
CYKL KOMÓRKOWY
Nagroda Nobla
w dziedzinie fizjologii i medycyny
rok 2001
„for the key regulators of the cell cycle”
Paul M. Nurse (Imperial Cancer Research Fund, London UK)
R.Timothy Hunt (Imperial Cancer Research Fund, London UK)
Leland Hartwell (University of Washington, Seattle, USA)
Nagroda Nobla
w dziedzinie chemii
rok 2004
„for the discovery of ubiquitine –mediated
protein degradation”
Aaron Ciechanover (Israel Institute of Technology, Haifa)
Avram Hershko (Israel Institute of Technology, Haifa)
Irwin Rose (University of California, USA)

CYKL
ŻYCIOWY
CYKL KOMÓRKOWY
Jest to ściśle określona sekwencja etapów,
przez które przechodzi komórka od chwili
powstania do zakończenia podziału
CYKL MITOTYCZNY
Jest to zespół procesów molekularnych i
zmian strukturalnych w komórce
rodzicielskiej, zachodzących w ściśle
określonej kolejności i kierunku, których
skutkiem jest precyzyjne podwojenie i
rozdział materiału genetycznego pomiędzy
dwie komórki potomne
Cykl mitotyczny składa się z faz:
G1, S i G2 – interfazy
oraz z fazy
M -
mitozy
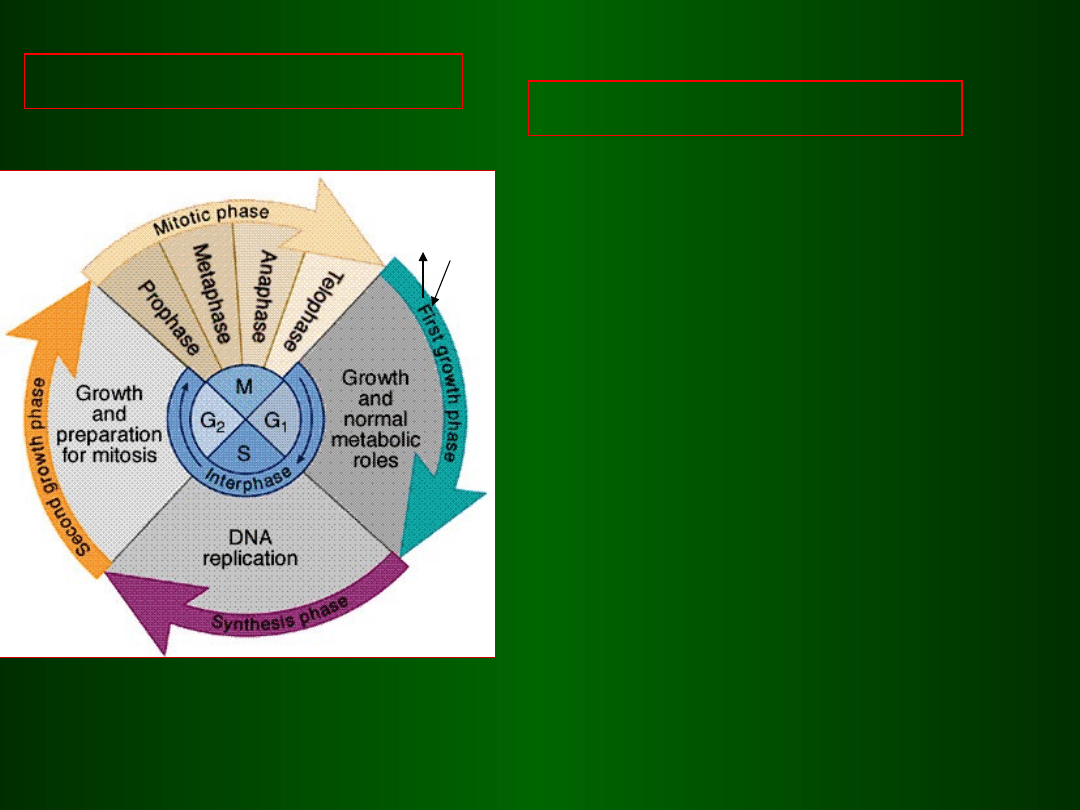
CYKL MITOTYCZNY
Interfaza=G1+S+G2
G1 faza wzrostu i intensywnego
metabolizmu komórkowego,
przedreplikacyjna naprawa DNA
S faza jednorazowej replikacji DNA
i syntezy białek histonowych
G2 faza wzrostu i syntezy białek
wrzeciona podziałowego,
naprawa zreplikowanego DNA
M faza mitozy
G
0
2C DNA (G1) 4C DNA
(G2)

• Komórki namnażające się, odnawialne, o charakterze
embrionalnym,
merystematyczne, macierzyste (G1)
• Komórki spoczynkowe, zdolne do ponownych podziałów (G0)
• Komórki zróżnicowane, ale zdolne do odróżnicowania i
ponownych podziałów
• Komórki zróżnicowane ostatecznie, niezdolne do ponownych
podziałów
• Komórki eliminowane na drodze apoptozy (zwierzęce) lub
programowanej
śmierci komórkowej (roślinne)
Kategorie losów komórek w organizmach
roślin i zwierząt
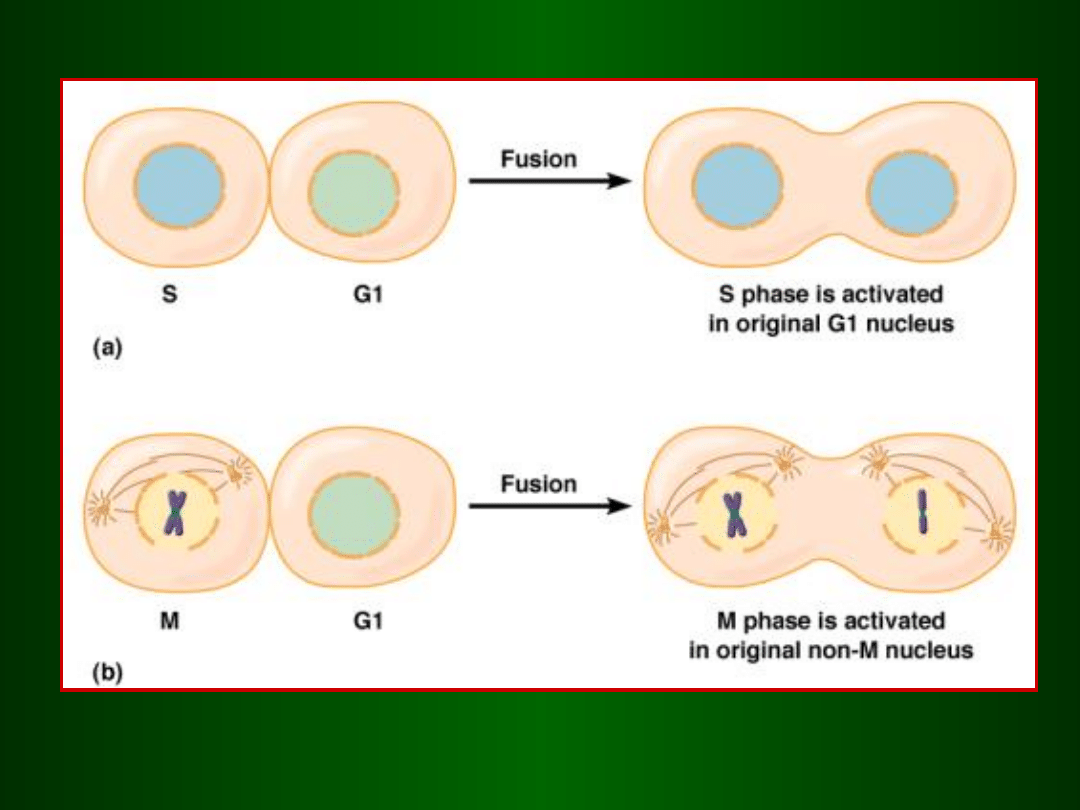
Dowód na to, że istnieją czynniki, które przełączają fazy cyklu
Fuzja komórek He-La

Do punktów kontrolnych należy regulacja i kontrola
przejścia komórki do
