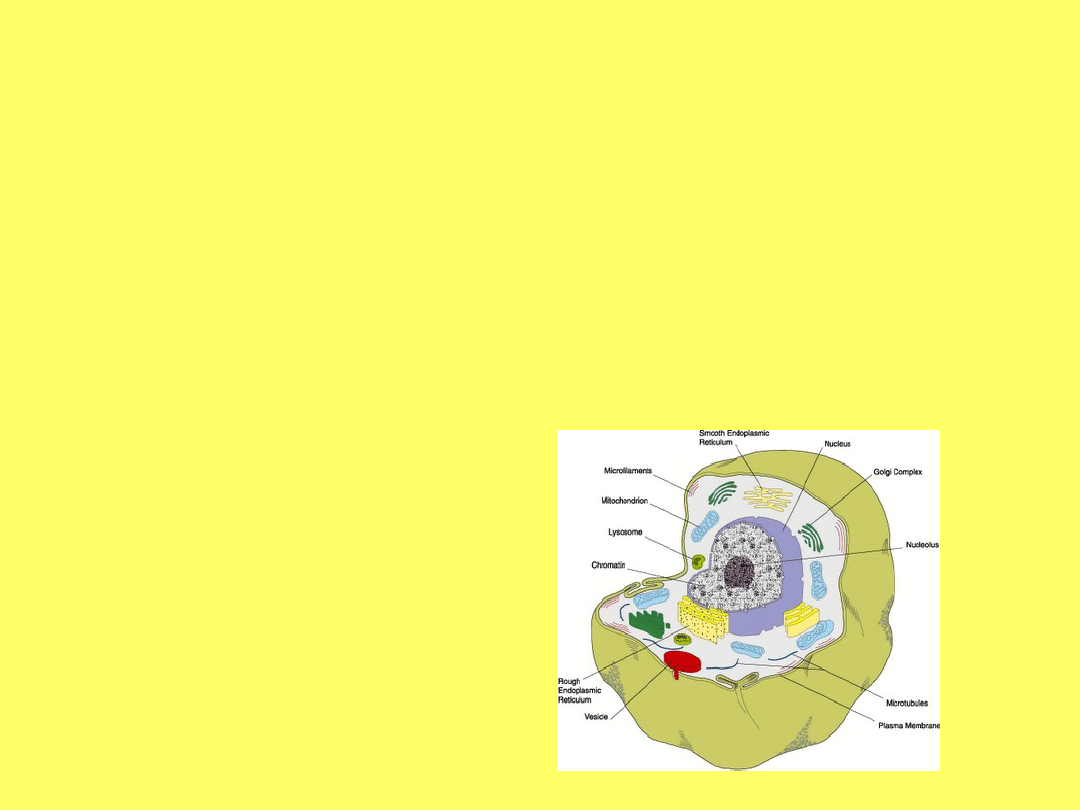
RECEPTORY BŁONY
KOMÓRKOWEJ I ORGANELLI
- BUDOWA I FUNKCJE
Szkudlarek Natalia
Przybyla Ewa
Grzelak Ilona
Marek Karolina

RECEPTORY BŁONOWE
• Bardzo ważną rolę w odbiorze informacji przez komórki
odgrywają białka receptorowe. Syntetyzowane są w
siateczce śródplazmatycznej ziarnistej. Przyczepiają się do
wewnętrznej powierzchni błony komórkowej i następnie w
procesie eksternalizacji białek receptorowych przenoszone
są na zewnętrzną powierzchnię. Po związaniu się
przekaźników chemicznych (transmitery, hormony, i inne
substancje) występujące w płynie zewnątrzkomórkowym z
receptorami na zewnętrznej powierzchni błony komórkowej
białka receptorowe oddziałują na inne białka błony
komórkowej. Wówczas białka nośnikowe przyśpieszają lub
zwalniają aktywny transport przez błony komórkowe,
otwierają się lub zamykają kanały dla dokomórkowych i
dokomórkowych prądów jonowych oraz następuje
aktywacja lub inaktywacja białek enzymatycznych.

• Białka receptorowe nie tylko oddziałują na
inne białka błony komórkowej, ale również
same transportują cząsteczki chemiczne z
płynu zewnątrzkomórkowego do cytoplazmy.
Cząsteczki insuliny, białka wytwarzane przez
komórki B wysp trzustkowych, po związaniu
się z receptorami insulinowymi na
zewnętrznej powierzchni błony komórkowej
dostają się do wnętrza komórki w procesie
internalizacji receptorów.

