Rozmnażanie bakterii
Bakterie rozmnażają się bezpłciowo poprzez prosty podział komórki macierzystej na dwie komórki potomne (rozszczepienie).
Po zreplikowaniu materiału genetycznego powstaje przegroda poprzeczna będąca wpukleniem błony komórkowej.
Charakterystyczny jest brak wrzeciona kariokinetycznego tworzonego podczas mitozy u Eucaryota.
pierwszy etap - podział materiału genetycznego (nukleoidu), uprzednio podwojonego w wyniku replikacji, ma on miejsce w fazie intensywnego wzrostu komórki
drugi etap - po podziale nukleoidu następuje podział komórki w wyniku tworzenia się przegrody poprzecznej (septum), wrastającej od zewnątrz do wnętrza komórki i dzielącej protoplast na dwie części
Na powstałej przegrodzie odkładane są składniki dobudowywanej ściany komórkowej bakterii.
Rozmnażanie bakterii przez podział prosty, w wyniku którego z każdej komórki macierzystej powstają dwie komórki potomne powoduje, że ich liczba wzrasta w postępie geometrycznym.
20 > 21 > 22 > 23 > ......................> 2n
gdzie n to liczba podziałów.
W zależności od formy morfologicznej typowej dla danego gatunku bakterii, po zakończonym podziale komórki mogą się rozdzielić lub pozostać w różnego typu formacjach morfologicznych np. laseczki mogą tworzyć łańcuszki, ziarniaki mogą pozostawać w agregacjach od dwoinek po gronkowce.
U bakterii nie stwierdzono rozmnażania płciowego.
Przekazywanie i wymiana materiału genetycznego zachodzi jedynie w stwierdzonych tu procesach płciowych, nie mających nic wspólnego z rozmnażaniem płciowym, jak to jest u Eucaryota.
Bakterie (prawie zawsze) są haploidalne, mogą one tworzyć zygoty ale nigdy przez fuzje całych komórek.
Zazwyczaj tylko część materiału genetycznego komórki dawcy zostaje przeniesiona do komórki biorcy (akceptora), z wytworzeniem niepełnej zygoty (merozygoty).
Chromosom biorcy i część chromosomu dawcy łączą się w parę i wymieniają między sobą odcinkami. Następne podziały chromosomów i komórek prowadzą do powstania potomstwa zawierającego zrekombinowane chromosomy.
Wyróżnia się trzy sposoby przekazywania informacji genetycznej u bakterii, określanego jako horyzontalny transfer genów, a są to:
koniugacja
transdukcja
transformacja
Te trzy typy różnią się jedynie sposobem przenoszenia.
Wniknięcie DNA dawcy do komórki biorcy i połączenie z jego DNA powoduje rekombinację DNA akceptora, a komórkę w której ten proces zaszedł nazywamy rekombinantem.
Rekombinacja może mieć charakter:
ogólny (homologiczny) - DNA dawcy zostaje włączony do DNA biorcy poprzez połączenie się w pary homologicznych sekwencji, pęknięcie i krzyżową wymianę (crossing-over). Warunkiem koniecznym tego procesu jest istnienie długich odcinków homologii sekwencji DNA obu partnerów.
zlokalizowany (zależny od określonego miejsca lub sekwencji) - wymaga jedynie krótkiego odcinka homologii DNA obu partnerów w celu rozpoznania się (np. integracja bakteriofaga ၬ).
Koniugacja
Bezpośrednie przeniesienie DNA z komórki do komórki w drodze ich bezpośredniego kontaktu.
Podczas koniugacji dawca i biorca wchodzą w ścisły kontakt, w którym uczestniczy duża grupa białek tworzących połączenie między koniugującymi komórkami - mostek koniugacyjny. Również białka znajdujące się w kanale uczestniczą w przenoszeniu DNA.
Koniugacja jest procesem polegającym na jednokierunkowym przekazaniu materiału genetycznego z komórki dawcy do biorcy, ponadto zarówno rekombinacja jak i segregacja genów zachodzi tylko w komórce biorcy.