kolejnych faz cyklu dopiero po pomyślnym ukończeniu fazy
poprzedzającej
Punkt kontrolny późnej fazy
G1
kontroluje przejście
G1/S ,
zwany
START
lub
PUNKT RESTRYKCYJNY
Decyduje on o wejściu komórki do cyklu mitotycznego
Punkt kontrolny późnej fazy
G2
kontroluje przejście
G2/M
Decyduje on o wejściu komórki do fazy
Mitozy
Punkt kontrolny wrzeciona podziałowego kontroluje
przejście
Metafaza/Anafaza
Decyduje on o precyzyjnym rozdziale wszystkich chromatyd
siostrzanych
(chromosomów potomnych) do dwóch przeciwległych
biegunów komórki
PUNKTY KONTROLNE (ZWROTNE) CYKLU
MITOTYCZNEGO(CHECKPOINTS)

Przejście przez punkty kontrolne jest
realizowane dzięki aktywacji tzw. przełączników
molekularnych
Są to dimery złożone z:
KINAZ ZALEŻNYCH OD CYKLIN
(CDK
= Cyclin Dependent Kinases)
i
CYKLIN
Cykliny regulują aktywność enzymatyczną kinaz
– bez połączenia z cyklinami kinazy nie działają
Przełączniki molekularne w poszczególnych
fazach cyklu są kompleksami różnych
CDK
i
różnych
CYKLIN
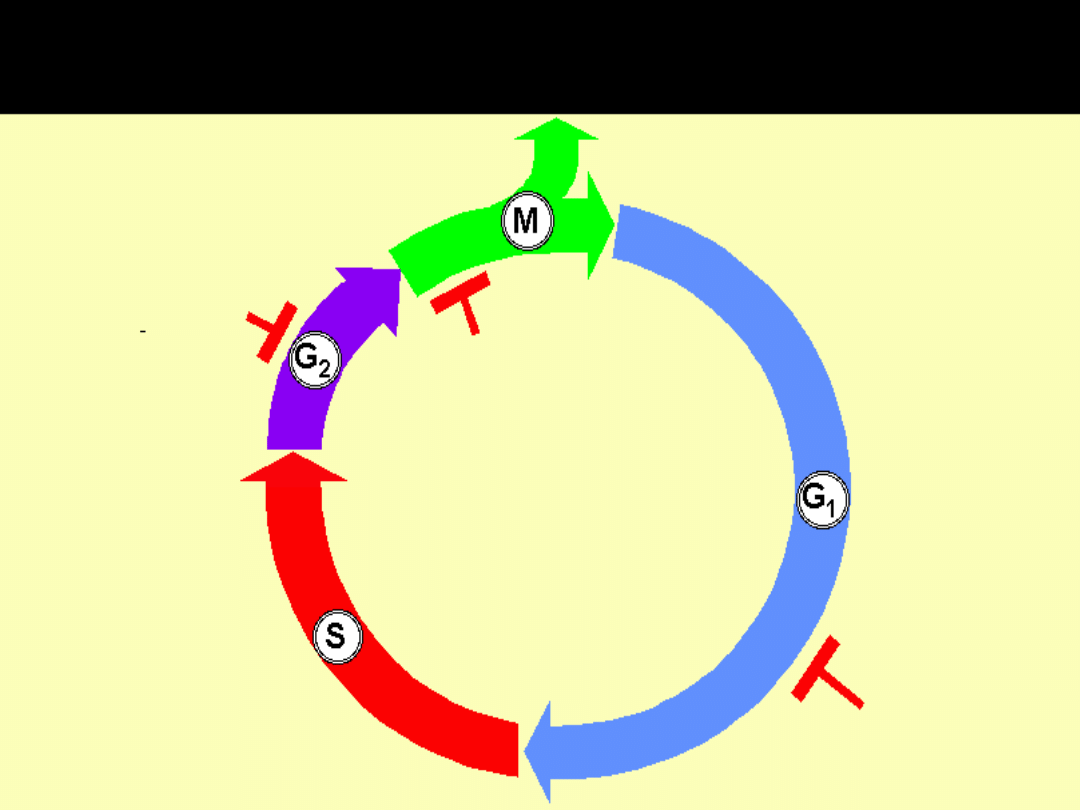