• Receptory błonowe mają zdolność swoistego wiązania się z
określonymi substancjami chemicznymi, zwanymi ligandami,
endogennymi oraz egzogennymi. W błonie komórkowej
występują białka receptorowe o najwyższym powinowactwie do
licznych substancji. Aktywne substancje chemiczne
występujące w płynie zewnątrzkomórkowym konkurują ze sobą
o miejsce wiązania się z receptorem. Z kilku substancji
chemicznych o podobnej budowie wiążą się z receptorem
cząsteczki tej substancji, która ma największe powinowactwo
do tego receptora. Może być niewiele receptorów wiążących
jedną substancję, w stosunku do jednostki powierzchni błony
komórkowej i nawet przy małym stężeniu cząsteczek w płynie
zewnątrzkomórkowym wszystkie receptory mogą być
połączone z ligandem. Dalsze zwiększenia stężenia cząsteczek
w płynie zewnątrzkomórkowym nie będzie miało przestają one
wiązać się z substancjami endogennymi. W ten sposób
receptory w błonie komórkowej wpływu na jej działanie na
komórkę, ze względu na małą pojemność wiązania.

• Po zadziałaniu na błonę komórkową
substancji egzogennych blokujących
określone receptory zostają na krótszy
lub dłuższy czas zablokowane i komórka
przestaje odbierać informacje za
pośrednictwem określonego receptora.
Biologicznie aktywne substancje
egzogenne wykazujące powinowactwo do
receptorów błonowych są antagonistami
określonych substancji endogennych.

ODBIOR INFORMACJI PRZEZ KOMÓRKĘ
Informacja odbierana przez receptory w błonie komórkowej
wywołuje reakcje:
-krótkotrwałe ( od ułamka milisekundy do kilkudziesięciu
milisekund) związane z ruchem jonów przez błonę komórkową, z
czym wiąże się z kolei wiąże się występowanie potencjału
elektrycznego;
-dłużej trwające (od milisekund do wielu godzin) w wyniku
aktywacji lub inaktywacji enzymów w błonie komórkowej i
wzmożenia lub osłabienia metabolizmu komórkowego;
-długotrwale (od wielu godzin do wielu tygodni ) kiedy dochodzi
ekspresji genów w jądrze komórkowym, w których zakodowana
jest informacja genetyczna potrzebna do syntezy enzymów
zmieniających metabolizm komórkowy.

Odbieranie i przetwarzanie informacji przez błonę
komórkową przebiega w trzech etapach:
• Rozróżnianie sygnałów – informację odbieraną stanowią
niezależne od siebie sygnały, czyli cząsteczki przekaźników
chemicznych, działające niezależnie od siebie na błonę
komórkową. Receptory błonowe odróżniają cząsteczki na nie
działające, np. cząsteczki acetylocholiny od cząsteczek
noradrenaliny;
• Przenoszenie sygnałów – białka receptorowe związane ze
swoistymi przekaźnikami chemicznymi oddziałują na białka
kanałów jonowych i białka enzymatyczne;
• Wzmacnianie sygnałów – sygnały odebrane przez komórkę,
czyli na wejściu układu, są zazwyczaj zbyt słabe, aby mogły
wpływać istotnie na metabolizm wewnątrzkomórkowy. Muszą
zostać odpowiednio wzmocnione. Takimi uniwersalnymi
wzmacniaczami licznych sygnałów odbieranych przez komórki
są enzymy wytwarzające drugi wewnątrzkomórkowy
przekaźnik chemiczny.

Drugi przekaźnik wewnątrzkomórkowy
• Krążące w płynach ustrojowych fizjologicznie aktywne,
endogenne substancje chemiczne, przede wszystkim hormony,
stanowią pierwszy przekaźnik informacji. Po odebraniu
przez receptory w błonie komórkowej informacji i jej
przeniesieniu dochodzi do aktywacji lub inaktywacji enzymów i
do zwiększenia lub zmniejszenia w komórkach stężenia
drugiego przekaźnika informacji.
• Do wewnątrzkomórkowych przekaźników informacji zalicza się:
cykliczny adenozynomonofosforan (cAMP), cykliczny
guanozynomonofosforan (cGMP), inozytolotrifosforan (IP3),
diacyloglicerol (DAG) i jony wapnia (Ca+).
• Pod wpływem cyklazy adenylanowej, występującej po
wewnętrznej stronie błony komórkowej, odczepiają się
pirofosforany od adenozynotrifosforanów i powstaje cykliczny
adenozyno-3’,5’-monofosforan, czyli cykliczny AMP .