Zdolność pełnienia roli dawcy zależy od obecności w jego komórce ruchomej cząsteczki DNA tzw. czynnika płciowego F (fertility z ang. płodność). Komórki nie posiadające tego czynnika (F-) mogą być jedynie biorcami.
Czynnik F jest zamkniętą, kolistą, zdolną do autonomicznej replikacji cząsteczką dwuniciowego DNA o wielkości ok. 100 tys. pz. Jest to więc pozachromosomalny czynnik dziedziczności - plazmid.
Plazmid F zawiera geny odpowiedzialne za proces koniugacji, a więc kodujące specjalne struktury na powierzchni komórki np. pilusy F (w liczbie 2-3).
DNA czynnika F penetruje komórkę biorcy po częściowym złączeniu się komórek krzyżujących się partnerów.
Tylko nieliczne bakterie z populacji F+ mogą przekazać chromosomowy DNA. Cechę tę mają komórki, w których nastąpiła integracja czynnika F do chromosomu biorcy. Takie komórki określamy jako Hfr (high frequency of recombinants = wysoka częstość rekombinantów).
Liczba rekombinantów powstałych w wyniku koniugacji z komórkami Hfr jako dawcami przewyższa tysiąckrotnie liczbę otrzymaną po koniugacji z komórkami F+.
Transdukcja
Proces przenoszenia fragmentu DNA z jednej komórki do drugiej przez bakteriofaga łagodnego (w czasie cyklu lizogenicznego). Fag taki wbudowuje część bakteryjnego DNA w swój własny kwas nukleinowy i przenosi go jako trwałą cechę na szczep biorcy. Zazwyczaj przenoszony jest tylko mały fragment DNA gospodarza (dawcy).
Proces przenoszenia fragmentu DNA z jednej komórki do drugiej przez bakteriofaga łagodnego (w czasie cyklu lizogenicznego). Fag taki wbudowuje część bakteryjnego DNA w swój własny kwas nukleinowy i przenosi go jako trwałą cechę na szczep biorcy. Zazwyczaj przenoszony jest tylko mały fragment DNA gospodarza (dawcy).
Fag replikujący się w bakterii w trakcie cyklu litycznego tnie bakteryjny chromosom. W trakcie składania nowych cząstek wirusowych zamiast DNA fagowego do główki fagowej może zostać omyłkowo zapakowany fragment DNA bakteryjnego.
Zainfekowanie taką cząstka wirusową kolejnej komórki bakteryjnej powoduje wprowadzenie do niej fragmentu DNA innej bakterii, a więc nie spowoduje ani lizogenii ani lizogennego wzrostu.
Wprowadzony obcy DNA albo włącza się do chromosomu w wyniku homologicznej rekombinacji, albo zostanie utracony.
Transdukcję dzielimy na:
Niespecyficzną (ogólną) - gdy każdy segment DNA gospodarza może zostać przeniesiony. W tym typie transdukcji fragment DNA gospodarza zostaje włączony do cząstki wirusowej albo jako odcinek dodatkowy albo zastępując w całości genom fagowy (wirus przenosi tylko fragment DNA gospodarza).
Specyficzną - ograniczoną do przekazania określonych segmentów DNA. Tylko niektóre geny fagowe są zastąpione przez geny gospodarza (np. fag lambda przenosi określone geny należące do operonów gal (wykorzystanie galaktozy) i bio (synteza biotyny)).
W obu przypadkach fagi transdukujące są zazwyczaj poronne np. często tracą zdolność do lizy komórek gospodarza
Transformacja
Jest to proces przekazania genów szczepom biorcy w formie rozpuszczalnego DNA wyekstrahowanego lub uwolnionego z komórki dawcy, z pominięciem łączenia się komórek dawcy i biorcy w pary.
Do wykrycia tego procesu przyczyniły się badania Griffitha nad dwoinką zapalenia płuc (Streptococcus pneumonie). Opisał on zmianę bezotoczkowego, łagodnego szczepu R w szczep otoczkowy, zjadliwy S.
Pobranie DNA ze środowiska.
