
Biologia komórki
Błony biologiczne
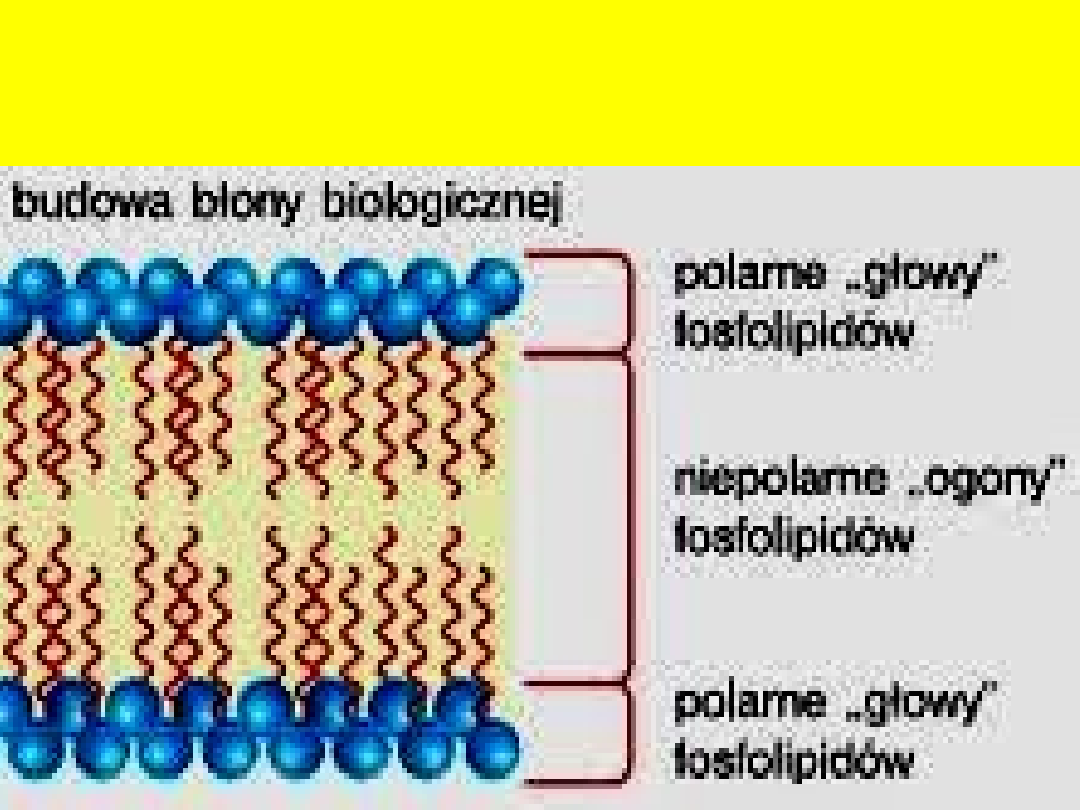
Fosfolipidy (fosfoglicerydy) są głównymi składnikami błon lipidowych
Fosfolipidy to estry glicerolu, kwasów tłuszczowych i kwasu ortofosforowego
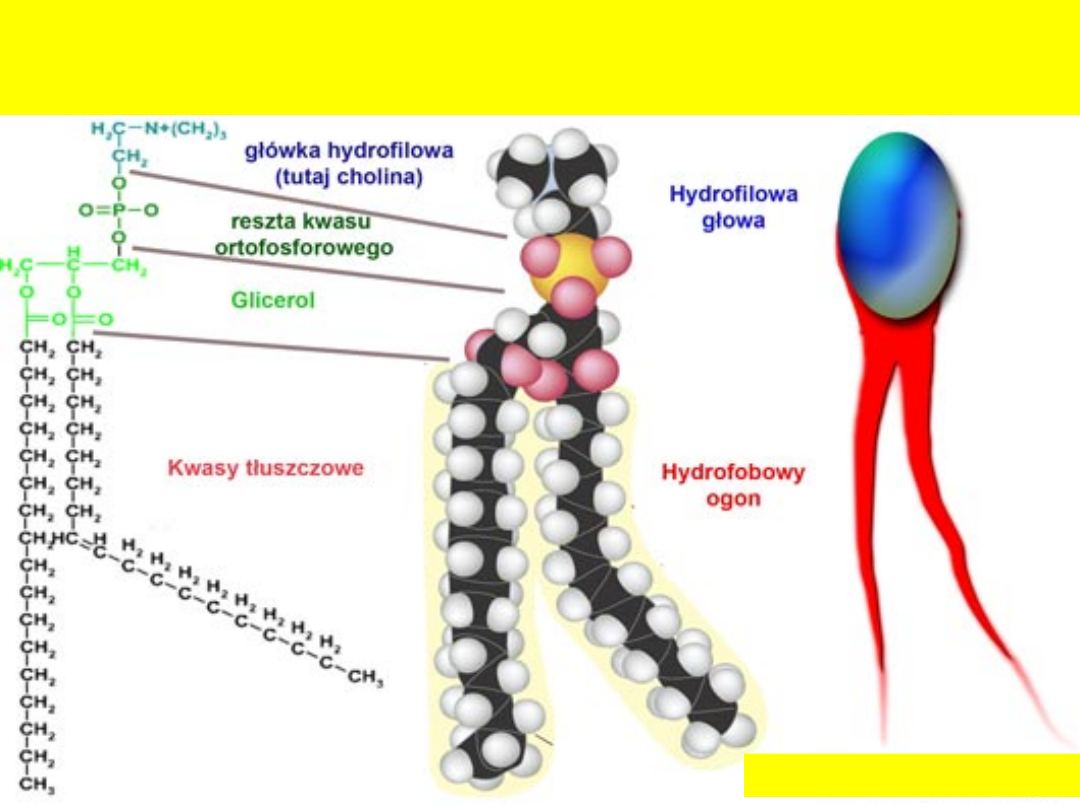
Fosfolipidy występują głównie w formie estru z dwoma kwasami tłuszczowymi
i jedną resztą fosforanową, do której przyłączona jest grupa hydrofilowa
Polarność fosfolipidu
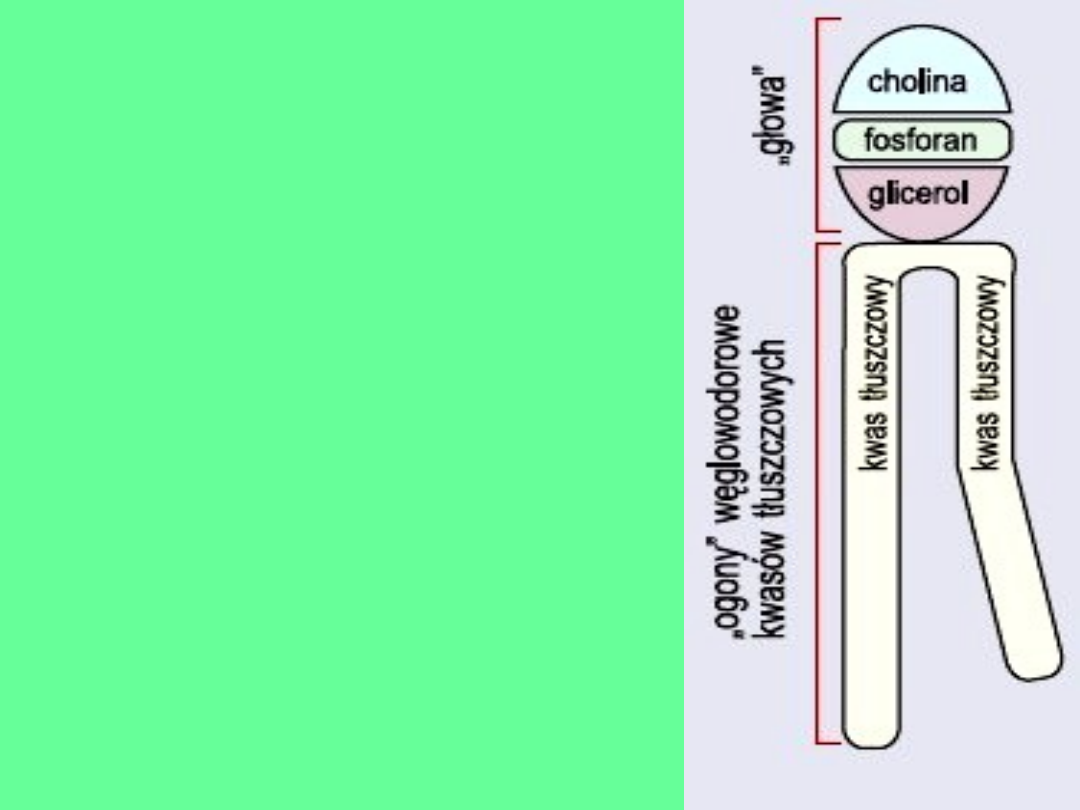
Fosfolipidy ze względu na swoje
właściwości amfipatyczne (jeden
koniec hydrofobowy, drugi
hydrofilowy, a odległość między oba