• Krążące w płynach ustrojowych fizjologicznie aktywne,
endogenne substancje chemiczne, przede wszystkim
hormony, stanowią pierwszy przekaźnik informacji. Po
odebraniu przez receptory w błonie komórkowej informacji i
jej przeniesieniu dochodzi do aktywacji lub inaktywacji
enzymów i do zwiększenia lub zmniejszenia w komórkach
stężenia drugiego przekaźnika informacji.
• Do wewnątrzkomórkowych przekaźników informacji zalicza
się: cykliczny adenozynomonofosforan (cAMP), cykliczny
guanozynomonofosforan (cGMP), inozytolotrifosforan (IP3),
diacyloglicerol (DAG) i jony wapnia (Ca+).
• Pod wpływem cyklazy adenylanowej, występującej po
wewnętrznej stronie błony komórkowej, odczepiają się
pirofosforany od adenozynotrifosforanów i powstaje
cykliczny adenozyno-3’,5’-monofosforan, czyli cykliczny
AMP .

• W przenoszeniu sygnałów pomiędzy receptorem a
związanym z ligandem i enzymem pośredniczą białka
błonowe G występujące w wewnętrznej warstwie błony
komórkowej. Ich właściwością jest zdolność wiązania się i
hydrolizowania guanozynotrifosforanów. Białka G składają
się z trzech podjednostek β, α i γ. W zależności od typu
białka G, działają one aktywująco lub inaktywująco na
cyklozę adenylanową. Po odebraniu sygnału od receptora w
białku aktywującym Gs następuje zmiana związanego z
białkiem Gs guaznozynodifosforanu (GDP) na
guanozynotrifosforan (GTP), odłączenie się jednostek β i γ i
połączenie podjednostki α z cyklozą adenylanową, pod
której wpływem tworzy się cAMP. Uwolnienie ligandu od
receptora powoduje odwrotny proces – zamianę związanego
z białkiem Gs guanozynotrifosforanu na
guanozynodifosforan, odłączenie się ze sobą trzech
podjednostek białka G s , czyli powrót do stanu spoczynku.
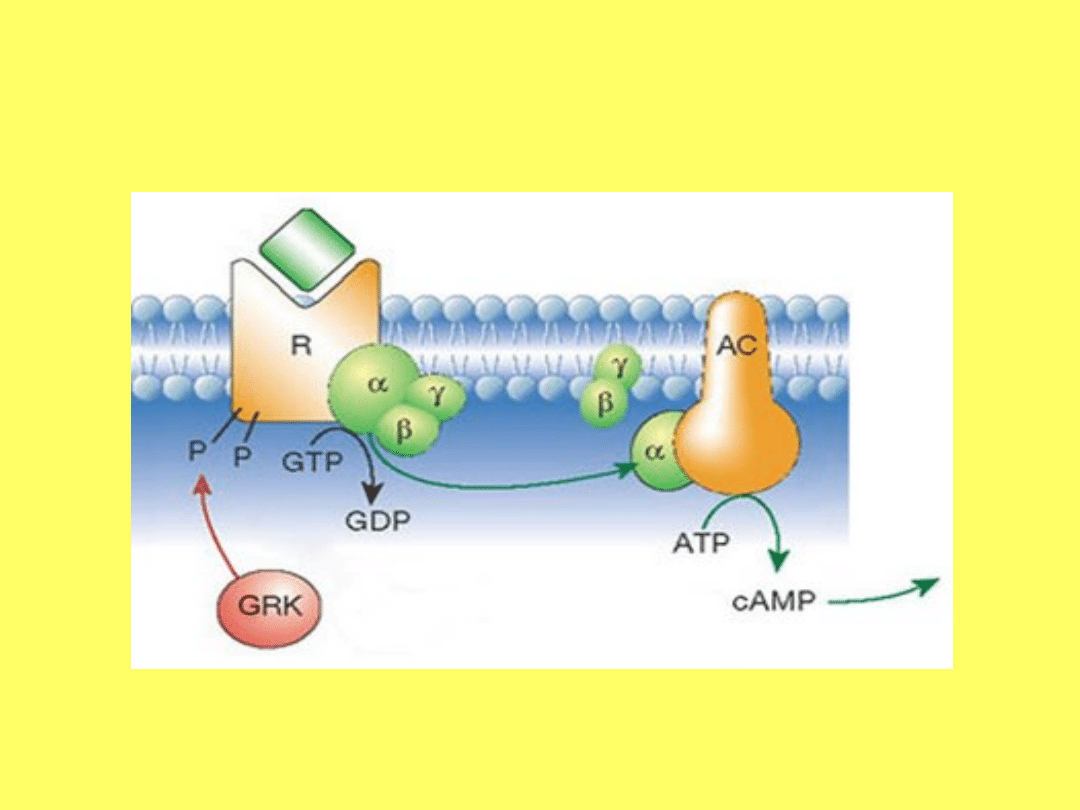

