
UAKTYWNIANIE
JEDNOSTAWOWEGO
APARATU RUCHU

UDZIAŁ INFORMACJI
AFERENTNYCH
Z RECEPTORÓW CZUCIA
W WYZWALANIU AKTYWNOŚCI
RUCHOWEJ

BUDOWA ŁUKU
ODRUCHOWEGO

Łuk odruchowy
- jest to
droga jaką przebywa impuls
nerwowy od receptora do
narządu wykonawczego

Składa się z 5-ciu części:
- receptora;
- drogi dośrodkowej (aferentnej);
- ośrodka nerwowego (analizatora);
- drogi odśrodkowej (eferentnej);
- narządu wykonawczego
(efektora).

Receptor
– wyspecjalizowana
komórka lub zespół komórek, które
pod wpływem działającego na nie
bodźca wyzwalają impulsy we
włóknie nerwowym.

Droga dośrodkowa –
druga część
łuku odruchowego. Tworzą ją
włókna nerwowe dośrodkowe
(aferentne), przewodzące impulsy
nerwowe do CUN.

Ośrodek nerwowy –
jest to skupienie
neuronów w ośrodkowym układzie
nerwowym zawiadujące określoną
funkcją.

Droga odśrodkowa –
łączy ośrodek
nerwowy z narządem
wykonawczym. Drogi odśrodkowe
są utworzone przez wypustki
neuronów skupionych
w jadrach nerwów czaszkowych
i jądrach nerwów rdzeniowych.

Narządy wykonawcze (efektory ) –
są zbudowane z komórek
pobudliwych czyli komórek mięśni
poprzecznie prążkowanych, mięśni
gładkich oraz komórek
gruczołowych.

Rodzaje łuków odruchowych:
- monosynaptyczny
(dwuneuronalny);
- bisynaptyczny (trójneuronalny);
- polisynaptyczny
(wieloneuronalny).

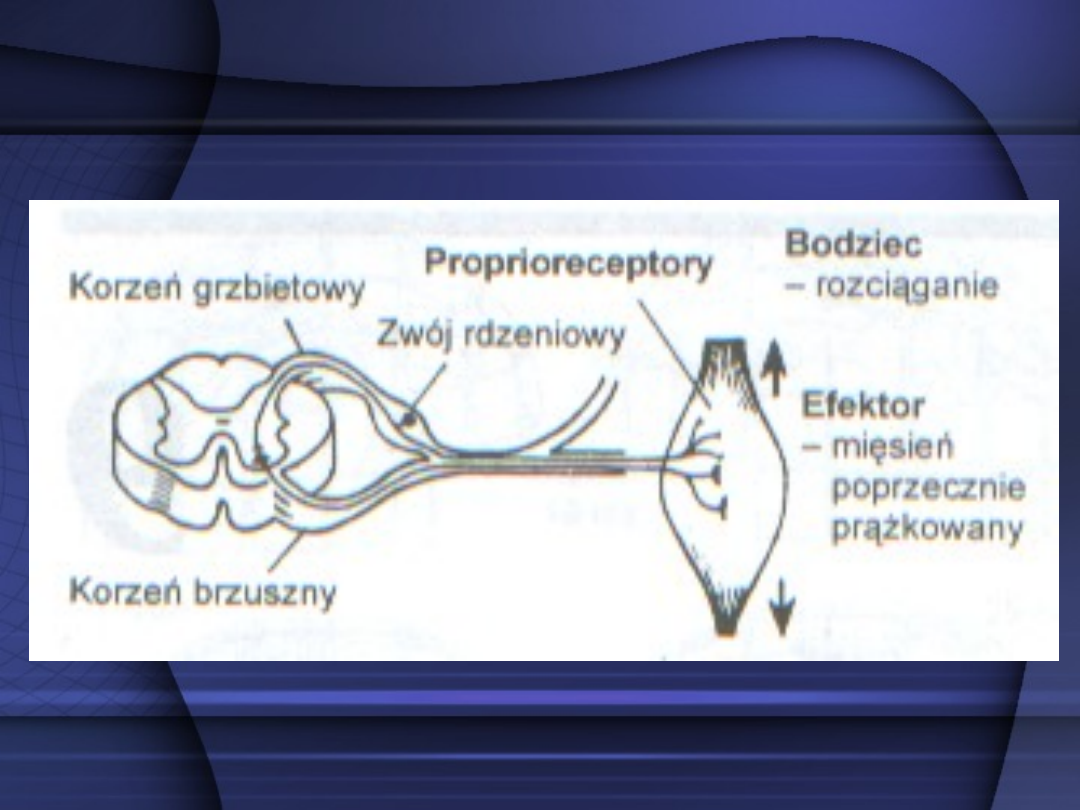
Łuk odruchowy monosynaptyczny –
zawiera jedną synapsę między drogą
dośrodkową a drogą odśrodkową.
Składa się on z neuronu
odbierającego impulsację
dośrodkową, którego ciało znajduje
się w zwoju rdzeniowym
i neuronu ruchowego w rogach
przednich rdzenia kręgowego,
wysyłającego impulsację odśrodkową.