końcami jest bardzo duża), w
środowisku wodnym skupiają się ze
sobą częściami hydrofobowymi
wystawiając do polarnej wody
końcówkę hydrofilową.
Wyobraźmy sobie ogromną liczbę szpilek, które
mają metalowe główki i plastikowe ostrza,
zbliżenie magnesu do rozsypanych szpilek
wymusza na nich przyjmowanie zawsze takich
samych układów przestrzennych.
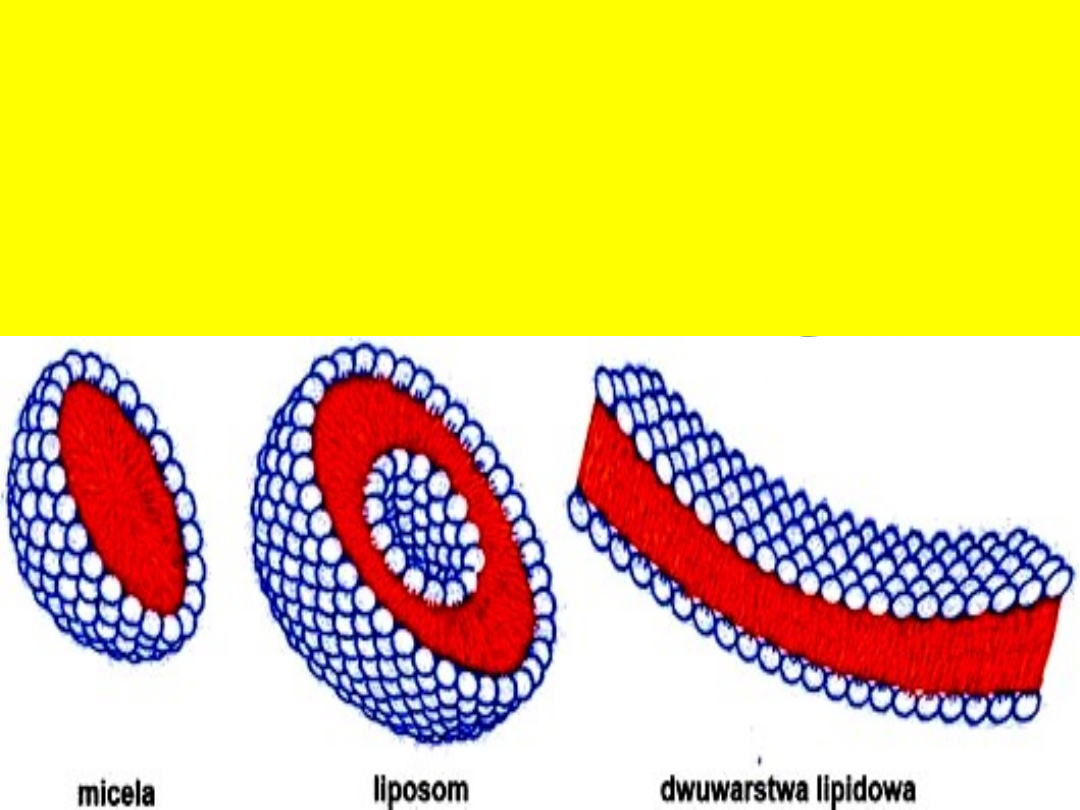
Dlatego fosfoglicerydy w środowisku
wodnym tworzą zawsze trzy typy
struktur: micele, liposomy i
dwuwarstwy lipidowe.
Proporcje i typy lipidów w warstwach wewnętrznej i zewnętrznej błony
komórkowej są odmienne.
