Ewolucja Hedgehoga
Autor tekstu: PZ Myers
Tłumaczenie: Małgorzata Koraszewska
Public Library of Science opublikowała niedawno wysoce spekulacyjną, ale bardzo ciekawą
pracę o tym, jak mógł wyewoluować szlak sygnałowy Hedgehoga. Jest to stosunkowo wczesny etap
testowania hipotezy i to właśnie powoduje (między innymi), że ta praca jest interesująca —
zazwyczaj widzimy jedynie końcowy wynik długiego zbierania danych, eksperymentowania
i sprawdzania, co znaczy, że literatura naukowa daje nieco skrzywiony obraz procesu naukowego,
pozwalając postronnemu obserwatorowi zobaczyć jedynie to, co zostało już wyklepane
i wypolerowane, ukrywa zaś surowsze szkice, które pozwoliłyby nam zobaczyć, jak to się zaczęło
i jak nabudowywało. Jest to pouczające szczególnie dla tych, którzy przeglądają „literaturę"
kreacjonistów, często prymitywnie małpującą produkty rzeczywiście działającej nauki, ale bez
solidnej podbudowy metodologicznej. W szczególności u kreacjonistów zupełnie nie ma procesu
grzebania w rzeczywistym świecie, żeby rozwijać pomysły, ponieważ zaczynają oni od wniosków.
Potraktujmy więc ten opis jako pracę w procesie poszukiwań — obserwujemy tu dynamikę
budowania dobrego, działającego modelu. Jak zwykle na solidnych podstawach potwierdzonych,
znanych dowodów stawia się rozsądną hipotezę opartą na faktach, a następnie proponuje szereg
dróg badawczych z przewidzianymi wynikami, które potwierdziłyby hipotezę.
Hedgehog jest niezwykle ważną rodziną cząsteczek sygnałowych w wielokomórkowych
zwierzętach. Jak można oczekiwać, jeśli jesteś wielokomórkowym zwierzęciem, twoje komórki
potrzebują mechanizmu na wzajemne przekazywanie sobie informacji podczas procesu rozwojowego
— komórki w tkance muszą pracować razem jako zespół i potrzebują sposobów na koordynowanie
tej pracy. Potrzebny jest im sposób na mówienie jednej komórki do drugiej: „Słuchajcie, jesteśmy
wszyscy częścią śledziony! Zbudujmy wspierające struktury dla krwinek!" W rozwoju potrzebne są
one także do wzorcowania; na przykład niektóre cząsteczki Hedgehog są używane do określania
miejsca linii środkowej zwierzęcia, żeby narządy budowały się bilateralnie. Zabierz Hedgehoga,