Transformacja naturalna - zachodząca w środowisku naturalnym
Transformacja sztuczna - indukowana np. przez szok chemiczny, elektryczny, który ma sprowokować bakterie do pobrania DNA do czego nie były one zdolne w warunkach naturalnych
Istnieje kilka doświadczalnych metod transformacji:
Wykorzystanie naturalnej zdolności (kompetencji) niektórych gatunków bakterii do pobierania obcego DNA. W ten sposób może być przekazana np. cecha prototrofizmu czy oporności na antybiotyki (Acinetobacter, Azotobacter, Bacillus, Haemophilus, Mycobacterium, Neisseria, Pseudomonas, Streptococcus).
Indukcja kompetencji przez specjalne, przedtransformacyjne zabiegi (ściśle określone zmiany warunków hodowlanych)
Protoplastyzacja komórek (Bacillus, Streptomyces), pozbawione ściany komórkowej po potraktowaniu ich polietylenoglikolem, są zdolne do pobrania plazmidowego DNA.
Elektroporacja - zwiększenie przepuszczalności błony cytoplazmatycznej pod wpływem pola elektrycznego.
Taksonomia bakterii
W zależności od kryteriów podział taksonomiczny bakterii może mieć charakter:
konwencjonlany (sztuczny nie opierający się na klasyfikacji filogenetycznej) K
molekularny M w tym filogenetyczny F
numeryczny
W mikrobiologii jednostką podstawową jest szczep. Jest to czysta kultura wyizolowanej bakterii.
Szczepy grupowane są w gatunki (species), gatunki w rodzaje (genus), rodzaje w rodziny (końcówka -aceae).
W klasyfikacji bakterii bierze się pod uwagę następujące cechy:
morfologia (komórki i kolonii na podłożu hodowlanym) - K
odczyn Gram - K
właściwości fizjologiczno-biochemiczne
wzrost w warunkach tlenowych i beztlenowych - K
sposób uzyskiwania energii (oddychanie, fermentacja, fotosynteza) - K
zależność wzrostu od temperatury i pH - K
wykorzystywane źródła energii (składniki pokarmowe) - K
siedlisko - K
interakcje między bakteriami (symbioza, antybioza, metabioza, pasożytnictwo)- K
inkluzje komórkowe, barwa, materiały otoczkowe - K
wytwarzanie endospor - K
skład ściany komórkowej, błony komórkowej (profil białek i lipidów) - K
właściwości serologiczne (skład oligosacharydów) - K
wrażliwość na antybiotyki - K
skład zasad w DNA (stosunek G:C) - M, F
hybrydyzacja DNA-DNA - M
zdolność do transformacji międzygatunkowej - M
sekwencja 16S rRNA lub 5S rRNA - F
Symbole K, M, F odpowiadają rodzajowi taksonomii
Taksonomia numeryczna - opiera się na zasadzie Adansona, wg której wszystkie uchwytne cechy organizmu są w klasyfikacji jednakowo ważne. W analizie tej należy brać pod uwagę jak największą liczbę cech.
Wycena cech jest jednoznaczna: obecność (+), brak (-).
Podobieństwo pomiędzy szczepami jest wyrażone współczynnikiem podobieństwa S, które oblicza się ze wzoru:
S = a + d/ (a+b+c+d)
a i d - to sumy cech wspólnych dla szczepów A i B (a - obie pozytywne, d - obie negatywne)
b - to suma cech pod względem których A jest pozytywny a B jest negatywny
c - to suma cech pod względem których A jest negatywny a B jest pozytywny
Współczynnik przyjmuje wartość między 1 i 0
S = 1 oznacza 100% podobieństwa
S ြ 0,02 oznacza całkowity brak podobieństwa
Najważniejsze grupy bakterii
W oparciu o morfologię, możliwość wzrostu w warunkach tlenowych/beztlenowych oraz odczyn Grama podzielono bakterie na kilka grup, z których omówione zostaną najważniejsze.
