
BŁONY BIOLOGICZNE
I.
Funkcje
II.
Ogólna budowa błon
III. Składniki lipidowe błon
IV. Białka integralne i peryferyjne
V.
Przepuszczalność błon biologicznych

I. FUNKCJE BŁON
a) Błony komórkowe (plazmatyczne)
1.
Oddzielenie środowiska wewnętrznego komórki od otoczenia;
2.
Pośrednictwo w integracji komórek z macierzą pozakomórkową;
3.
Utrzymywanie gradientu stężeń pewnych metabolitów pomiędzy
wnętrzem komórki a środowiskiem zewn.;
4.
Ochrona komórki;
5.
Zapewnienie kształtu;
6.
Receptory błonowe – odbieranie sygnałów spoza komórki,
rozpoznawanie się komórek;
7.
Transport różnych substancji do wnętrza i na zewnątrz komórki.

I. FUNKCJE BŁON
b) Błony wewnątrzkomórkowe:
•
Podział wnętrza komórki na liczne przedziały (są silnie
rozbudowane);
•
Wykorzystanie procesów transportu wewnątrz komórki do
wytwarzania i gromadzenia energii;
•
Umożliwienie zachodzenia przeciwstawnych procesów w
komórce;
•
Magazynowanie jonów i różnych substancji w pewnych okolicach
komórki;
•
Transport cząsteczek w określonych kierunkach wewnątrz
komórki.

II. OGÓLNA BUDOWA BŁON
• Grubość ok. 7-10nm;
• Składają się gł. z lipidów i białek, w
niektórych błonach występują dodatkowo
składniki cukrowe związane z białkami
(glikoproteiny) lub z lipidami (glikolipidy);
• Zrąb błon stanowi tzw. dwuwarstwa
lipidowa (warstwy E i P).

III. SKŁADNIKI LIPIDOWE BŁON
Są to:
1. Glicerofosfolipidy (50-90%)
2. Sfingolipidy (do 10%)
3. Cholesterol

III. SKŁADNIKI LIPIDOWE BŁON
Fosfolipidy i sfingolipidy są związkami amfipatycznymi -
posiadają hydrofilową głowę (zwykle gr. fosforanowa,
wiążąca estrowo inny związek polarny – serynę,
etanoloaminę, cholinę) i hydrofobowy ogon (łańcuchy
węglowodorowe kwasów tłuszczowych lub aminoalkoholi –
sfingozyny lub dihydrosfingozyny) .
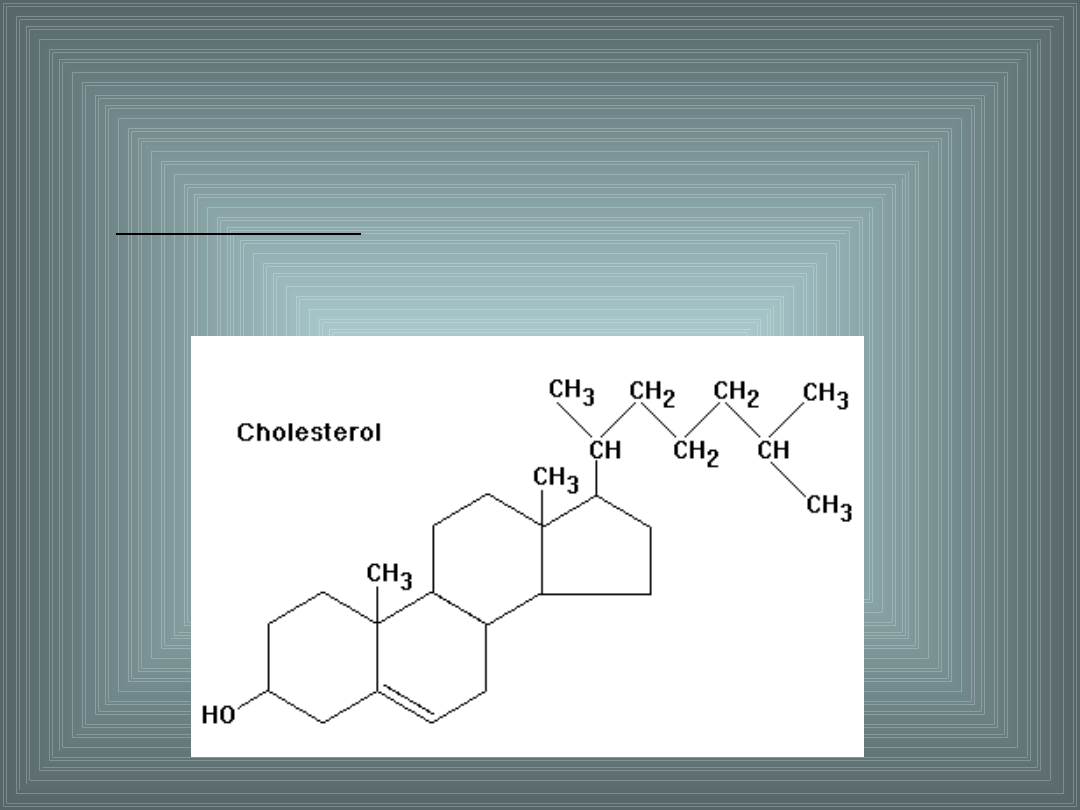
Cholesterol jest najbardziej hydrofobowym składnikiem
błon. Nie posiada hydrofilowej głowy - charakterystycznej
dla większości lipidów błonowych i hydrofobowego ogona.