• Związanie się innego ligandu z innym receptorem
wywołuje wytwarzanie się po wewnętrznej stronie
błony komórkowej inozytolotrifosforanu i
diacyloglicerolu. Receptory związane z ligandem
aktywują fosfolipazę C, która hydrolizuje 4,5-
difosforan fosfatyloninzytolu do
inozytolotrifosforanu (IP3) i diacylkoglicerolu (DAG).
W przenoszeniu sygnałów pomiędzy receptorem i
fosfolipazą C pośredniczy również białko błonowe
G. Inozynotrifosforan wywołuje w komórce przepływ
jonów wapnia z siateczki śródplazmatycznej do
cytoplazmy, diacyloglicerol aktywuje zaś kinazę
białkową C i zwiększa metabolizm komórki.

• Jony wapnia uważane są również za „drugi przekaźnik”
wewnątrzkomórkowej informacji. W płynie
zewnątrzkomórkowym (ECF) jony Ca2+ występują jako jony
wolne lub związane z zewnętrzną powierzchnią błony
komórkowej. Do komórek napływają co najmniej trzy różne
kanały dla dokomórkowego ich prądu. Wolne jony Ca2+ po
wejściu do komórki wiążą się z błoną komórkową po stronie
wewnętrznej oraz z błonami siateczki śródplazmatycznej i
błonami mitochondrialnymi, jak również z białkiem
występującym w cytoplazmie – kalmoduliną, zbudowana
ze 148 aminokwasów (16 700m.cz.). Kompleks kalmodulina
– jony wapnia przyłącza się do enzymów
wewnątrzkomórkowych, przede wszystkim do kinaz
białkowych i je aktywuje. Po wejściu do komórek jonów
Ca2+ następuje przyspieszenie egzocytozy, w tym również
uwalniania transmiterów z zakończeń komórek nerwowych.

Przetwarzanie i przenoszenie informacji
Zarówno w środowisku zewnętrznym jak i wewnętrznym
zachodzą stałe zmiany. Nieznaczna nawet zmiana w
środowisku jest źródłem sygnałów odbieranych przez
receptory.
Każdy sygnał niesie informacje ze środowiska zewnętrznego
lub wewnętrznego. W obrębie receptorów zachodzi
przetwarzanie informacji w sposób analogowy i cyfrowy, jak
również jest stale przenoszona – w procesie tym ilość
informacji się nie zmenia

Przenoszenie informacji w
sposób ciągły (analogowy)
Odbywa się np. za pośrednictwem
cząsteczek krążących we krwi
hormonów.
Proporcjonalnie do wzrostu koncentracji
cząsteczek hormonów w określonym
czasie wzrasta ilość przenoszonej
informacji.
Przenoszenie to zachodzi na drodze
hormonalnej.

Przetwarzanie i przenoszenie
informacji w sposób
przerywany (cyfrowy)
Zachodzi np. we włóknach nerwowych.
Narządy receptorowe spełniają funkcje
przetworników zmieniających informacje ze
środowiska zewnętrznego lub wewnętrznego
na salwy impulsów nerwowych
przewodzonych przez włókna nerwowe.
Poszczególne impulsy przewodzone przez o
samo włókno nerwowe (kanały informacyjne)
nie różnią się między sobą. Informacja
zakodowana jest w częstotliwości z jaką
impulsy nerwowe przewodzone są przez
włókno nerwowe.
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
Wyszukiwarka
Podobne podstrony:
Wykład 7 Biolkom Błony komórkowe i organelli
komórka, Organelle komórki, Organelle komórki
Blony komorkowe
09 Biochemia blony komorkowe
Struktura błony komórkowej, biochemia
błony komórkowe
BŁONY KOMÓRKOWE, Studia - materiały, semestr 6, Fizjologia żywienia
71 Potencjal blony komorkowej
BŁONY KOMÓRKI
Błony komórkowe, Fizjoterapia, Fizjologia
blony komorki
Transport przez błony komórki
BŁONY KOMÓRKOWE
0202 04 03 2009, wykład nr 2 , Budowa i funkcje błony komórkowej oraz transport przez błony(1)
1 cw BLONY KOMORKOWEid 9083 Nieznany (2)
więcej podobnych podstron