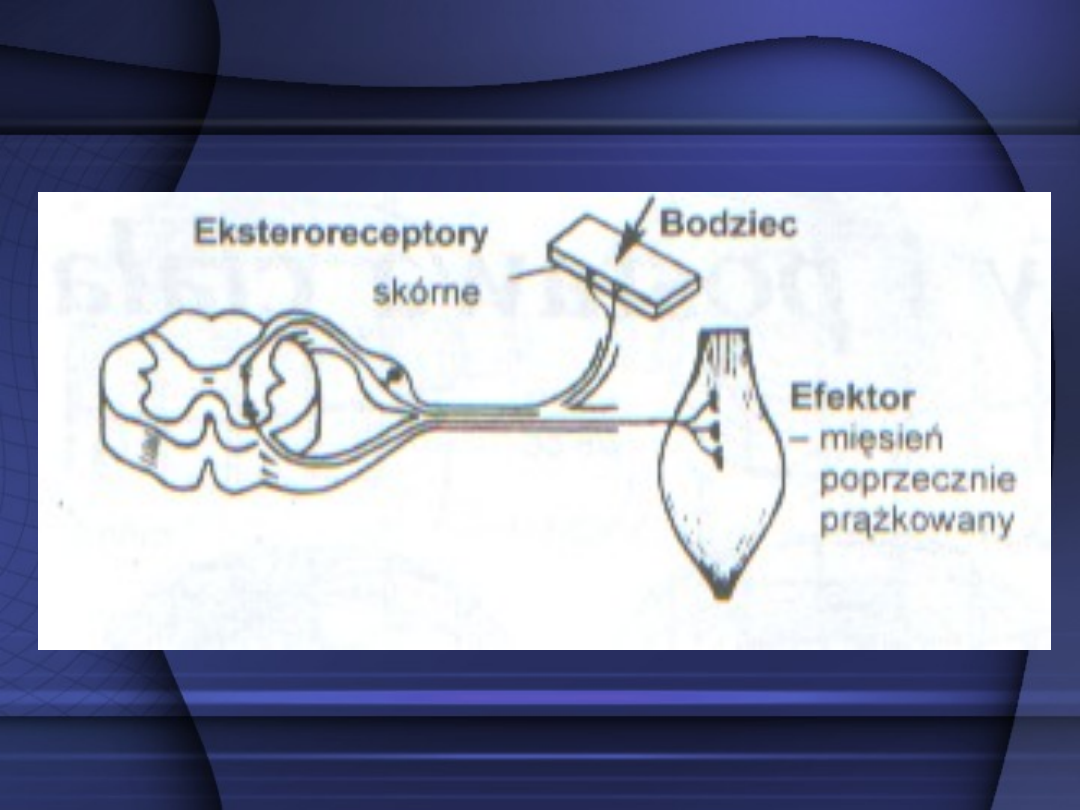
Łuk odruchowy bisynaptyczny –
zawiera dwie synapsy między drogą
odśrodkową a drogą dośrodkową. W
jego skład wchodzi jeden neuron
pośredniczący. Łuk odruchowy
bisynaptyczny jest jednocześnie
łukiem trójneuronalnym.