Jego jedynym hydrofilnym składnikiem jest gr. –OH
związana z C
3
– wbudowanym z strukturę pierścienia.
Pozostałe elementy składowe cholesterolu mają charakter
hydrofobowy.
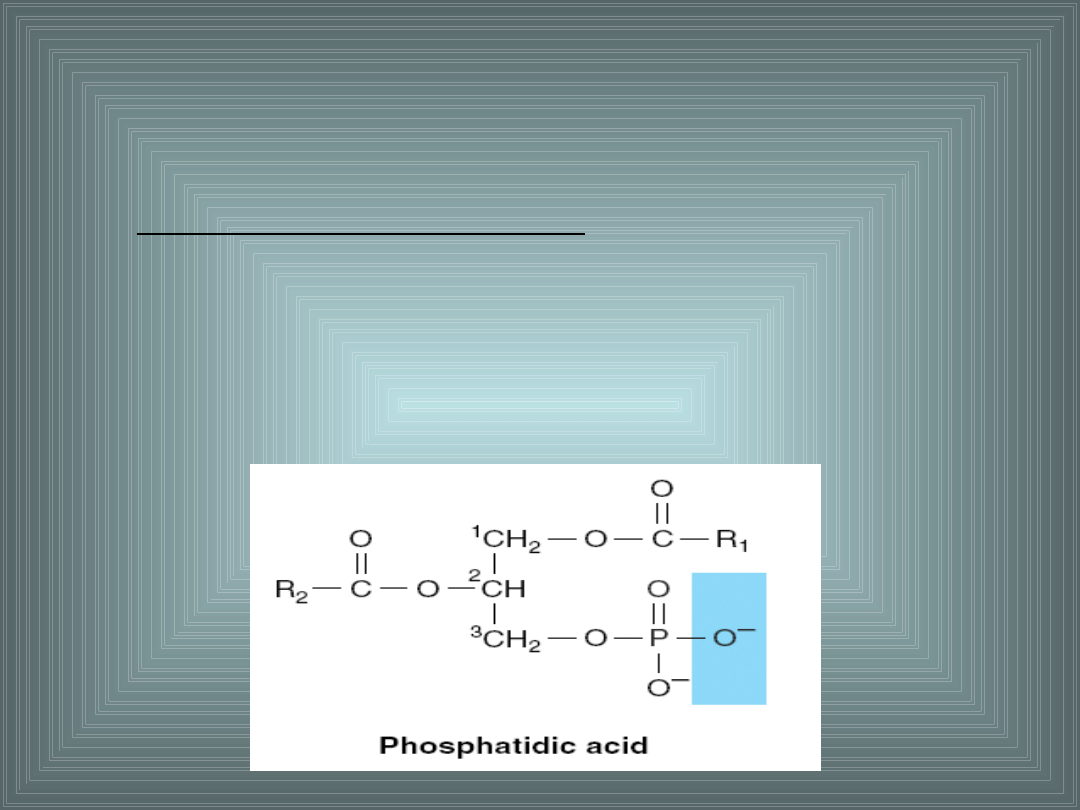
III. SKŁADNIKI LIPIDOWE BŁON
1. GLICEROFOSFOLIPIDY.
Są to estry glicerolu z kwasem fosforowym i kwasami
tłuszczowymi.
Kwas fosfatydowy – najprostszy fosfolipid, prekursor
pozostałych fosfolipidów.