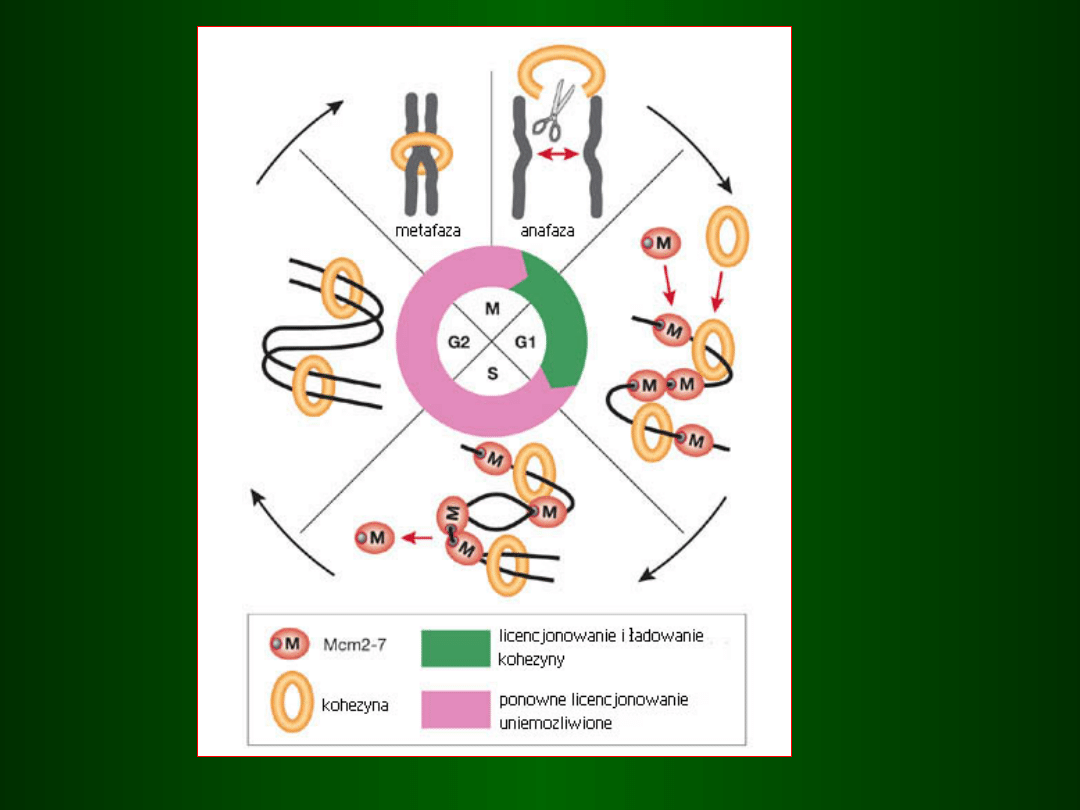
Punkty kontrolne i sprzężenie zwrotne
w regulacji cyklu komórkowego
mitoza
„niezmontowane” wrzeciono
rozmiar komórki,
poziom metabolizmu,
uszkodzone DNA
synteza DNA
niezreplikowane lub
uszkodzone DNA
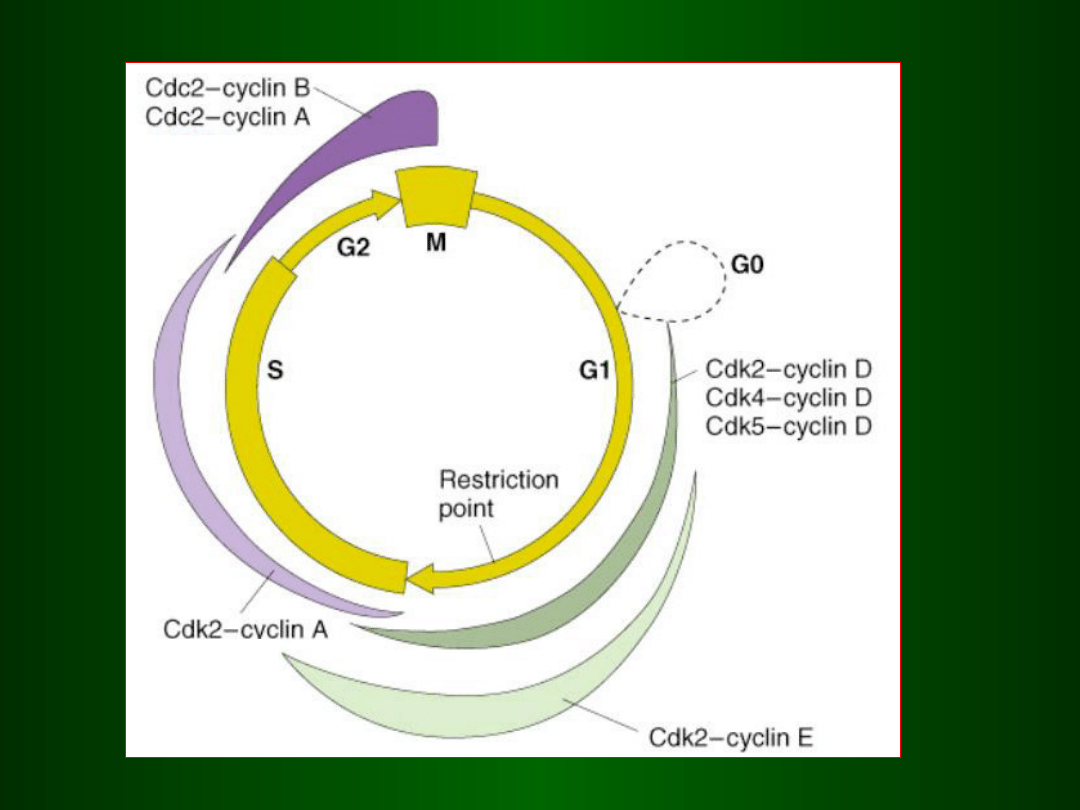

uruchamia następujące po sobie fazy cyklu
mitotycznego,
działając w punktach kontrolnych np. pomiędzy:
G1 / S
tzw.
SPF
(S-phase Promoting Factor)
G2 / M
tzw.
MPF
(M-phase Promoting Factor)
Jest dimerem składającym się z dwóch białek:
K I N A Z Y CYKLINOZALEŻNEJ
i
C Y K L I N Y
Dla każdej fazy cyklu charakterystyczne są różne kinazy
i różne cykliny
Poziom kinaz podczas cyklu mitotycznego jest stały,
a cyklin zmienia się cyklicznie
PRZEŁĄCZNIK
MOLEKULARNY
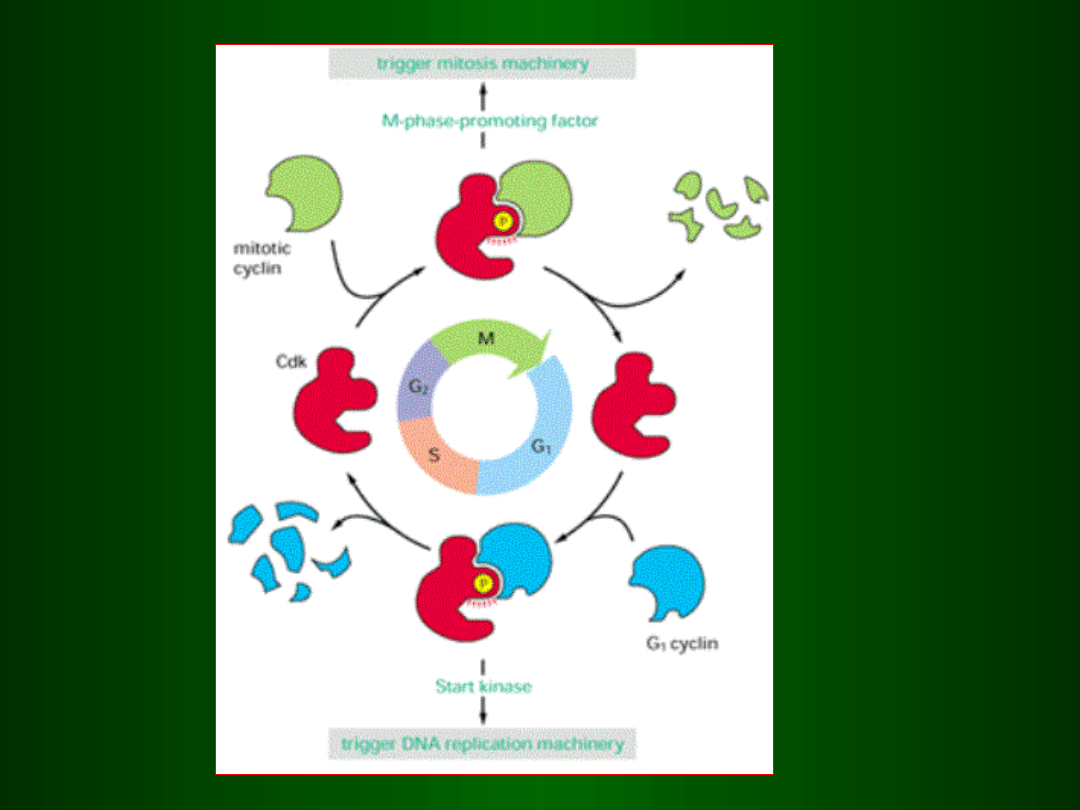
MPF
SPF

Faza G1
•
Wzrost komórki
• Synteza cyklin D i E
• Uzyskiwanie „licencji” chromatyny na replikację (ładowanie
na DNA
kompleksów OR i RLF)
• Kontrola i naprawa uszkodzeń DNA (białko p53 i p21)
• Fosforylacja białka RB przez SPF (punkt restrykcyjny)
