
BŁONY KOMÓRKOWE
ZŁOŻONE I ZORGANIZOWANE
UKŁADY ZBUDOWANE GŁÓWNIE Z
LIPIDÓW I BIAŁEK

ROLA BŁON
1. O
ddzielają komórki od środowiska
- wysoce selektywne bariery – specyficzne kanały, przenośniki, pompy
- zawierają specyficzne RECEPTORY BODŹCÓW ZEWNĘTRZNYCH – kontrola
przepływu informacji między komórką a środowiskiem – ruch
bakterii do pożywienia, reakcja na hormony, percepcja światła.
- generowanie sygnałów o charakterze chemicznym lub elektrycznym, np.
przy przenoszeniu bodźców nerwowych.
2.
Wytyczają granice organelli
– tworzą odrębne PRZEDZIAŁY w
komórce
3. W błonach zachodzą PROCESY PRZEKSZTAŁCANIA ENERGII,
wymagające uporządkowania enzymów i innych białek w specyficzne
sekwencje:
- fotosynteza
- fosforylacja oksydacyjna
4. Transport –
w obrębie retikulum endoplazmatycznego.

WSPÓLNE CECHY BŁON
1. Są strukturami warstwowymi, gr. ok. 6-10 nm
2. Zbudowane są gł. z lipidów i białek, których stosunek wagowy
sięga od 1:4 do 4:1. Zawierają także cukrowce związane z
lipidami i białkami
3. Lipidy błonowe są stosunkowo małymi cząsteczkami zawierającymi
reszty hydrofobowe i hydrofilowe.
4. W charakterystycznych funkcjach błon pośredniczą specyficzne
białka, które działają jako pompy, kanały, przenośniki,
receptory, transformatory energii i enzymy.
Białka błonowe mogą być zanurzone w dwuwarstwie lipidowej, która
tworzy właściwe środowisko dla działania tych białek.
5. Błony – zespoły cząsteczek związanych
NIEKOWALENCYJNIE
.
6. Błony są
ASYMETRYCZNE
i obie jej strony są zawsze różne.
7. Błony są strukturami
PŁYNNYMI
.
8. Większość błon jest
SPOLARYZOWANA ELEKTRYCZNIE
, z ładunkiem
ujemnym od strony wnętrza zamkniętego przedziału (zazwyczaj
– 60 mV). Potencjał błonowy ma kluczowe znaczenie w
procesach transportu, przekształceń energii i pobudliwości.
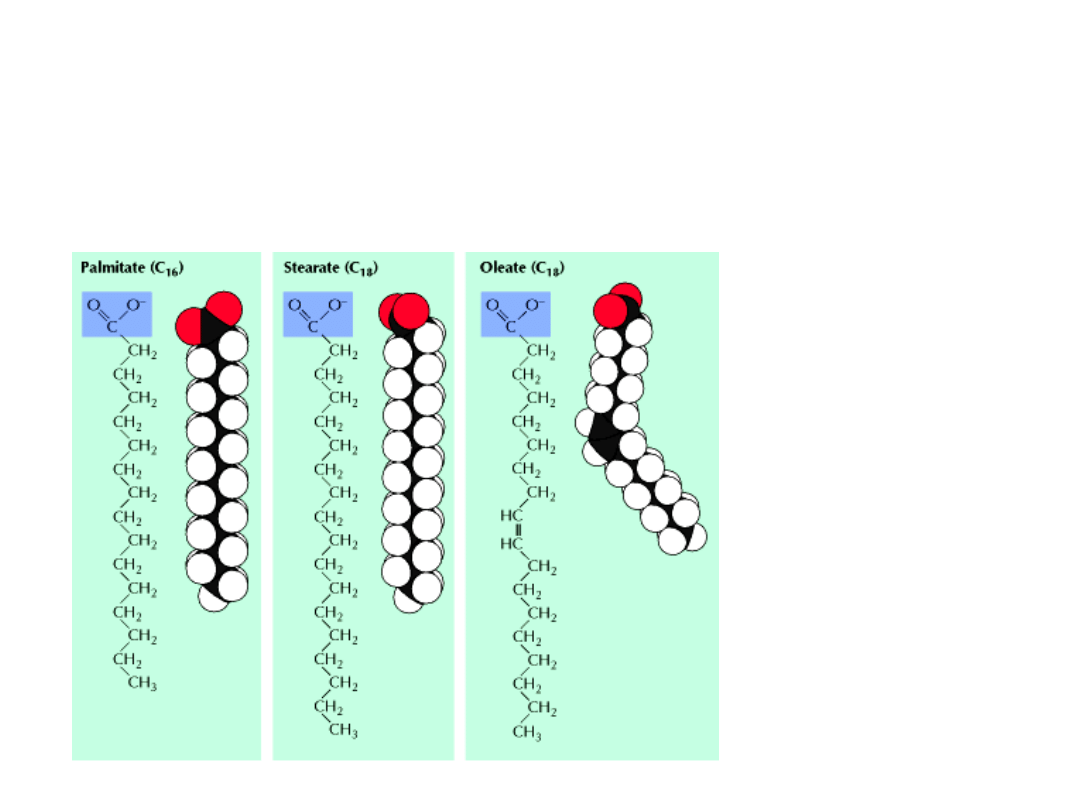
LIPIDY BŁONOWE
GŁÓWNYM składnikiem lipidów błonowych są
KWASY TŁUSZCZOWE
.
Są to łańcuchy węglowodorowe o różnej długości i stopniu nasycenia,
zakończonymi grupami karboksylowymi.
Łańcuchy kwasów tłuszczo-
wych zawierają parzystą liczbę
atomów węgla (14-24),
najczęściej 16 – 18.
Zwierzęce kw. tłuszczowe –
łańcuchy są nierozgałęzione.
Kwasy tłuszczowe mogą być
- nasycone
- nienasycone – wiązanie
podwójne przyjmuje prawie
zawsze konfigurację cis.
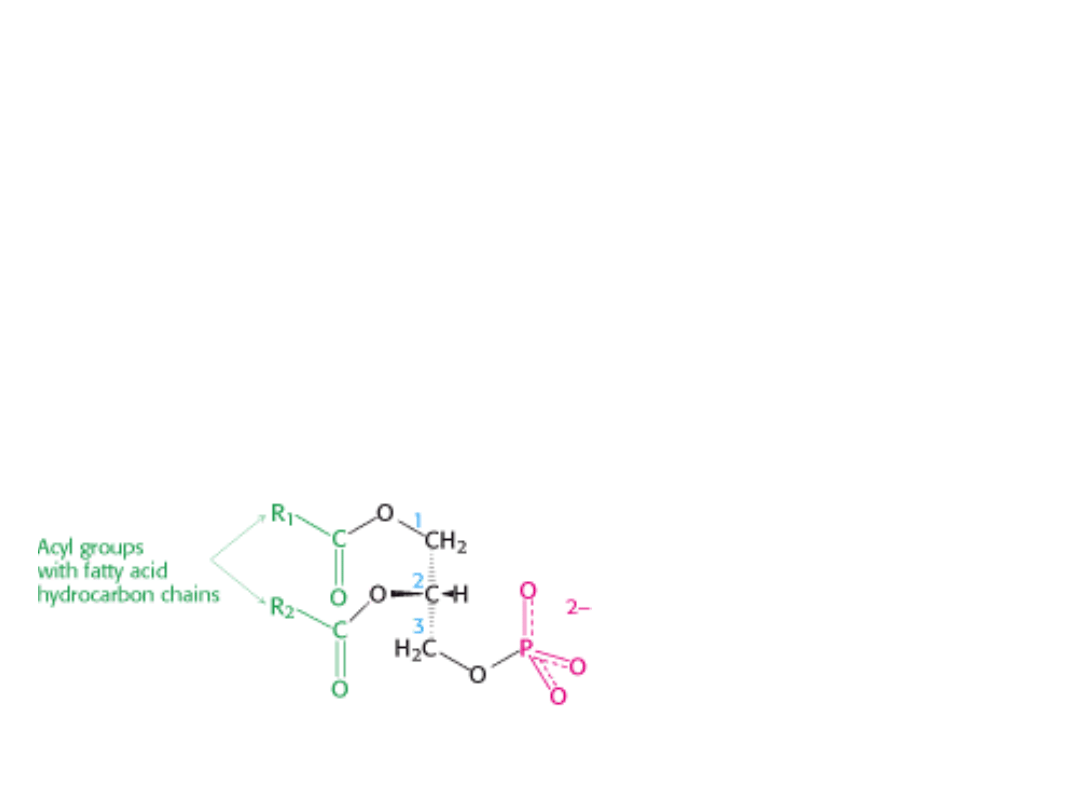
W błonach biologicznych występują 3
typy lipidów:
FOSFOLIPIDY,
GLIKOLIPIDY, CHOLESTEROL
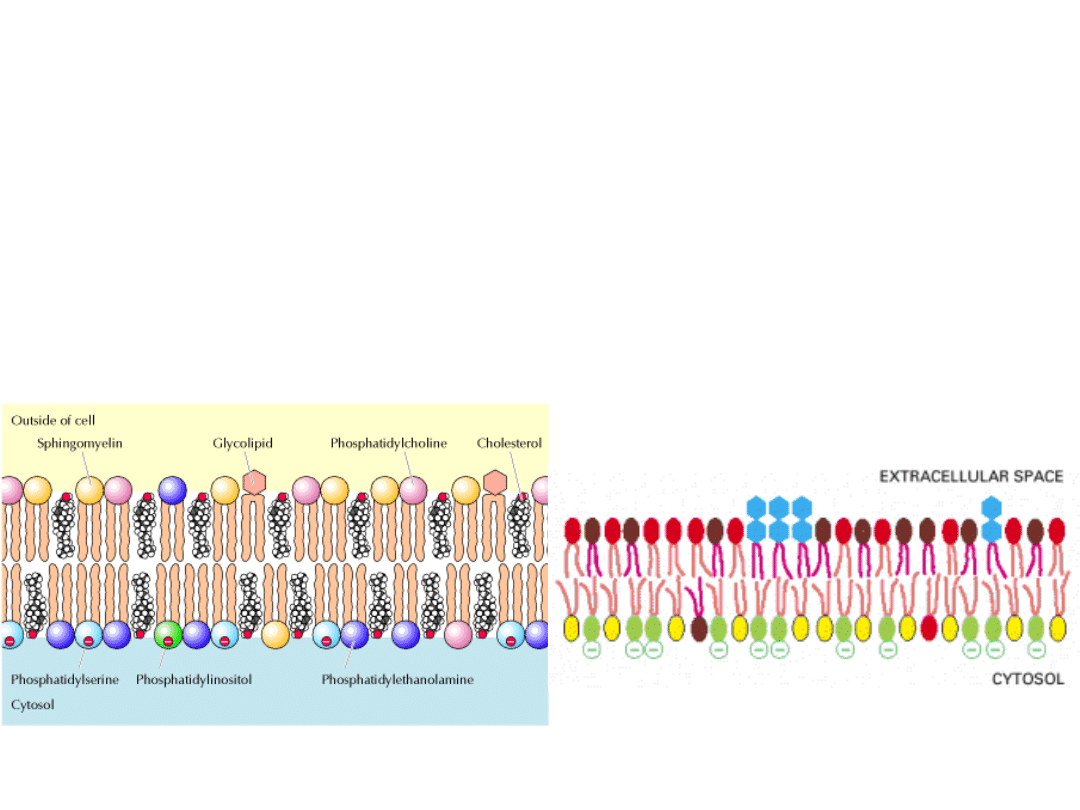
FOSFOLIPIDY
Są to pochodne GLICEROLU (fosfoglicerydy) lub SFINGOZYNY
(sfingomielina).
FOSFOGLICERYDY: szkielet cząsteczki – reszta glicerolu
2 łańcuchy kwasu tłuszczowego
połączone
wiązaniami
estrowymi z węglami C-1 i C-2 glicerolu
fosforylowany alkohol.
Najprostszym fosfoglicerydem jest FOSFATYD. W błonach występują
tylko
śladowe ilości fosfatydów, lecz pełnią one kluczową rolę jako
intermediaty w biosyntezie innych fosfoglicerydów.
Fosfatyd
diacyloglicerolo-3-fosforan
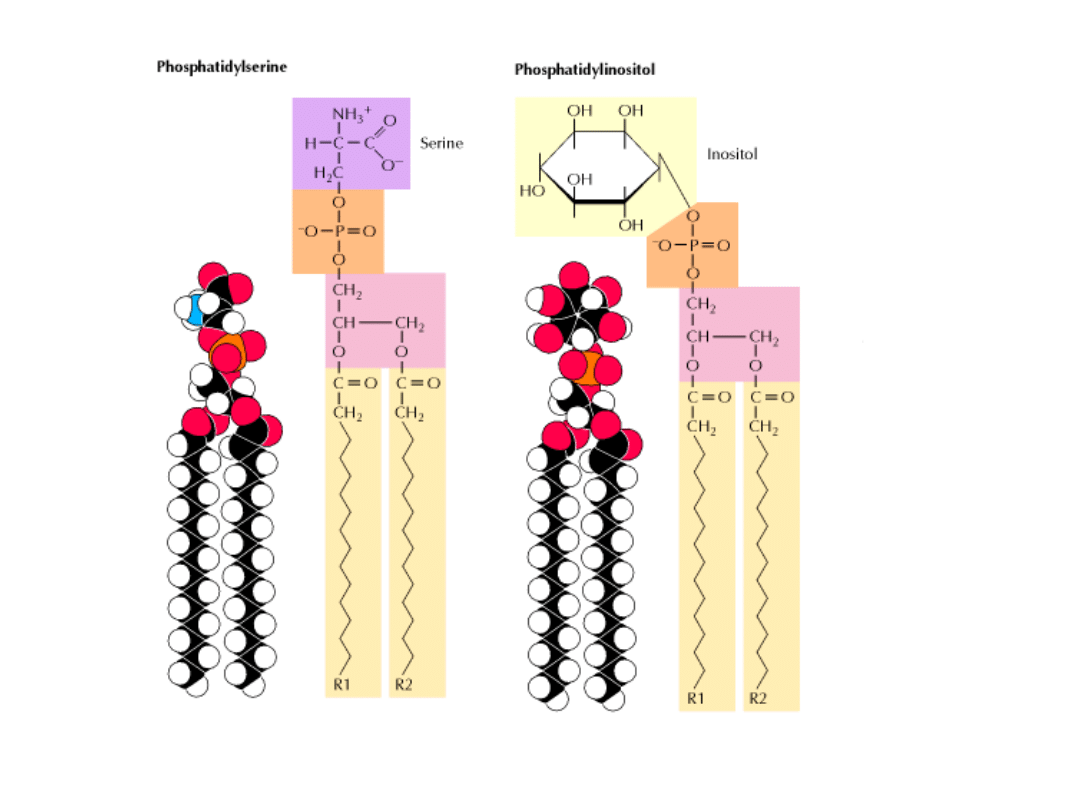
Większość fosfoglicerydów jest
pochodnymi fosfatydów:
grupa fosforanowa jest zestryfikowana
grupą hydroksylową jednego z alkoholi
seryny, etanoloaminy, choliny, glicerolu
i inozytolu.