a zwierzę traci wyczucie tej ważnej linii dzielącej i buduje jedno oko zamiast dwóch, i umieszcza je
tam, gdzie go nie powinno być.
Sama cząsteczka Hedgehog jest informacją wysłaną do innych komórek, by rozpoczęły zmiany
w ekspresji genów w komórkach docelowych. Potrzebna do tego jest komórka źródłowa
z maszynerią do wydzielania informacji, cząsteczki (Hedgehog) o unikatowym charakterze, która
stworzy z tego specjalny klucz, receptor lub czujnik w komórce docelowej oraz efektor, który może
być modyfikowany przez czujnik i który może następnie modyfikować ekspresję genu (zazwyczaj
dzięki kaskadzie cząsteczek pośredniczących), aby zmienić wzór rozwoju komórki. Tutaj jest
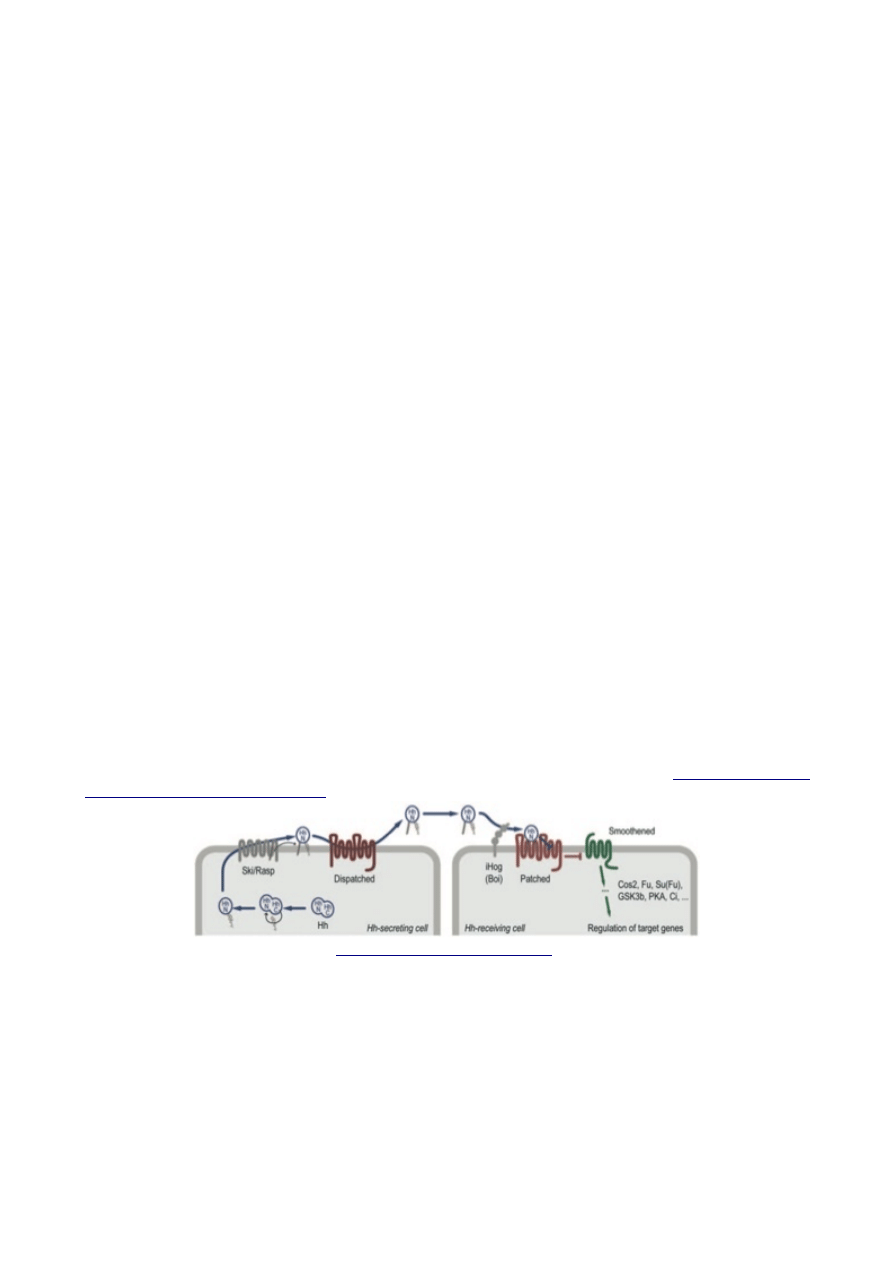
schemat tego procesu dla Hedgehoga, a jeśli to nie starczy, proszę przeczytać
Schematic overview of the Hedgehog signaling pathway. Signal-secreting cells (left)
release the morphogen protein Hh after modifying it through the addition of two lipid
molecules. A C-terminal cholesterol moiety is added via the activity of an intein
domain within Hh itself, whereas the protein Ski/Rasp attaches an N-terminal palmitic
acid. Lipid-modified Hh is released from the producing cell with the aid of the Disp
protein. Signal-receiving cells (right) bind Hh via the transmembrane protein Ptc,
perhaps with the assistance of the iHog/Boi family of proteins. Hh binding to Ptc leads
to the de-repression of the GPCR-related protein Smo. Smo subsequently initiates
intracellular signal transduction events, which involve proteins such as Cos2, Fu, and
Su(fu), that lead to changes in target gene expression. The inhibition of Smo by Ptc is
Racjonalista.pl
Strona 1 z 5

of particular interest here; it occurs nonstoichiometrically, in a manner that appears to
rely on a catalytic activity in Ptc.
Tutaj jest obsada ról, żeby pomóc wszystko uporządkować:
ˇ Po stronie wydzielania jest zestaw cząsteczek zaangażowanych w eksport Hedgehoga,
a także w modyfikowanie białka; jednym z głównych aktorów jest cząsteczka Dispatched. Nie
powiem nic więcej o tej części szlaku, ale proszę zauważyć podobieństwa między Dispatched i
Patched:
są
one
częścią
tej
samej
rodziny.
