
Fizjologia mięśni,
skurcz mięśniowy

Molekularny mechanizm skurczu (sprzężenie
elektromechaniczne)
- pobudzenie motoneuronu
- pobudzenie dociera do jednostki motorycznej
- uwalnianie ACh do przestrzeni synaptycznej
- depolaryzacja sarkolemy, a także kanalików T i L pod wpływem ACh
- uwolnienie jonów Ca
2+
z pęcherzyków końcowych (kanaliki L)
- jony Ca
2+
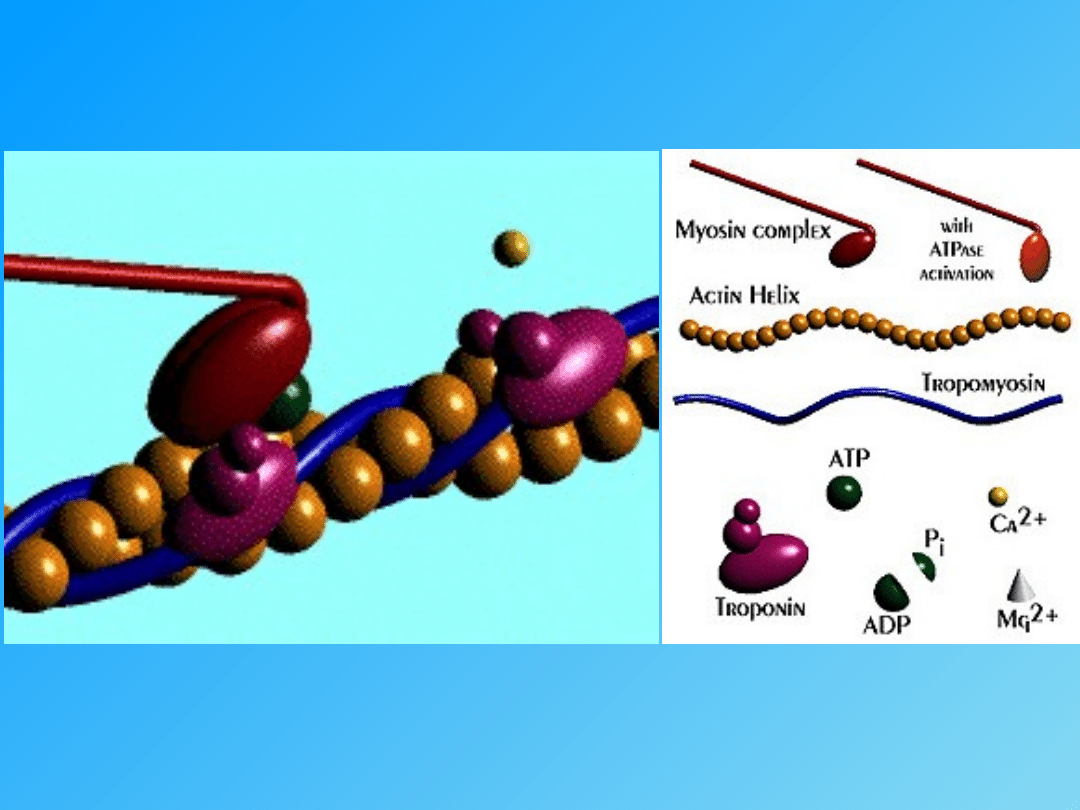
łączą się z troponiną C, co odblokowuje na nici aktyny miejsca
uchwytu mostków poprzecznych miozyny
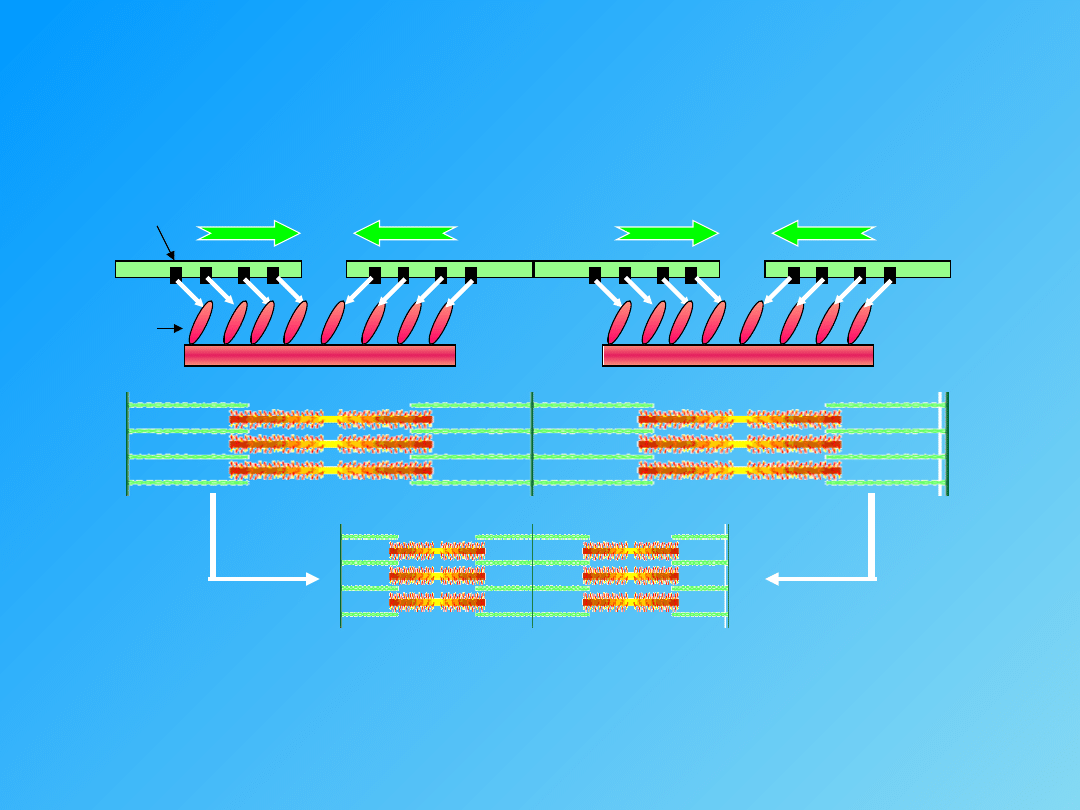
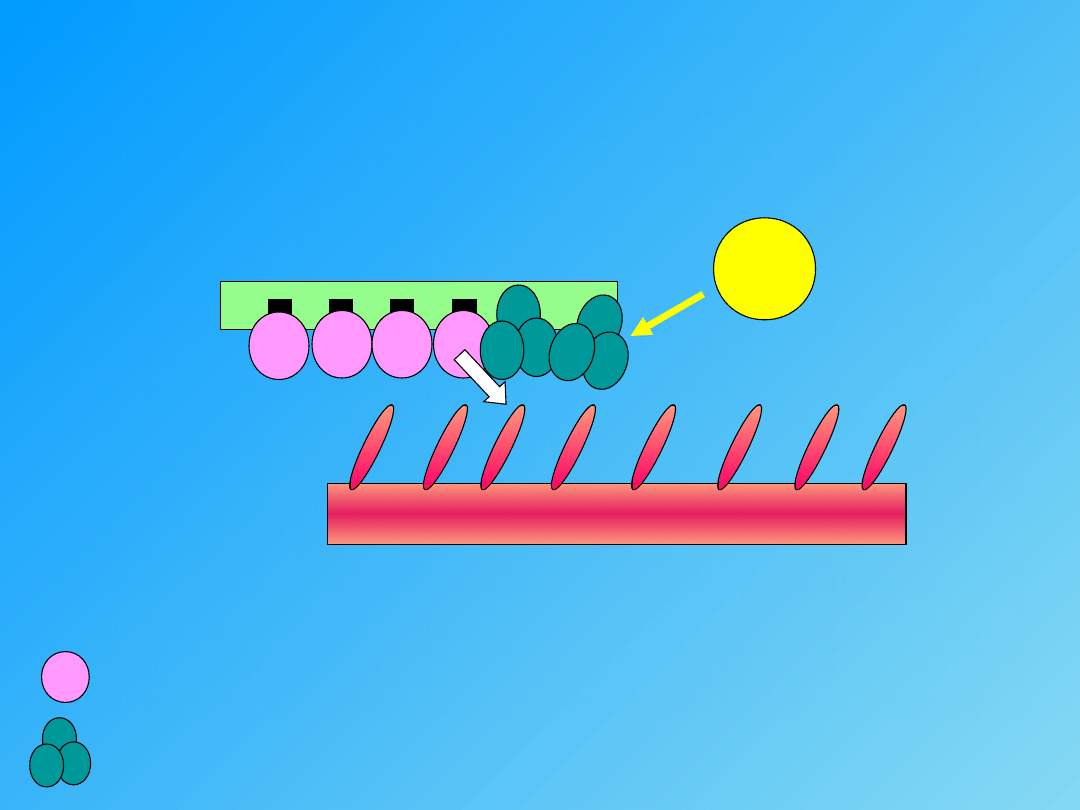