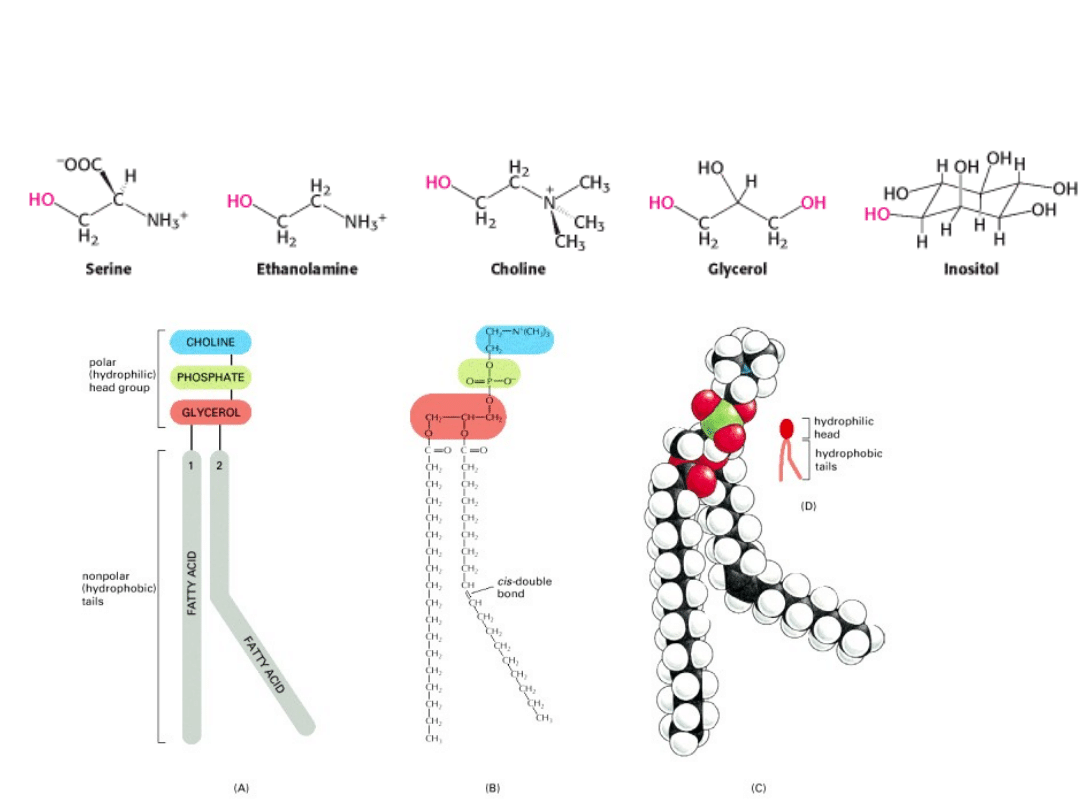
PODSTAWOWE
ALKOHOLE ESTRYFIKUJĄCE GR. FOSFORANOWĄ
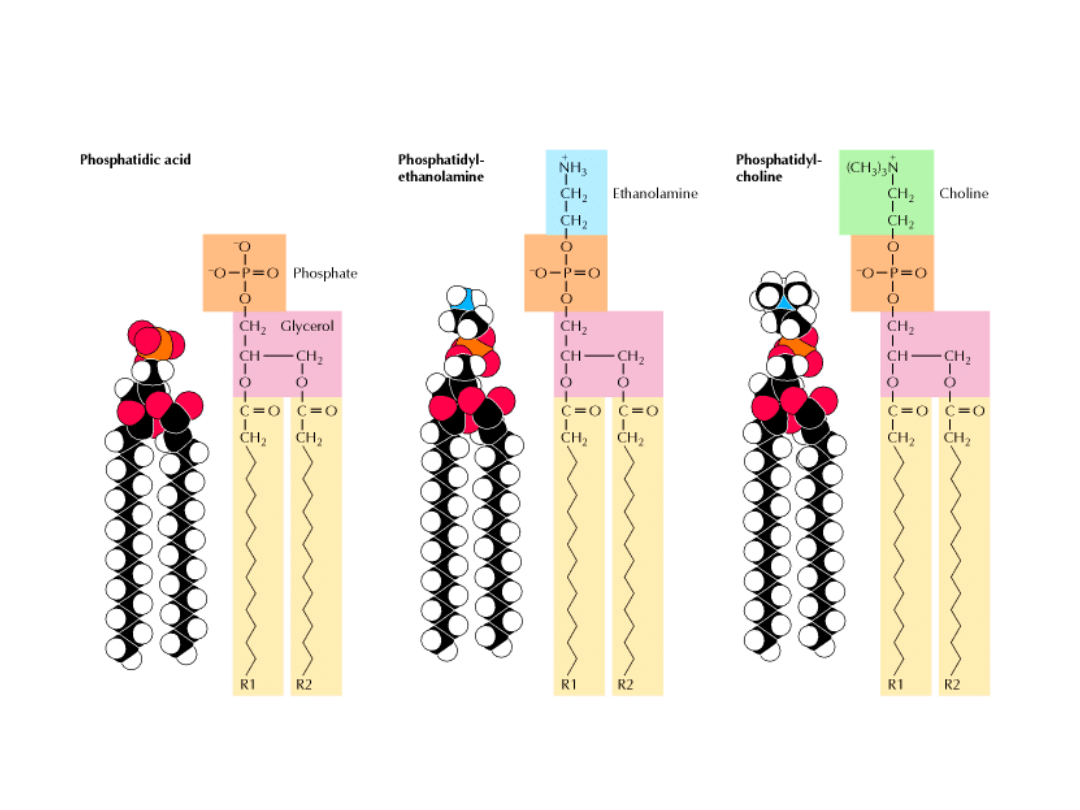
PODSTAWOWE FOSFOLIPIDY
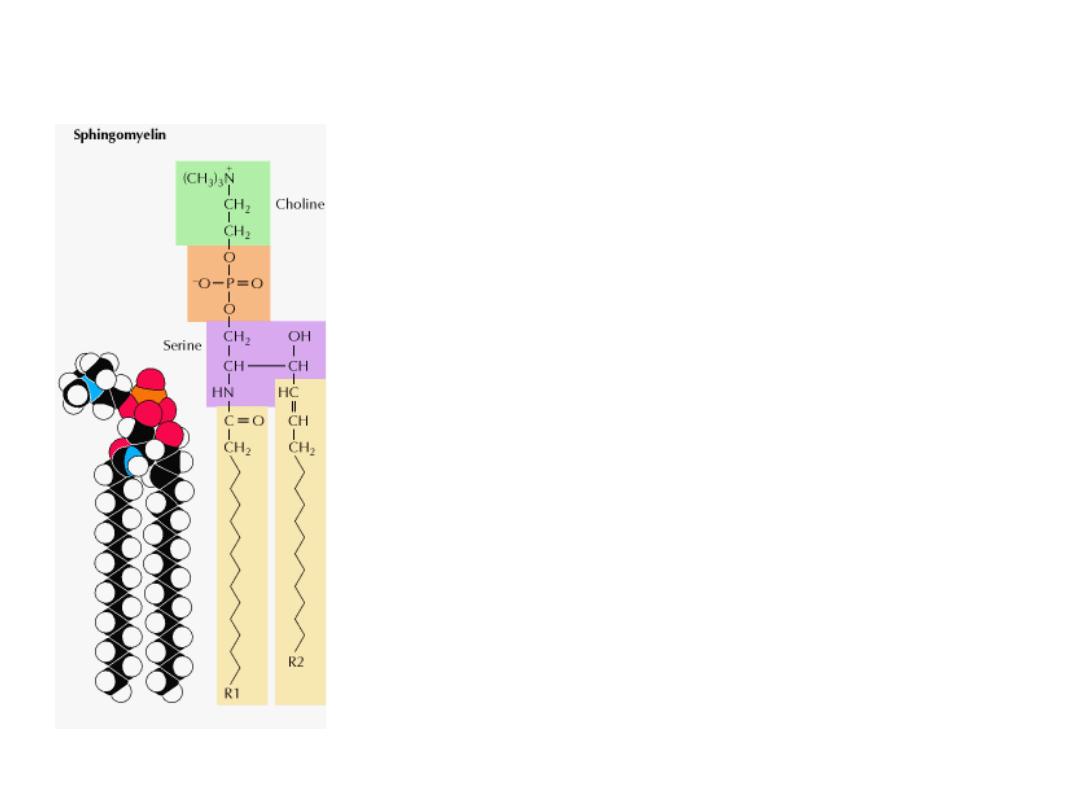
SFINGOMIELINA
Jest to jedyny fosfolipid błon, który nie jest pochodną
glicerolu.
Szkielet sfingomieliny stanowi SFINGOZYNA (na rysunku
BŁĄD: część fioletowa + R2 = sfingozyna).
Grupa amidowa szkieletu sfingozyny łączy się z kwasem
tłuszczowym poprzez WIĄZANIE AMIDOWE.
Pierwszorzędowa grupa hydroksylowa sfingozyny jest
zestryfikowana FOSFOCHOLINĄ.
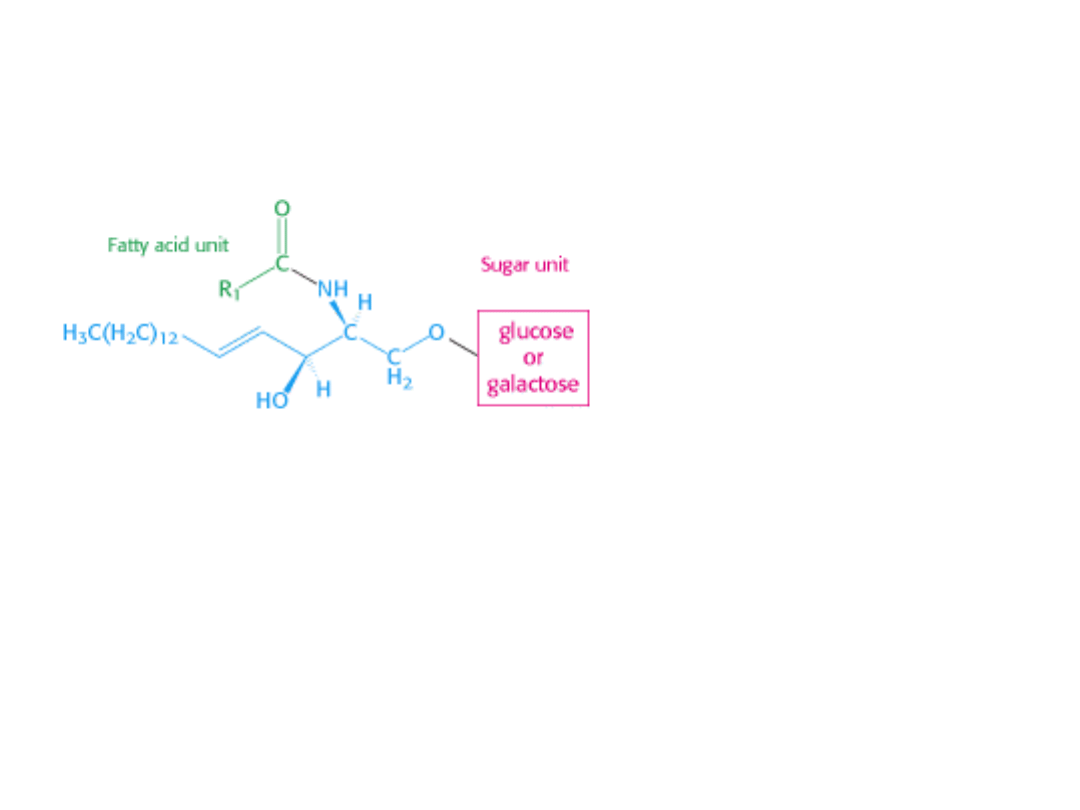
GLIKOLIPIDY
Są to lipidy zawierające cukier. Są pochodnymi SFINGOZYNY.
Cerebrozyd
Zamiast fosfocholiny w glikolipidach
z pierwszorzędową resztą
hydroksylową sfingozyny łączy się
jedna lub więcej cząsteczek cukru.
CEREBROZYD – najprostszy glikolipid,
zawiera 1 cząsteczkę cukru: glukozę
lub galaktozę.
GANGLIOZYDY – zawierają rozgałęziony łańcuch złożony aż z 7 reszt cukrowych.
Glikolipidy są asymetrycznie zorientowane w błonie – reszty cukrowcowe występują
JEDYNIE po zewnątrzkomórkowej stronie błony.