III. SKŁADNIKI LIPIDOWE BŁON
Synteza glicerofosfolipidów
- polega na przyłączeniu kwasu fosfatydowego z
CDP-diacyloglicerolu do odpowiedniego alkoholu, lub na
wiązaniu 1,2-diacyloglicerolu z estrem fosforanowym
odpowiedniego alkoholu, odłączanego od CDP-alkoholu;
w końcowym etapie odłącza się CMP
- są syntetyzowane w gładkiej siateczce
endoplazmatycznej, stąd transportowane do aparatu
Golgiego, a następnie do błon organelli komórkowych i
do błony plazmatycznej, gdzie stają się ich składnikami
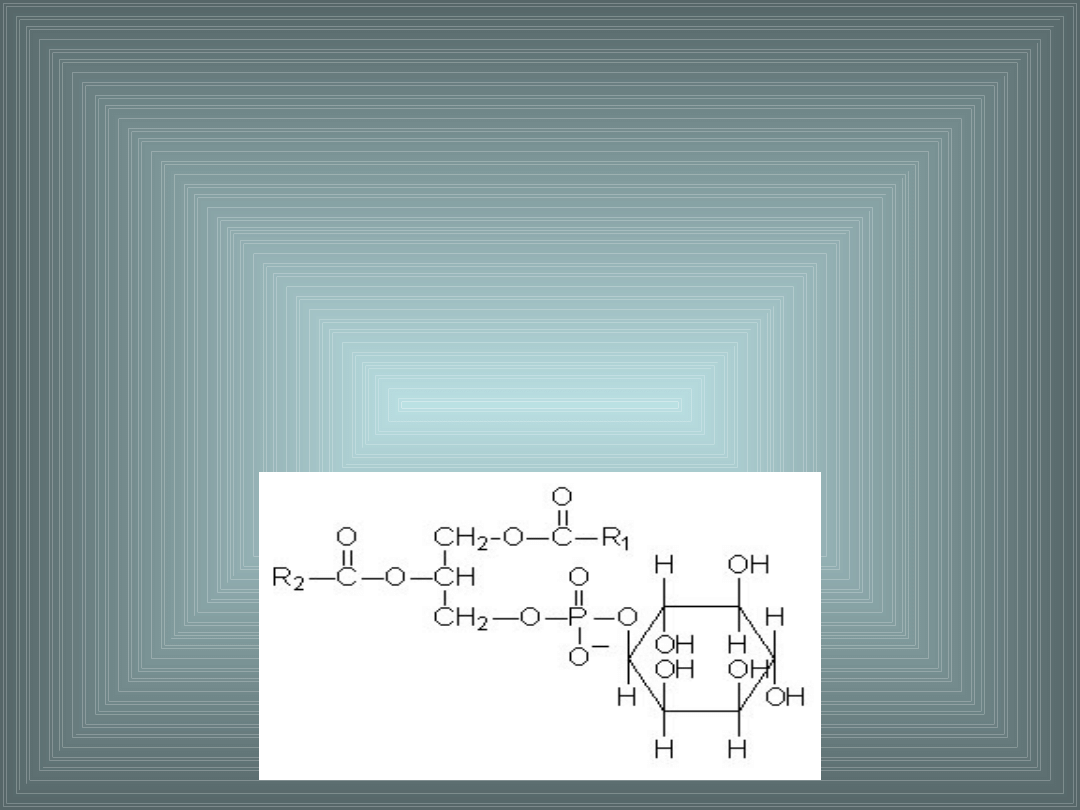
III. SKŁADNIKI LIPIDOWE BŁON
FOSFATYDYLOINOZYTOL:
- uczestniczy w zakotwiczaniu białek w błonach biologicznych
poprzez połączenie łańcucha oligosacharydowego białka