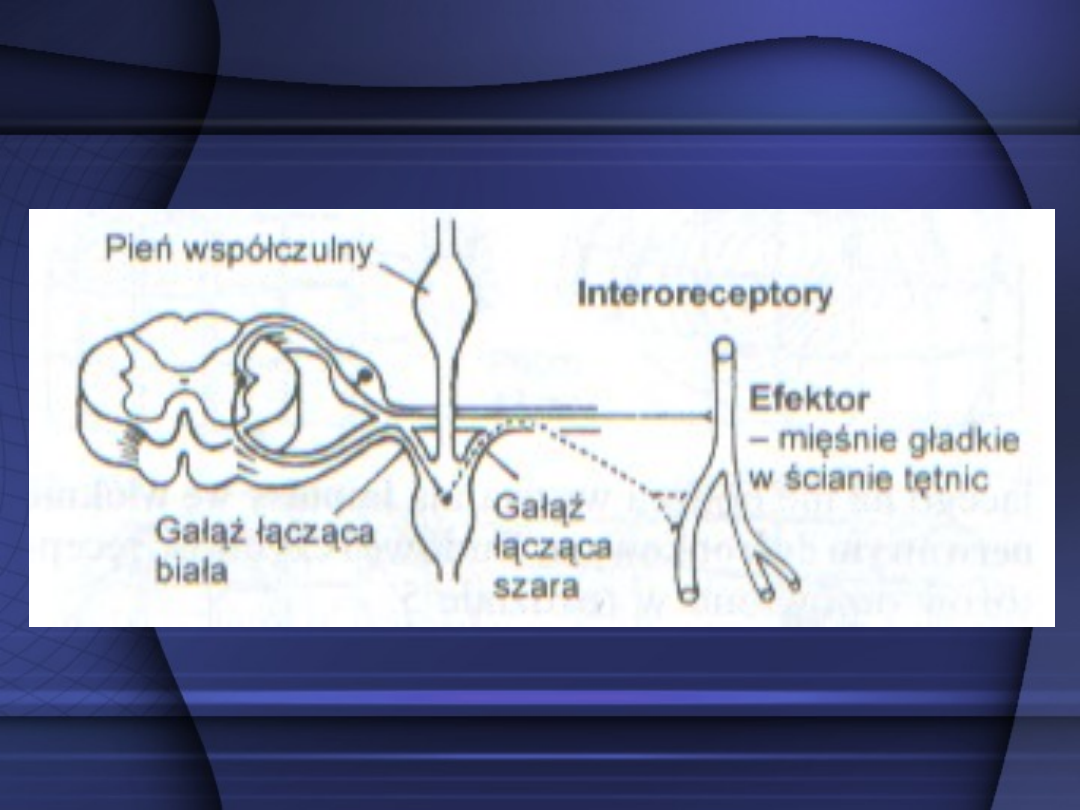
Łuk odruchowy polisynaptyczny –
ma
liczbę synaps większą od dwóch.
Jest to łuk odruchowy
wieloneuronalny. Łuk odruchowy
autonomiczny, z ośrodkiem w
rdzeniu kręgowym, różni się od łuku
odruchowego w układzie nerwowym
somatycznym tym, że zawiera dwa
neurony w drodze odśrodkowej.
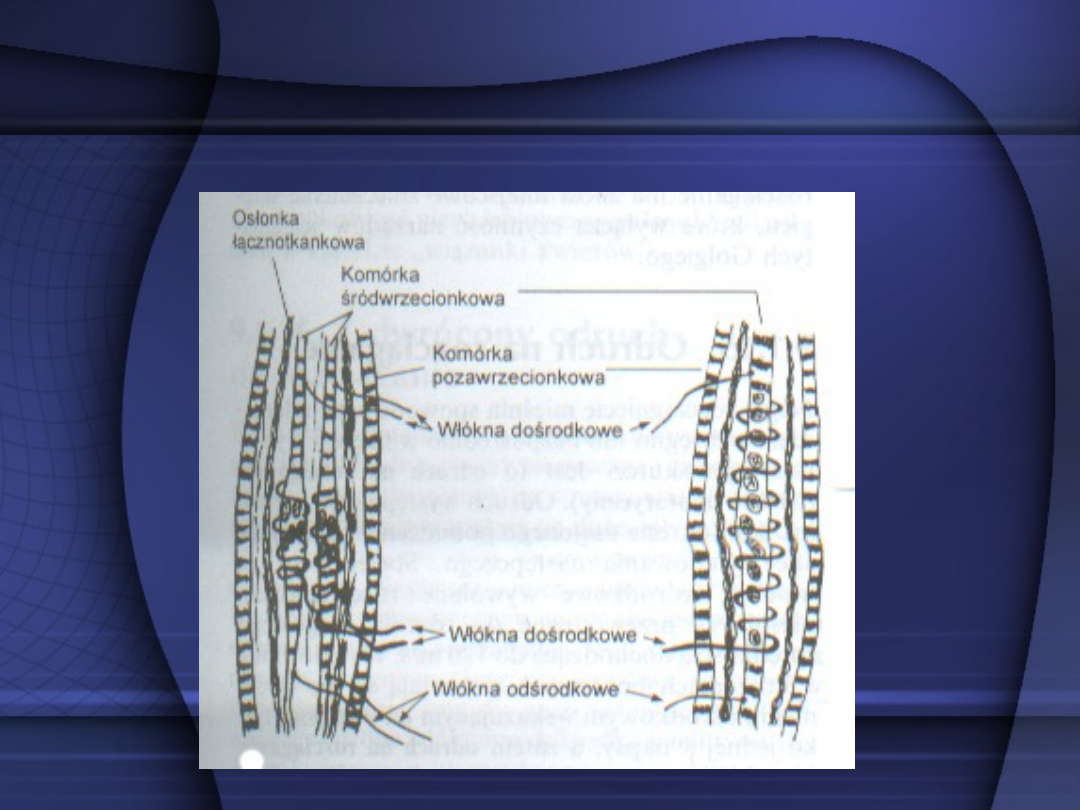
Wrzeciona nerwowo-mięśniowe

