DSC00720
wstałykompleks Os-GDP ma zdolność do łąpzenia się z podjednos tką py, co pozwala na odtworzenie się białka G>. Omawiany cykl białka Gs zamyka oddysocjowanie GDP od podjednostki p W świetle przedstawionych przemian staje się oczywistym, że działanie receptora związanego z ligandem powinno polegać na przyspieszaniu reakcji odpowiedzialnej za powstawanie kompleksu ou-GTP aktywującego cyklazę adeny łanową. Dodać należy, że wbrew uprzednim przekonaniom dimer py powstający podczas dysocjacji białka Gs może mieć dodatkowy wpływ na odpowiedź komórkową poprzez stymulację innego efekt ora niż kompleks ou-GTP. Transdukcję sygnału z udziałem białka G przedstawiono schematycznie na rysunku 28.6.
Dotychczasowe badania pozwoliły na poznanie z jakimi receptorami współdziałają określone rodzaje białek G w transdukcji sygnału poprzez błonę komórkową. I tak wiadomo, że białka G, odpowiedzialne są za aktywację cyklazy adeny łanowej oraz oddziaływanie na kanał wapniowy i sodowy w przypadku stymulacji receptorów (J-adrenergicznych, TSH i glukagonu. Białka G, będące drugim rodzajem białek G współpracują z receptorami: da-adreneigicznymi, Mz-muska-rynowym oraz Met-enkefalinowym-GTP i są odpowiedzialne za hamowanie aktywności cyklazy adenylanowej. aktywowanie fosfolipazy C i Az oraz funkcjonowanie kanałów K* i Ca2*. Białka Gq są związane z receptorami: oti-adrenergicznymi i Mi-muskarynowymi wpływającymi na aktywację fosfolipazy C-j)i, -jfe i -fh. Dotychczas nie udało się ustalić zarówno receptorów, jak i efektorów związanych z rodziną białek Giz.
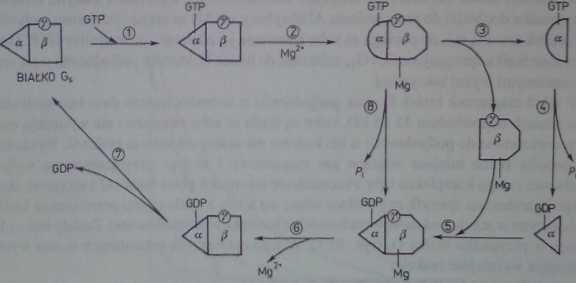
Rys. 28.6. Hipotetyczny cykl przemian białka Q« w stanie podstawowym komórki. Cykl przemian białka G> zbudowanego z podjednostek a. p i Y wiąże się z ich dysocjacją i reasocjacją połączoną ze zmianą właściwości biologicznych. (Na schemacie podjednostki CU. P i Y zaznaczono odpowiednio przez trójkąt, prostokąt i kółko). W wyniku przyłączenia sk GTP do podjednostki cu białka G« (reakcja 1) oraz jonów Mg2* do podjednostki p (reakcja 2) dochodzi do zmian konfiguracyjnych w ich budowie zaznaczonych na schemacie przez zmianę kształtu podjednostki a z trójkątnego aa półokrągły, a podjednostki p z prostokątnego na sześci oboczny. powodujących przejście pierwszej z nich w stan „aktywny*. w którym jest zdolna do aktywacji cyklazy adenylanowej. Od tak powstałego kompleksu oddysocjowuje aktywna podjednostka cu związana z GTP (reakcja 3). która przechodzi w stan nieaktywny z momentem hydroli tycznego odłącze-nia fosforanu (Pi) od sprzężonego z nią GTP (reakcja 4). Powstały kompleks podjednostki a z GDP łączy się ponownie z kompleksem podjednostki P i y oraz Mg2* (reakcja 5). W następnych dwóch reakcjach dochodzi do odłączenia sięjono magnezu i GDP od odpowiednich podjednostek. co prowadzi do przejścia w stan. od którego rozpoczęto omawianie przemian. Reakcja 8 uwzględnia możliwość hydrolizy GTP przed dysocjacją białka G„ co umożliwiłoby ominięcie reakcji 3-*5 w przebiegucylefo- Przyjmuje się. Ze kompleks receptor -ligand, może wpływać na przyspieszenie reakcji 7 i l.iiym samym powodować rwięłozone powstawanie kompleksu a-GTP
Wyszukiwarka
Podobne podstrony:
83065 IMG336 Wpływ czynników pozatermicznych na komfort cieplny. Organizm ludzki ma zdolność do przy
DSC00722 (3) Endospory (przetrwał ni ki) Zdolność do wytworzenia endospor ma tylko mała grupa bakter
□ Poszukujemy odpowiedzi na pytania: ■ Czy firma ma zdolność do
Kapitał to kategoria ekonomiczna, która ma zdolność do wzrostu, do pomnażania się. Różnica wartości,
Kształtowanie struktury kapitału Kapitał - rzecz, która ma zdolność do samoistnego
zagad 2 Ciężar dowodu Pojęcie stosunku prawnego Zdolność prawna os. fizycznych i pr. Zdolność do czy
DSC00760 (3) Oddziaływanie wirusów * Szybkie rozmnażanie oraz zdolność do rozprzestrzeniania się w p
56 Artur GUZOWSKI, Andrzej SOBCZYK tele-operacji wynosi do 1500m. Manipulator ma zdolność do podejmo
Nadzór immunologiczny - ludzie Najistotniejsze znaczenie dla nadzoru immunologicznego ma zdolność do
I Zdolność do zwijania języka w rurkę to dominująca cecha autosomnlna (/?, #*). Franek nie ma zdolno
Każda żywa komórka roślinna ma zdolność do pobierania i oddawania wody. U podstaw tych zjawisk leżą
przykladowe testy 9 A Test nr 1 I. Każdy człowiek od chwili urodzenia ma a zdolność do czynności pra
czynności prawne wykonują za nie przedstawiciele ustawowi; czynność prawna osoby, która nie ma zdoln
c) centrosom/ centrum komórkowe/ centmm ruchowe/ centrosfera- ma zdolność do polimeryzacji mikrotubu
traktatowa podmiotów art.6 KW o PT: „Każde państwo ma zdolność do zawierania traktatu.” Ta zdolność
więcej podobnych podstron