Grupa:
1. Ziarniaki (cocci), formy kuliste
Ziarniaki Gram(+)
tlenowe: Micrococcus, Staphylococcus, Streptococcus, Leuconostoc, Pediococus
beztlenowe: Sarcina, Peptococcus,Ruminococcus
Ziarniaki Gram(-) - są tu bakterie glebowe, patogeny orazi zasiedlające przewód pokarmowy, błony śluzowe ssaków.
tlenowe: Neisseria, Moraxella, Acinetobacter,
beztlenowe: Acidaminococcus, Veilonella
Pałeczki (proste bakterie cylindryczne)
Gram(+)
tlenowe G(+) nie tworzące endospor: Lactobacillus, Listeria,
tlenowe maczugowce i promieniowce: Corynebacterium, Arthrobacter, Brevibacterium, Eubacterium, Bifidobacterium, Mycobacterium, Nocardia, Actinomyces, Micromonospora, Streptomyces.....
tlenowe pałeczki i ziarniaki tworzące endospory: Bacillus, Sporolactobacillus, Sporosarcina, Thermoactinomyces
beztlenowe pałeczki i ziarniaki tworzące endospory: Clostridium, Desulfomaculatum, Oscillospira
Tlenowe bakterie fototroficzne - sinice (Cyanophyta)
fototrofy oksygenowe, zawierają chlorofil a i inne barwniki (takie same jak w chloroplastach roślin)
duże zróżnicowanie morfologiczne, występują zarówno formy jedno- jak i wielokomórkowe
niektóre poruszają się ruchem ślizgowym
niektóre gatunki posiadają w nici ściśle wyspecjalizowane komórki - heterocysty - odpowiedzialne za wiązania azotu atmosferycznego, co powoduje, ze są one organizmami pionierskimi, budowa komórki, ściana komórkowa zawierająca mureinę, rybosomy 70S - świadczą o tym, że sinice są gramujemnymi bakteriami
szeroko rozpowszechnione w wodach słodkich, słonych jak i na lądzie, wykazują zdolność do wzrostu w ekstremalnych środowiskach (np. kwaśne pH 4,0, gorące 700C źródła)
niektóre gatunki żyją w symbiozie (np. Nostoc, Anabaena)
Klasyfikacja:
Grupa I SINICE CHROOKOKALNE (jednokomórkowe pałeczki i ziarniaki)
Komórki pojedyncze lub zagregowane otoczkami bądź śluzem. Dzielą się przez podział poprzeczny lub pączkowanie.
Synechococcus, Gleocapsa, Gleothece, Gleobacter
Grupa II SINICE PLEUROKAPSULARNE -formy jednokomórkowe, tylko te, które rozmnażają się przez podziały wielokrotne, w wyniku tego procesu w dzielącej się komórce macierzystej powstaje wiele małych komórek - beocytów.
Pleurocapsa, Dermocarpa, Myxosarcina
Kolejne trzy grupy :
tworzą nici złożone z łańcuchów komórek,
poruszają się ruchem ślizgowym
wzrost nici ma charakter interkalarny tzn. zachodzi przez podział komórek w obrębie nici, a nie na wierzchołkach
mogą rozmnażać się poprzez hormogonia - fragmenty nici powstałe w wyniku ich rozerwania.
Grupa III SINICE NITKOWATE NIE TWORZĄCE HETEROCYST - nici wyłącznie wegetatywne
Oscillatoria, Spirulina, Phormidium
(drgalnica
Grupa IV SINICE NITKOWATE TWORZĄCE HETEROCYSTY - w warunkach niedoboru azotu, niektóre komórki różnicują się i tworzą heterocysty, a w pewnych warunkach mogą powstawać akinety, grubościenne komórki spoczynkowe.
Anabaena, Nostoc, Calotrix,
Grupa V SINICE NITKOWATE TWORZĄCE HETEROCYSTY - od grupy IV różnią się sposobem podziału komórki - w więcej niż w jednej płaszczyźnie.