wiązaniem
O-glikozydowym z inozytolem fosfatydyloinozytolu
- występuje gł. w warstwie P
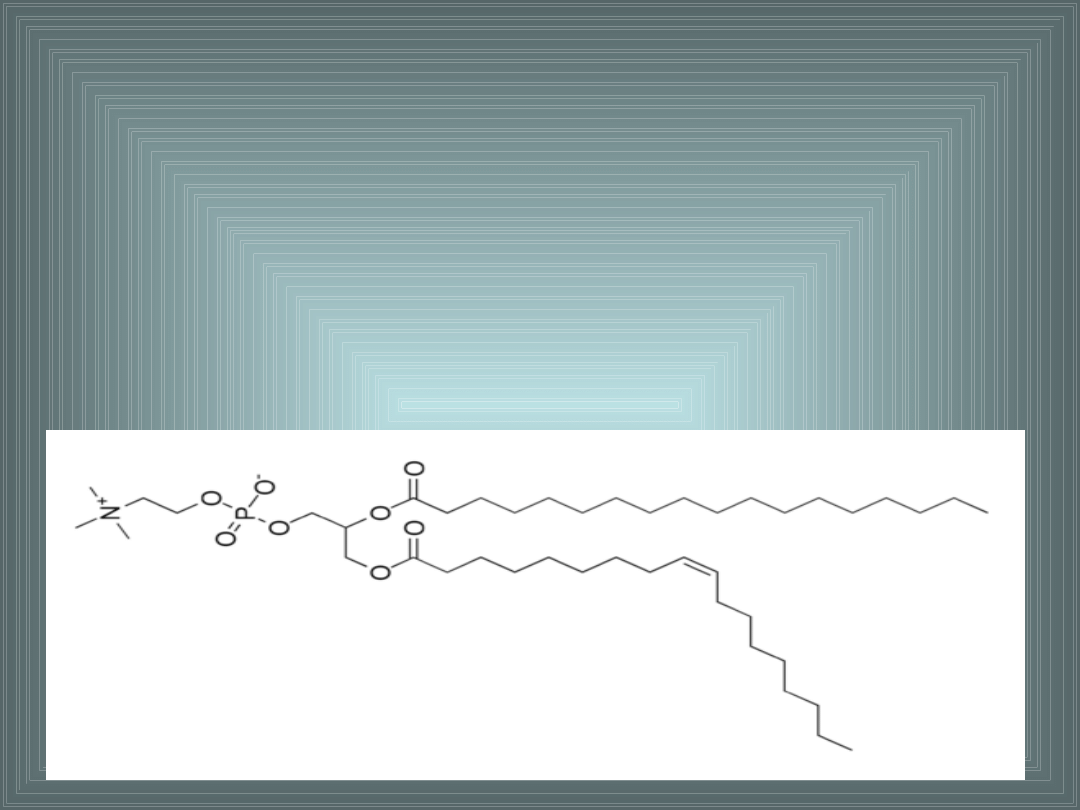
III. SKŁADNIKI LIPIDOWE BŁON
FOSFATYDYLOCHOLINA (lecytyna):
- reszta fosforanowa zestryfikowana choliną
- występuje gł. w warstwie E dwuwarstwy lipidowej
- przeważa ilościowo nad innymi lipidami błonkomórkowych
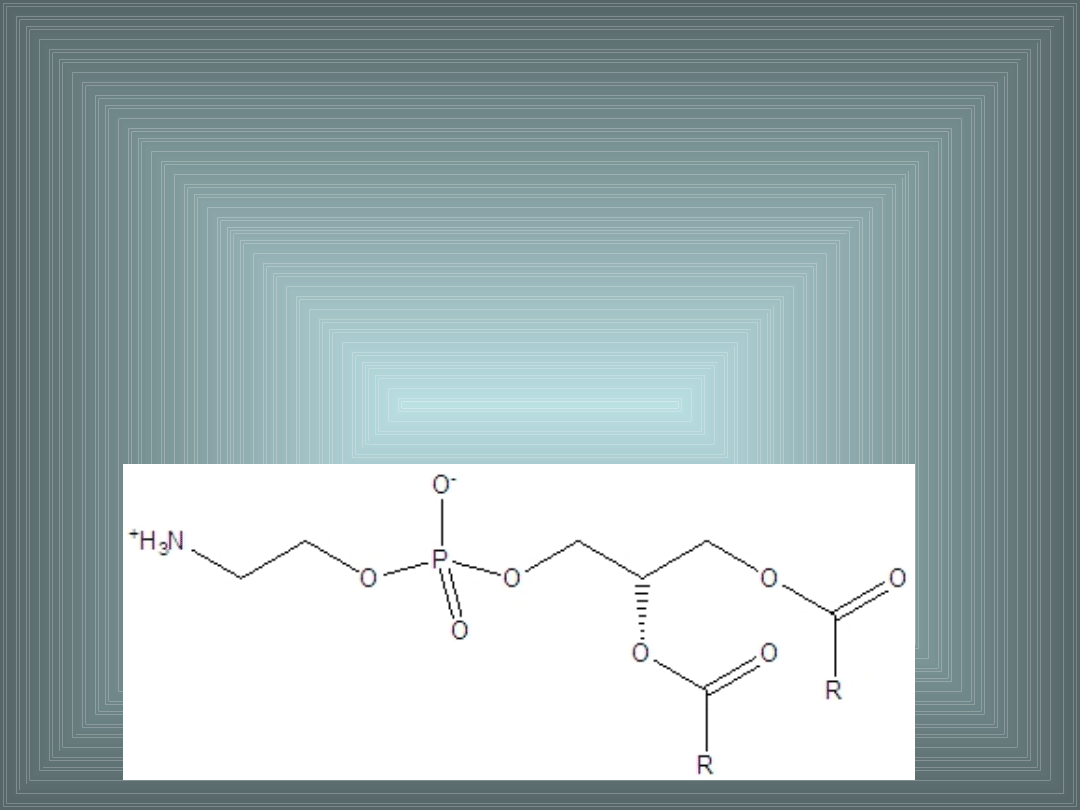
III. SKŁADNIKI LIPIDOWE BŁON
FOSFATYDYLOETANOLOAMINA(kefalina):
- reszta fosforanowa zestryfikowana etanoloaminą
- występuje gł. w warstwie P dwuwarstwy
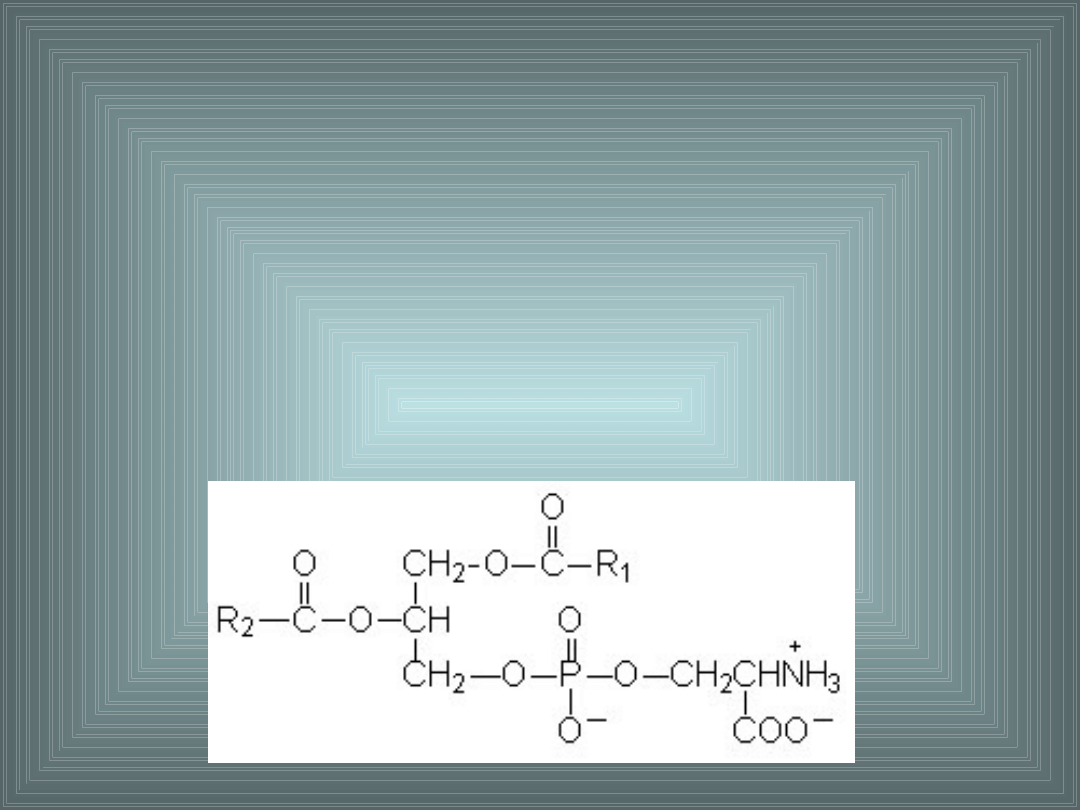
III. SKŁADNIKI LIPIDOWE BŁON
FOSFATYDYLOSERYNA:
- reszta fosforanowa zestryfikowana seryną
- występuje w warstwie P