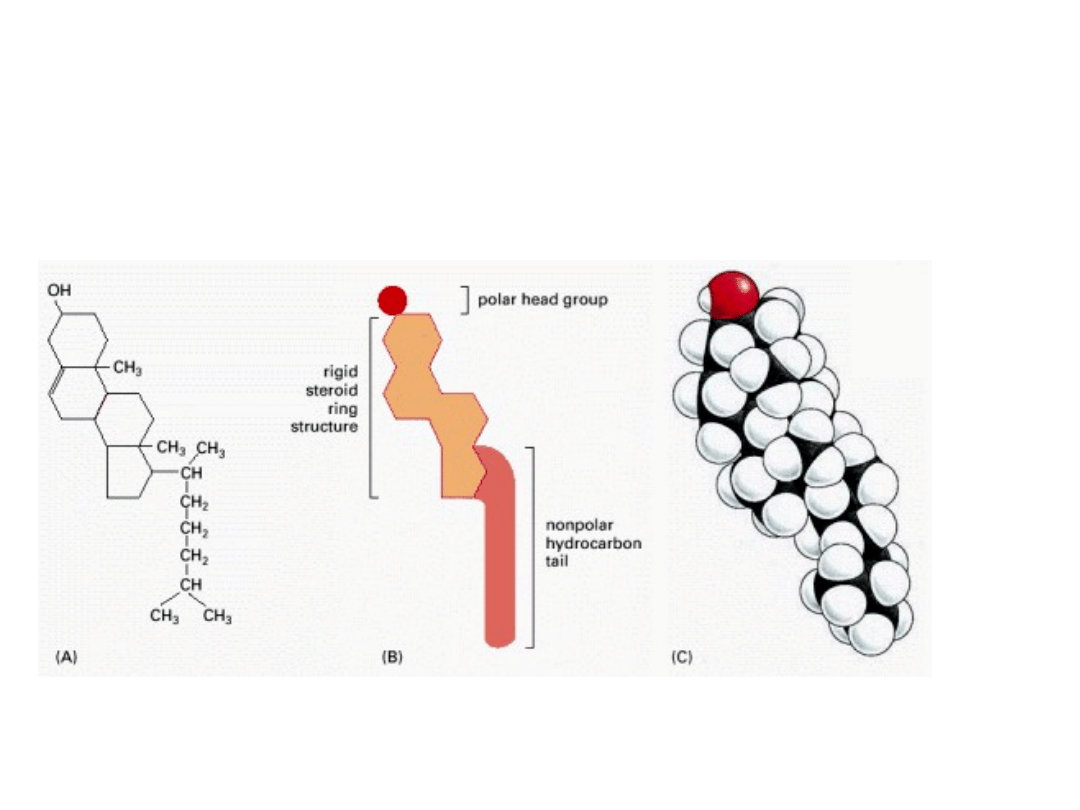
CHOLESTEROL
Występuje jedynie u organizmów eukariotycznych i i jest właściwie we wszystkich
błonach organizmów zwierzęcych, chociaż jego zawartość znacznie się waha w
zależności od typu błony. W niektórych komórkach nerwowych stanowi on ok.
25%, natomiast prawie nie występuje w błonach wewnątrzkomórkowych.
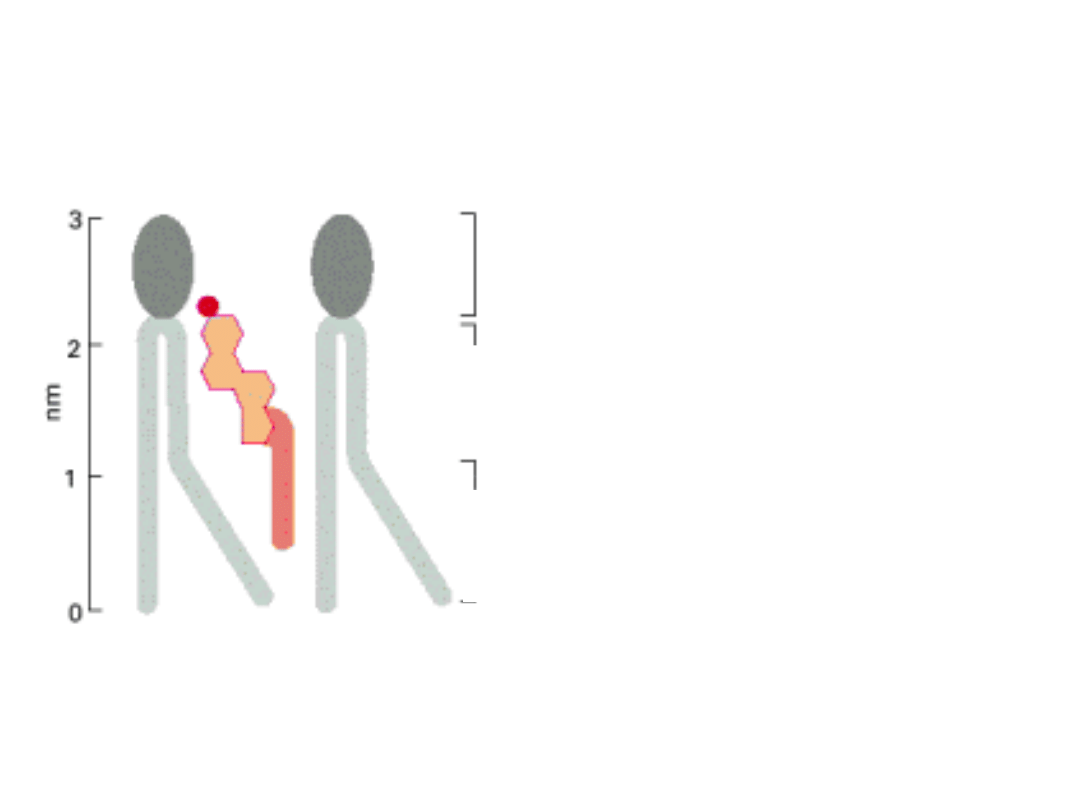
LIPIDY BŁONOWE SĄ CZĄSTECZKAMI
AMFOFILOWYMI
polarne
głowy
rejon hydrofobowy
usztywniony
pierścieniem
steroidowycm cholesterolu
bardziej płynny
rejon
hydrofobowy
W błonie cząsteczka cholesterolu jest ułożona równolegle do łańcuchów kwasów
tłuszczowych fosfolipidów. Grupa hydroksylowa oddziałuje z sąsiadującymi grupami
polarnych głów fosfolipidów.

Lipidy błon
Jednostki hydrofobowe
Jednostki
hydrofilowe
FOSFOGLICERYD
Y
SFIGOMIELINY
GLIKOLIPIDY
CHOLESTEROL
Łańcuchy kwasów tłuszczowych
Łańcuch kwasu tłuszczowego i
węglowodorowy łańcuch
sfingozyny
Łańcuch kwasu tłuszczowego i
węglowodorowy łańcuch
sfingozyny
Cała cząsteczka z wyjątkiem
grupy
OH
Ufosforylowany alkohol
Fosfocholina
Jedna lub więcej reszt
cukrowych
Grupa OH przy C-3
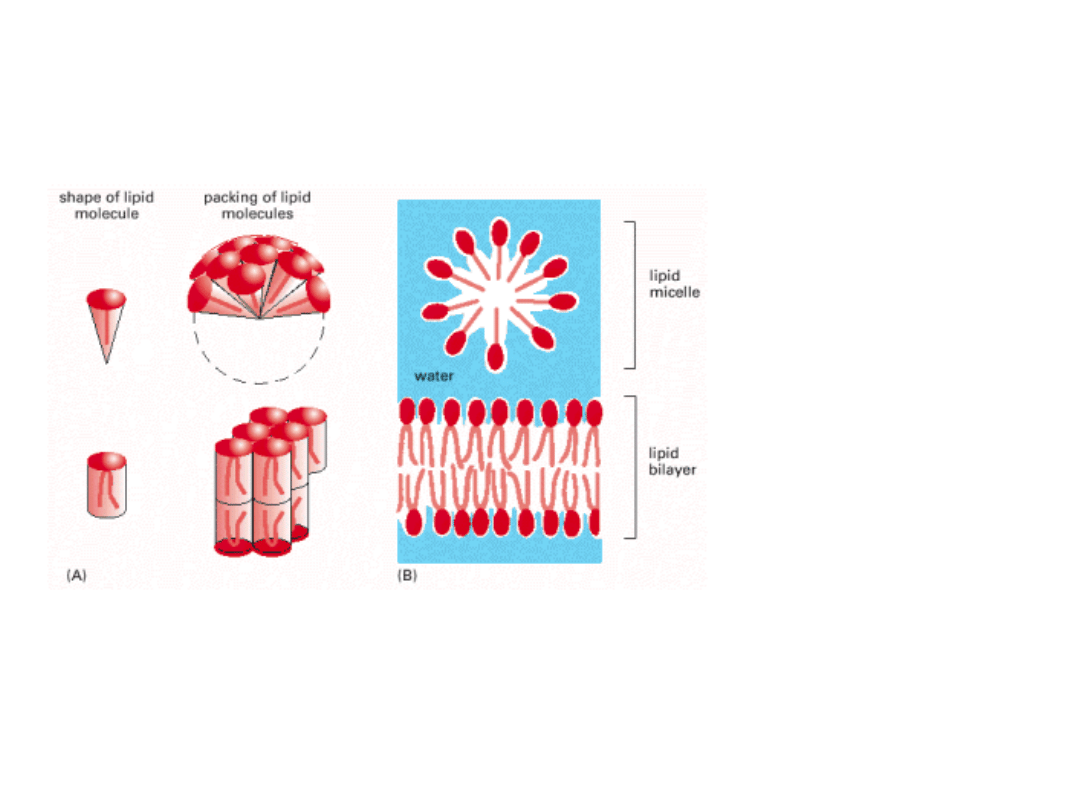
FOSFOLIPIDY I GLIKOLIPIDY W
ŚRODOWISKU WODNYM
Micele są rzadko tworzone przez
fosfolipidy i glikolipidy .
Gł. sole kwasów tłuszczowych.
Dwuwarstwa lipidowa – tworzo-
na gł. przez fosfolipidy
i glikoliopidy.
Tworzenie się dwuwarstwy lipidowej jest procesem samoistnym. Główną siłą orgfanizują-
cą stanowią ODDZIAŁYWANIA HYDROFOBOWE.
Ponadto: oddziaływania VAN DER WAALSA między łańcuchami węglowodorowymi
oddziaływania ELEKTROSTATYCZNE i WODOROWE między grupami polarnymi
głów fosfolipidów a cząsteczkami wody.
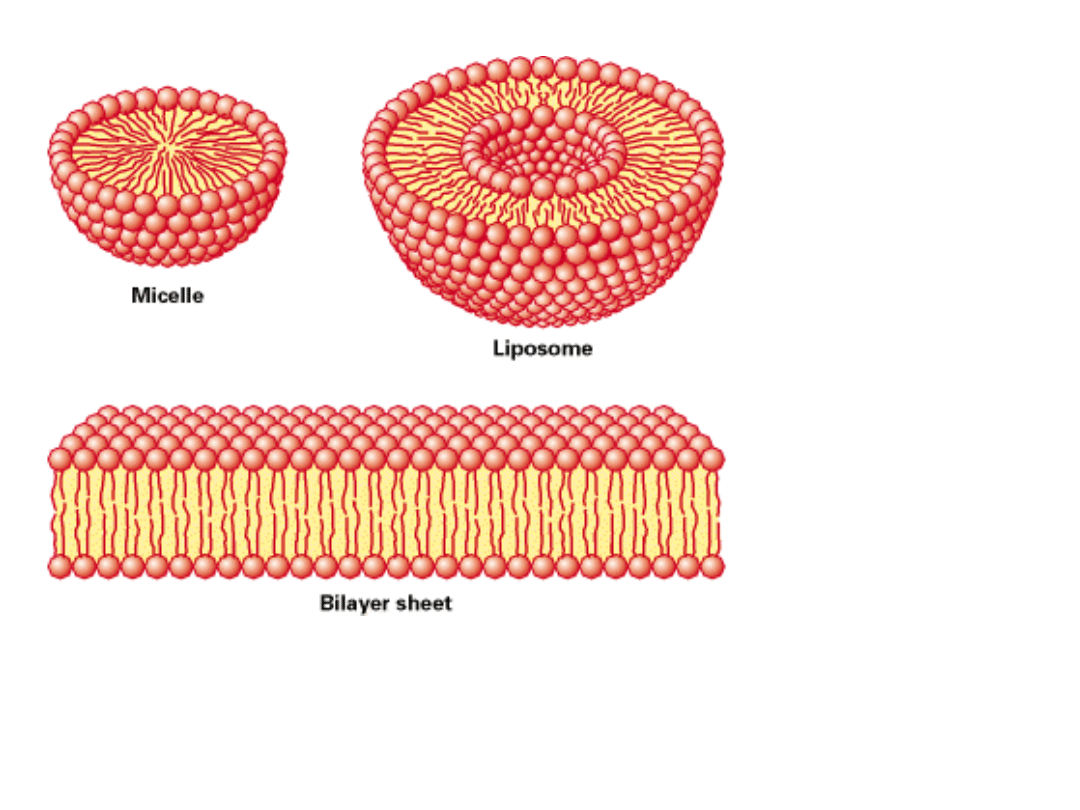
Dwuwarstwy lipidowy mają naturalną tendencję do tworzenia dużych powierzchni i do
ZASKLEPIANIA SIĘ, aby nie pozostawiać krawędzi z wystawionymi łańcuchami
węglowodorowymi –
tworzą zamknięte błoną przedziały.