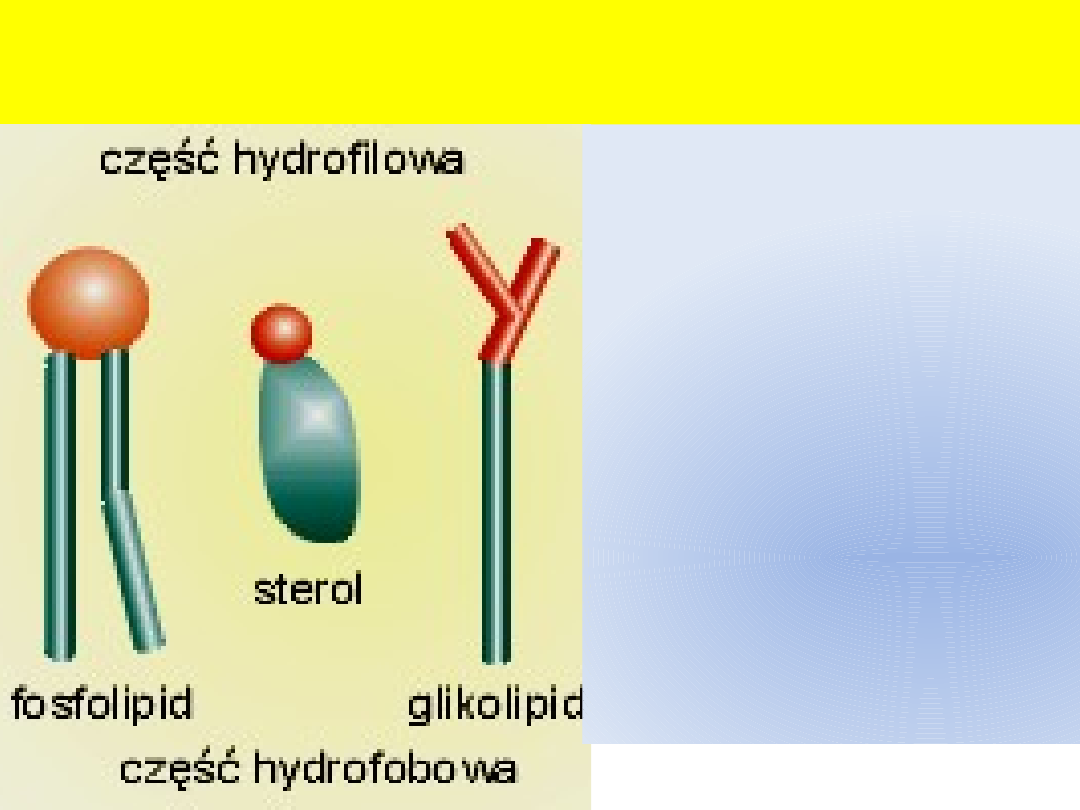
Cząsteczki chemiczne budujące błony
biologiczne
Amfifilowość,
amfipatyczność
- cecha
wskazująca na polarność
budowy cząsteczki
chemicznej.
Jeden z biegunów cząsteczki
jest hydrofilowy, drugi
hydrofobowy.
Amfipatyczność
fosfolipidów jest podstawą
ich samoorganizacji w
struktury dwuwarstwowe,
charakterystyczne dla błon
biologicznych.
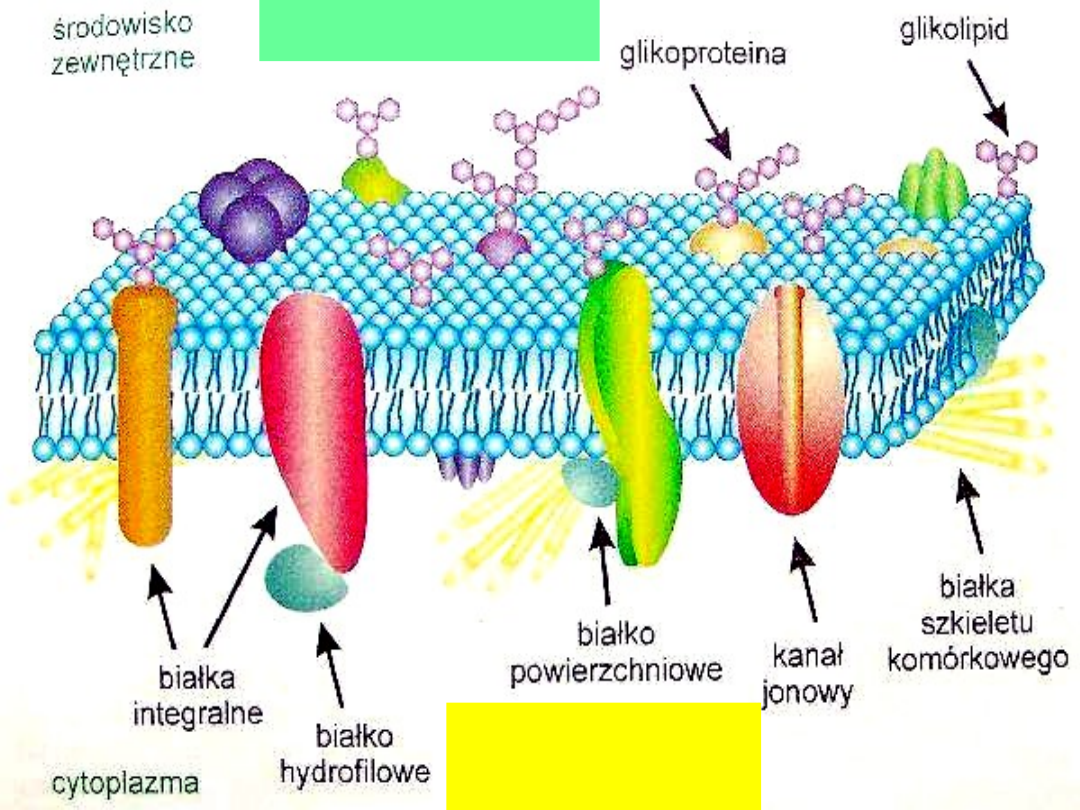
Budowa błon