Fischerella
Fikobilisomy
Krążkowate struktury przyczepione do zewnętrznej powierzchni tylakoidów
- Zawierają:
fikocyjaninę, fikobiliproteinę (75%),
fikocjanobilina
fikoerytrobilina
allofikocyjaniny (12%)
fikoerytryna
bezbarwne polipeptydy
Archaea
Archaea dzielimy na trzy grupy:
Metanogeny
Ekstremalne halofile
Ekstremalne termoacidofile
Archeony:
żyją w środowiskach ekstremalnych takich jak
gorące źródła (Sulfolobus, Pyrococcus)
duże zasolenie (Halobacterium)
bezwzględnie beztlenowe - bakterie metanogenne (Methanobacterium)
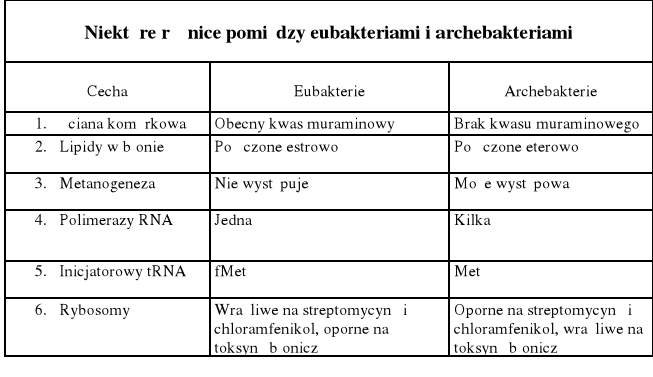
rybosomy (70S) Archeabakterii są tej samej wielkości jak u Eubacterii, lecz opornością na antybiotyki (streptomycyna, chloramfenikol) oraz wrażliwością na toksynę błoniczą przypominają rybosomy eukariotyczne
chromosom Archea jest podobnie jak u bakterii kolistą, pojedynczą cząsteczką DNA nie otoczoną błoną, lecz często mniejszą niż np. u E. coli
inny niż u bakterii skład ściany komórkowej i błony
- lipidy błony cytoplazmatycznej u Archea nie są związane estrowo (jak u bakterii i eukariota), ale są połączone z glicerolem eterowo, ponadto są długołańcuchowe i mogą być rozgałęzione
osłony (ściana komórkowa) Archea nie zawierają peptydoglikanu, występuje tu tzw. pseudomureina, zawierająca zamiast kwasu N-acetylomuraminowego, kw N-acetylotalozoaminouronowy, ponadto występuje tu warstwa S będąca dwuwymiarowym, parakrystalicznym układem białek lub glikoprotein na powierzchni komórek, u niektórych Archeabacterii na zewnątrz błony cytoplazmatycznej występuje gruba warstwa polisacharydowa
Wyszukiwarka
Podobne podstrony:
spr 2, biotechnologia inż, sem2, MO
MO lab4, biotechnologia inż, sem2, MO
analiza sanitarna popr, biotechnologia inż, sem2, MO
MO, biotechnologia inż, sem2, MO
met ozn mikroorg2, biotechnologia inż, sem2, MO
wymagania MO, biotechnologia inż, sem2, MO
SPRAWOZDANIE Z LABOLATORIUM Z FIZYKI I BIOFIZYKI cw.5, biotechnologia inż, sem2, FiB, laborki, spraw
MS Cz 10 A 1 5, biotechnologia inż, sem2, MŚ
Wnioski Stokes, biotechnologia inż, sem2, FiB, laborki, sprawka
MS Cz 10 A 1 4, biotechnologia inż, sem2, MŚ
Sprawozdanie STOCK, biotechnologia inż, sem2, FiB, laborki, sprawka
Sprawozdanie BERNULLI-1, biotechnologia inż, sem2, FiB, laborki, sprawka
Dyfrakcja, biotechnologia inż, sem2, FiB, laborki, sprawka
MS Cz 10 A 1 6, biotechnologia inż, sem2, MŚ
MS Sr 14 B 4 4, biotechnologia inż, sem2, MŚ
Dyfuzja, biotechnologia inż, sem2, FiB, laborki, sprawka
tarcie, biotechnologia inż, sem2, FiB, laborki, sprawka
Przewodniki, biotechnologia inż, sem2, FiB, laborki, sprawka
więcej podobnych podstron