Faza S
•
Jednorazowa replikacja DNA
• Synteza białek histonowych
• Ubikwitynacja i degradacja w proteasomach cyklin
fazy G1
• Usuwanie kompleksów RLF po zakończeniu syntezy
DNA w
replikonie
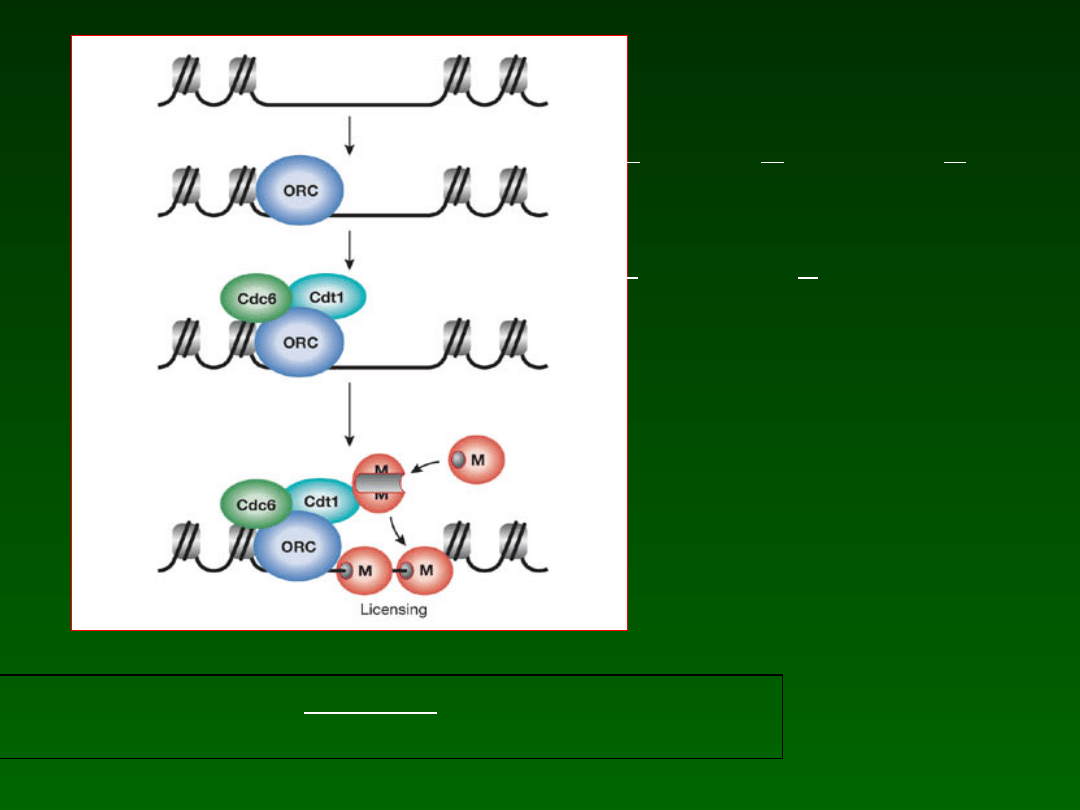
Faza G1
Powstawanie i ładowanie kompleksów OR i RLF
ORC
Origin of Replication Complex
RLF
Replication Licencing Factor
(Cdc6,Cdt1, M)
OR obecne podczas całego cyklu
RLF ładowane podczas G1, a
usuwane podczas S, po replikacji
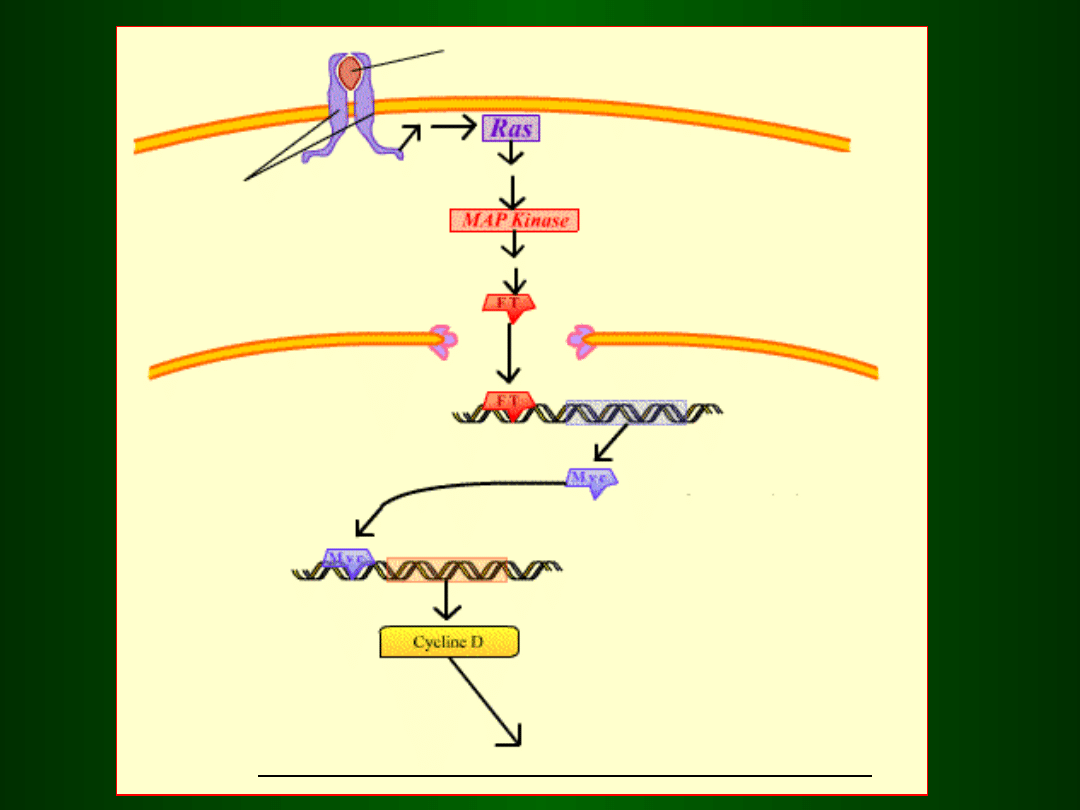
czynnik wzrostowy
receptor (dimer)
środowisko zewnętrzne
cytoplazma
jądr
o
aktywacja czynników transkrypcyjnych
czynnik transkrypcyjny
gen cykliny D
wejście do fazy G1 i jej kontynuacja
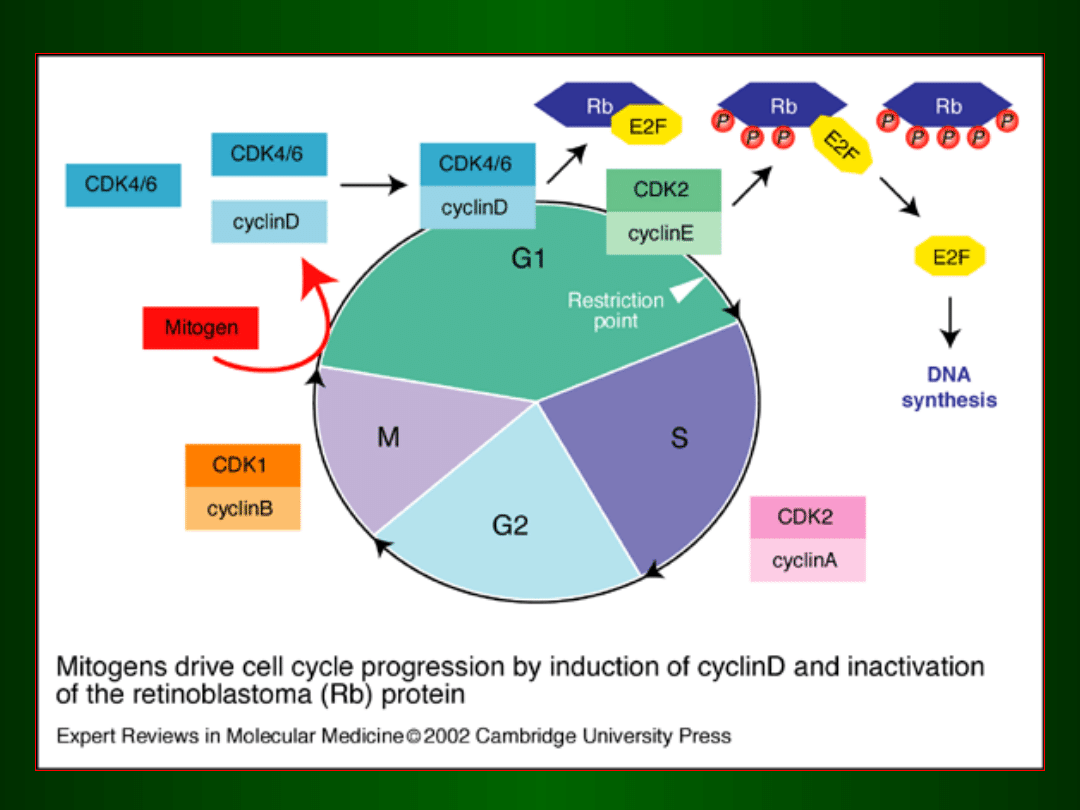
ekspresja
genów
fazy S
Działanie CDK 4/6 / Cykliny D na
białko Rb
Inaktywacja czynnika transkrypcyjnego
E2F poprzez związanie go z białkiem Rb
Uwolnie
nie
czynnik
a E2F
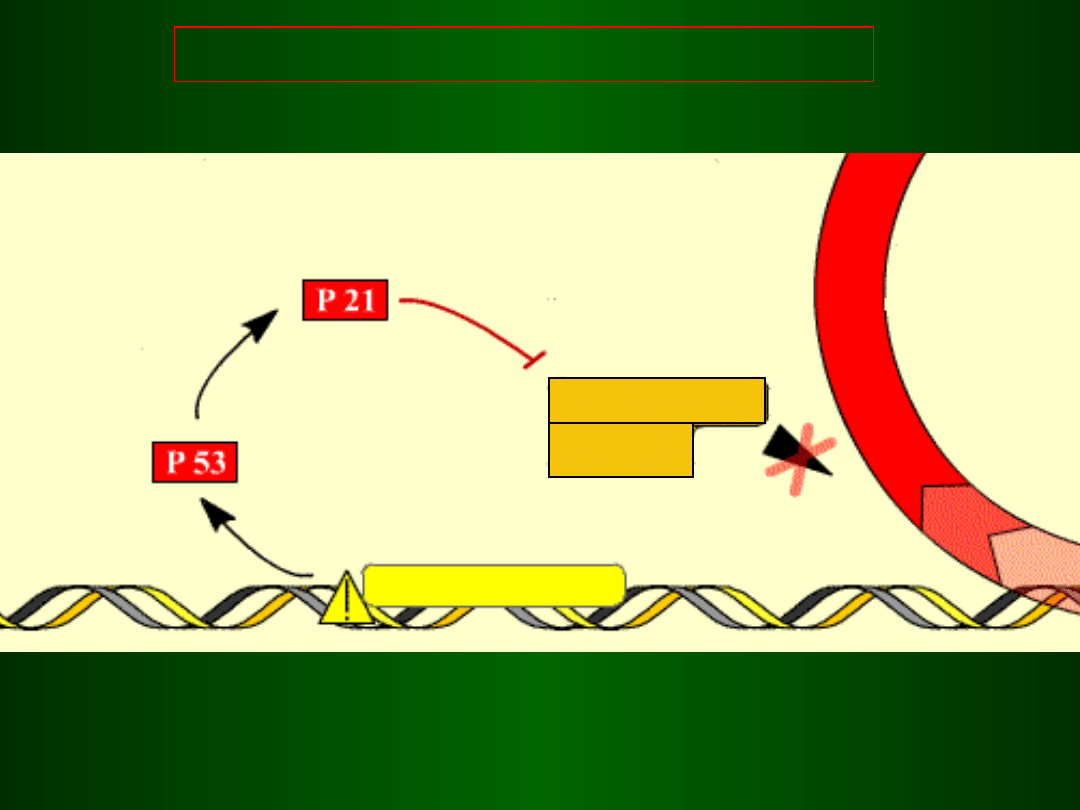
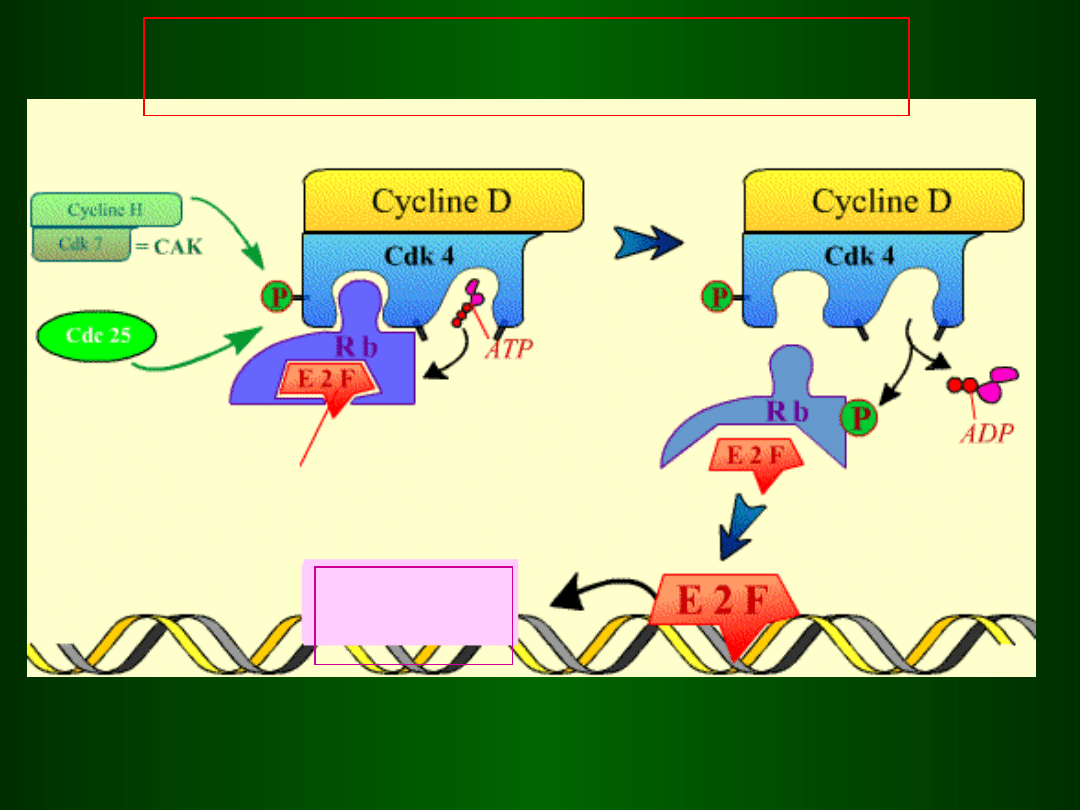
Blokada inicjacji fazy S przez p53
DNA