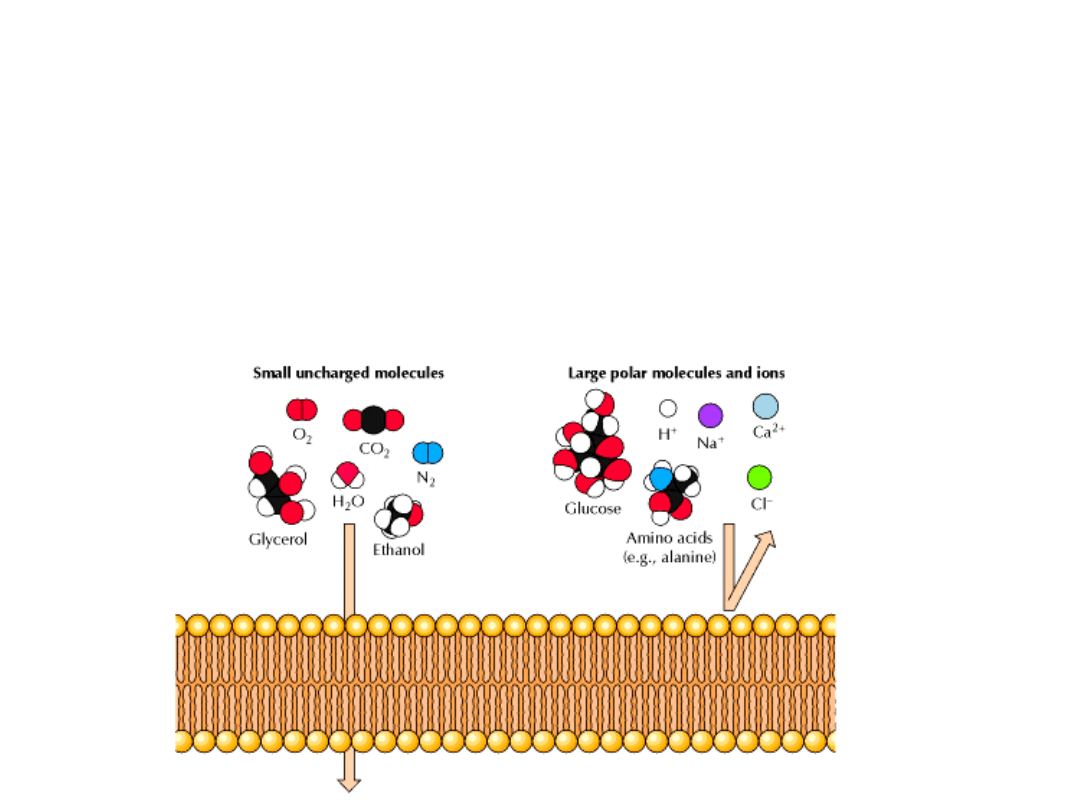
DWUWARSTWA LIPIDOWA JAKO
BARIERA
Dwuwarstwa jest nieprzepuszczalna dla jonów i większości cząsteczek
polarnych (wyjątki: woda, etanol).
PRZYCZYNY:
1. cząsteczka przechodząc przez dwuwarstwę musi utracić otoczkę
hydratacyjną,
2. następnie musi ulec rozpuszczeniu w węglowodorowym rdzeniu,
3. na koniec dyfunduje na drugą stronę błony.
Pozbywanie się otoczki hydratacyjnej jest termodynamicznie niekorzystne.
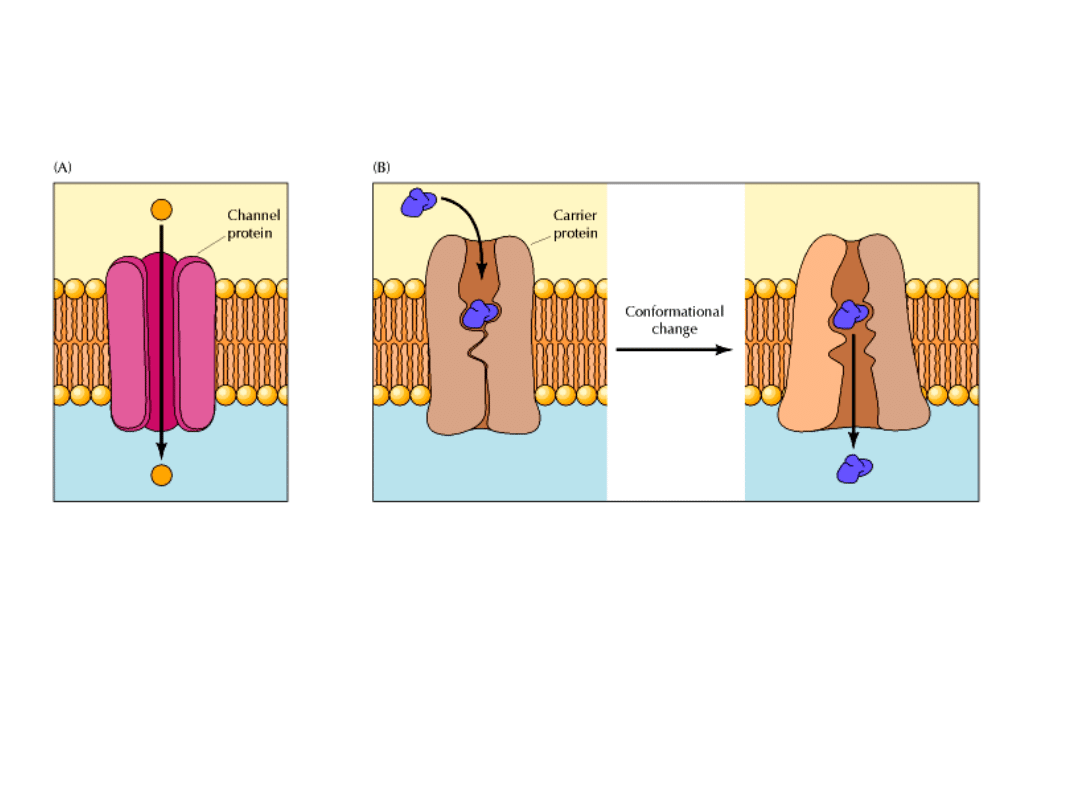
KANAŁY I POMPY BŁONOWE
Przenikanie wielu cząsteczek polarnych możliwe jest dzięki białkom tworzącym kanały
w błonie lub funkcjonującym jako pompy.
KANAŁY –
umożliwiają szybki przepływ jonów przez błony w kierunku termodynamicznie
korzystnym. Jest to przykład TRANSPORTU BIERNEGO lub DYFUZJI UŁATWIONEJ.
POMPY – wykorzystując źródła energii swobodnej, takie jak ATP lub światło prowadzą
transport jonów lub cząsteczek w kierunku termodynamicznie niekorzystnym.
Jest to przykład TRANSPORTU AKTYWNEGO.
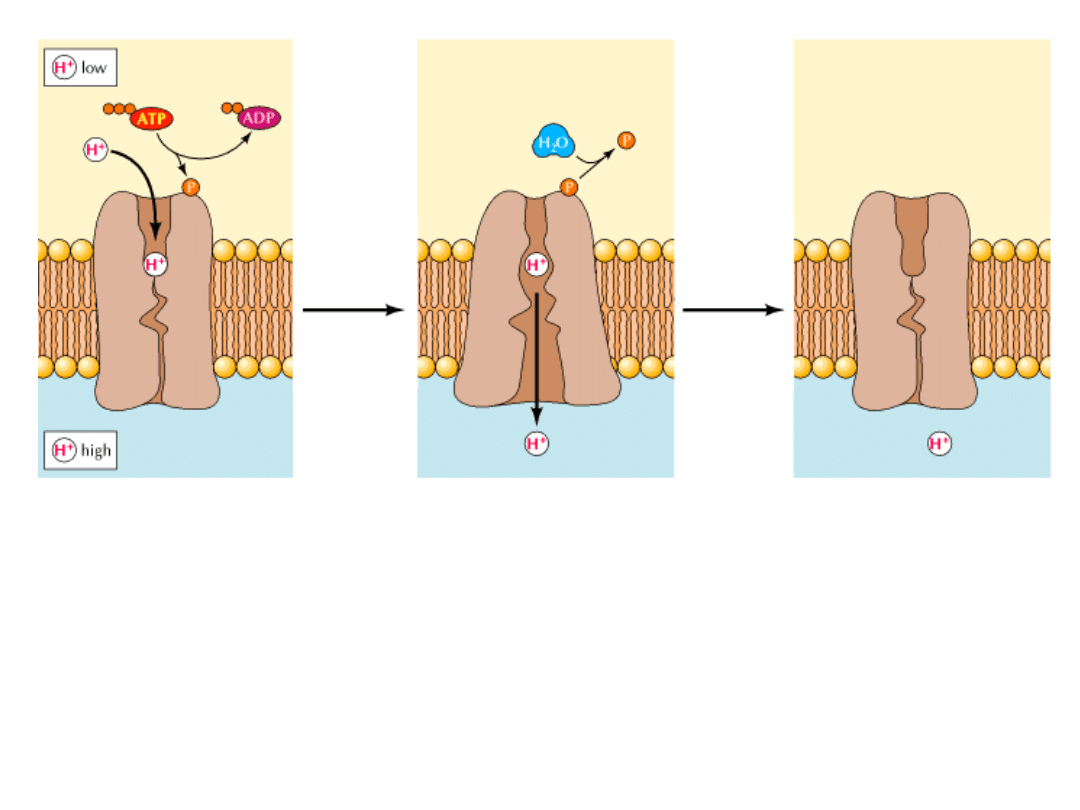
Przykładem mogą być
pompy zależne od ATP
.
Białka te podlegają zmianom konformacyjnym podczas wiązania i hydrolizy ATP, co
powoduje, że związany jon może być przetransportowany przez błonę.
Fosforylacja i defosforylacja pompy są sprzężone ze zmianami orientacji i powinowactwa
miejsc wiązania jonów.
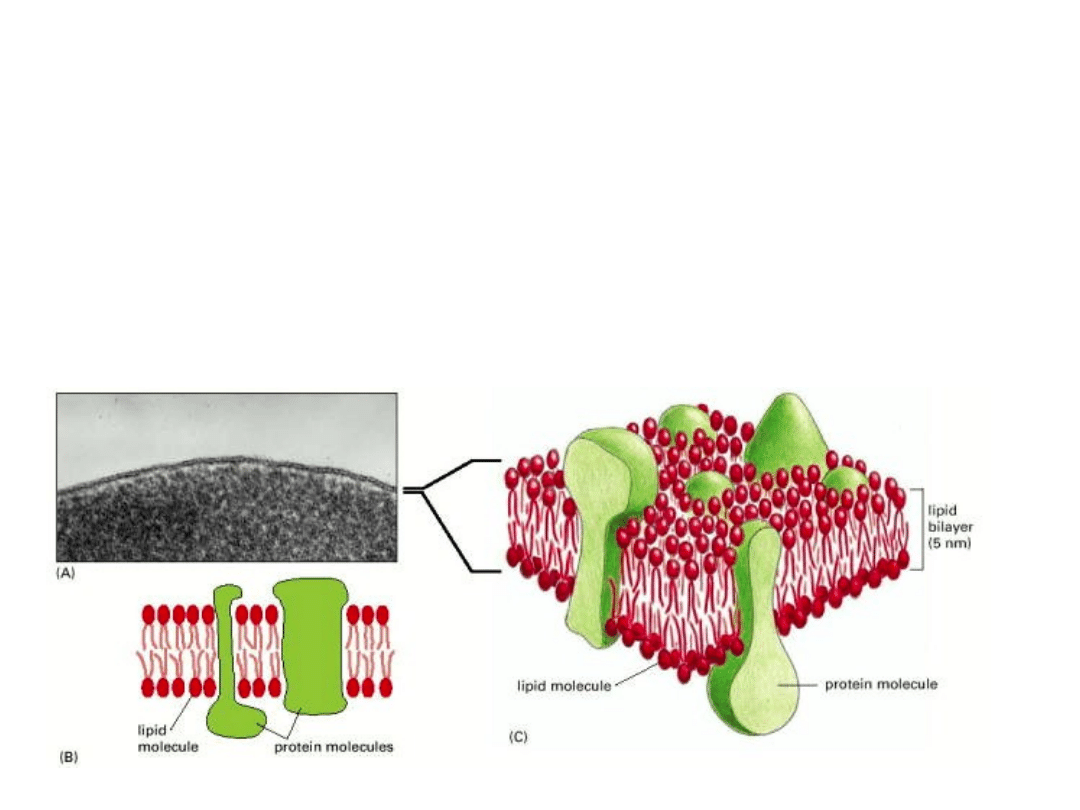
BIAŁKA BŁONOWE
Błony biologiczne różnią się pod względem zawartości białek.
MIELINA- błona otaczająca niektóre włókna nerwowe, działa jako izolator.
Białka stanowią jedynie 18%.
BŁONY PLAZMATYCZNE – znacznie bardziej aktywne – ok. 50% białek
BŁONY WEWNĘTRZNE MITOCHONDRIÓW I CHLOROPLASTÓW – największa
zawartość białek – ok. 75%. (bardzo duża aktywność metaboliczna).