- przyłączenie mostków poprzecznych do aktyny, a także rozkład ATP
(energia)
- przesunięcie nitek aktyny między nitki
miozyny
SKURCZ
- wychwyt jonów Ca
2+
przez kanaliki podłużne L i transport ich do
pęcherzyków końcowych
A
M
A
mostki poprzeczne
miejsce aktywne
T
m
T
m
T
m
T
m
Ca
+
+
T
I
C
T
I
C
T
m
T
I
C
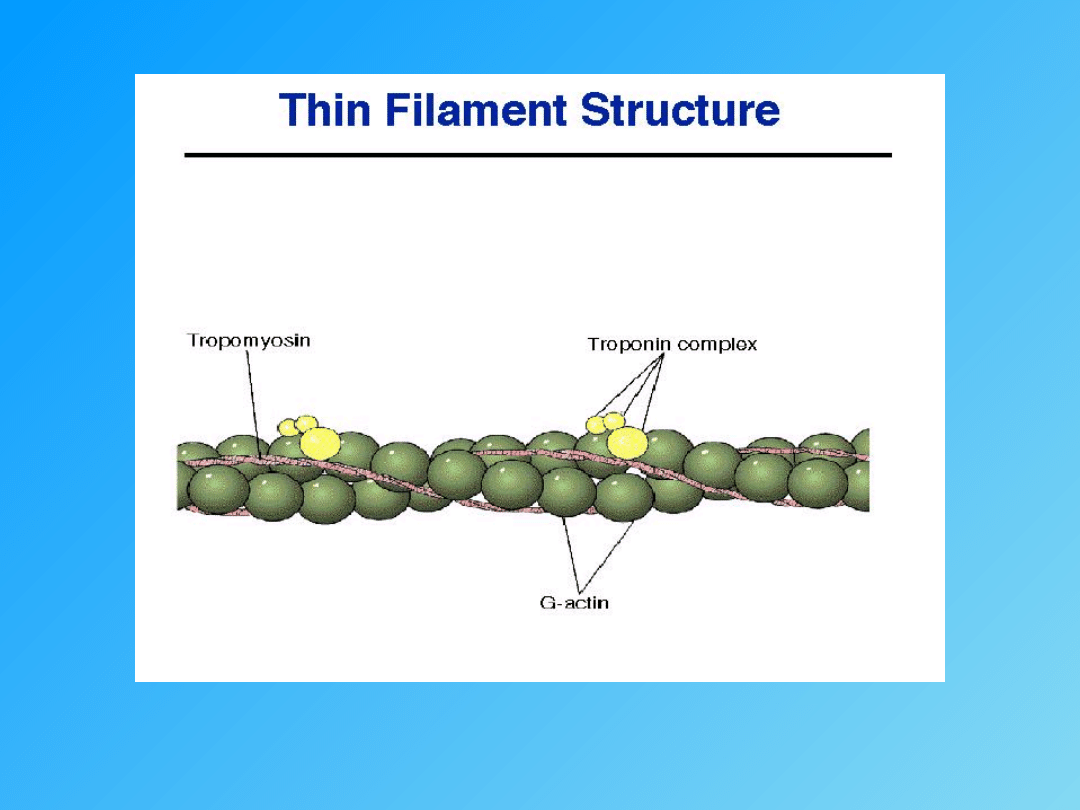
- tropomiozyna
- troponina

Źródła tlenu w mięśniach
1. Ciśnienie parcjalne O
2
w komórce.
2. Utlenowana mioglobina.
• do wytwarzania energii (ATP) w procesach tlenowych (glikoliza tlenowa) mięśnie nie
obciążone potrzebują 250 ml O
2
/min.
• w czasie ciężkiej pracy mięśni wartość ta wzrasta do 4 dm
3
O
2
/min
• krew jest w stanie dostarczyć mięśniom 3 dm
3
O
2
/min
• zaczyna się beztlenowa synteza ATP (glikoliza beztlenowa) w warunkach długu
tlenowego
Dług tlenowy (niedobór tlenu) – zaciągany od rozpoczęcia pracy do przystosowania się
ukł. krążenia i ukł. oddechowego do większego zapotrzebowania na O
2
. Oddawany w
po zakończeniu pracy mięśni – jeszcze trwa wzmożona praca serca i ukł. oddechowego.

Energia skurczu pochodzi z rozkładu ATP
ATP → ADP + Pi + 7,8 kcal/mol
ADP → AMP + Pi + 7,8 kcal/mol
Resynteza ATP:
1. fosfokreatyna
hydroliza
kreatyna + Pi + E
2. glikogen → glukoza
r. beztlen.