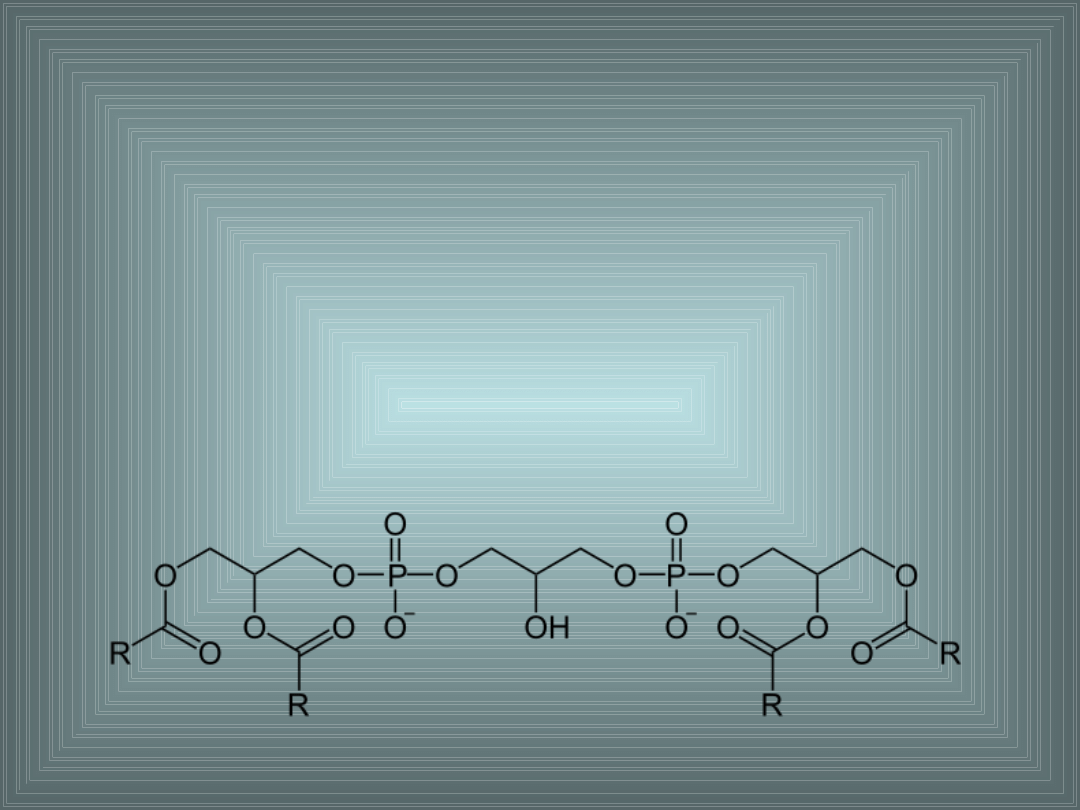
III. SKŁADNIKI LIPIDOWE BŁON
KARDIOLIPINA:
- difosfatydyloglicerol, ważny składnik wewnętrznej błony
mitochondrialnej i błon bakteryjnych
- odgrywa ważną rolę w prawidłowym funkcjonowaniu enzymu
oksydazy cytochromu C (COX)
III. SKŁADNIKI LIPIDOWE BŁON
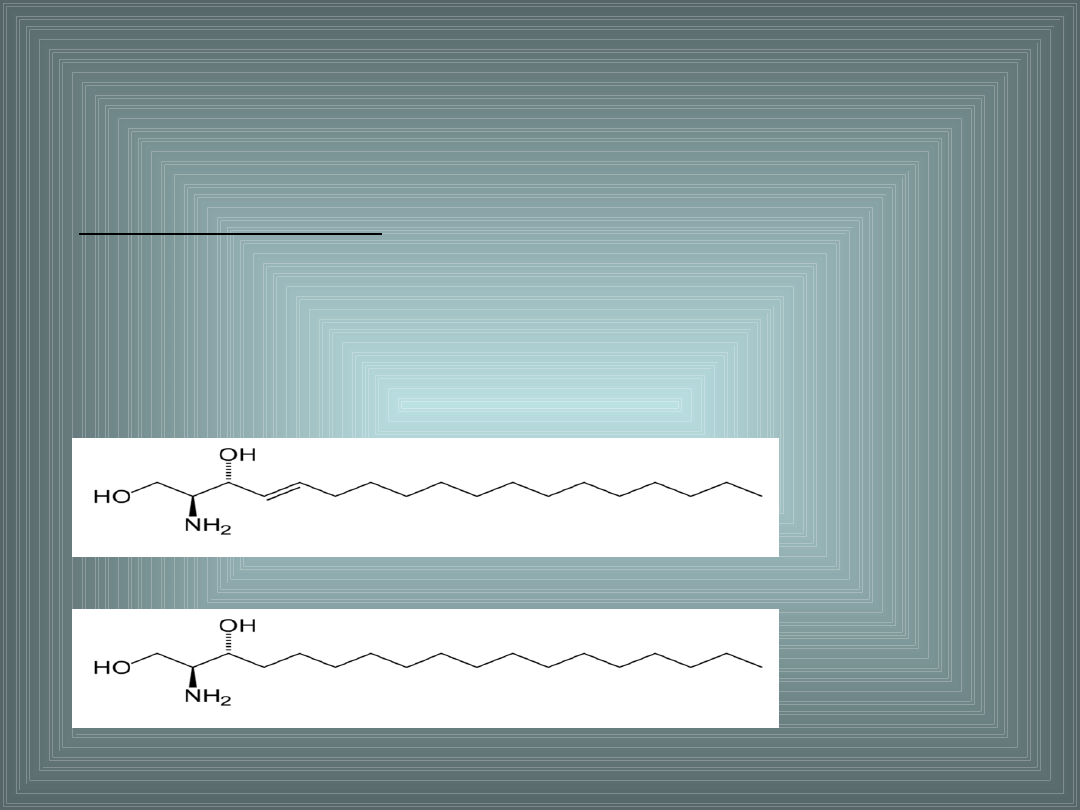
2. SFINGOLIPIDY.
Należą do fosfolipidów, zamiast glicerolu zawierają aminoalkohol:
sfingozynę lub dihydrosfingozynę.

III. SKŁADNIKI LIPIDOWE BŁON
Sfingolipidy są obecne w błonie komórek nerwowych, gdzie
uczestniczą w mechanizmie przekazywania sygnałów
elektrycznych i rozpoznawaniu molekularnym.
Chronią powierzchnię komórek przed niebezpiecznymi czynnikami
otoczenia przez tworzenie wytrzymałej mechanicznie i chemicznie
najbardziej zewnętrznej warstwy błon komórkowych
Są syntezowane w aparacie Golgiego, po czym są modyfikowane
w endozomach. Ich transport z aparatów Golgiego do błon
komórkowych odbywa się przez cytoplazmę w formie
monomerycznej. Sfingolipidy nie występują w błonach
wewnątrzkomórkowych otaczających mitochondrium i reticulum.

III. SKŁADNIKI LIPIDOWE BŁON
W wyniku wiązania kwasu tłuszczowego (najczęściej
palmitynowego, stearynowego, lignocerynowego i
nerwonowego) z gr. aminową sfingozyny lyb
dihydrisfingozyny wiązaniem amidowym, powstaje
ceramid, który jest prekursorem sfingomielin a także
glikolipidów, tzw. cerebrozydów.