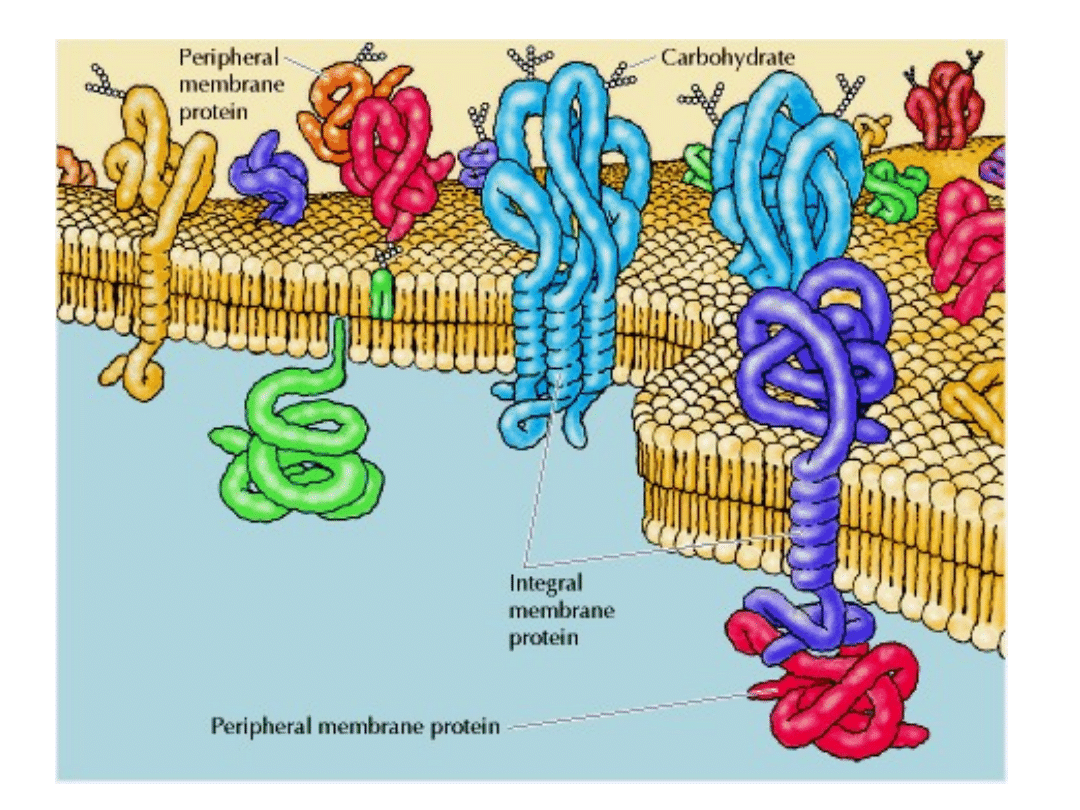
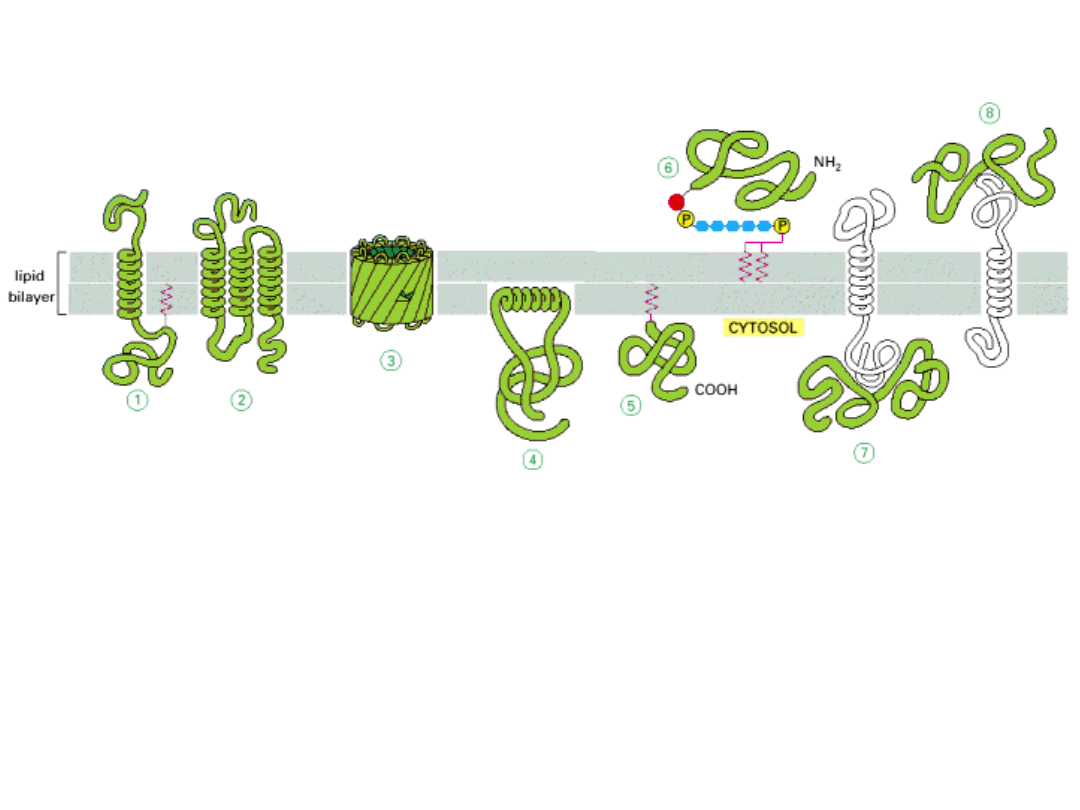
BIAŁKA BŁONOWE MOGĄ BYĆ W RÓŻNY SPOSÓB ZWIĄZANE Z BŁONĄ
INTEGRALNE białka błonowe – transbłonowe (1-3)
lub zakotwiczone w błonie hydrofobowymi rejionami (4) lub zakotwiczone lipidami
związanymi kowalencyjnie z białkami (5,6)
PERYFERYJNE białka błonowe (7,8) – związane są z błoną poprzez oddziaływanie z
innymi białkami błonowymi.
OGÓLNIE – w błonie zanurzone są rejony hydrofobowe białek, natomiast na zewnątrz błony
są eksponowane rejony hydrofilowe.
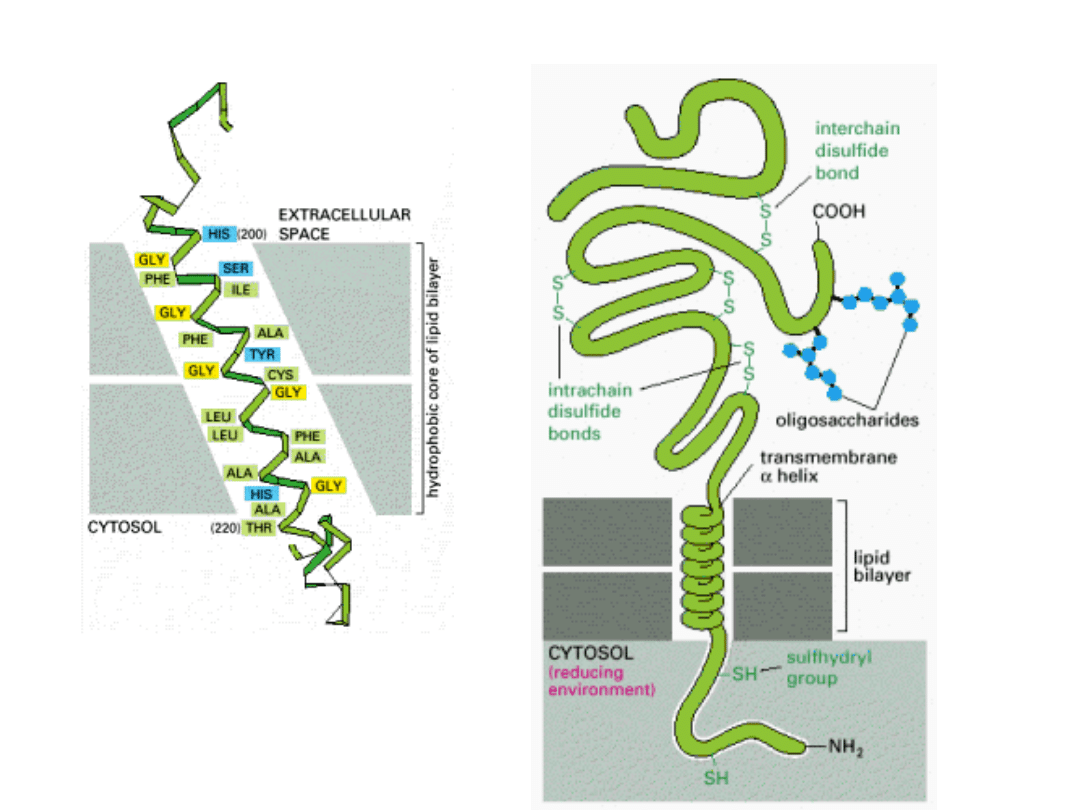
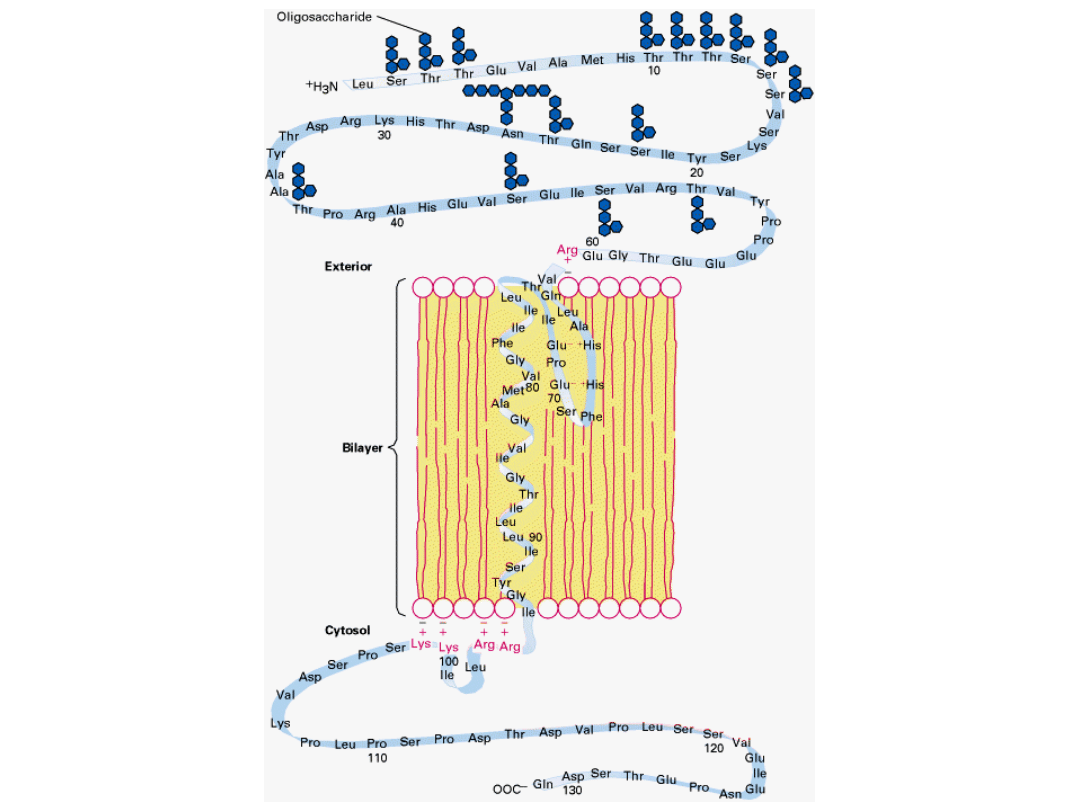
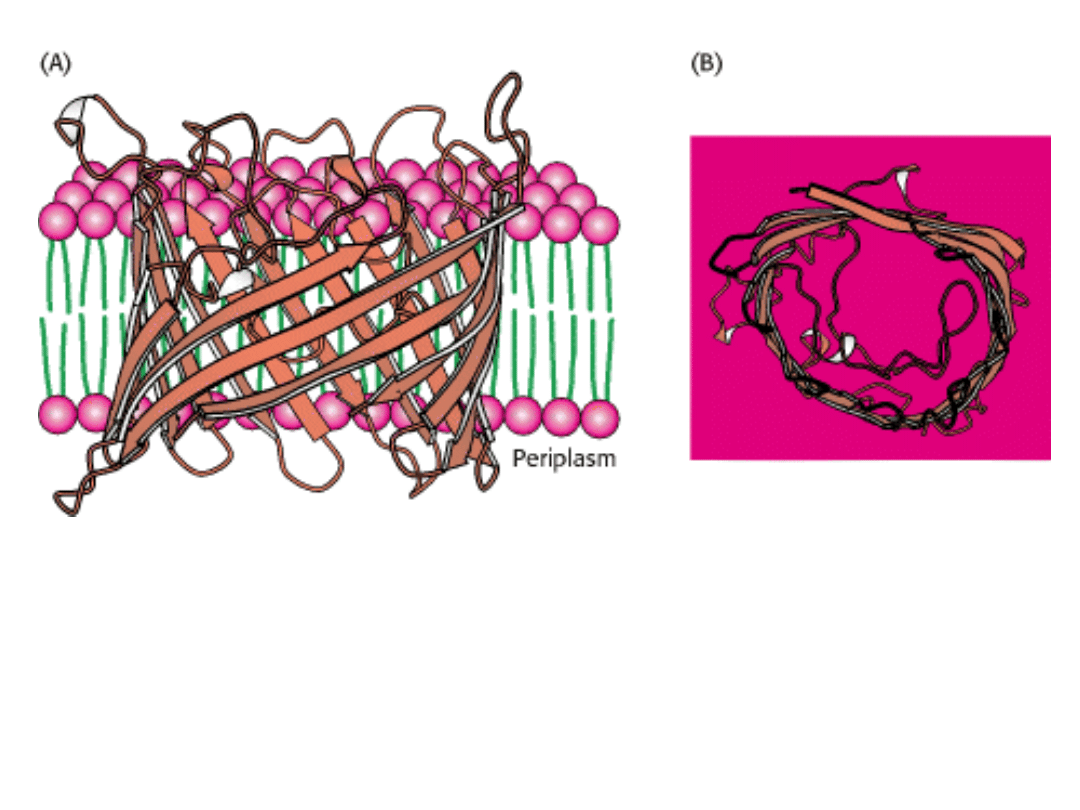
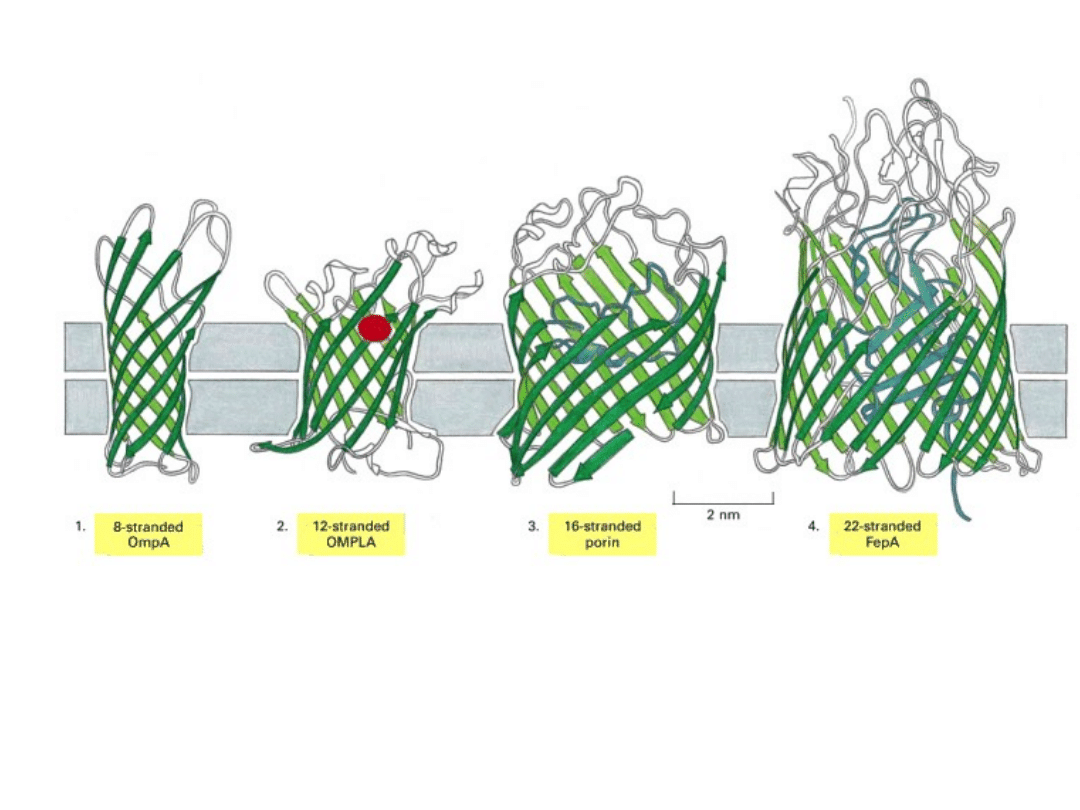
Białka kanałowe mogą być tworzone przez nici β.