p53 wiąże się z
DNA
i aktywuje
transkrypcję
genu p21
(inhibitor cyklin D
i E)
Wzrost poziomu i aktywacja
Białka p53 (czynnik transkrypcyjny)
uszkodzone DNA
brak fazy S
p21 blokuje aktywność
kompleksu
i CDK4/6 Cyklina D
Cyklina D
cdk 4/6
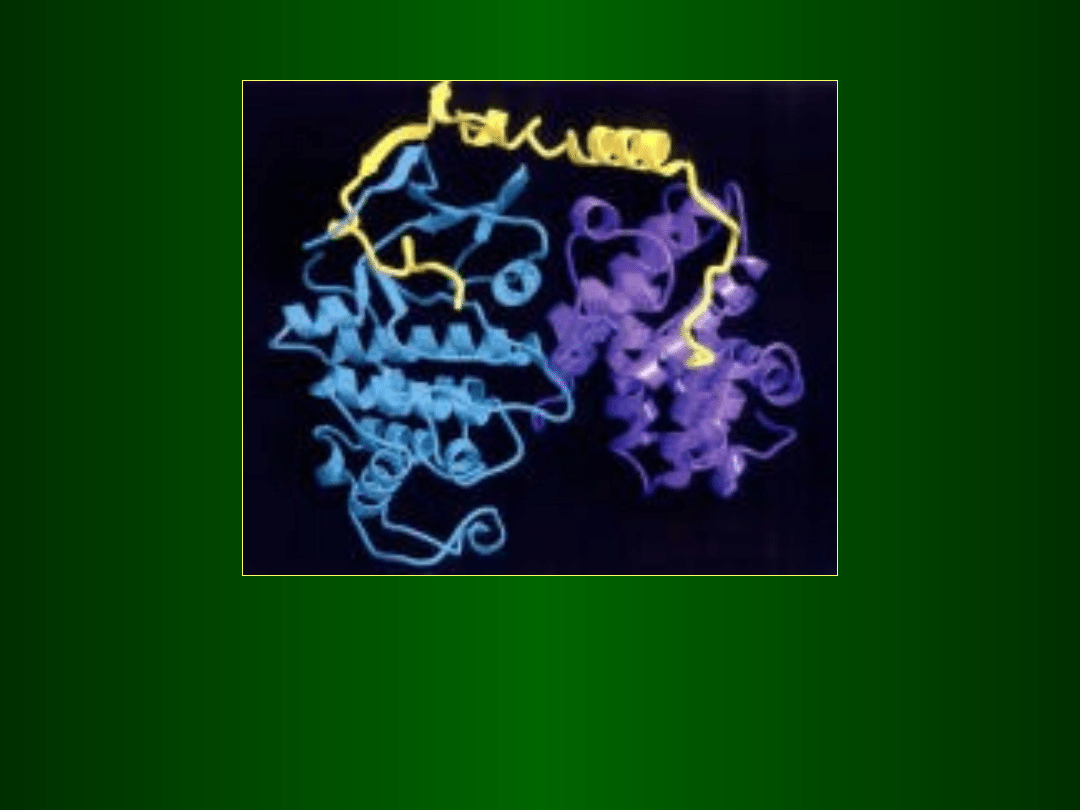
Kinaza Cdk4
(niebieska)
/ cyklinaD
(fioletowa)
zablokowane
przez bialko inhibitorowe p21
(żółty)

Faza G2
Wzrost komórki
Synteza cykliny B
Synteza białek aparatu mitotycznego (tubuliny )
Kontrola prawidłowości zreplikowanego DNA (białko p53
i p21))
Aktywacja MPF przez cdc25 fosfatazę
Faza M
Powstanie dwubiegunowego wrzeciona mitotycznego
Kondensacja i ruchy chromosomów mitotycznych
zależne od MT
Rozproszenie i ponowne odtworzenie otoczki
jądrowej i jąderka
Inaktywacja MPF (Metafaza/Anafaza) i ubikwitynacja
i degradacja w
proteasomach sekuryny, cykliny A
Ubikwitynacja i degradacja cykliny B – wyjście z
mitozy

MPF = CDK1 + Cyklina B
CDK1
jest składnikiem enzymatycznym dimeru
(fosforyluje inne białka enzymatyczne, strukturalne,
regulatorowe
itp.)