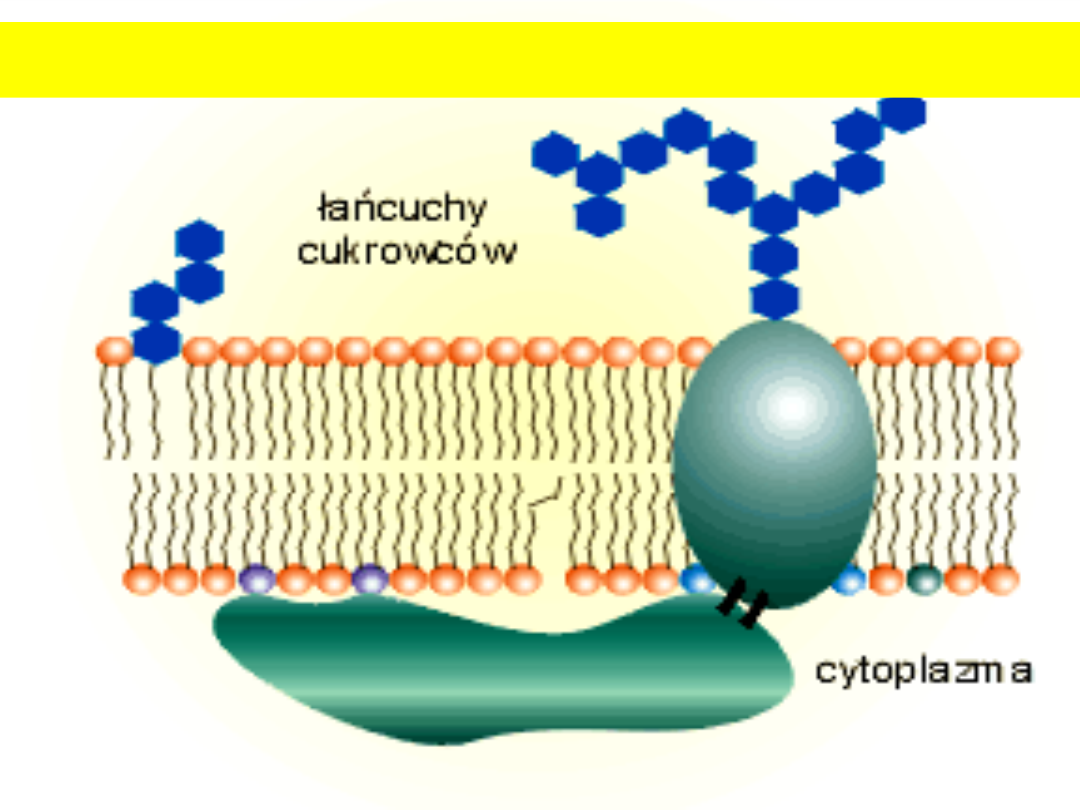
glikolaliks
Kora
komórki
Glikolipidy
zawierają cukier,
którym najczęściej
jest galaktoza lub
laktoza
.
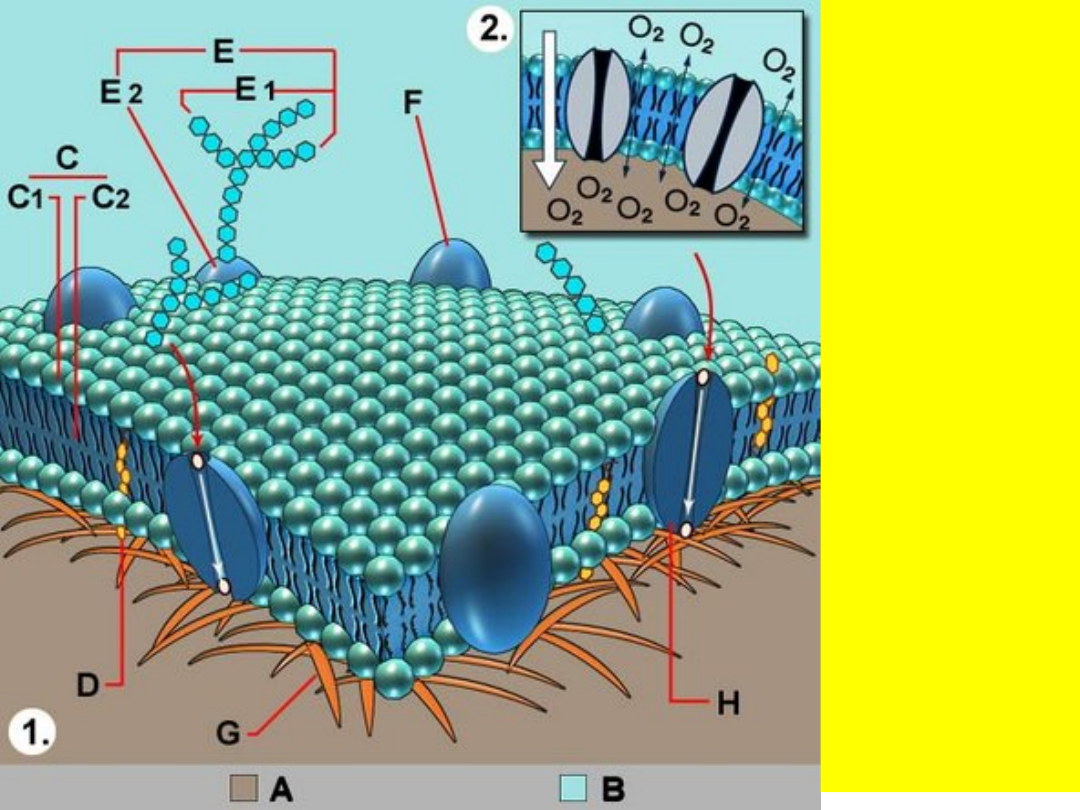
1
. Schemat
błony
komórkowej:
A -
cytoplazma,
B -
płyn
pozakomórko
-wy,
C
-
fosfolipid,
C1
- głowa
cząsteczki,
C2
- ogon
cząsteczki,
D
- cholesterol,
E -
glikoproteina
E1
cukier,
E2
- białko,
F
-
receptor
białkowy,
G
-
cytoszkielet,
H
transport
białka.