Zewnętrzna powierzchnia poryny jest stosunkowo niepolarna i oddziałuje z
węglowodorowym rdzeniem błony.
Wnętrze kanału jest wyraźnie hydrofilowe i wypełnione cząsteczkami wody.
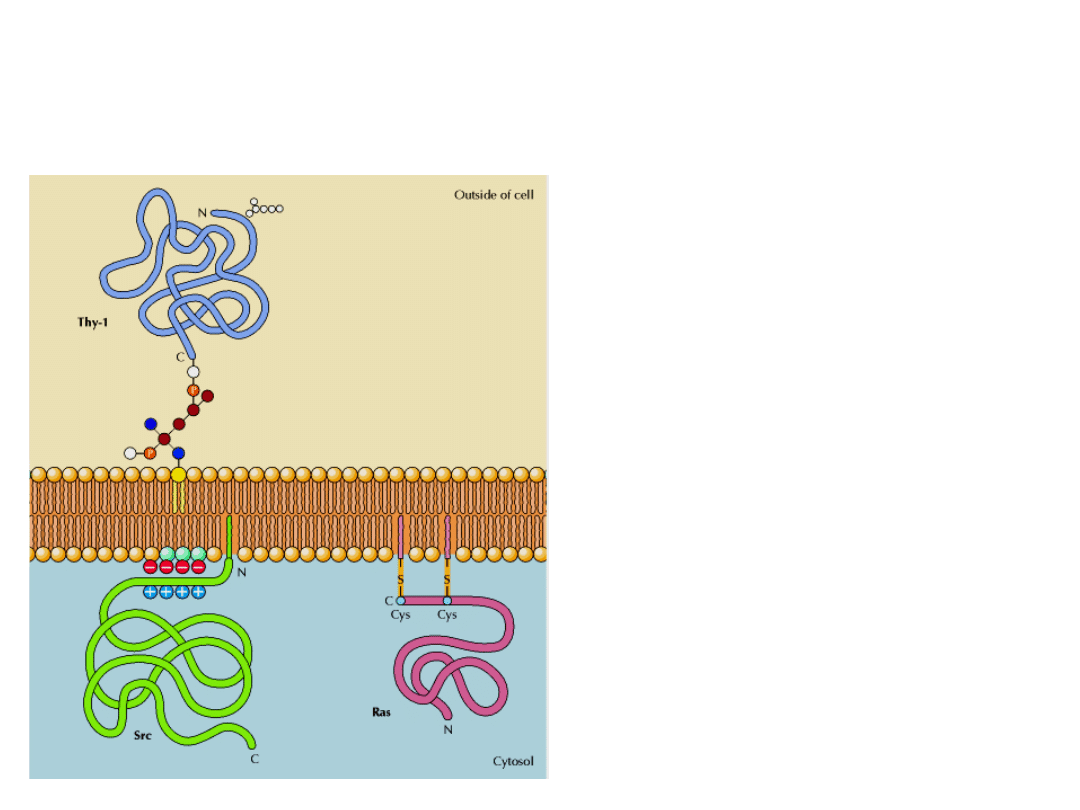
NIEKTÓRE BIAŁKA WIĄŻĄ SIĘ Z BŁONĄ
POPRZEZ KOWALENCYJNIE PRZYŁĄCZONE DO
NICH GRUPY HYDROFOBOWE
1. Ugrupowanie glikolipidowe
( kotwica GPI) przymocowane do
końca karboksylowego łańcucha
polipeptydowego.
2. Grupa palmitoilowa, połączona poprzez
wiązanie tioestrowe ze specyficzną resztą
cysteiny lub grupa farnezylowa przyłączona
do reszty cysteiny na C-końcu białka.
PŁYNNOŚĆ BŁON
BŁONY BIOLOGICZNE NIE SĄ
STRUKTURAMI
SZTYWNYMI.
Białka i lipidy znajdują się w ciągłym
ruchu.
Ruch białek w płaszczyźnie poziomej –
do
kilku μM/min.
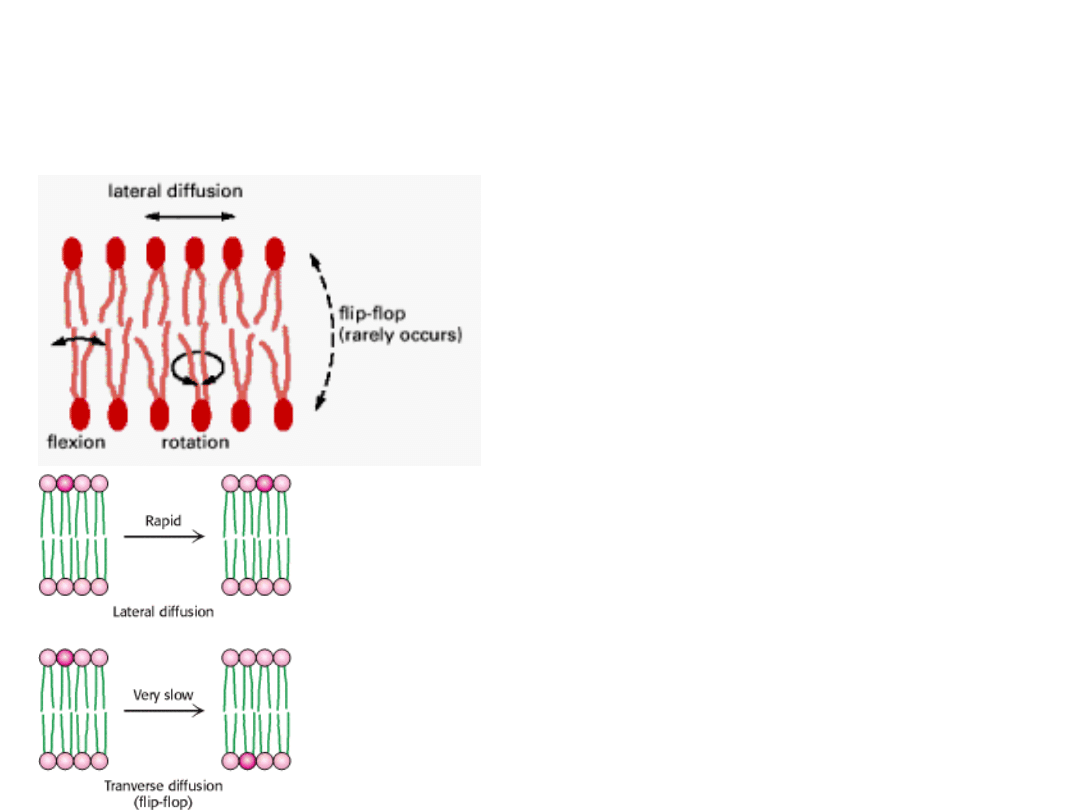
RUCHLIWOŚĆ LIPIDÓW
BŁONOWYCH
Lipidy są generalnie bardziej ruchliwe od białek.
Typy dyfuzji:
1. DYFUZJA BOCZNA (lateralna) – szybka
2. DYFUZJA POPRZECZNA (flip-flop) –
bardzo powolna
Czasami konieczne jest przemieszczanie
cząsteczki fosfolipidu w poprzek błony.
Dzieje się tak np. podczas wzrostu komórki.
Nowo syntetyzowane fosfolipidy gromadzą się
po wewnętrznej stronie dwuwarstwy – ta strona
staje się powiększona.
Funkcję przenoszenia lipidów w poprzek błony
pełnią enzymy –
FLIPAZY
, które selektywnie
przenoszą specyficzne cząsteczki fosfolipidów.
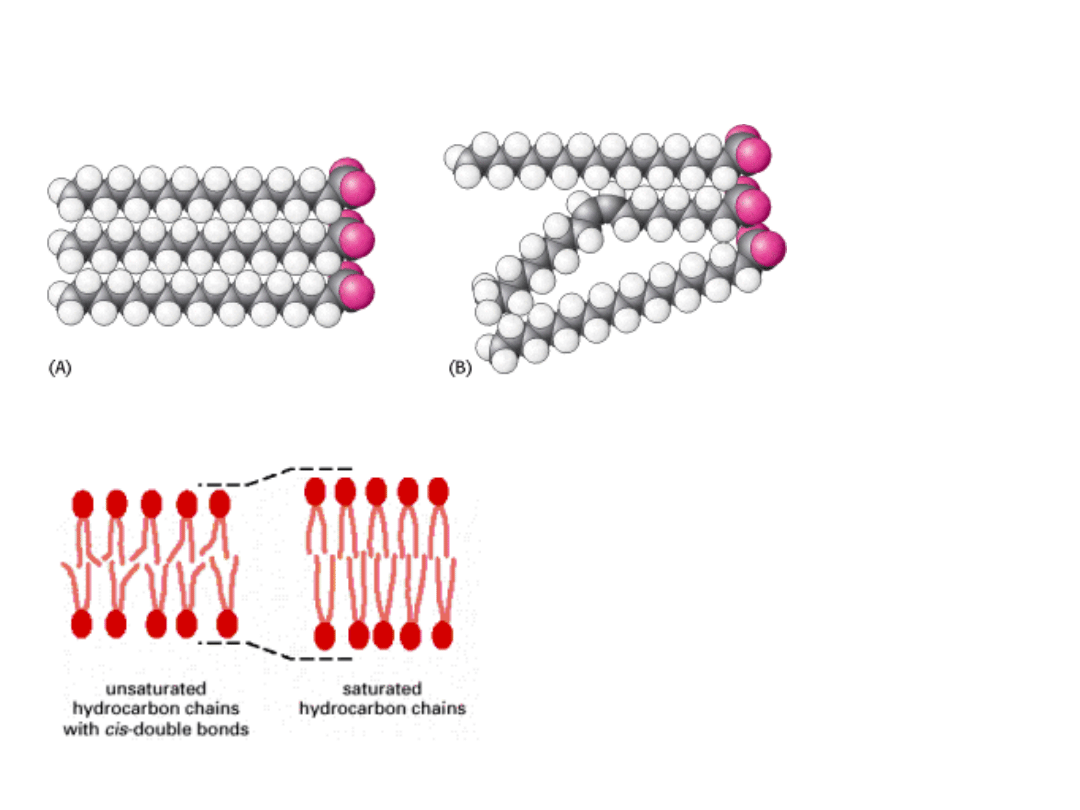
PŁYNNOŚĆ BŁON
Na płynność błon wpływają:
1. Długość łańcucha węglowodorowego – im
dłuższe, tym mniejsza płynność
2. Obecność wiązań podwójnych – obniżają
temperaturę topnienia
3. Obecność cholesterolu – wciskając się
między łańcuchy węglowodorowe zapobiega
ich krystalizacji.
Z drugiej strony sztywne pierścienie
steroidowe ograniczają ruchy łańcuchów
węglowodorowych na większą skalę.