Synonimy CDK1:
p34
(bialko o c.cz. 34 kD)
Cdc2,
ponieważ jest kodowana przez gen Cdc 2
(cell division cycle)
gen Cdc 2 –
to jeden z najstarszych i szczególnie
konserwowanych
genów w ewolucji, prawie
niezmieniony od 500 mln lat
C y k l i n a
B
jest białkiem regulatorowym
jest kodowana przez gen Cdc13
Cyklina
cdc13
reguluje aktywność
kinazy cyklinozależnej
cdc2
Charakterystyka MPF (Nagroda Nobla 2001)
Aktywny MPF:
CDK1 jest zdefosforylowana przez fosfatazę Cdc25
przy resztach aminokwasowych tyrozyny 15 (Tyr15) i treoniny 14
(Thr14)
Cyklina B – maksimum koncentracji
Aktywny MPF fosforyluje następujące białka strukturalne:
Histony - skutkiem jest kondensacja chromosomów od profazy do
metafazy
Laminy blaszki jądrowej - skutkiem jest fragmentacja otoczki jądrowej
w profazie
Białka MAP - skutkiem jest powstanie wrzeciona mitotycznego
Nukleoliny - skutkiem jest rozproszenie jąderka w profazie
MPF jest aktywny w przejściu G2/M
MPF jest inaktywowany w
przejściu
Metafaza/Anafaza,
degradacja cykliny B w
anafazie
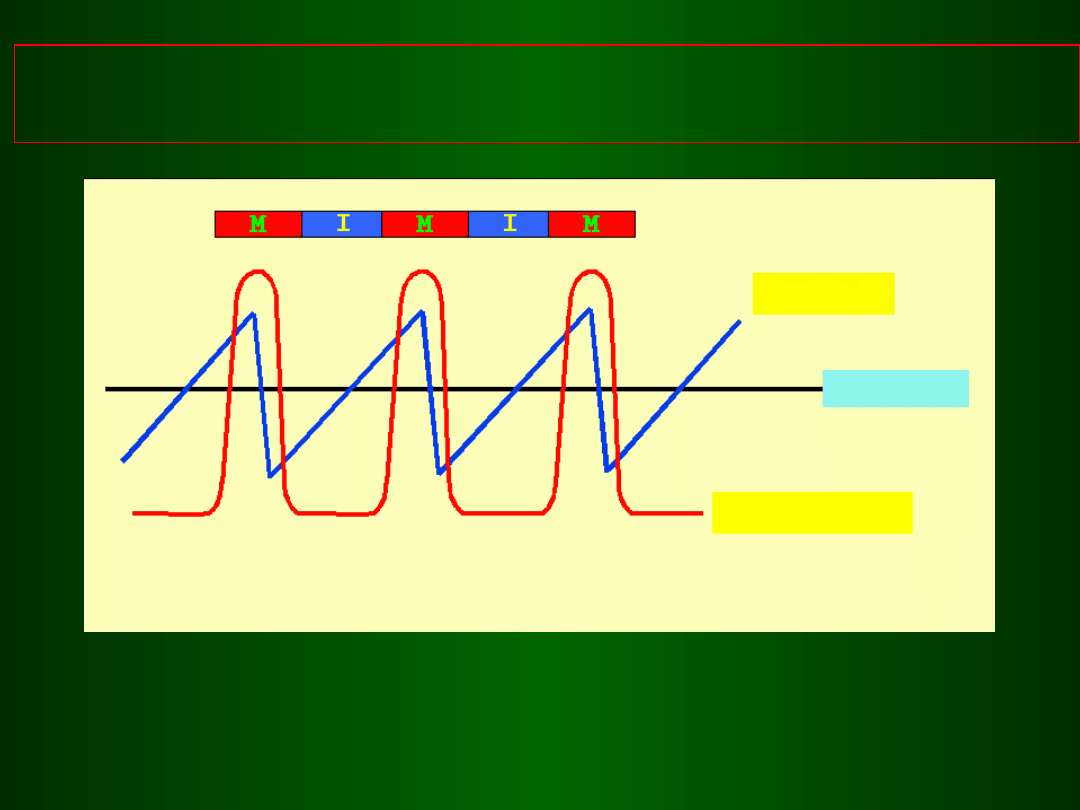
Zależność pomiędzy poziomem cykliny B i
aktywnością MPF
Cyklina B
próg
aktywność MPF
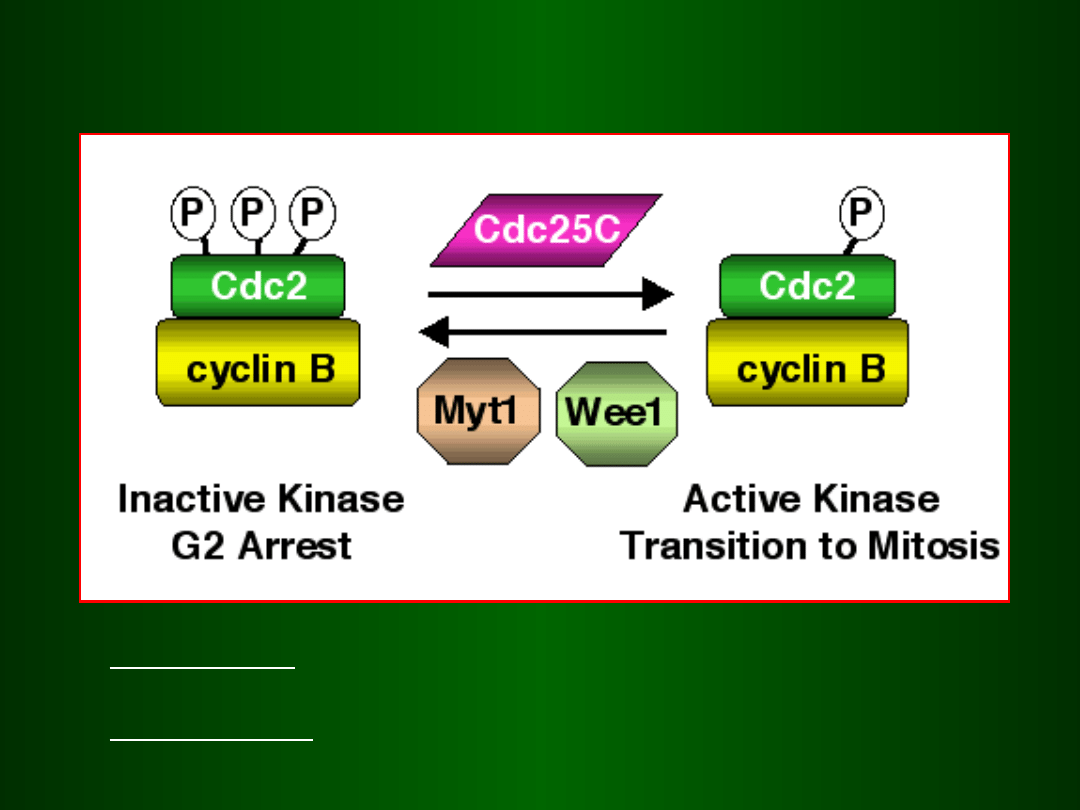
Aktywacja i inaktywacja MPF
Aktywacja: Cdc25C = fosfataza białkowa,
defosforylacja przy tyr15 i thr14
Inaktywacja: Wee1 = kinaza inaktywująca,
fosforylacja przy tyr15 i thr14
