30743 P1520540

mH wu\\u\ luli W\uuv\\Iwa wiwiiiiBWHi ii^iiiIhhwi^hiiih • Im-mii> ~««i«
* Wl\\\W'\w\\ih) HlilliuMiBH Miniihvi'Hi\Viłi jiii nim llfHHitijiH Mmiv Ihiiiiiii IiiiwhI hMlł lipni*" \\U>wwm\n i'u nim nhrnmt<h Mmii lummilm^Hi iiliWNltiijH wi^h miimih iu^§nVliB ł'l\nł>Uw*nv»'l\ u |M4wnłivnvi'li mmIiIh lntlnnl*Hhl) ll|Śiffej$(|$
nvwv i^n iinwim mBiui ^\mj jńHl! |ii#<\yhKN iIimIh (himIi 11111 i i m Ih ha1 Minii n>Y^\VYW v\h\I'Mv\\\uv\ \\m wi\i|pwi it.immumli iHiiIitmiwij (jui^ i ii thMh
**- A-
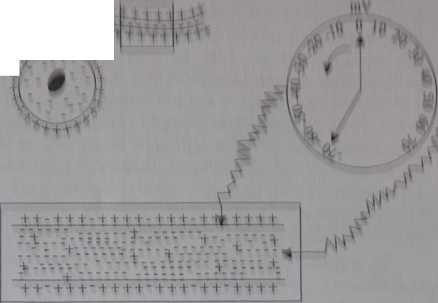
Rys. 1.4. Rozmieszczenie kationów (koić? czerwony) i anionów (kolor zielony) po whą stronach błony komórkowej neuronw w stanie spoczynku- Na wyoinkn nerwu, mieUzy wnętrzem a zewnętrzna powierzchnia włókna nerwowego potencjometr wskazuje noplecio równe -70 mV (prąd spoczynkowy)
uporządkowanie ładunków elektrycznych - czyli polaryzacja - jest wynikiem nie tylko dyfuzji jonów przez błonę komórkową, zmierząjącej do wyrównania stężeń, ale również aktywnego procesu enzymatycznego. Polaryzacja elektryczna komórki i utrzymanie stałej różnicy potencjałów elektrycznych po obu stronach błony komórkowej jest skutkiem działania dwu podstawowych struktur białkowych: białkowego kanału jonowego dla K+, umożliwiającego wchodzenie potasu do wnętrza komórki oraz enzymu - ATP-azy, zależnej od Na+i K+, przenoszącej 3 jony Na+ na zewnętrzną powierzchnię błony komórkowej i 2 jony K+ do wnętrza komórki. Kanały jonowe dla sodu, potasu, chloru i innych jonów, jako efektory docelowe receptora (rys. 1.2), są otwierane lub zamykane w wyniku reakcji hormonu lub mediatora z receptorem jonotropowym (str. 14). Otwieranie i zamykanie kanału jonowego jest nazywane bramkowaniem. W efekcie aktywnego transportu, we wnętrzu neuronu gromadzą się, w przeważającej koncentracji, naładowane ujemnie i nie dyfundujące aniony pochodzenia organicznego (fosforany organiczne, aminokwasy), a na zewnątrz neuronu - dodatnio naładowane kationy sodu (rys. 1.4). Dzięki tym procesom powstaje różnica potencjałów między wnętrzem neuronu a zewnętrzną jego powierzchnią. Wynosi ona -70 mV (rys.
Jest to prąd spoczynkowy o napięciu -70 mV. Zachowanie różnicy
16
Wyszukiwarka
Podobne podstrony:
67094 Obraz7 W lym mlujm u iiIiiIrjHzyiih m/wu/mi dotykalny Iwa),iing
majesttic12ilustracja TmC mmtC nOv»c **r u, m>TCF JECST V*mh) totnutim rzxMrmi n* :ouaM fm.fM &nb
majesttic12ilustracja TmC mmtC nOv»c **r u, m>TCF JECST V*mh) totnutim rzxMrmi n* :ouaM fm.fM &nb
3 (76) 7 tń Mh I 1 a 4 II Olchy iUIIDyiMlU/IM* HA Olchy i /iiiitu/iiMiiiia IIAl « /iimn/iriny u)
Wolbrom. 1(> luli^jo 2018CERTYFIKAT E-SYS II M Morem Firek nini<r»/ym certyfikatom z.i wiarJcz
Zdjęcie052 -Vm Vn I */fl mwv( w# l Mh‘ m j»wn I • i 4N«# Nivn 00 i w »
r jfflgn wlkll y m /JnU I lilii V /Ul Biiłl jtf MH ■gffc 1 V Im r mii 1
majesttic12ilustracja TmC mmtC nOv»c **r u, m>TCF JECST V*mh) totnutim rzxMrmi n* :ouaM fm.fM &nb
majesttic12ilustracja TmC mmtC nOv»c **r u, m>TCF JECST V*mh) totnutim rzxMrmi n* :ouaM fm.fM &nb
STA7ESMAN W«dn*d«y. May 9.1964 iwa filii n Y.ii lilii !?{jM
P1520562 pwti Mtih wwpuiąwvchi ąąii vąiuą(l$iuvvi*Ii m ią #włvłu
więcej podobnych podstron