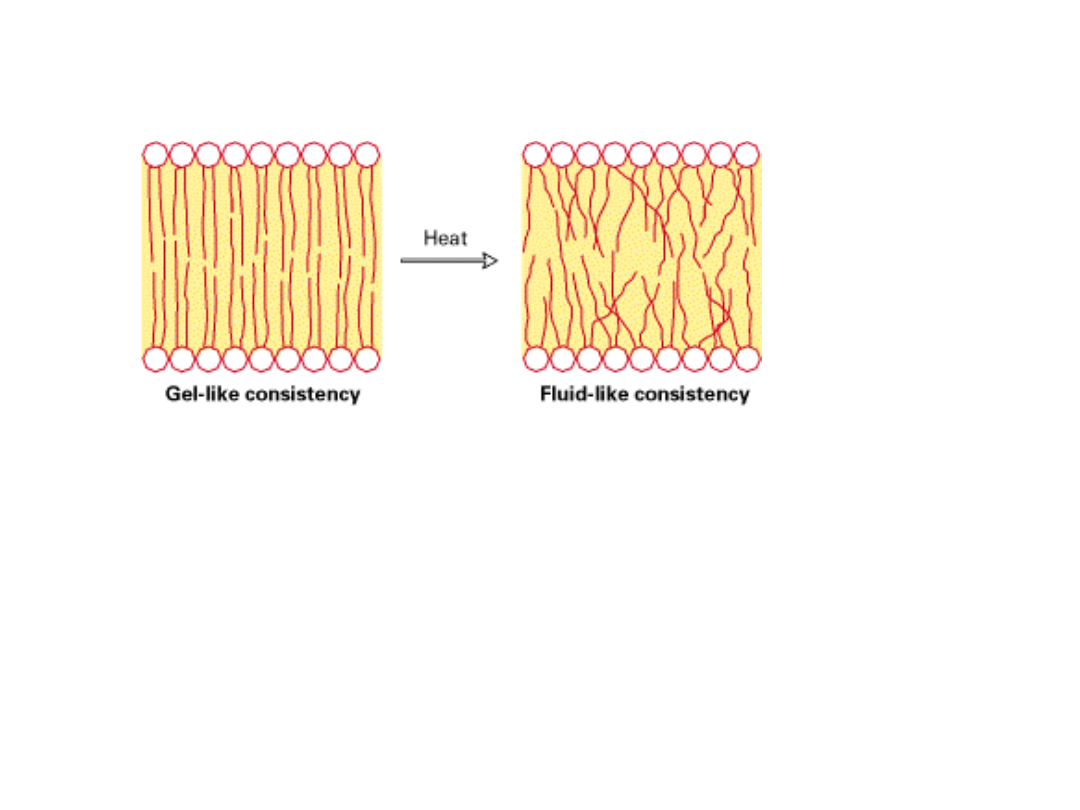
ZMIANY ADAPTACYJNE BŁON
Pod wpływem zmian temperatury zmienia się konsystencja błon.
Adaptacja organizmów:
1. U ryb zamieszkujących wody odznaczające się dużymi wahaniami sezonowymi
temperatur (roczne wahania temp. 30 10
o
C ) dochodzi do zmian stopnia
nasycenia fosfolipidów dzięki działaniu enzymu DESATURAZY. Spadek temperatury
indukuje enzym wzrost ilości łańcuchów z wiązaniami nienasyconymi.
2. U bakterii – zmiany poziomu syntezy łańcuchów węglowodorowych z wiązaniami
nienasyconymi w zależności od temperatury otoczenia.
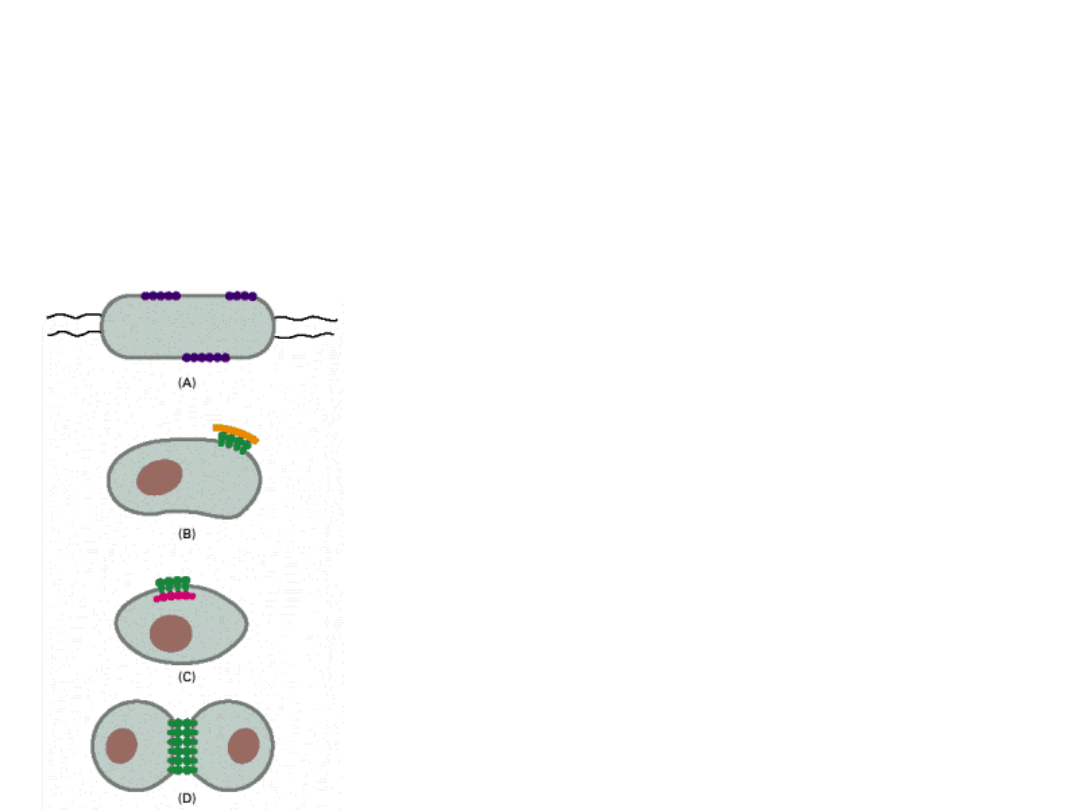
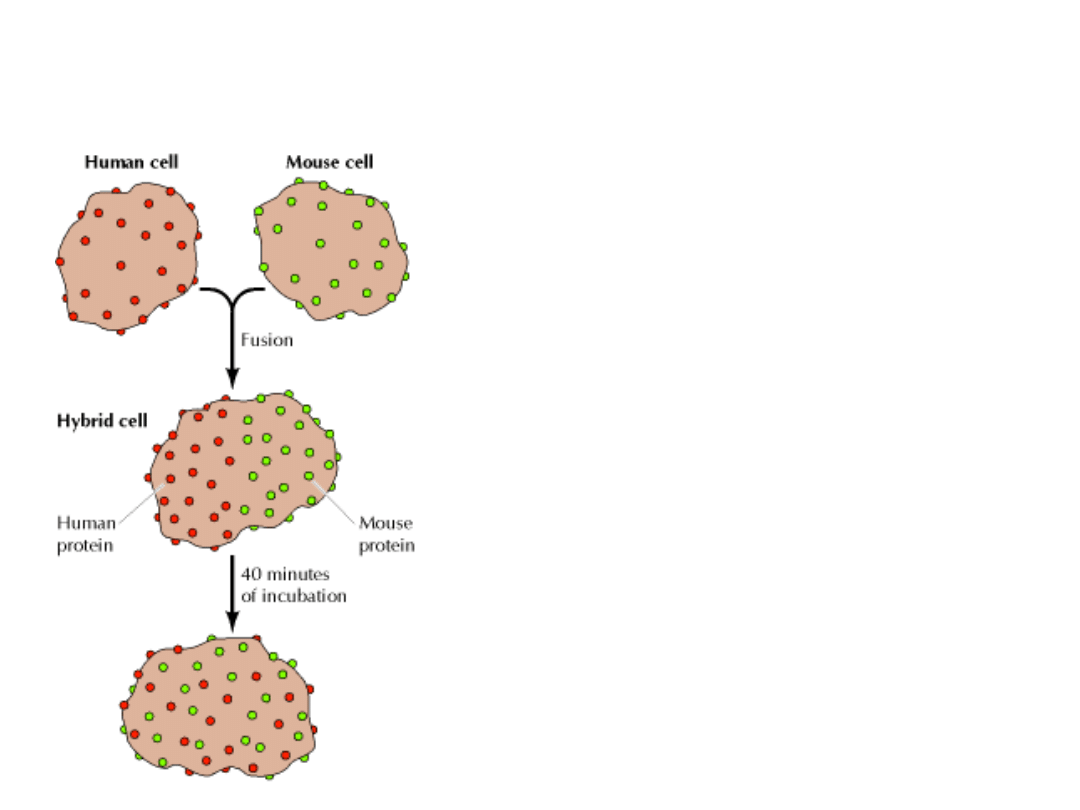
RUCHLIWOŚĆ BIAŁEK
Białka są na ogół mniej ruchliwe od lipidów i różnią się znacznie między sobą
ruchliwością.
Niektóre są bardzo ruchliwe (prawie jak lipidy), np. RODOPSYNA; inne są prawie
nieruchome, np. FIBRONEKTYNA, zakotwiczona do włókien aktyny.
A. Białka mogą tworzyć wielkie agregaty w błonie (np.
bakteriorodopsyna w błonie purpurowej bakterii
Halobacterium).
B. Białka mogą być wczepione w cząsteczki substancji
zewnątrzkomórkowej.
C. Białka mogą być włączone w korę komórki (sieć
włóknistych białek stanowiących przybłonową część
cytoszkieletu).
D. Białka mogą oddziaływać z innymi białkami na
powierzchni sąsiadujących komórek
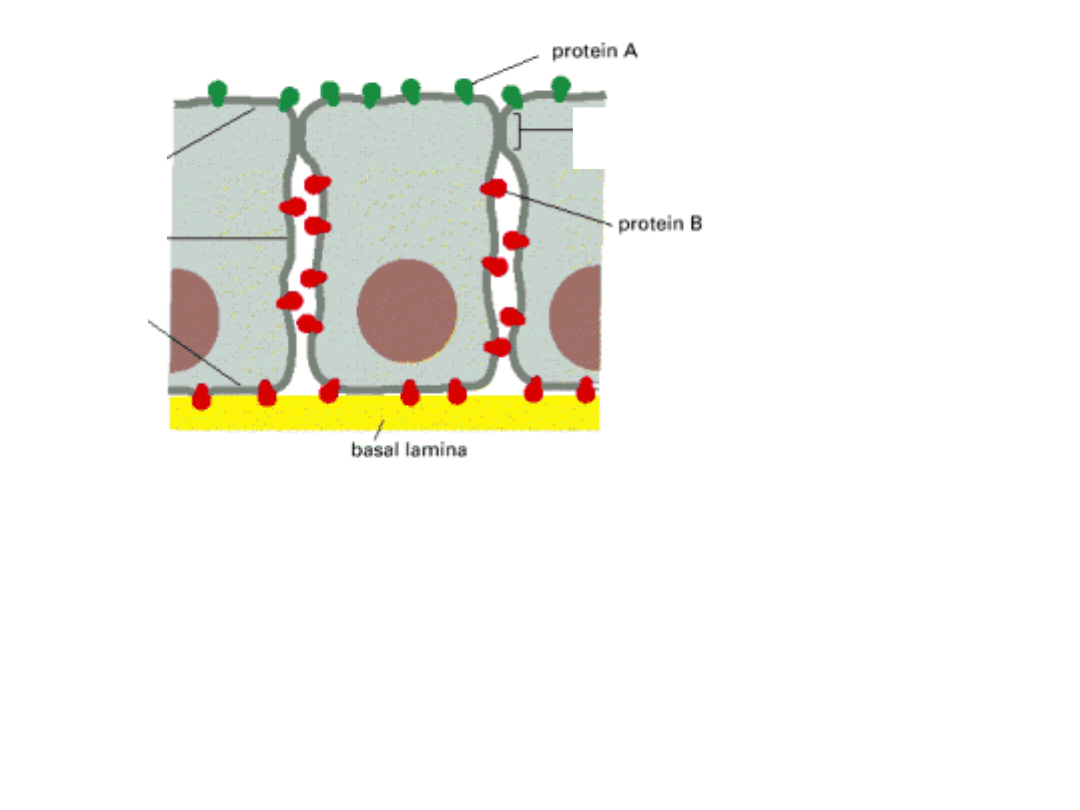
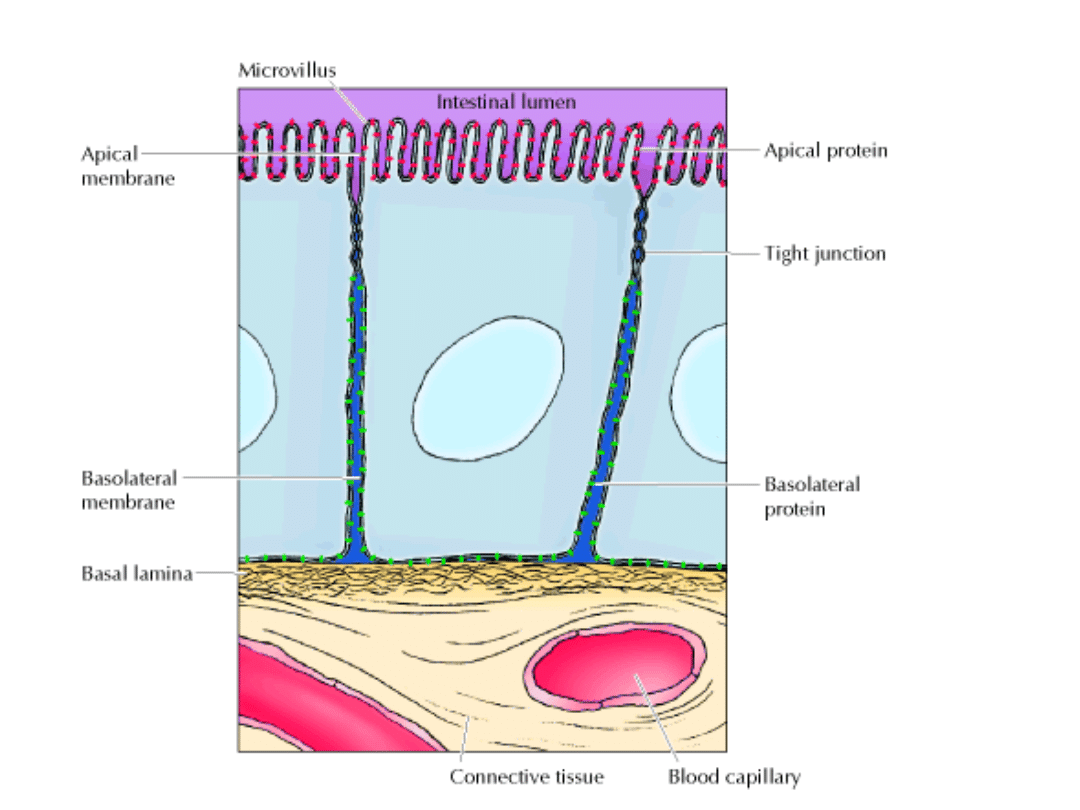
SCHEMAT NABŁONKA JELITA
Białko A (w błonie szczytowej) i białko B( w błonach bocznych i podstawnej)
mogą dyfundować bocznie w obrębie swych domen błony, ale ich przejście do
innych domen jest uniemożliwione przez specjalne złączenia komórkowe
o nazwie „ZŁĄCZENIA ZAMYKAJĄCE”.
podstawna
błona
komórkowa
szczytowa
błona
komórkowa
boczna błona
komórkowa
ZŁĄCZENIA ZAMYKAJĄCE – pas utworzony
przez białka łączące -ŚCISŁE ZESPOLENIE -
-
białko transportujące substancje
odżywcze z jelita
-
wyprowadza
rozpuszczone
substancje z komórki
nabłonkowej do krwiobiegu.