2
. Transport
przez błonę
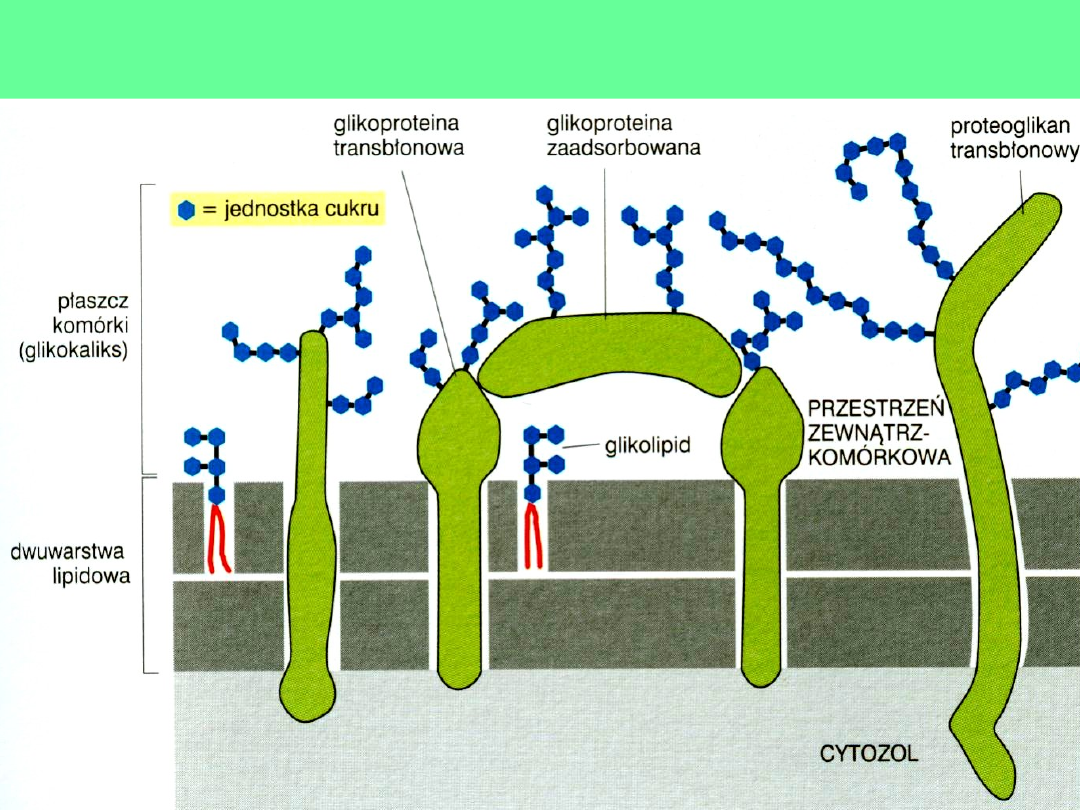
Glikokaliks- cukrowe znaczniki błony

Błony fosfolipidowe
są
płynną mozaiką. Wiele
składników błonowych jest w
stanie ciągłej dynamicznej
płynności.
Błony komórkowe nie są
homogenne.
Składniki błon organelli
komórkowych oraz błony
komórkowej różnią się od
siebie.
Systemy te tworzą przedziały
wewnątrzkomórkowe,
utrzymujące różnice pH i
składu jonowego w
poszczególnych częściach
komórki.
Komponenty błonowe są
asymetryczne.
Stan skupienia i
składowe błon
fosfolipidowych

Flipazy
– enzymy znajdujące się wewnątrz błon
fosfolipidowych, przenoszące fosfolipidy z jednej warstwy
błony do drugiej.
Odgrywają istotną rolę podczas regeneracji błony oraz w
utrzymaniu asymetrii błon - asymetrycznego
rozmieszczenia fosfolipidów w każdej z dwóch warstw
danej błony
.
Energii potrzebnej do działania tych białek dostarcza
sprzężony proces hydrolizy ATP.
W normalnych warunkach
flip-flop
, czyli proces
przerzucenia fosfolipidu z jednej strony błony na
drugą
, trwa bardzo długo ze względu na amfipatyczny
charakter tych lipidów i samą budowę błon.
Flipazy
skracają czas trwania tego procesu
z kilku dni
do kilku minut.
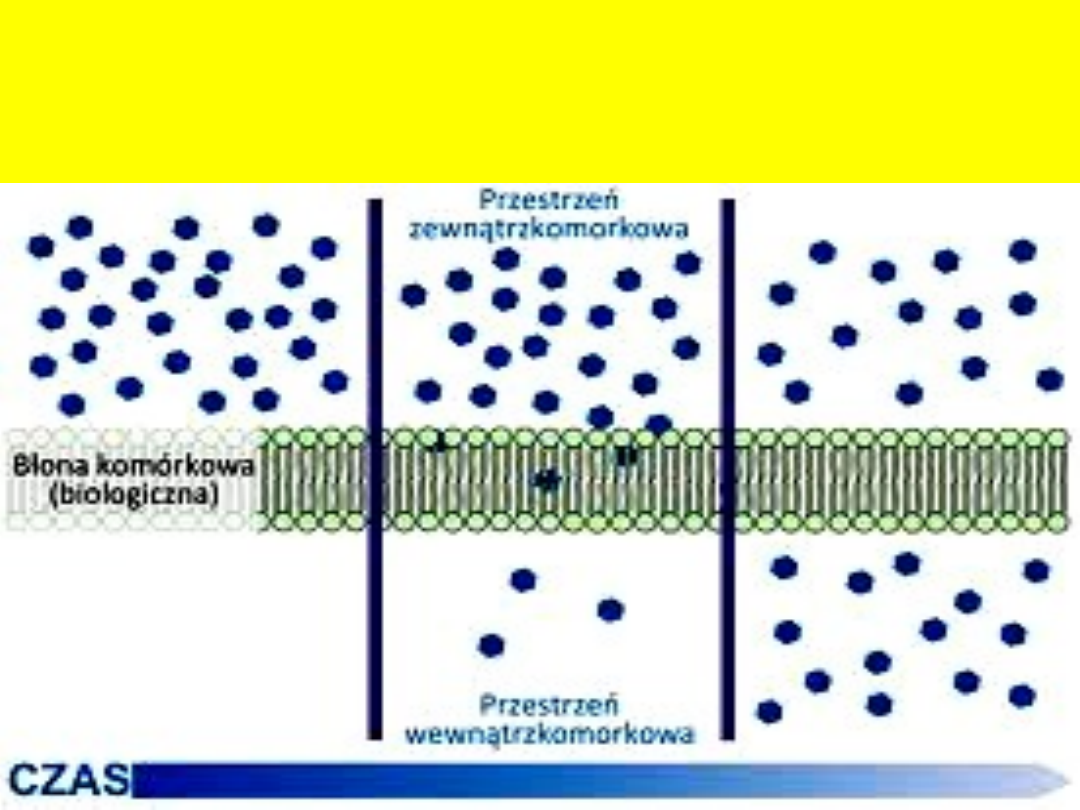
Osmoza – dyfuzja rozpuszczalnika przez błonę półprzepuszczalną
rozdzielającą dwa roztwory o różnym stężeniu. Osmoza spontanicznie
zachodzi od roztworu o wyższym stężeniu substancji rozpuszczonej do
roztworu o niższym, czyli prowadzi do wyrównania stężeń obu roztworów
.

Roztwór o niższym stężeniu nazywa się
hipotonicznym, roztwór o wyższym stężeniu -
hipertonicznym.