Narządy ścięgniste Golgiego
Impulsacja nerwowa dośrodkowa przewodzona od wrzecionek
nerwowo-mięśniowych (Ia i IIa) i od narządów ścięgnistych
Golgiego (Ib i IIb). Górne zapisy (I) uzyskano w czasie
rozciągania mięśnia z jednakową siłą, dolne zapisy (II) zaś w
czasie skurczu pojedynczego mięśnia. Stałe rozciąganie
mięśnia z niewielką siłą pobudza tylko wrzecionka nerwowo-
mięśniowe, skurcz mięśnia zaś pobudza narządy ścięgniste
Golgiego, które wysyłają salwę impulsów.

Receptory stawowe (proprioreceptory)
Receptory te należą do wolno
adaptujących się i informują o
stanie mięśnia a więc o aktywnym
skurczu, rozciągnięciu biernym,
czyli o długości włókien, oraz o
napięciu.

ODRUCHY - RODZAJE
I DEFINICJA

ODRUCH
- jest to odpowiedź
narządu wykonawczego (efektora)
wywołana przez bodziec działający
na receptor
i zachodząca za pośrednictwem
ośrodkowego układu nerwowego.

- odruch miotatyczny (rozciągowy,
„odruch własny mięśnia”)-
jest reakcją
skurczową na nagłe rozciągnięcie
mięśnia spowodowane uderzeniem w
ścięgno lub bezpośrednio
w mięsień. Odruch ten występuje po
bardzo krótkim czasie utajonego
pobudzenia
i nie wykazuje wyładowania
następczego.

- odruch kolanowy (rzepkowy)-
dotyczy korzeni L2, L3,
L4
. Po
uderzeniu młotkiem w ścięgno
rzepkowe obserwuje się skurcz
mięśnia czworogłowego uda.
Należy zwrócić uwagę na
osłabienie lub wzmożenie
odruchów.


-
odruch Hoffmanna (H)-
dotyczy C7,
C8
;
aby go wywołać należy zgiąć gwałtownie
końcowy paliczek palca wskazującego
lub środkowego pacjenta. Po zwolnieniu
palca napięte ścięgno mięśnia zginacza
szybko się rozluźnia. Zgięcie kciuka
wskazuje na wzmożenie odruchów (może
być obecne u osoby zdrowej z żywymi
odruchami ścięgnistymi).