2 ATP + kw. mlekowy
r. tlenowy.
38 ATP
Wydajność energetyczna mięśnia:
• 20 – 25% → praca
• 75 – 80% → ciepło
Równanie Hilla:
Energia całkowita = En. aktywacji + En. skurczu + En. pracy + ciepło
KREW
MIĘŚNIE
WĄTROBA
glukoza
kwas mlekowy
w. beztlen.
kwas mlekowy
20% + O
2
80%
E
glikogen
glukoza
Cykl Coricha (Coriego)

Reakcje energetyczne zachodzące podczas skurczu mięśnia
1. Rozpad ATP (starcza na 10 skurczów).
2. Rozpad fosfokreatyny dla resyntezy ATP (starcza na 50 skurczów = 10-20 sek.).
3. Beztlenowy rozpad glikogenu (glikoliza) dla resyntezy ATP i fosfokreatyny
(rozpoczyna się po 30 sek. od rozpoczęcia pracy mięśnia).
4. Fosforylacja tlenowa glukozy i wolnych kwasów tłuszczowych (głównie okres
odnowy i wypoczynku).

Ciepło mięśni
1. Ciepło spoczynkowe – związane z podstawowymi procesami metabolicznymi
2. Ciepło początkowe
ciepło aktywacji – tworzy się przez cały czas skurczu;
„aktywacja” miozyny
ciepło skracania
ciepło rozkurczu
3. Ciepło wypoczynku – resynteza ATP

Znużenie mięśni
PRZYCZYNY ZMĘCZENIA:
1. Wyczerpanie się zapasów energetycznych.
2. Zatrucie produktami metabolizmu (kwas mlekowy).
3. Wpływ ośrodków ruchowych.
• Najpierw męczy się komórka czuciowa (pobudzić można inną kom. i uzyskać skurcz),
• potem męczy się płytka motoryczna (można uzyskać jeszcze skurcz działając
bezpośrednio na mięsień),
• na końcu męczy się mięsień.

Rodzaj pracy, a zmęczenie mięśni
1. Praca intensywna – organizm nie nadąża dostarczać O
2
tworzy się bardzo duży dług
tlenowy, działający przede wszystkim na neurony, które sygnalizują zmęczenie.
2. Praca długotrwała – jest to praca przy uregulowanym dopływie O
2
; zaczynają się
wyczerpywać zapasy energetyczne.
3. Praca statyczna – wzrasta napięcie mięsni, stały ucisk na naczynia krwionośne
zmniejsza odżywianie mięśnia.

Cechy zmęczenia
- dłuższa latencja
- zmniejszona amplituda skurczu
- dłuższy czas rozkurczu
- zmniejszona pobudliwość
- zmniejszona maksymalna siła skurczu
- zmniejszone zapasy energetyczne
czas
siła skurczu

Elastyczność mięśnia
MIĘSIEN WYKAZUJE:
1. Fizyczne cechy ciał martwych:
a) Zdolność do reagowania odkształceniem na działającą siłę (ale niezgodnie z
prawem Hooka, tzn. równy ciężar dodany nie powoduje jednakowego przyrostu
długości).
b) Zdolność do powrotu do pierwotnej formy po odjęciu siły rozciągającej.
2. Szczególne właściwości elastyczne – pod wpływem siły rozciągającej lub
pobudzenia nerwowego może zmieniać swoje napięcie nie zmieniając długości.
mięśnie
ciała martwe
długość ∆ l
obciążenie P

% max. siły skurczu
100
długość mięśnia
L max
Mięsień rozkurczony kurczy się słabo. Wstępne rozciągnięcie mięśnia zwiększa
siłę skurczu (najkorzystniejsze ułożenie włókien aktyny i miozyny względem
siebie).
L max – najkrótsza długość mięśnia rozwijającego
max. siłę (ok. 20 % rozciągnięcia).