Gdy roztwory pozostają w równowadze osmotycznej,
mówi się że są względem siebie izotoniczne.
Zjawisko odkryto, gdy badano plazmolizę komórek skórki liści
zapasowych cebuli – dotyczyło więc żywych komórek, w
których błoną półprzepuszczalną jest błona komórkowa, a
roztworami były: zawartość komórki i płyn pod szkiełkiem
nakrywkowym preparatu mikroskopowego.
Później nauczono się wytwarzać sztuczne błony
półprzepuszczalne (np. celofan) i okazało się, że samo zjawisko
nie jest specyficzną cechą życia. Może zachodzić w różnych
rozpuszczalnikach i dotyczyć różnych substancji
rozpuszczonych.
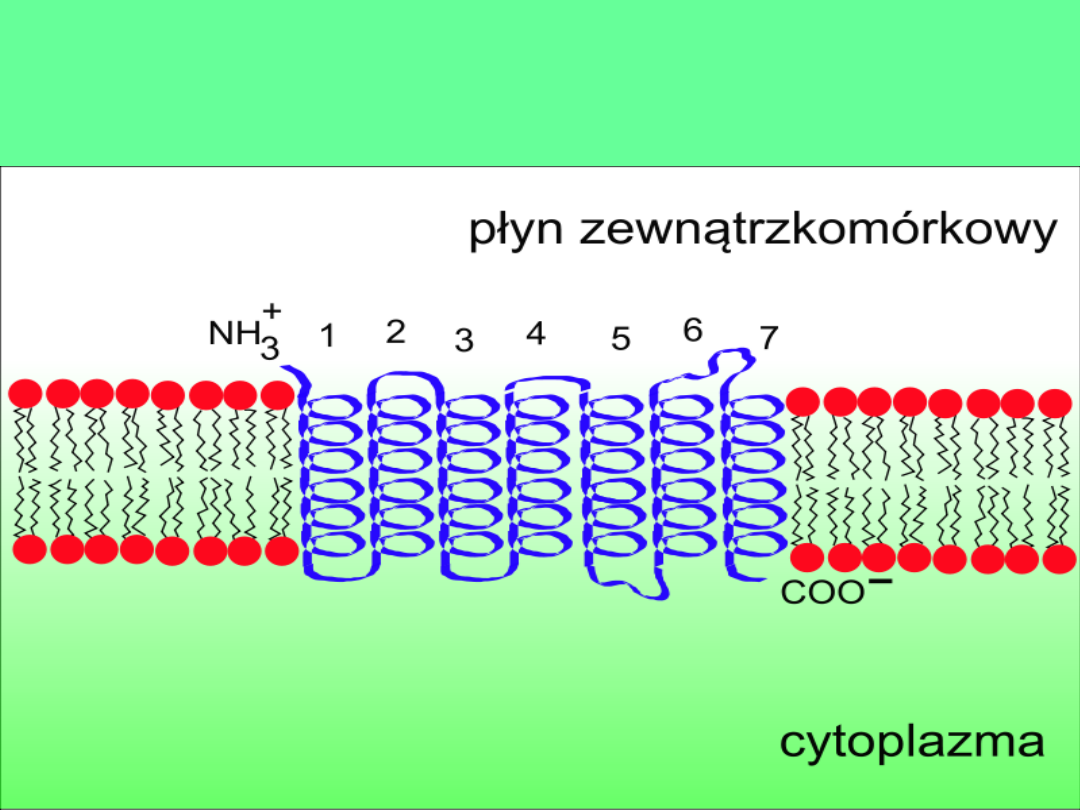
Schemat receptora błonowego z siedmioma
domenami (helisami) transbłonowymi
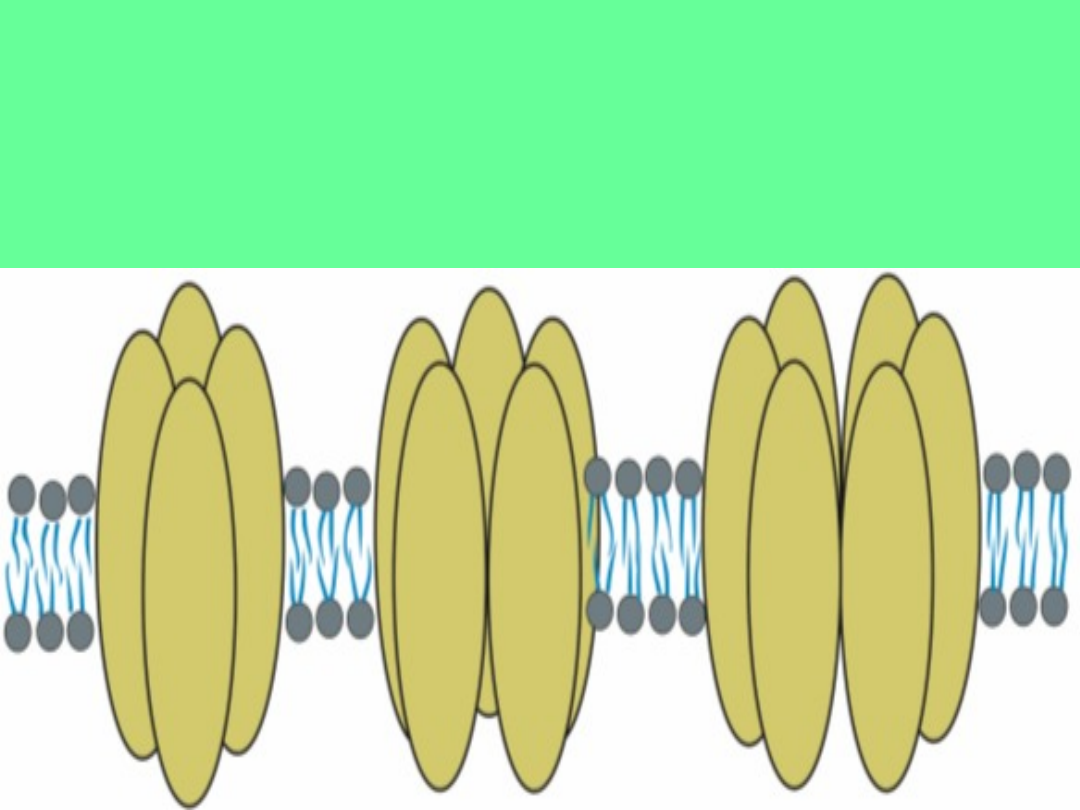
Kanały błonowe
składają się z 4, 5 lub 6 podjednostek.