Synteza sfingomieliny:
ceramid + fosfatydylocholina -> sfingomielina +1,2-diacyloglicerol
Synteza cerebrozydów:
ceramid + UDP-galaktoza -> cerebrozyd + UDP
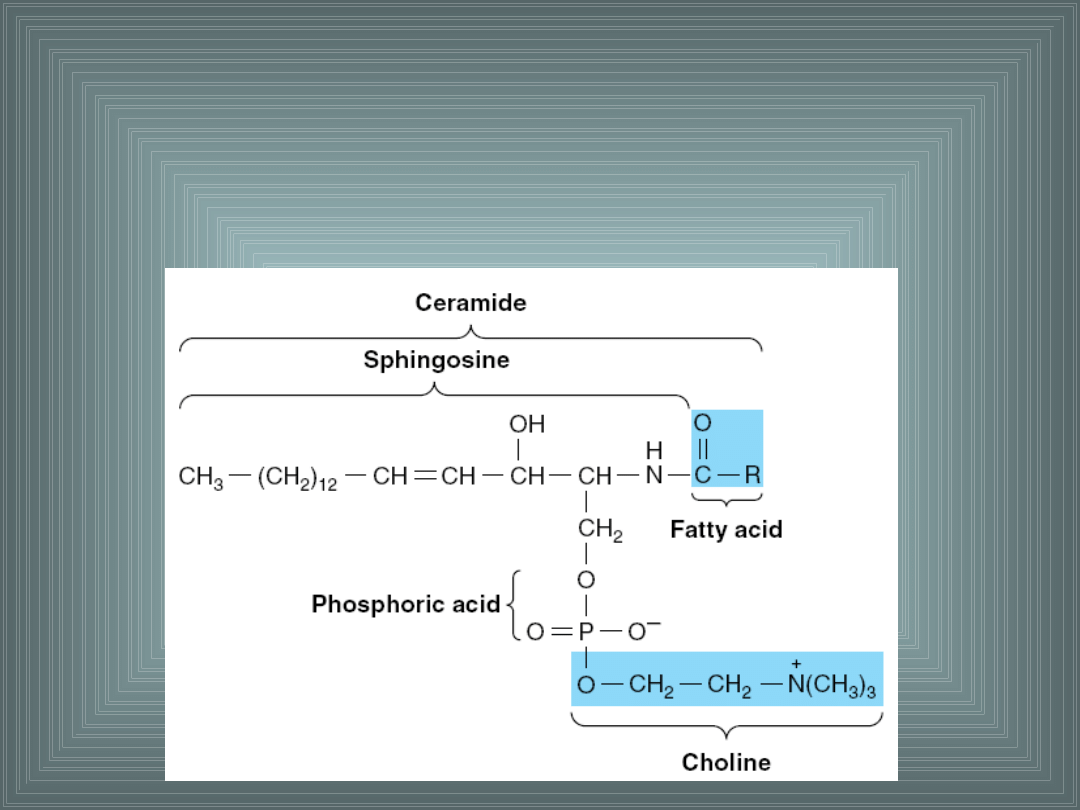
III. SKŁADNIKI LIPIDOWE BŁON
Sfingomielina –
występuje w dużych ilościach w mózgu i
tkance nerwowej. Posiada fosfocholinę połączoną z grupą
alkoholową.
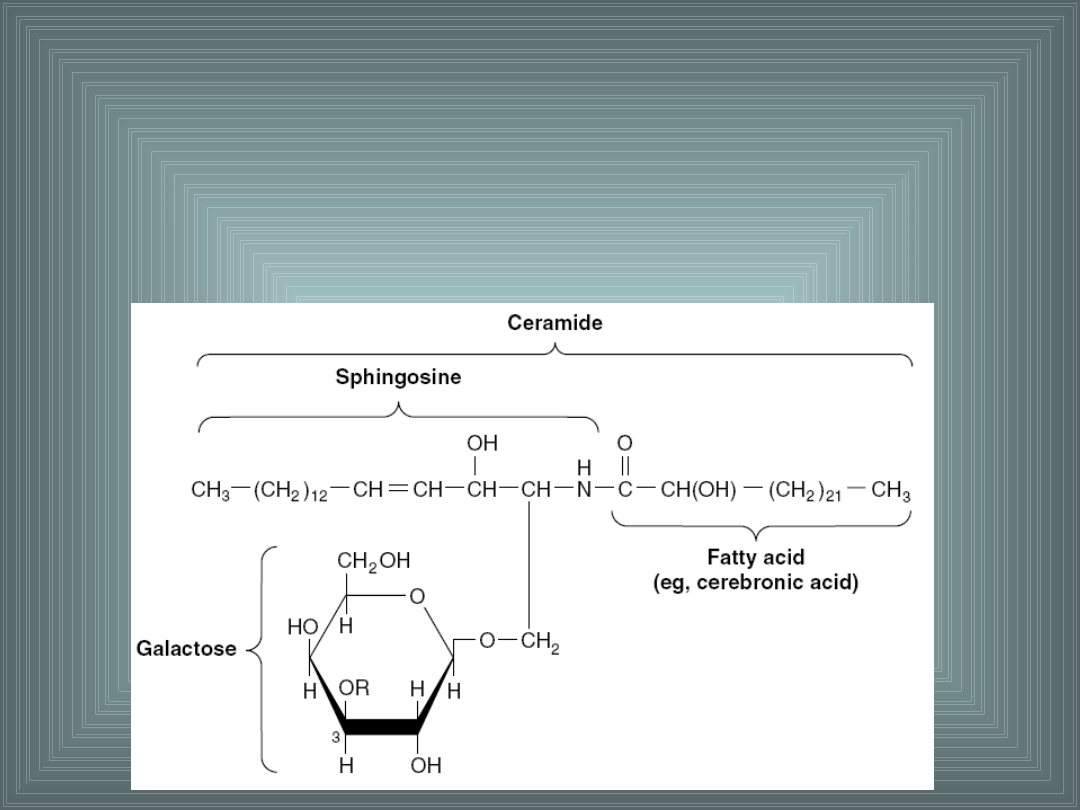
III. SKŁADNIKI LIPIDOWE BŁON
Cerebrozydy –
ich grupa alkoholowa jest połączona z
pojedynczą resztą heksozy (glukozą lub galaktozą). Występują
w dużej ilości np. w mózgu i włóknach nerwowych, a także we
krwi.