Zależność między długością, a siłą skurczu mięśnia
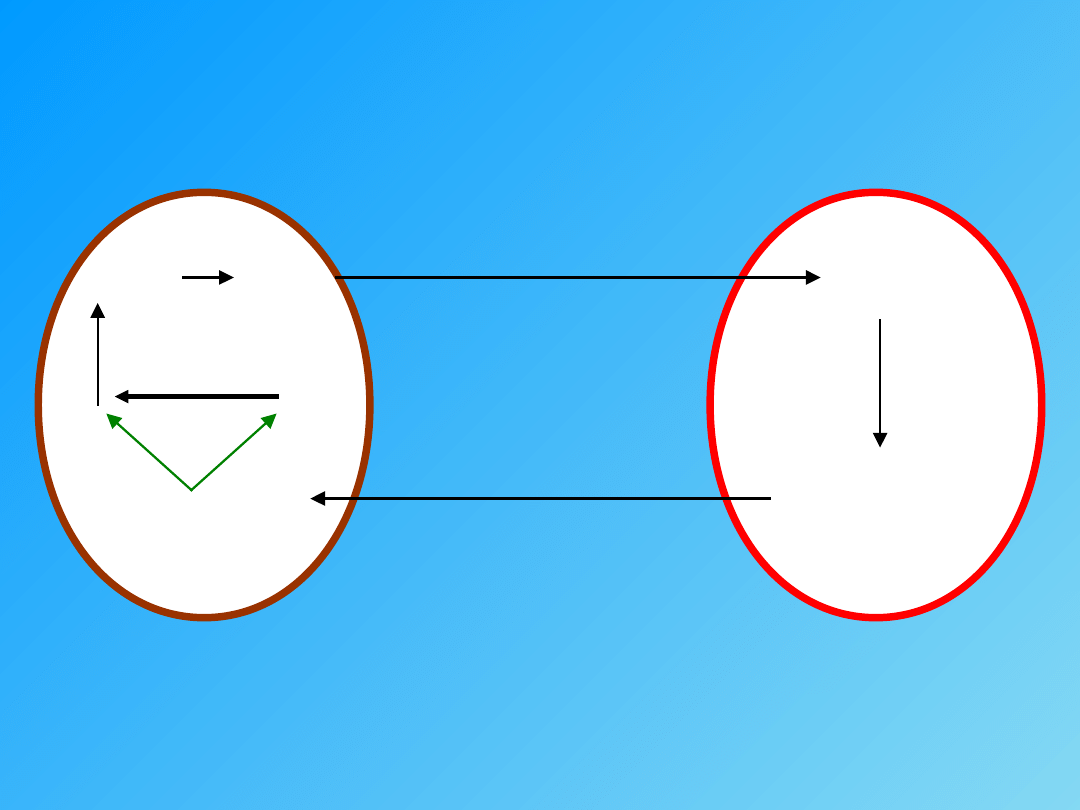
Rodzaje skurczów
K
E
SKURCZ IZOTONICZNY
(takie samo napięcie, zmiana długości) Skurcz bez
obciążenia.
E
K
V = max
∆ L ≠ 0
SKURCZ IZOMETRYCZNY
(taka sama długość, zmiana napięcia) Obciążenie
przekraczające max. siłę mięśnia.
K
E
1 t
1 t
E
K
F = 0
W = F x L = 0
V = 0
∆ L = 0
F = max
W = 0
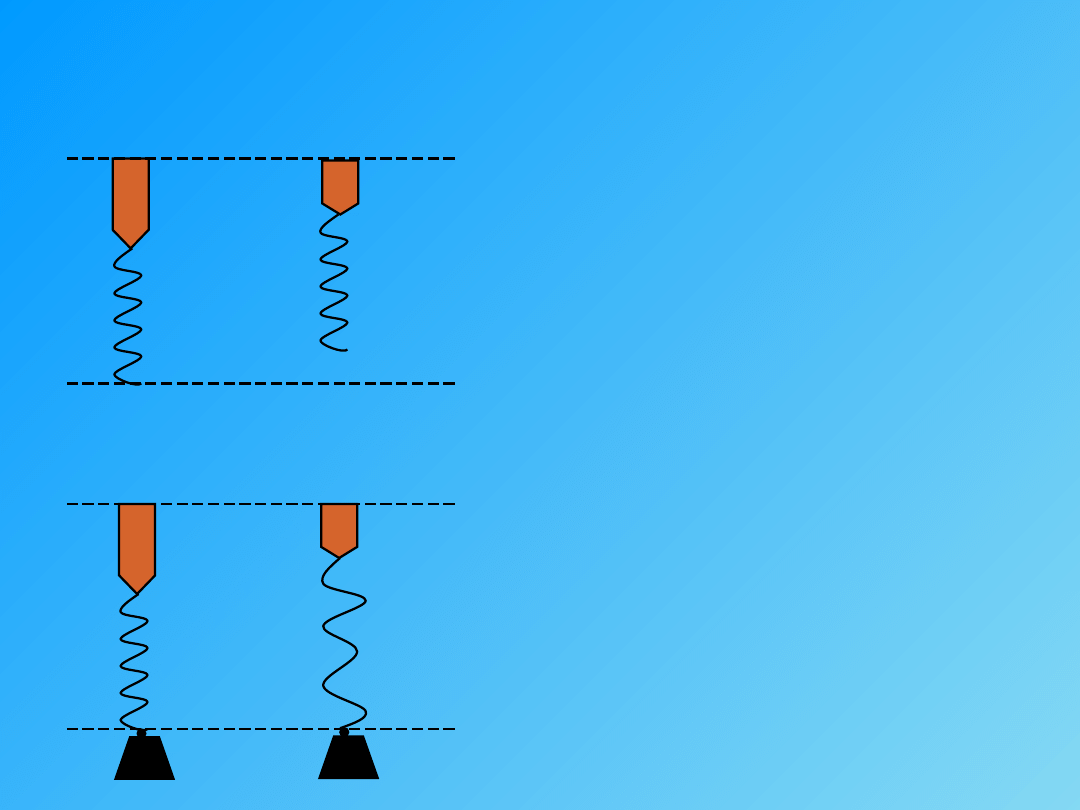
SKURCZ AUKSOTONICZNY
(mieszany, fizjologiczny, efektywny). Skurcz przy
pewnym obciążeniu.
