzić przy udziale białek G.
Jak już wspomniano, IP3 wiąże się z receptorami-kanałami wapniowymi w błonach gładkiej siateczki śródplazmatycżriej. W wyniku związania IP3 kanały wapniowe są otwierane, co powoduje wzrost stężenia Ca2* w cytosolu i aktywację kinaz zależnych od Ca2*/CaM. DG aktywuje natomiast kinazy C [PKC). PKC są kinazami serynowortreoninówymi. Aktualnie znane są4różne izoenzymy PKC: o, fi, y \ C DG aktywuje tylko PKCav fi i y. PKCf jest natomiast aktywowana przez trifosforan fosfatydyloinozytoiu (PIP3), który powstaje w wyniku fosforylacji PIP2 przez swoistą kinazę PPS.
PKC wykazują zdolność do fosforylacji wielu różnych białek błonowych, cytoplazmatycznych oraz jądrowych i w związku z tym przypisuje się im indukcję wielu różnych efektów fizjologicznych. Przykłady białek będących substratami PKC podano w Tabeli 3.
Wyszukiwarka
Podobne podstrony:
skanowanie0011 !2MH Cykl lyclowy komórki I ]®j jtttfjtliłl / / / Hy< , 0,1,1. Schemat przebieg
Wzór ryciny komórki ziarniste śc Ryc. 1. Schemat budowy pęcherzyka jajnikowego ssaka (na podstawie
Wzór ryciny komórki ziarniste śc Ryc. 1. Schemat budowy pęcherzyka jajnikowego ssaka (na podstawie
Wzór ryciny komórki ziarniste śc Ryc. 1. Schemat budowy pęcherzyka jajnikowego ssaka (na podstawie
14 J. Jirout Ryc. 4. Schemat mechanizmu nachylenia ku przodowi dolnych kręgów szyjnych, powiązanego
20583 skanowanie0016 Błona komórkowa jonem błony, a jedynie wiązać się z nią za pomocą kowalencyjneg
73796 skanowanie0014 64 AuREAKCJA KOMÓRKI Ryc. 13. Mechanizmy aktywacji szlaku przekazywania sygnałó
skanowanie0003 pienia ostatecznej reakcji komórki. Schemat taki prezentuje Ryc. Informator I rzędu I
więcej podobnych podstron