III. SKŁADNIKI LIPIDOWE BŁON
3. Cholesterol.

IV. Białka INTEGRALNE I PERYFERYCZNE
1) Białka integralne:
- są trwale związane z błoną
-
dzielimy je na białka transbłonowe i nieprzebijające błony
-
regulują transport substancji niezbędnych do życia komórki
-
tworzą trwałe kompleksy z lipidami oraz białkami
powierzchniowymi
-
funkcje: tworzą kanały, przenośniki, pompy, receptory, enzymy,
białka strukturalne, białka uczestniczące w przemianach
energetycznych w komórce, białka adhezyjne i wiele innych
-
występują dwa mechanizmy zakotwiczania ich w błonie:
a)
białka posiadające domeny zbudowane z aminokwasów o
resztach hydrofobowych o strukturze α-helisy, które zakotwiczają
je w dwuwarstwie
b)
białka posiadające kowalencyjnie dołączone grupy lipidowe

IV. Białka INTEGRALNE I PERYFERYCZNE
2) Białka peryferyjne:
- są związane z błona tylko po jednej stronie przez
oddziaływania elektrostatyczne pomiędzy różnoimiennymi
ładunkami powierzchniowymi
- ich usunięcie nie pozbawia błon zdolności do pełnienia ich
funkcji

V. PRZEPUSZCZALNOŚĆ BŁON
Błona komórkowa jest nieprzepuszczalna dla większości
cząsteczek hydrofilnych. Uniemożliwia „ucieczkę” metabolitów
z wnętrza komórki, ale też utrudnia wnikanie do komórki
niezbędnych składników i usuwanie zbędnych produktów
metabolizmu.
Dyfuzja – przenikanie substancji przez błonę zgodnie z gradientem
stężeń; dyfuzja jest tym szybsza im cząsteczka jest mniejsza,
bardziej hydrofobowa i elektrycznie obojętna.
Tlen cząsteczkowy i CO
2
szybko dyfundują.
Nienaładowane cząsteczki polarne dyfundują, gdy są dostatecznie
małe (woda, etanol – szybko, glicerol – wolniej, glukoza – nie
dyfunduje prawie wcale).

V. PRZEPUSZCZALNOŚĆ BŁON
Wszystkie jony i cząsteczki naładowane nie są przepuszczane
przez błonę, nawet jeśli są bardzo małe. Ich ładunek oraz
płaszcz wodny sprawiają, że nie mogę przenikać przez
hydrofobową warstwę błony.
Dlatego do transportu przez błonę cząsteczek naładowanych,
cukrów, aminokwasów, nukleotydów i innych metabolitów
niezbędne są obecne w błonie białka transportujące, tworzące
kanały i pompy.
Źródła: Biochemia Harpera, Murray; Seminaria z cytofizjologii, Zabel; Biochemia, Bańkowski
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
Wyszukiwarka
Podobne podstrony:
Transport przez błony biologiczne, Studia, I rok, Wykłady z biofizyki
Błony biologiczne
Biochemia wykład 12 Błony biologiczne
wyklad 3 Transport przez blony biologiczne 1
54 Wlasciwosci blony biologicznej i blon molekularnych
2 b Transport lekow przez blony biologiczne
Błony biologiczne architektura i funckja sackman 1
12 BIOCHEMIA blony biologiczne
14. Transport cząsteczek przez błony biologiczne, Studia, biologia
Blony biologiczne id 75238 Nieznany (2)
błony biologiczne
Blony biologiczne K, dydaktyka, biologia-praktyki, II gimnazjum
BŁONY BIOLOGICZNE
BIOCHEMIA 5 Blony biologiczne
Błony biologiczne i transport błonowy
33 TRANSPORT BIERNY I TRANSPORT AKTYWNY JONOW SODU I POTASU PRZEZ BLONY BIOLOGICZNE
Błony Biologiczne
więcej podobnych podstron