K
E
E
K
K
E
V ≠ 0
∆ L ≠ 0
F ≠ 0
W ≠ 0
Relaksacja napięcia (opóźnienie sił elastycznych) – jest to opóźnienie z jakim mięsień
rozwija siłę (elastyczną) skracając się i pokonując opór (obciążenie).
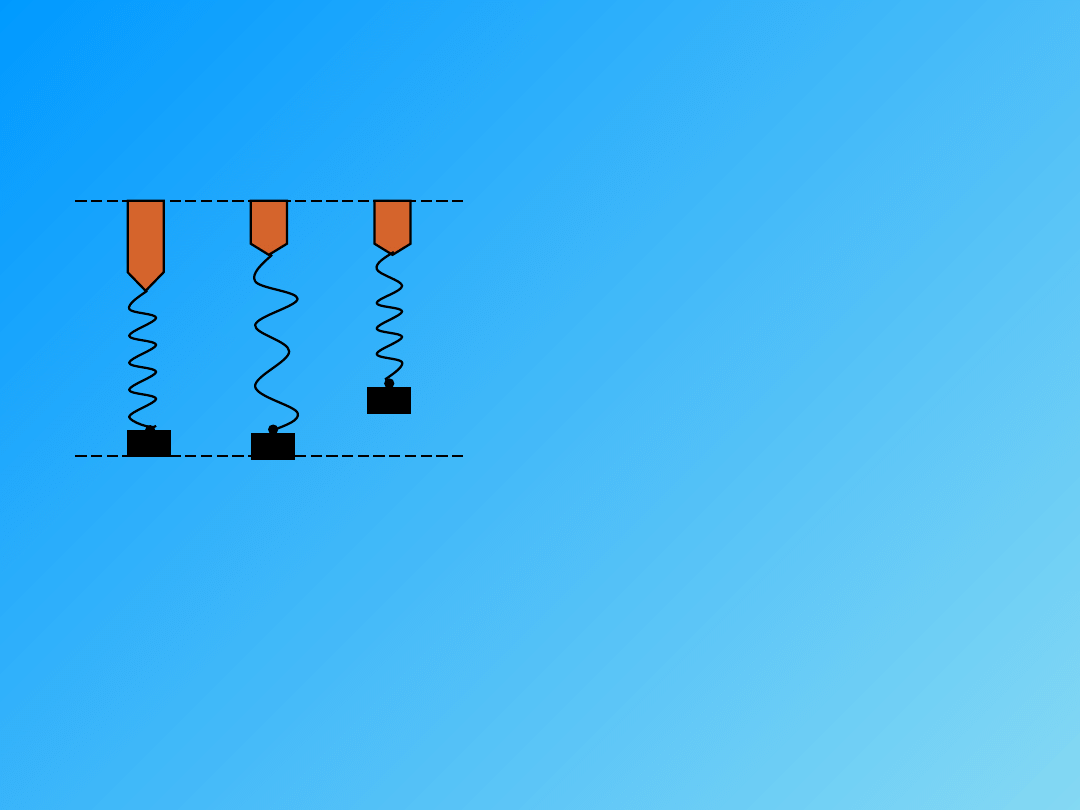
Zależność między szybkością skurczu, a obciążeniem mięśnia
obciążenie
szybkość
skurczu
Mięsień kurczy się szybciej, im mniej jest obciążony.