- odruch cofania (zginania)-
jest reakcją
na bodźce uszkadzające skórę lub
tkankę podskórną (bodźce
nocyceptywne). Przykładowo w obrębie
kończyny dolnej dochodzi do skurczu
mięśni zginaczy
i jednoczesnego rozluźnienia
prostowników.
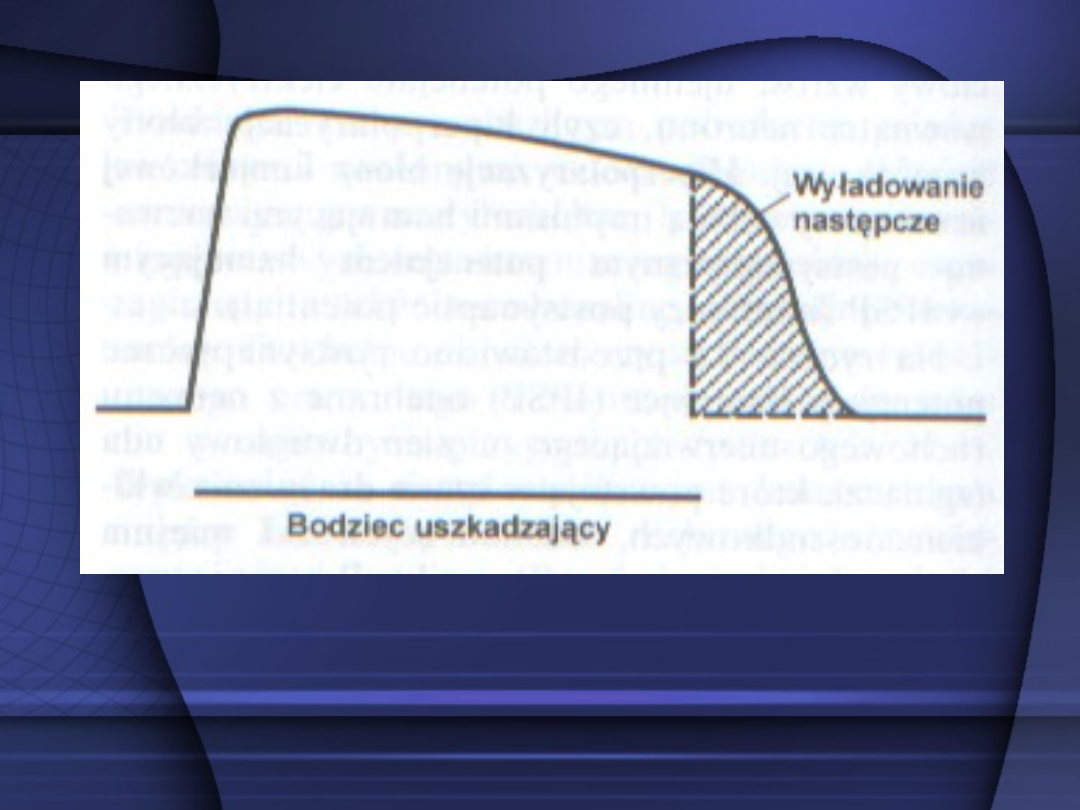
Zapis skurczu odruchowego mięśnia zginacza
wykazujący wyładowanie następcze po
zadziałaniu bodźca uszkadzającego

- skrzyżowany odruch prostowania-
jest nieodłącznie związany z
odruchem zginania. Zazwyczaj
reakcji odruchowej kończyny
podrażnionej towarzyszy odruchowy
skurcz prostowników kończyny
kontralateralnej. Zachodzi on przy
jednoczesnym rozluźnieniu zginaczy
tej kończyny. Skrzyżowany odruch
prostowania usztywnia kończynę w
celu utrzymania pozycji ciała.

ORGANIZACJA
NEURONALNA RDZENIA
KRĘGOWEGO

Istota szara każdego odcinka
rdzenia kręgowego zawiera wiele
tysięcy neuronów, które dzielą się
na neurony czuciowe, ruchowe,
pośredniczące
i neurony układu autonomicznego.