ˇ Hedgehog jest informacją. Jest to małe białko z przyczepionym lipidem, cholesterolem. Jest
wydzielane do przestrzeni międzykomórkowej, skąd może przenikać do sąsiadujących komórek.
ˇ Patched jest receptorem. Kiedy wiąże Hedgehoga nie wchodzi już w reakcję z następną cząsteczką
na szlaku, Smoothendem. Kiedy nie wiąże Hedgehoga, hamuje Smoothend, nie pozwalając mu na
wykonanie
jego
zadania.
ˇ Smoothend jest efektorem. Kiedy Patched mu nie przeszkadza, włącza łańcuch białek, które wiodą
do aktywacji określonych genów w jądrze.
Jest to rodzaj podwójnie negatywnej regulacji: Hedgehog hamuje Patched, powstrzymując go
od hamowania Smoothend, który włącza geny. Jest to także skomplikowana maszyna molekularna
w stylu Rube Goldberg, która powierzchownie robi wrażenie, jakby było bardzo trudno ją
wyewoluować — właśnie tego rodzaju, którym uwielbiają zanudzać kreacjoniści. Nic jednak nie
przychodzi z nikąd i okazuje się, że każda jej część ma bardzo ciekawych ewolucyjnych
poprzedników.
Zacznijmy od Patched. To białko jest prastare; spokrewnionych członków tej rodziny genów
możemy znaleźć w bakteriach prokariotycznych, które oczywiście nie są wielokomórkowe i nie są
zwierzętami. Co robi ono w bakteriach? Jest to transporter, mała pompa, której komórka używa do
wydzielania nadmiaru produktów ubocznych metabolizmu, a szczególnie klasy lipidów o nazwie
hopanoidy, które są cząsteczkami podobnymi do sterydów. Nad waszymi głowami powinno się teraz
zapalić małe światełko: pamiętamy, że Dispatched jest zaangażowany w wydzielanie Hedgehoga,
który jest kowalentnie modyfikowany przez dodatek podobnej do sterydu cząsteczki.
A co z Smoothend? Także jest członkiem wybitnej rodziny i ma cechy wspólne z receptorami
sprzężonymi z białkiem G (GPCR). Te cząsteczki są wszędzie; wspomniałem ich rolę w przenoszeniu
sygnałów
w
(http://scienceblogs.com/pharyngula/2006/09/rhabdome
ric_and_ciliary_eyes.php) i
(http://scienceblogs.com/pharyngula/2006/09/cats_can
dy_and_evolution.php). Zasadniczo są to cząsteczki w błonie komórkowej, które potrafią wykryć
bodziec i uruchomić kaskadę aktywacji białek, prowadzącą do zmian w metabolizmie komórkowym.
To tyle i ta hipoteza powinna być zrozumiała. Ewolucja zmieszała i dopasowała kawałki
z różnych szlaków, żeby zbudować wyrafinowany szlak sygnalizacyjny w szeregu małych, łatwych
kroków, z których każdy dawał organizmowi sensowną korzyść. Tutaj jest hipotetyczna ilustracja
jego ewolucji.