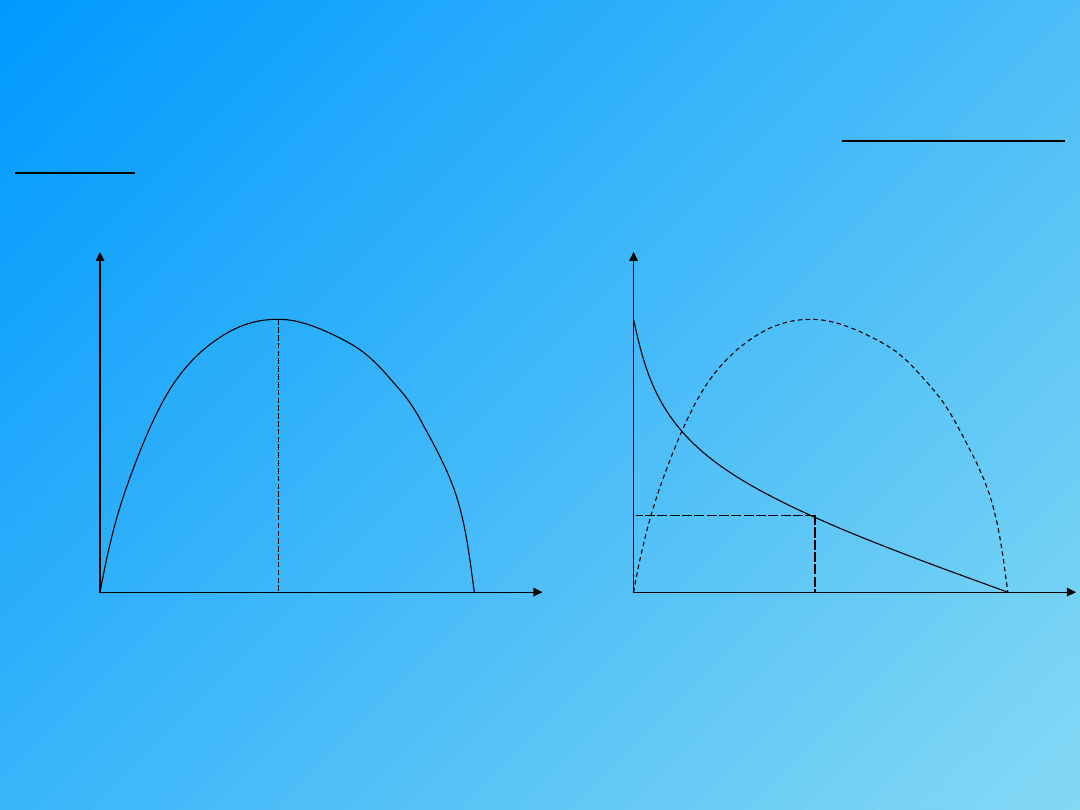
Zależność między wartością obciążenia mięśnia, która pozwala się skracać, a
wartością która zapewnia rozwinięcie pewnej siły to prawo średnich
obciążeń.
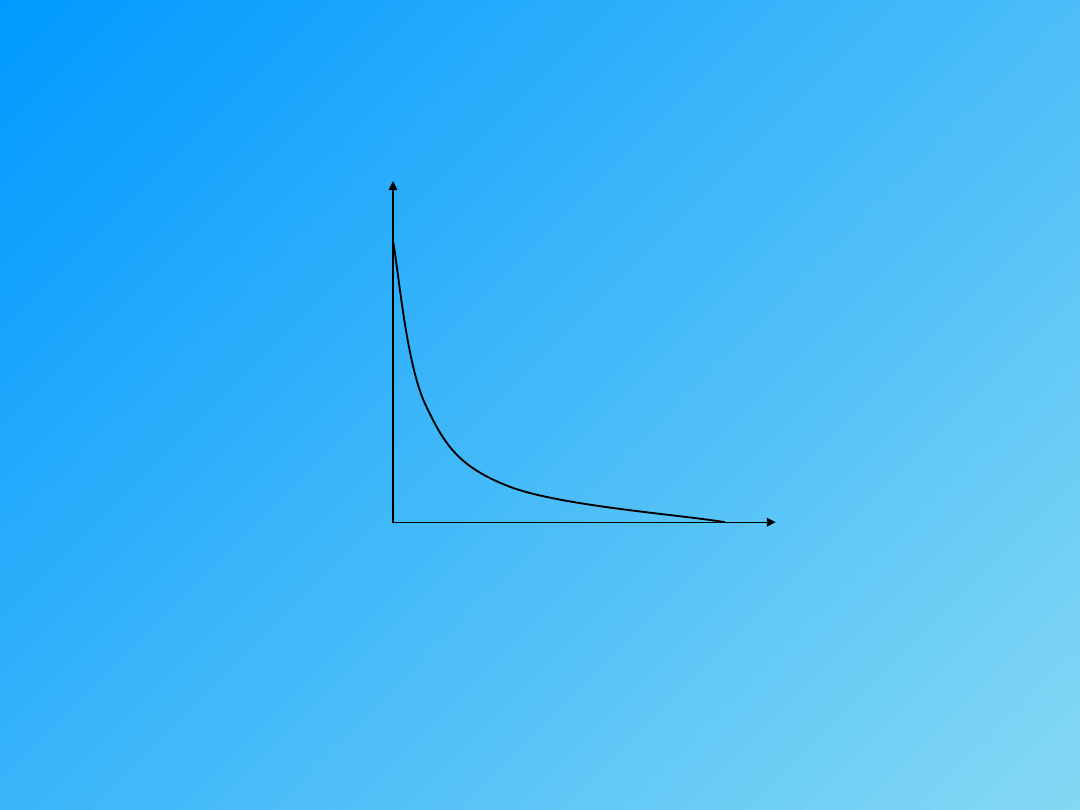
P (obciążenie)
W (% max. pracy)
½ P
Maksymalna praca wykonywana jest przy ½ obciążenia maksymalnego
P (obciążenie)
½ P
W
V max.
1/3 V
i
prędkości
skurczu 1/3 V max.

Cechy mięśni gładkich
- „pływający” potencjał spoczynkowy (od - 25 do -75 mV)
- zmienna amplituda potencjałów czynnościowych
- aktywacja pod wpływem pobudzenia nerwowego (gł. naczynia), ale też pobudzenie 2
sąsiednich komórek mięśnia przez niskooporowe złącza
- powolność skurczu i rozkurczu (toniczność skurczu)
- silny automatyzm (kurczą się bez impulsów z OUN lub mają własne ośrodki
automatyczne np. skurcz przez rozciągnięcie - ruchy robaczkowe jelit wywołane przez
przesuwający się pokarm)
- wykazują obecność podwójnego unerwienia – sympatycznego i parasympatycznego
(napędu i hamulca)
- silnie zaznaczona elastyczność typu plastycznego – duża zmiana długości, a mała
napięcia (pęcherz moczowy)
- wrażliwe na bodźce chemiczne, szczególnie na aminy biogenne
- nieuporządkowany układ miofibrylli – brak poprzecznego prążkowania, ale za to siła
skurczu nie słabnie nawet przy znacznym rozciągnięciu mięśnia