Wraz z liczbą domen wzrasta średnica kanału.
Środkowa oś symetrii wyznacza por, czyli bramkowaną i
wypełnioną wodą drogę dla jonów
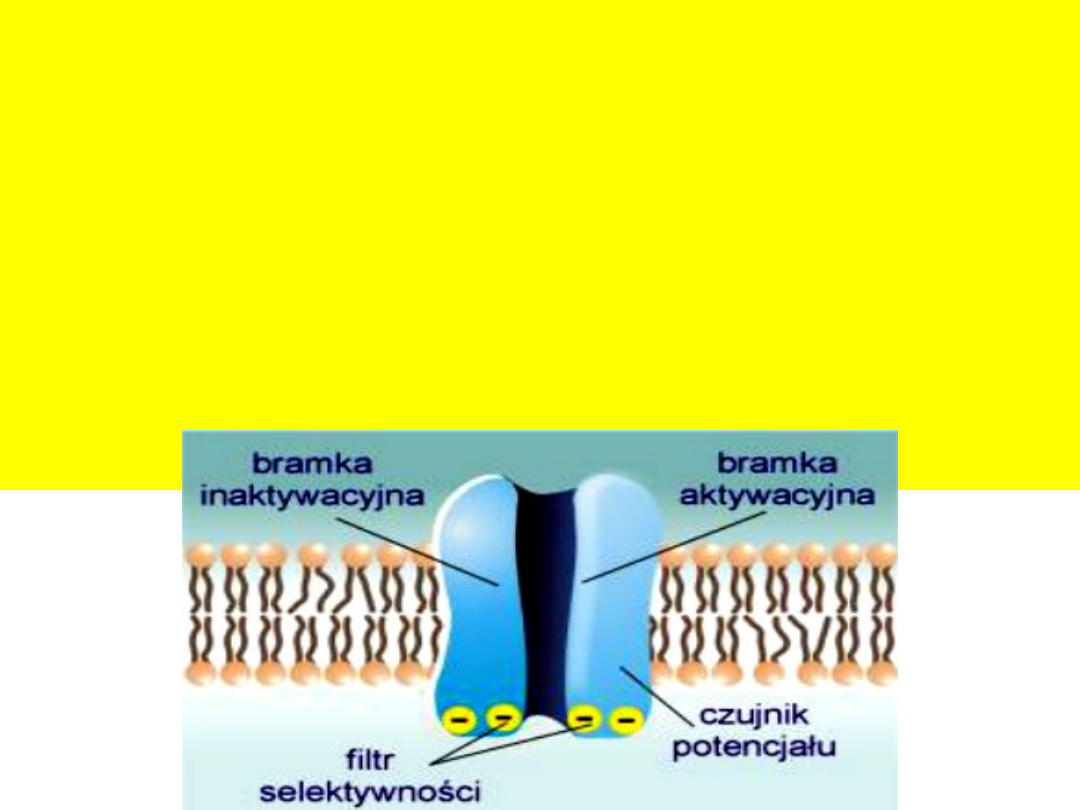
Kanały jonowe
to białka integralne posiadające zdolność do
kontrolowanego przepuszczania jonów. Por wodny ulega otwarciu lub
zamknięciu w zależności od czynników zewnętrznych.
Ze względu na rodzaj czynnika otwierającego (aktywującego) kanały jonowe,
dzieli się je na 3 zasadnicze grupy:
- kanały bramkowane napięciem,
- kanały bramkowane ligandem,
- kanały bramkowane naprężeniem mechanicznym.
Istotną cechą kanałów jest ich selektywność. Mówi się o kanałach
kationowych lub anionowych, a gdy kanały są jeszcze bardziej
„wyspecjalizowane” to określamy je jako sodowe, potasowe itd.
Określenie: np. kanał sodowy – oznacza jedynie, iż kanał ten najlepiej
przepuszcza jony sodu. Oprócz nich, choć znacznie gorzej, mogą przez ten
kanał przechodzić także inne kationy.

Potencjałem błonowym
lub membranowym nazywa
się różnicę potencjałów pomiędzy wnętrzem komórki i jej
otoczeniem (V
m
= V
w
- V
z
). Potencjał po zewnętrznej stronie błony
wynosi zero.
Napięcie pomiędzy obu stronami błony jest
potencjałem wnętrza komórki mierzonym względem jej otoczenia.
Większość żywych
komórek utrzymuje
stałą wartość różnicy
potencjałów
pomiędzy swym
wnętrzem a
otoczeniem
Płyny wewnątrz- i
zewnątrzkomórkowy różnią się
istotnie pod względem
stężenia w nich
poszczególnych jonów
.
Przez
błonę komórkową ustala się
gradient stężenia oraz
gradient ładunków
elektrycznych poszczególnych
jonów
W płynie wewnątrzkomórkowym
przeważają organiczne aniony, które
nie mają zdolności przechodzenia
przez błonę komórkową. Błona
komórkowa ulega więc polaryzacji:
po jej wewnętrznej stronie występuje
ładunek ujemny - wynikający z
nagromadzenia anionów, natomiast
na zewnątrz komórki - ładunek
dodatni
.

Potencjał spoczynkowy
to stały potencjał wnętrza komórki
względem jej otoczenia.
Zdolność komórki do utrzymywania stałej wartości potencjału
spoczynkowego związana jest bezpośrednio z istnieniem różnicy
stężeń niektórych jonów pomiędzy wnętrzem i otoczeniem komórki.
Dla większości komórek jonami "najważniejszymi„ z punktu widzenia
potencjału spoczynkowego są jony sodu, potasu oraz chlorkowe
.
Rodzaj jonu
Stężenie zewnątrz komórki
Stężenie wewnątrz komórki
K
+
4 mM
140 mM
Na
+
144 mM
ok. 10 mM
Cl
-
114 mM
ok. 4 mM
Na zewnątrz komórki stężenie jonów sodowych i chlorkowych jest
większe niż wewnątrz komórki, natomiast stężenie jonów potasu jest
większe wewnątrz komórki.
Stała wartość potencjału błonowego może być utrzymana jedynie wtedy, gdy
całkowity ładunek przepływający przez błonę jest równy zeru (w przeciwnym
razie następowałaby zmiana ładunku błony i związana z tym zmiana
potencjału).