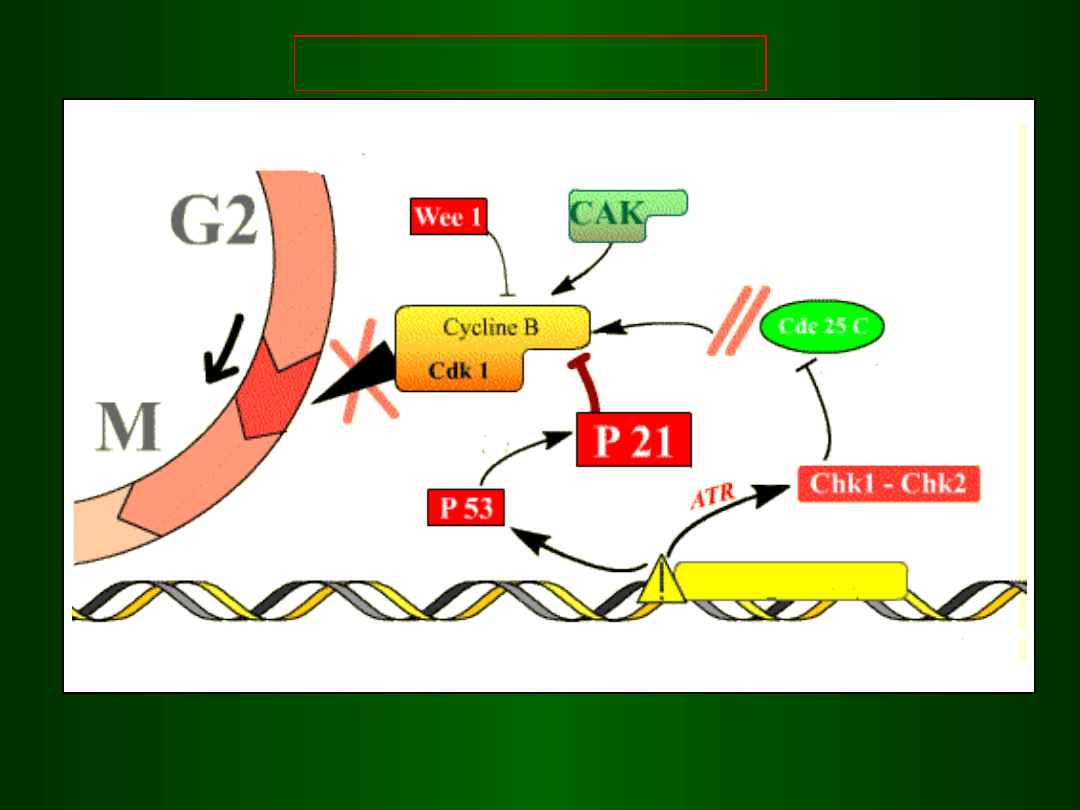
Punkt kontrolny G2/M
DNA
Błąd w
replikacji
synteza
sekwestracja
cdc
25
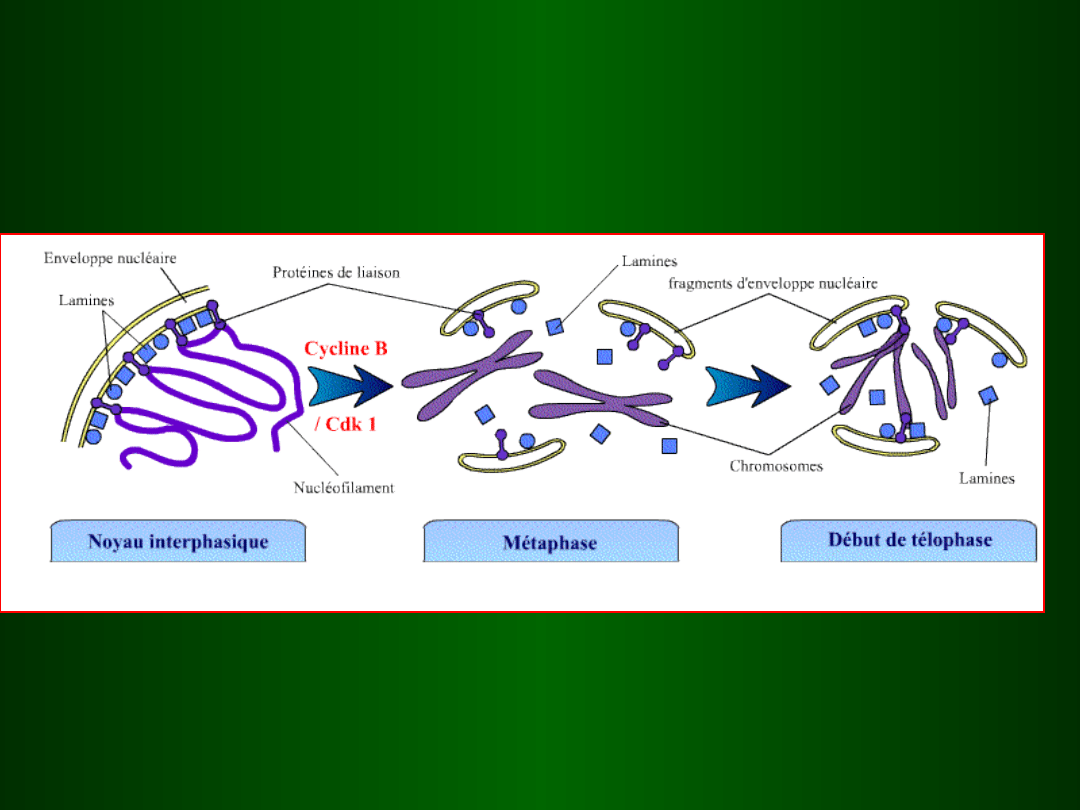
jądro
interfazowe
metafaza
wczesna
telofaza
MPF aktywny
MPF
nieaktywny
Uszkodzenie DNA – odpowiedź komórki
______________________ __________________________________
______________________ __________________________________
ATRIP / Rad26
sensor
ATM / ATR (kinazy)
p53 chk1 (kinaza)
przekaźnik
apoptoza
p21
Cdc25C
Cdc25A
CyklinaD/CDK4/6 CyklinaB/CDK1
efektor
(SPF) (MPF)
__________________________________________________
G1 S G2 M
_______________________________________
3’
3’
5’
5
’
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
Wyszukiwarka
Podobne podstrony:
Biologia Komorki Cykl Komorkowy Nieznany (2)
Cykl komórkowy
CYKL KOMÓRKOWY
komórka, Cykl komórkowy, Cykl komórkowy
Cykl komórkowy, czyli powtórka do sesji
CYKL KOMÓRKOWY, biologia komórki
Cykl komórkowy 1
Cykl komórkowy i starzenie się komórek
16.CYKL KOMÓRKOWY I JEGO REGULACJA, studia-biologia, Opracowane pytania do licencjatu
Cykl komorkowy, farmacja, I sem, biologia
W8 Cykl komórkowy (asus Komputer's conflicted copy 2012 05 26)
W11 Cykl komorkowy
CYKL KOMORKOWY, Biotechnologia i, Rok I, Biologia komórki, Biologia komorki materialy
cykl komórkowy
Prelekcja 2 Cykl komórkowy, biologia
Cykl komorkowy 2007 2008
więcej podobnych podstron