5282744613
forów fitochromów roślin wyższych, wykazywało fotoodwracalność i ulegało auto fosforylacji na reszcie histydyny [18]. Proces ten zachodził intensywniej po naświetleniu daleką czerwienią. Ponadto forma Pr cph 1 fosforylowała rep 1, podczas gdy forma Pfr nic wykazywała takiej aktywności (Ryc. 2).
Nie wiadomo, czy fitochrom roślin wyższych wykazuje podobne właściwości, jednakże zarówno forma Pr, jak i Pfr fitochromu owsa może fosforylować białko rcpl [19]. W tej sytuacji fosforylacji ulegała jednak nie reszta asparaginy, jak to ma miejsce w przypadku kinaz histydynowych, a reszta seryny, co sugeruje, że fitochrom zachowuje się jak kinaza Ser/Thr. W związku z tym postuluje się, że fitochrom roślin wyższych zachował aktywność kinazy histy-dynowej, w której reszta fosforanowa jest intensywnie przenoszona na serynę [21, 22].
Możliwość funkcjonowania fitochromu jako kinazy serynowo-treoninowej postulowano już wcześniej, kiedy u mszaka Ceratodon purpureus zidenty-
Skłoniło ich to do sugestii, że ufosforylowana tyrozyna może służyć jako miejsce wiązania białek mających domeny SH2, rozpoznających jedną z dwóch specyficznych sekwencji aminokwasowych: pTyr-hydrofilowy-hydrofilowy-hydrofobowy lub pTyr-hydrofobowy-dowolny-hydrofobowy. Autorzy doliczyli się sześciu takich sekwencji w fitochro-mie A owsa.
III-2. Drobnocząsteczkowc elementy szlaku sygnałowego
Charakterystycznymi reakcjami rośliny na światło czerwone i daleką czerwień są: ekspresja genów białek PSI i PSU (fnr, cab) oraz genów odpowiedzialnych za biosyntezę antocyjanów (chs).
Stosując mikroiniekcje do subepidcrmalnych komórek hipokotyla mutanta pomidora — Aurea specyficznych agonistów i antoganistów białek G wykazano, żc jednym z pierwszych etapów przekazania sy-


cab-GUS
Ekspresja
chs-GUS
Ekspresja
fnr-GUS
Ekspresja
cab-GUS
cGMP
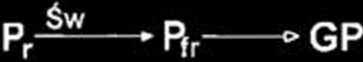
cGMP
Światło ■ ■»
fikowano białko fitochromowe, którego N-końcowa domena wykazywała cechy pośrednie pomiędzy fito-chromem A i B Arabidopsis, a domena C-końcowa posiadała sekwencje aminokwasowe homologiczne do eukariotycznych kinaz Scr/Thr/Tyr [5, 23, 24].
Właściwości kinazy Ser/Thr wykazują także flto-chromy uzyskane z owsa i z algi zielonej (Mesotae-nium caldariorum), przy czym bardziej aktywną formą pod względem autofosforylacji jest Pfr. Formy Pr i Pfr w równym stopniu, fosforylują histon H1, który z kolei stymuluje autofosforylację głównie formy Pr [19].
Fosforylację reszt tyrozyny w fitochromie w badaniach in vitro wykazali Sommer i wsp. [25].
Ekspresja
chs-GUS
Ekspresja
fnr-GUS
Fksnrpęia R-vc* 3* Schemat przedstawiający udział substancji
drobnocząstcczkowych w szlakach sygnałowych fitochromu (za 28, zmieniono]. Szczegóły w tekście. 1. Represja łańcucha transdukcji sygnałów zależnego od cGMP przez szlak obejmujący Ca27CaM. 2. Represja szlaku sygnałowego zależnego od Ca27CaM oraz od cGMP i Ca2VCaM przez kaskadę z udziałem cGMP. Oznaczenia strzałek: pełne — fotokon-wersja fitochromu; puste — kolejne etapy przekazywania sygnałów, tępo zakończone — negatywna regulacja jednego szlaku przez inny. GP — białka G.
gnału fitochromowego jest aktywacja tych heterotri-mcrycznych białek (Ryc. 3) [26]. W doświadczę-niach z wykorzystaniem Ca i aktywowanej kalmo-duliny stwierdzono, że cząsteczki te częściowo zastępują fitochrom A stymulując ekspresję cab-GUS oraz syntezę białek kompleksu fotosyntetycznego PSII — reakcje, których brakiem cechuje się mutant. Natomiast koiniekcje cGMP, jonów Ca2+, oraz inhibitorów i aktywatorów białek G wykazały, że cGMP aktywuje ekspresję chs-GUS, a w połączeniu z wapniem —fnr-GUS [27-29].
Na podstawie tych doświadczeń wyciągnięto wniosek, że cGMP działa jako wtórny przekaźnik na szlaku prowadzącym od fitochromu do syntezy anto-
187
POSTĘPY BIOCHEMII 47(2), 2001
Wyszukiwarka
Podobne podstrony:
Fizjologia 3 - Morfologia i anatomia aparatu fotosyntetycznego roślin wyższych -
Piasek z luźną roślinnością zielną - typowe siedlisko gniazdowania sieweczki rzecznej na wyższych wy
reakcja przebiegała intensywniej w przypadku formy Pfr [33J. Rośliny transgcniczne, w których PKS 1
5 (1497) 4.9. Sacharydy 155 komórkowej roślin wyższych, np. drewna (45-50%) i bawełny (90%). Otrzymy
Katedra Farmakognozji Uniwersytetu Medycznego w ŁodziANALIZA FITOCHEMICZNA roślinnych substancji
74714 Zdjęcie4136 Charakterystyka ogólna Występują w roślinach wyższych, głownio w postaci sol Kwasó
DSC00670 ZAWIĄZKI PLASTYDÓW Ryt 26,2. Wzajemna przemiana plastydów u roślin wyższych. Strzałkami prz
System waskularny odpowiadający za daleki transport w roślinach wyższych składa się z dwóch oddzieln
I Chlorofil b występuje u zielenic jest typowy dla roślin wyższych. 2.Rhodophyta
I .Chlorofil b występuje u roślin wyższych i zielenicach nie występuje u brunatnie
I Chlorofil b występuje u zielenic jest typowy dla roślin wyższych. 2.Rhodophyta
I .Chlorofil b występuje u roślin wyższych i zielenicach nie występuje u brunatnie
więcej podobnych podstron