Neurony ruchowe (motoneurony) –
są
zgrupowane w jądrze ruchowym,
w rogu przednim. Należą do nich
neurony ruchowe alfa i gamma.
Wypustki neuronów alfa unerwiają
komórki mięśniowe poprzecznie
prążkowane (miocyty). Pobudzenie
neuronów alfa wywołuje skurcz
mięśnia szkieletowego.

Neurony gamma -
są rozrzucone między
neuronami alfa, są od nich mniejsze
i stanowią 30% wszystkich neuronów
ruchowych. Odchodzące od nich
włókna unerwiają komórki mięśniowe
znajdujące się we wrzecionkach
nerwowo – mięśniowych, które są
receptorami czułymi na rozciąganie
mięśni.

Pobudzanie neuronów gamma nie
wywołuje skurczu całego mięśnia,
a jedynie skurcz komórek
mięśniowych wrzecionek nerwowo
– mięśniowych, zmieniając ich
pobudliwość.

Neurony pośredniczące –
występują
w całej istocie szarej rdzenia kręgowego.
Liczba tych neuronów jest 30×większa od
liczby neuronów ruchowych. Skupienie
neuronów pośredniczących między
rogiem tylnym a przednim zwane jest
jądrem pośrednim. Neurony
pośredniczące tworzą między sobą liczne
połączenia. Aksony niektórych neuronów
pośredniczących kończą się na
neuronach ruchowych.

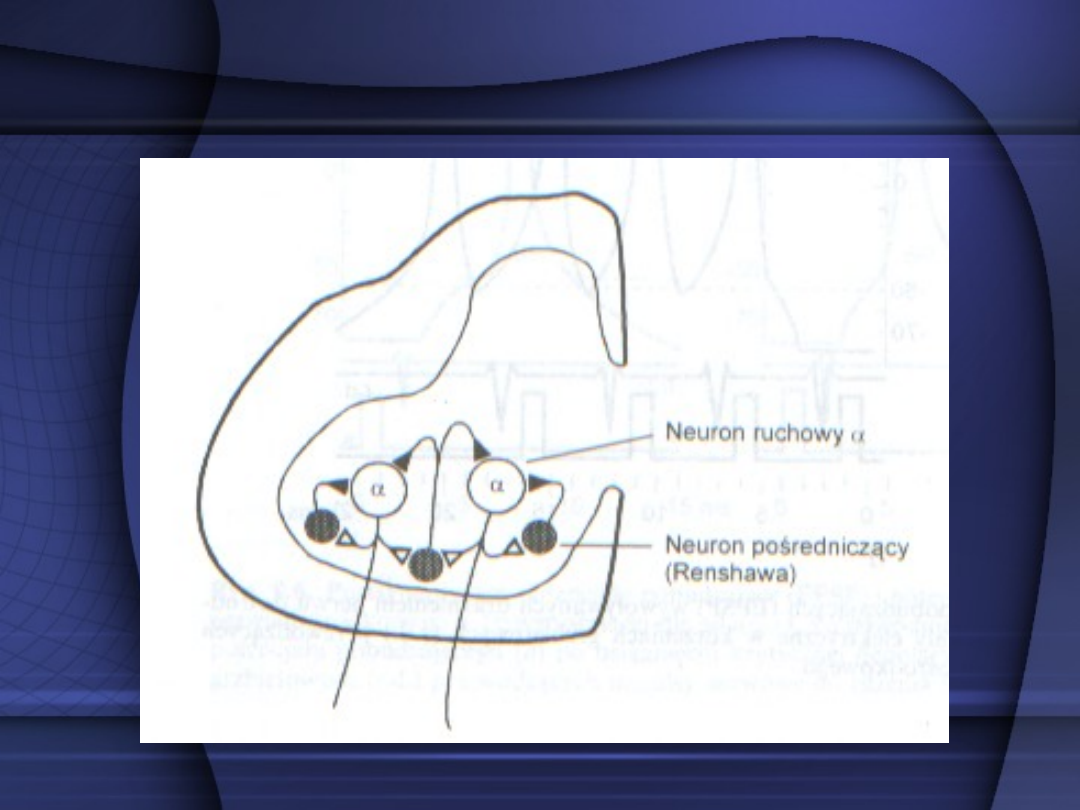
Osobną grupą są komórki
znajdujące się w rogu przednim,
zwane
neuronami pośredniczącymi
(Renshawa)
, które uczestniczą w
hamowaniu zwrotnym neuronów
ruchowych.