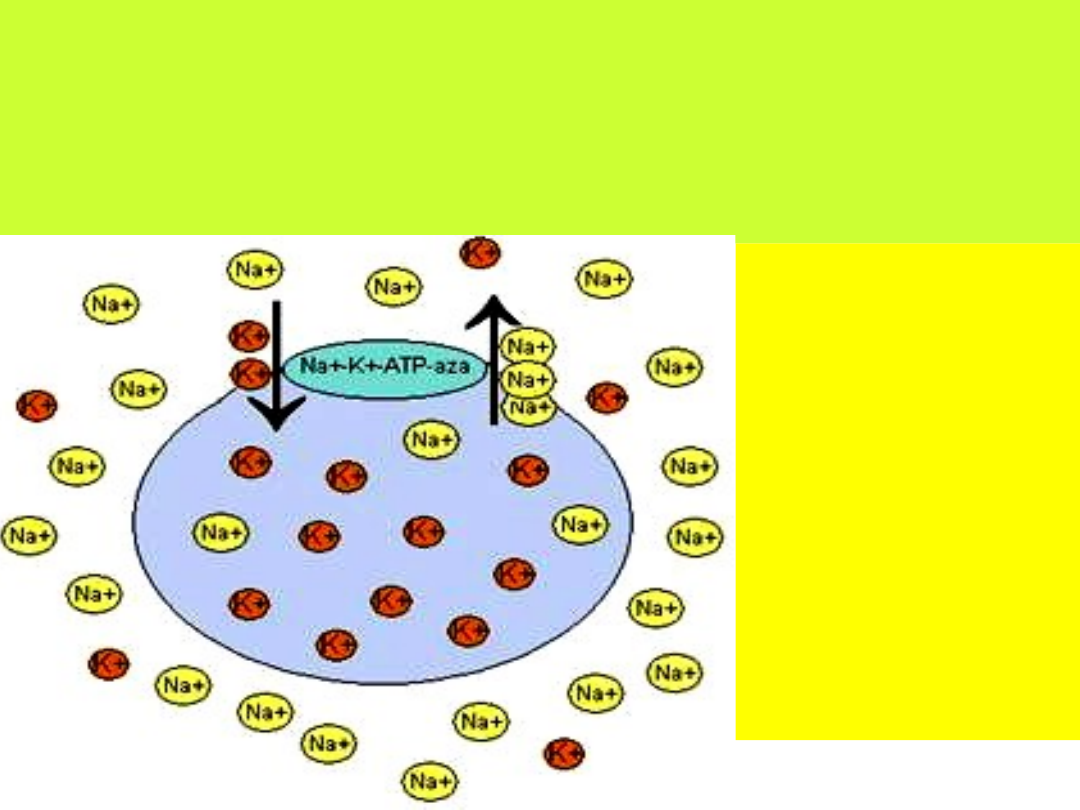
Pompa Na
+
↔K
+
Aby utrzymać wysokie stężenie jonów potasowych oraz niskie stężenie jonów sodowych
wewnątrz komórki, konieczny jest aktywny transport tych jonów przez błonę komórkową
wbrew gradientowi ich stężeń.
Enzym zwany ATP-azą sodowo-potasową zapewnia odpowiednie rozmieszczenie jonów i
utrzymanie ich gradientu przez błonę komórkową.
Jony sodowe zgodnie z
gradientem wnikają do
wnętrza komórki przez
kanały sodowe. Jednak
wbudowana w błonę
komórkową ATP-aza
sodowo- potasowa,
wiąże jony sodu i
wypompowuje je na
zewnątrz.
Jednocześnie
po zewnętrznej stronie
błony komórkowej
enzym przyłącza jony
potasu i wpompowuje
je do wnętrza komórki.

Zjawiska pobudliwości i pobudzenia
komórkowego.
Błonowy potencjał
spoczynkowy w komórkach
pobudliwych
Cechą komórek pobudliwych jest posiadanie stałego,
ujemnego, spoczynkowego potencjału błonowego. Jego
wartości podane są poniżej.
Rodzaj komórki:
Neuron:
Wartość potencjału błonowego: -60mV do -80mV
Komórka mięśnia poprzecznie prążkowanego
:
Wartość potencjału błonowego: -80mV do -90mV
Otwieranie i zamykanie kanałów jonowych zależy
bezpośrednio od tego w jakim stanie czynnościowym
znajduje się błona komórkowa.

Pobudliwość jest zdolnością komórki do
odpowiedzi na bodziec
.
Tkanki pobudliwe zbudowane są z komórek, mających zdolność do
szybkiej reakcji na bodźce.
Zalicza się do nich: tkankę mózgową i nerwy zbudowane z komórek
nerwowych, oraz tkankę mięśniową gładką, poprzecznie prążkowaną
oraz sercową zbudowaną z komórek mięśniowych.
Bodźce np. w postaci krążących w przestrzeni zewnątrzkomórkowej
substancji chemicznych (ligandów) mają zdolność wiązania się ze
specyficznymi dla siebie receptorami.
To połączenie może skutkować:
- otwarciem kanałów jonowych, co umożliwia przepływ odpowiednich
jonów przez błonę komórkową;
- aktywacją enzymów znajdujących się w błonie komórkowej
.
Odpowiedź na bodziec w postaci opisanych wyżej zmian błony
komórkowej zachodzi bardzo szybko - w ułamku sekundy.
Następujące po niej zmiany w metabolizmie komórki
zachodzą znacznie wolniej.
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Cząsteczki chemiczne budujące błony biologiczne
- Slide 8
- Slide 9
- Glikokaliks- cukrowe znaczniki błony
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
Wyszukiwarka
Podobne podstrony:
RECEPTORY BLONY KOMORKOWEJ I ORGANELLI
0202 04 03 2009, wykład nr 2 , Budowa i funkcje błony komórkowej oraz transport przez błony(1)
wykład4 reg cyklu kom i organelle komórkowe
Wykład piąty biologia komórki
EgzaminMikrobPytania2008, chemia organiczna, biologia ewolucyjna-wykłady, genetyka, biologia komórki
Fizjologia zwierząt wszystkie opracowania, chemia organiczna, biologia ewolucyjna-wykłady, genetyka,
biol.medyczna, wykład - Ogólna budowa komórki, Ogólna budowa komórki
Wykład 7 Embriologia Błony płodowe
komórka, Organelle komórki, Organelle komórki
Biochemia wykład 12 Błony biologiczne
Blony komorkowe
09 Biochemia blony komorkowe
Struktura błony komórkowej, biochemia
błony komórkowe
WYKLADY z biomórki, biologia komórki
więcej podobnych podstron