- w porównaniu z mięśniami szkieletowymi tylko 10 % białek kurczliwych, mniej ATP i
fosfokreatyny – stąd słabość skurczu
- brak zbiorników dla Ca
2+
- wykorzystują tylko Ca
2+
zewnątrzkomórkowe (przy
pobudzeniu tworzy się dokomórkowy prąd wapniowy).
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
Wyszukiwarka
Podobne podstrony:
FIZJOLOGIA MIĘŚNIA SERCOWEGO – AKTYWNOŚĆ SKURCZOWA MIĘŚNIA SERCOWEGO - instrukcja, Medycyna i Zdrowi
krótkie wykłady kraba seby z fizjologi Mięśnie, skurcz mięśnia
fizjologia-mięsnie ręki, AWF KATOWICE, FIZJOLOGIA
Wybrane zagadnienia z fizjologii mięśni i fizyczne właściwości mięśni, Biomechanika
fizjologia mięśnie
WŁAŚCIWOŚCI FIZJOLOGICZNE MIĘŚNIA SERCA, ZDROWIE, Kardiologia
Fizjologia mięśni 2
Fizjologia mięśni szkieletowych - laborki, ZDROWIE, MEDYCYNA, Anatomia i fizjologia człowieka, Anato
Fizjologia mięśni - fizjologia zwierząt. 5fantastic.pl , Ćwiczenia
fizjologia miesni
39 BUDOWA I FIZJOLOGIA MIĘŚNI SZKIELETOWYCH
fizjo mail, Fizjologia mieśni gładkich, B
FIZJOLOGIA CZŁOWIEKA ĆWICZENIA, Fizjologia mini poprzecznie prkowanych, Fizjologia mięśni poprzeczni
FIZJOLOGIA - mięśnie cd, Wykłady, FIZJOLOGIA
fizjologia miesni poprzecznie prazkowanych, Studia, Neurobiologia
FIZJOLOGIA MIĘŚNI
więcej podobnych podstron