Neurony należące do układu
autonomicznego są zgrupowane w rogu
bocznym. Neurony części współczulnej
układu autonomicznego znajdują się w
odcinkach Th i trzech górnych L rdzenia.
Neurony ośrodkowe części
przywspółczulnej układu
autonomicznego zgrupowane są w
drugim, trzecim i czwartym odcinku
części S rdzenia.

ROLA PROPRIOCEPTORÓW
W CZYNNOŚCIACH
ODRUCHOWYCH

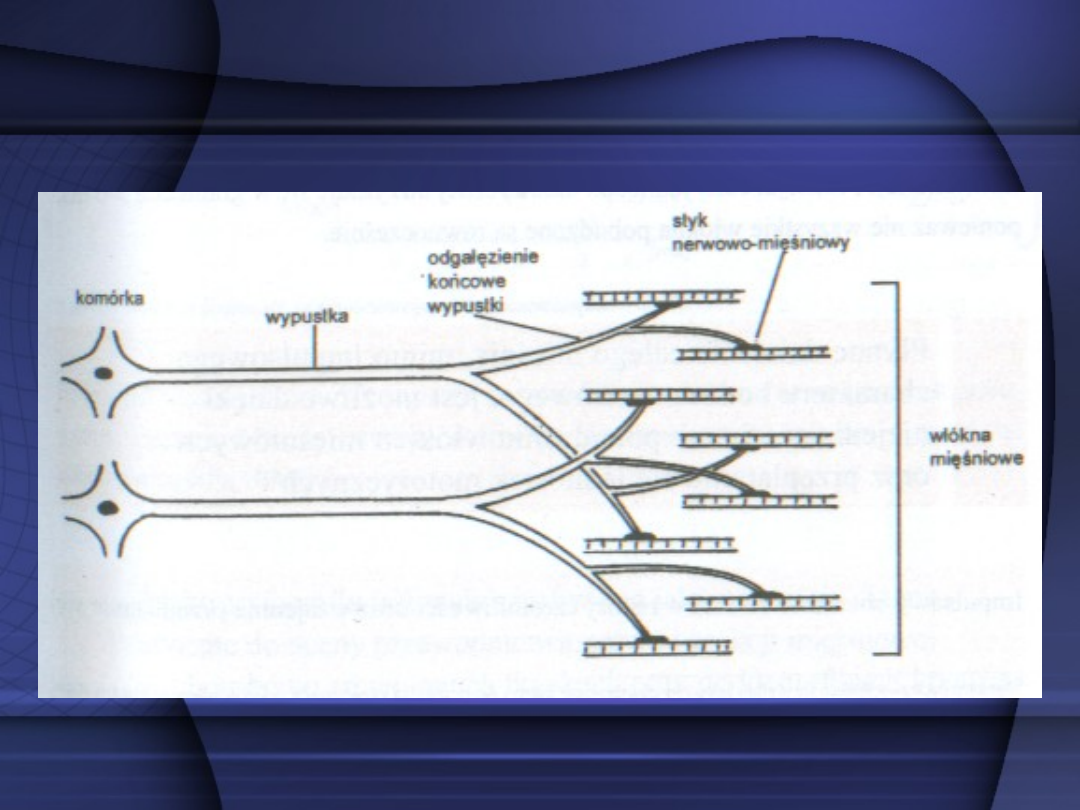
Odgałęzienia każdego aksonu
neuronu ruchowego przednich
rogów istoty szarej rdzenia
kręgowego unerwiają kilka do
kilkuset komórek mięśniowych.
Grupę komórek mięśniowych oraz
unerwiający je neuron nazywa się
jednostką motoryczną
.

Pomiędzy poszczególnymi komórkami
mięśniowymi nie ma żadnych połączeń
umożliwiających przekazywanie pomiędzy
nimi stanu czynnego. Są one od siebie
całkowicie izolowane. Wobec tego stan
czynny powstający w jednostce motorycznej
na skutek pobudzenia jej motoneuronu
ogranicza się do niej i nie jest przekazywany
na inne jednostki. W czasie fizjologicznego
skurczu tylko część jednostek motorycznych
zostaje aktywowana. W skutek tego istnieje
możliwość regulacji siły skurczu przez zmianę
liczby (rekrutacji) aktywowanych jednostek.