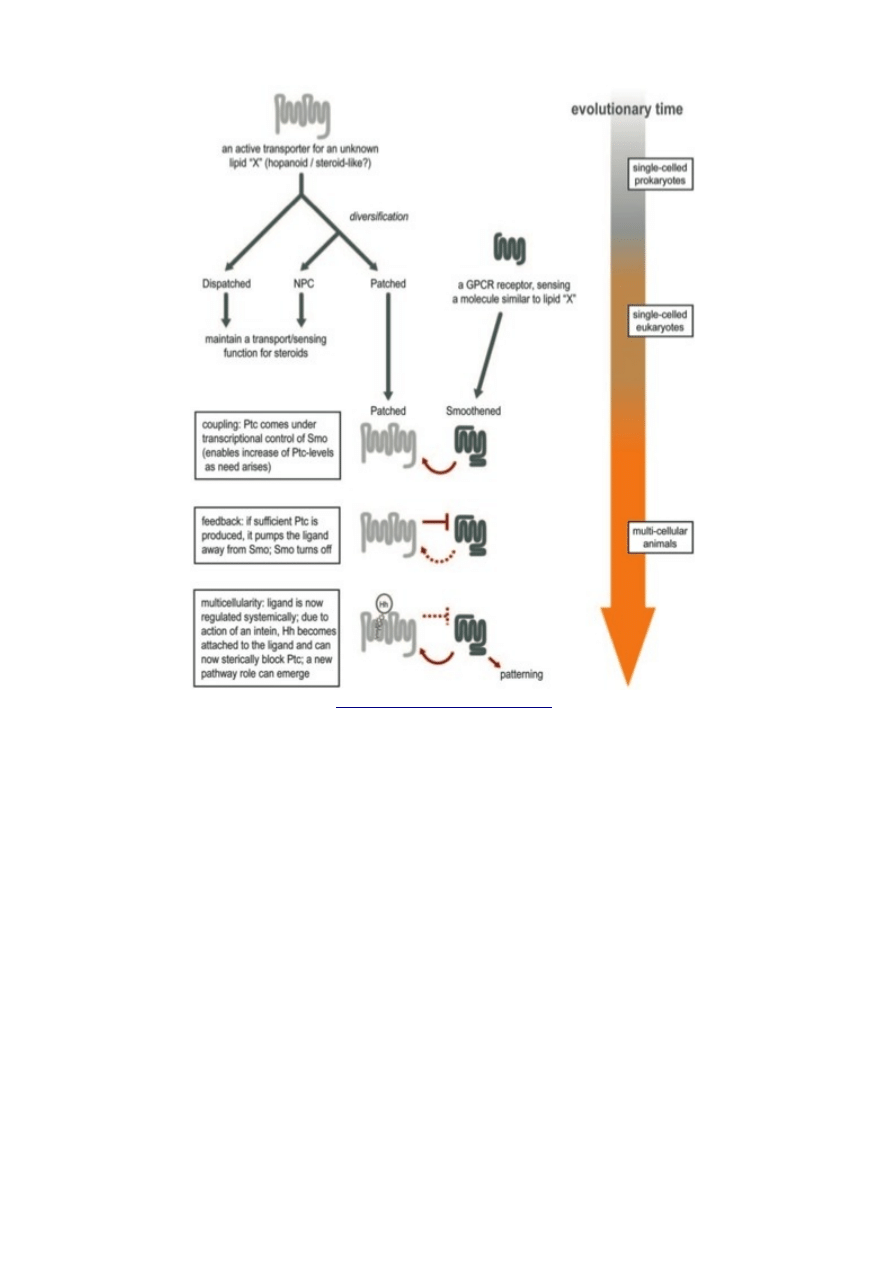
A parsimonious scenario for the evolution of the Ptc/Smo system. We hypothesize that
during the transition to multicellularity, a pre-existing lipid homeostasis system took
on a new function in signaling. Initially, an ancient lipid transporter diversified; one of
its descendents came under the transcriptional control of a GPCR that sensed the
same lipid (i.e., forming a negative homeostatic feedback loop). Then, the fortuitous
addition of a protein moiety to the lipid in question brought the system under the
control of gene expression; a neighboring cell could now secrete the lipid at will (by
coupling it to the protein moiety). Because the combined lipid-protein molecule would
block the transporter, this meant that the sending cell was capable of changing the
perceived homeostatic state of the receiving cell, which would have established
a graded (quantitative) mode of cell-cell communication.
To jest proste. Prokarioty miały białko Patched do wypompowywania lipidu X; miały także
GPCR Smoothend, który potrafił wyczuć lipid X i który odgrywał rolę w regulacji genów
zaangażowanych w metabolizm X.
Jednym z genów zaangażowanych w metabolizm X, regulowanym przez Smoothend , jest
Patched. To jest krok łączący i ma on sens z punktu widzenia doboru. Jeśli komórka pływa w wielkiej
ilości X, włącz białko, które pomoże pozbyć się nadmiaru X.
Następnym krokiem jest dodanie sprzężenia zwrotnego. Jeśli wytwarzane jest mnóstwo
cząsteczek Patched, byłoby wydajne zatrzymanie tej produkcji, tak więc ewolucja faworyzuje
ewolucyjne powiązanie między Patched i Smoothend. Zapobiegnie to nieokiełznanej produkcji białek
w tym konkretnym szlaku ponad to, co jest potrzebne.