ASYMETRIA BŁON
Występuje asymetria błon pod względem strukturalnym i funkcjonalnym.
1. Każda powierzchnia zawiera inne składniki i wykazuje odmienną
aktywność biologiczną
2. Rozmieszczenie lipidów jest również asymetryczne, ale w mniejszym
stopniu
niż białka (wyj. glikolipidy).
GLIKOLIPIDY i GLIKOPROTEINY umieszczone są zawsze po
zewnątrzkomórkowej stronie błony.
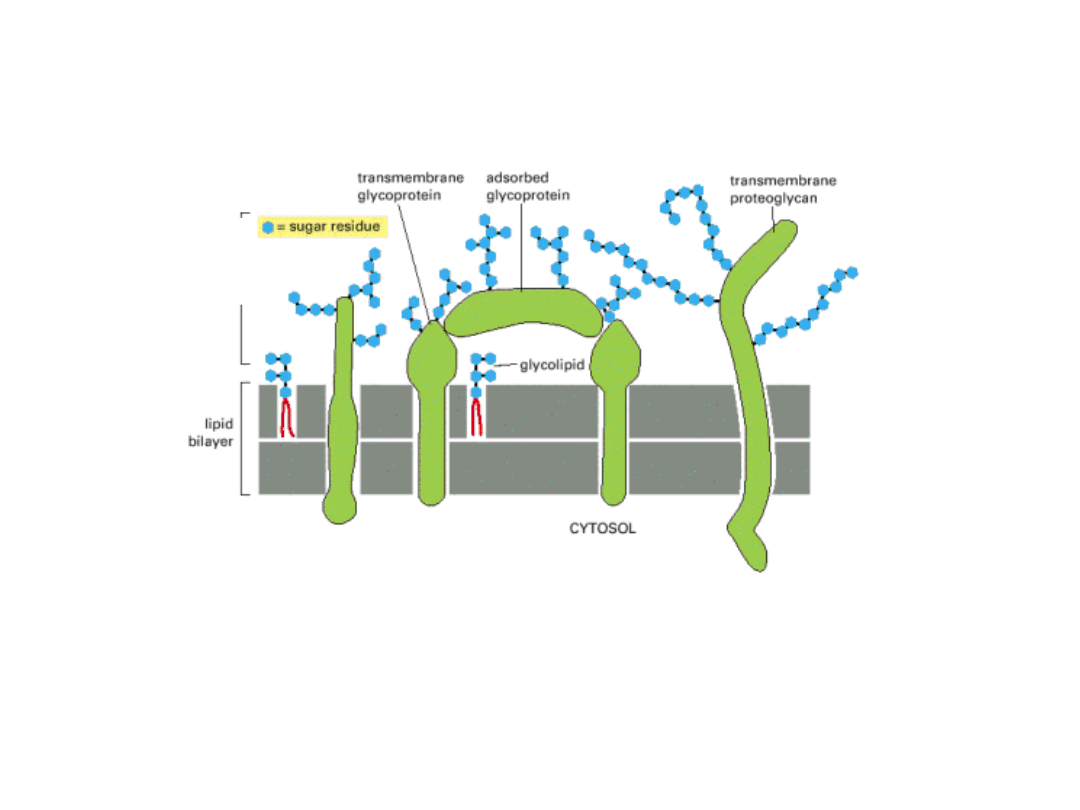
POWIERZCHNIA KOMÓRKI POKRYTA
JEST CUKROWCAMI
Wiele lipidów i białek leżących na zewnątrzkomórkowej stronie błony ma przyłączone
kowalencyjnie CUKRY.
Większość białek przyłącza krótkie łańcuchy cukrowe – oligosacharydy –
GLIKOPROTEINY
Jeśli białko zawiera 1 lub więcej długich łańcuchów polisacharydowych –
PROTEOGLIKAN
.
GLIKOKALIKS
– płaszcz cukrowcowy na powierzchni komórki, tworzony przez wszystkie
cukrowce wchodzące w skład glikoprotein, proteoglikanów i glikolipidów.
Płaszcz
komórkowy-
GLIKOKALIKS

ROLA GLIKOKALIKSU
1. ELEMENT OCHRONNY – chroni przed uszkodzeniami mechanicznymi i
chemicznymi.
Oligosacharydy i polisacharydy pochłaniają wodę wzrost śliskości
powierzchni
komórki. Dzięki temu komórki są ruchliwe, np.. białe krwinki mogą się
przeciskać
przez wąskie przestrzenie.
Poza tym krwinki się nie zlepiają i nie przyklejają do ścianek naczyń
krwionośnych.
2. ROLA WE WZAJEMNYM ROZPOZNAWANIU SIĘ KOMÓREK I ADHEZJI
Pewne białka –
LEKTYNY
- są wyspecjalizowane w rozpoznawaniu
poszczególnych
bocznych łańcuchów cukrowcowych i ich wiązaniu.
Glikokaliks może służyć jako rodzaj wyróżniającego okrycia,
rozpoznawanego
przez inne komórki.
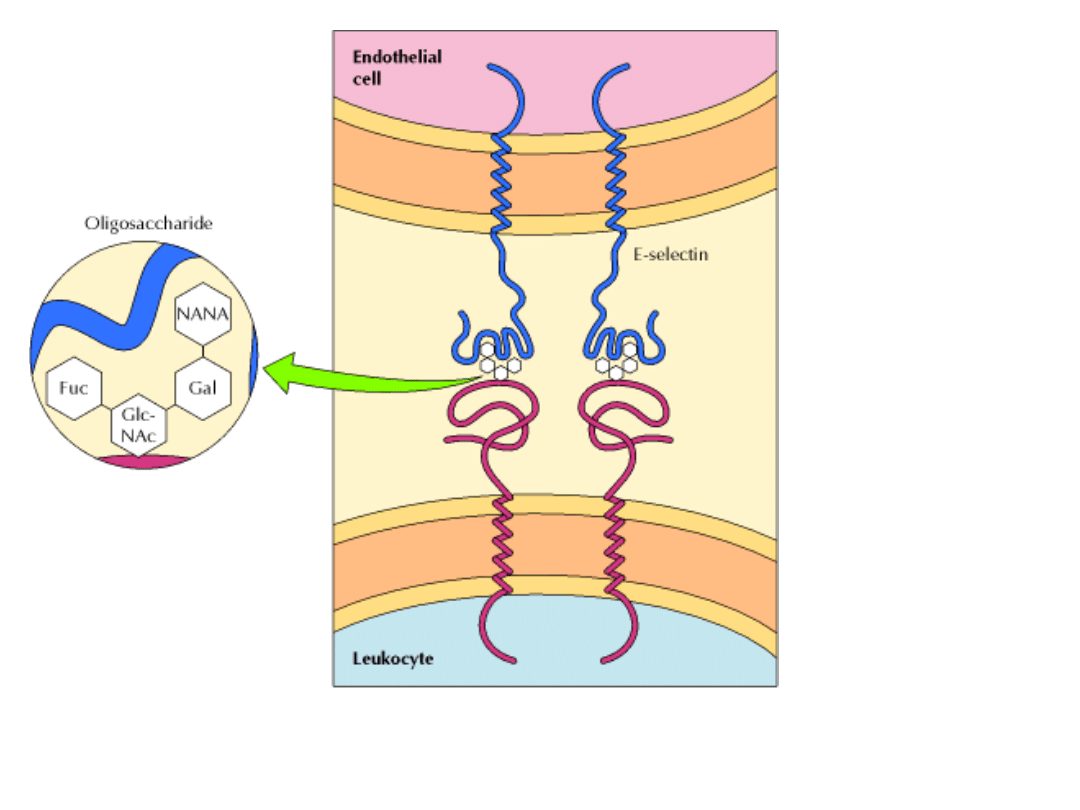
Przykładem może być rozpoznawanie charakterystycznych
oligosacharydów na
powierzchni komórkowej leukocytów.
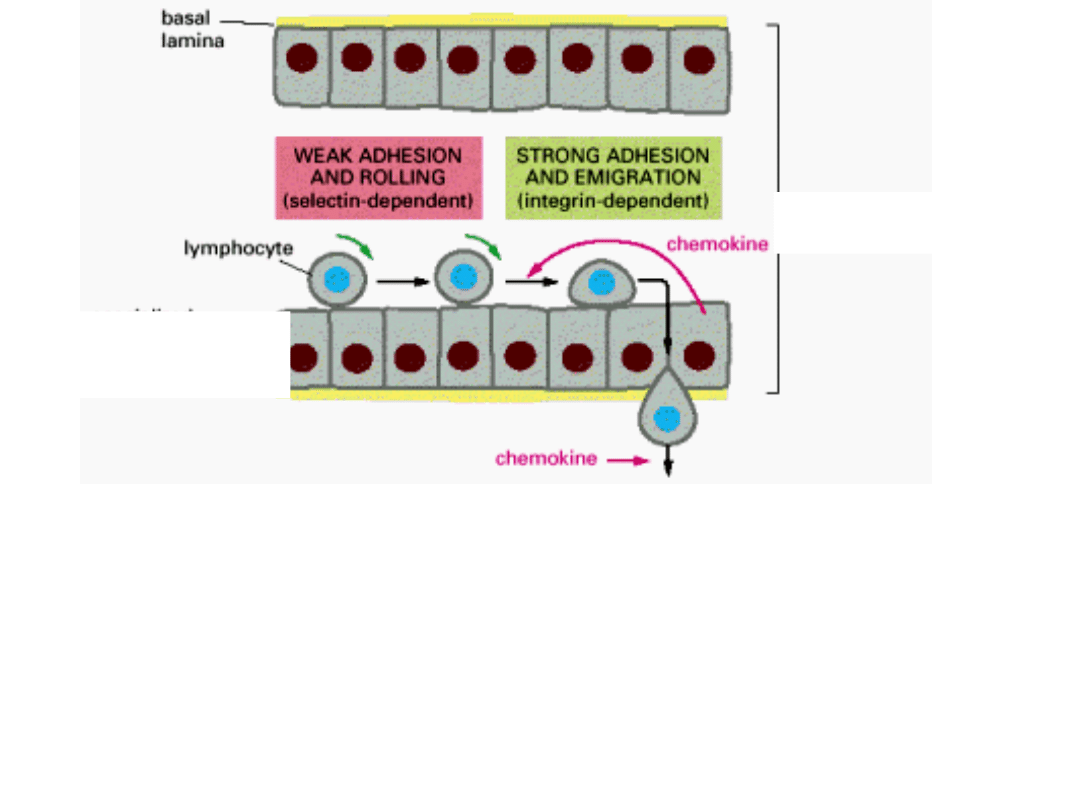
naczynie
krwionośne
wyspecjalizowana
komnórka
śródbłonka
1. Chemiczne sygnały z miejsca zakażenia komórki śródbłonka wytwarzają
wyspecjalizowane białka sródbłonowe – LEKTYNY.
2. Lektyny rozpoznają poszczególne grupy cukrów glikolipidów i glikoprotein powierzchni
limfocytów krążących we krwi.
3. Stymulowane przez chemokiny limfocyty mocno przylegają do powierzchni komórek
śródbłonka
4. Limfocyty przeciskają się między komórkami śródbłonka i docierają do miejsca
zakażenia.
INNY PRZYKŁAD – Oligosacharydy glikokaliksu biorą udział w rozpoznawaniu komórki
jajowej przez plemnik.
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
Wyszukiwarka
Podobne podstrony:
Blony komorkowe
09 Biochemia blony komorkowe
Struktura błony komórkowej, biochemia
RECEPTORY BLONY KOMORKOWEJ I ORGANELLI
błony komórkowe
BŁONY KOMÓRKOWE, Studia - materiały, semestr 6, Fizjologia żywienia
71 Potencjal blony komorkowej
BŁONY KOMÓRKI
Błony komórkowe, Fizjoterapia, Fizjologia
blony komorki
Transport przez błony komórki
0202 04 03 2009, wykład nr 2 , Budowa i funkcje błony komórkowej oraz transport przez błony(1)
1 cw BLONY KOMORKOWEid 9083 Nieznany (2)
Błony komórkowe
61 Dyfuzja czasteczek bialek w plaszczyznie blony komorkowej
więcej podobnych podstron