Końcowy krok pojawia się, kiedy szlak zostaje dokooptowany w ewolucji wielokomórkowości.
Racjonalista.pl
Strona 3 z 5

Lipid X jest prymitywnym sygnałem powiększenia szlaku komórkowego do pewnego poziomu
i zastopowania go; komórki mogą użyć tego sygnału, żeby oszukać swoje sąsiadki przez zalewanie
ich X, powodując włączenie u nich określonego zestawu genów. Jeszcze sprytniejsze jest to, że przez
modyfikowanie X kowalentnie związanym białkiem (co powstrzymuje komórkę od przetwarzania X)
mogą wytworzyć silniejszy sygnał. Ta część wymaga niejakiego zwrotu postrzegania; jesteśmy
stronniczy wobec białek i zazwyczaj uważamy białko za ważniejszą część, modyfikowaną przez
dodanie lipidu… to zaś sugeruje, że pierwotnie lipid był sygnałem, a białko dodatkiem.
Podoba mi się to. To ładny model i ma wiele sensu. Bazuje także na ustalonych wzorcach
homologii i funkcji molekularnej, nie jest więc po prostu wzięty z powietrza — istnieją dobre powody,
by sądzić, że jest to rozsądne wyjaśnienie.
Nauka nie zatrzymuje się tutaj, zadowalając się zaledwie prawdopodobieństwem. Następnym
krokiem jest przetestowanie idei i autorzy czynią cały szereg przewidywań tego, co powinni
zobaczyć, jeśli ich idea jest słuszna.
1. Powinny istnieć głębsze podobieństwa między transporterami bakteryjnymi i Patched.
Wiadomo, że białka bakteryjne funkcjonują jako trimery (trzy identyczne białka muszą się
zgrupować, żeby wykonać zadanie), a także używają gradientu stężenia protonów do napędzania
swojej
akcji
pompującej.
Czy
Patched
ma
podobne
wymagania?
2. Czy w podobny sposób można modyfikować funkcjonalnie bakteryjne transportery przez dodanie
blokujących
białek
do
ich
pożywek?
3. U zwierząt wielokomórkowych Patched może nadal funkcjonować pod nieobecność cholesterolu u
Hedgehoga; nabył mianowicie specjalnego powinowactwa do białka, a nie do lipidu. Czy w Patched
może jednak nadal istnieć szczątkowe miejsce wiążące dla cholesterolu?
4. Istnieją pewne ciekawe filogenetyczne dziwactwa, które powinno się zbadać. Na przykład nicienie
mają Patched, ale nie mają Smoothend. Autorzy przewidują, że Patched u C. elegant powinien być
w stanie współdziałać z Smoothend muszki owocowej. (Moim zdaniem jest to trochę ryzykanckie
przewidywanie: nicienie mogły utracić Smoothend i teza, że ich szlak reprezentuje krok pośredni
w ewolucji
tej
ścieżki,
jest
trochę
niebezpieczna).
5. Autorzy stawiają również hipotezę dotyczącą dokładnego mechanizmu funkcji
Patched/Smoothend. Ponieważ przypuszcza się, że Patched pochodzi z pompy, która odpycha
cząsteczkę od komórki, powinna istnieć ściśle umiejscowiona interakcja między tymi dwoma
białkami — nie powinny działać tak, że Patched przenosi do Smoothendu pośredniczącą, małą
cząsteczkę.
6. Jedną z propozycji, jak Patched i Smoothend mogą wchodzić ze sobą w interakcję, jest lokalna
dynamika lipidów — Patched zmienia środowisko lipidowe na swoim małym obszarze błony,
wysysając jakiś specjalny składnik, a Smoothend reaguje na tę zmianę.
To wszystko jest ciekawe, z masą interesujących szczegółów do rozplątania i zanalizowania.
Widzę także, że jest to program badawczy, który wykorzystuje biochemię, biologię molekularną,
bakteriologię i biologię porównawczą, żeby odpowiedzieć na ciekawe pytania biologii rozwojowej —
mam wrażenie, że będę musiał wrócić do szkoły, żeby odświeżyć wiedzę o podstawach, by móc
dotrzymać kroku w mojej dziedzinie.
Proszę także zauważyć, co jest ważne w uprawianiu nauki. To jest całkowicie spekulacyjny
model ewolucji konkretnego szlaku, ale wymaga on: 1) uznania istniejącej informacji, 2) planu
sprawdzania hipotezy i rozszerzenia wiedzy. Może być w wielu punktach błędny, ale proces
rozumienia, gdzie zawiódł, pomoże dopracować następny model.
Źródło: Hausmann, G, von Mering C, Basler K, (2009) The Hedgehog Signaling Pathway:
Where Did It Come From? „PLoS Biol" 7(6): e1000146.doi:10.1371/journal.pbio.1000146
(http://scienceblogs.com/pharyngula/2009/07/the_evol ution_of_hedgehog.php).
Pharyngula, 21 lipca 2009
Ur 1957. Amerykański profesor biologii na uniwersytecie w Minnesocie, prowadzi również
(Publikacja: 26-07-2009)

(http://www.racjonalista.pl/kk.php/s,6701)
Contents Copyright
©
2000-2009 Mariusz Agnosiewicz
Programming Copyright
©
2001-2009 Michał Przech
Autorem portalu Racjonalista.pl jest Michał Przech, zwany niżej Autorem.
Właścicielami portalu są Mariusz Agnosiewicz oraz Autor.
Żadna część niniejszych opracowań nie może być wykorzystywana w celach
komercyjnych, bez uprzedniej pisemnej zgody Właściciela, który zastrzega sobie
niniejszym wszelkie prawa, przewidziane
w przepisach szczególnych, oraz zgodnie z prawem cywilnym i handlowym,
w szczególności z tytułu praw autorskich, wynalazczych, znaków towarowych
do tego portalu i jakiejkolwiek jego części.
Wszystkie strony tego portalu, wliczając w to strukturę katalogów, skrypty oraz inne
programy komputerowe, zostały wytworzone i są administrowane przez Autora.
Stanowią one wyłączną własność Właściciela. Właściciel zastrzega sobie prawo do
okresowych modyfikacji zawartości tego portalu oraz opisu niniejszych Praw
Autorskich bez uprzedniego powiadomienia. Jeżeli nie akceptujesz tej polityki możesz
nie odwiedzać tego portalu i nie korzystać z jego zasobów.
Informacje zawarte na tym portalu przeznaczone są do użytku prywatnego osób
odwiedzających te strony. Można je pobierać, drukować i przeglądać jedynie w celach
informacyjnych, bez czerpania z tego tytułu korzyści finansowych lub pobierania
wynagrodzenia w dowolnej formie. Modyfikacja zawartości stron oraz skryptów jest
zabroniona. Niniejszym udziela się zgody na swobodne kopiowanie dokumentów
portalu Racjonalista.pl tak w formie elektronicznej, jak i drukowanej, w celach innych
niż handlowe, z zachowaniem tej informacji.
Plik PDF, który czytasz, może być rozpowszechniany jedynie w formie oryginalnej,
w jakiej występuje na portalu. Plik ten nie może być traktowany jako oficjalna
lub oryginalna wersja tekstu, jaki zawiera.
Treść tego zapisu stosuje się do wersji zarówno polsko jak i angielskojęzycznych
portalu pod domenami Racjonalista.pl, TheRationalist.eu.org oraz Neutrum.eu.org.
Wszelkie pytania prosimy kierować do
Racjonalista.pl
Strona 5 z 5
Wyszukiwarka
Podobne podstrony:
Ewolucja marketingu era produkcyjna, sprzedazowa, marketingowa Rynek definicja
Systemy teoretyczne socjologii naturalistycznej – pozytywizm, ewolucjonizm, marksizm, socjologizm pp
Ewolucja wszechśwaita i kosmologii
Ewolucja nowe
ewolucja integracji europejskiej 2011
Dowody za obiektywno¶ci± ewolucji z zakresu morfologii porównawczej 1 cz
Ewolucja techniki sekcyjnej – od Virchowa do Virtopsy®
Historyczne uwarunkowania ewolucji E coli
powiązania ewolucyjne t antytetyczna
ewolucja2
23 Argasinski, Metody teorii gier ewolucyjnych(2009)
Powstanie i ewolucja zycia
ERY, ewolucjonizm
więcej podobnych podstron