Siła skurczu każdej komórki mięśnia
może być regulowana przez stopień
jej spoczynkowego rozciągnięcia i
zmianę częstotliwości pobudzeń,
ponadto siła skurczu całego mięśnia
może być regulowana przez
zmianę liczby pobudzonych
jednostek motorycznych.

Włóknami nerwowymi – ruchowymi –
przenoszone są bodźce elektryczne
z układu nerwowego do mięśnia
(włókien mięśniowych). Przejście
włókna nerwowego do komórki
mięśniowej ma złożoną strukturę.
Bodziec elektryczny przenoszony przez
włókno mięśniowe jest krótkotrwały,
w stosunku do jednostek motorycznych
asynchroniczny i o dużej częstotliwości,
dochodzącej do
50 Hz.

Czas trwania pojedynczego impulsu
potencjału czynnościowego
pojedynczego włókna wynosi
1- 2
milisekund
. Potencjał całej jednostki
motorycznej utrzymuje się w
granicach od
5 do 6 milisekund
ponieważ nie wszystkie włókna
pobudzane są jednocześnie.

Płynne działanie całego mięśnia,
mimo impulsowego charakteru
bodźca nerwowego, jest możliwe
dzięki niejednoczesnemu
pobudzeniu włókien mięśniowych
oraz przeplataniu się jednostek
motorycznych.

CZYNNIKI MIĘŚNIOWE.
WŁAŚCIWOŚCI
MECHANICZNE MIĘŚNI.

ZALEŻNOŚĆ POMIĘDZY DŁUGOŚCIĄ MIĘŚNIA
A WIELKOŚCIĄ ROZWIJANEJ SIŁY.
Pojedyncze włókienko wyzwala maksymalną
siłę przy długości sarkomeru
od 2 do
2,25 μm
, czyli wówczas, gdy wszystkie
połączenia między mostkami miozynowymi
i nitkami aktyny są związane. Siła ta maleje
wraz z jego rozciągnięciem, co tłumaczy się
tym, że w jego trakcie zwiększa się liczba
połączeń mostkowych. Maleje ona również
wówczas, gdy sarkomer skraca się i nitki
miozyny dochodzą do linii Z.

Sarkomer – odcinek wydzielony liniami Z.
Schemat podłużnego przekroju włókienek
mięśniowych : grubych – miozynowych i
cienkich – aktynowych.

Jeżeli za wyjściową długość sarkomeru
przyjmuje się wartość spoczynkową
lo=2,25 μm to przy jego
rozciągnięciu do 3,65 μm (62 %
długości spoczynkowej), jego siła
spadnie do 0. Maksymalnie skrócony
sarkomer ma wymiar 1,27 μm czyli
może się skrócić o 44 %. Wówczas
jego siła też spada do 0.

Mięsień ma elementy czynne, zdolne do
wyzwalania siły, i elementy bierne, jak
ścięgna i powięzie oraz pozostałe
tkanki łączne. Własności sprężyste
tych elementów muszą być
uwzględnione, jeśli rozważana jest
zdolność mięśnia do rozwijania siły,
gdyż za ich pośrednictwem jest ona
przenoszona na kość. Ścięgno musi
przenosić przynajmniej taką siłę, jaką
rozwija brzusiec.

Elementy bierne mają pewną sprężystość,
która polega na odzyskiwaniu pierwotnych
kształtów po usunięciu naprężeń
mechanicznych wywołujących
odkształcenie. Może ono być spowodowane
działaniem elementów kurczliwych lub przy
ich biernym zachowaniu przez siłę
zewnętrzną. Skutkiem tego odkształcenia
jest pojawienie się wewnętrznych sił
sprężystości oraz związanej z ich istnieniem
potencjalnej energii mechanicznej, tzw.
energii sprężystości.

SZYBKOŚĆ ZMIAN DŁUGOŚCI (PRĘDKOŚCI)
MIĘŚNIA. ZALEŻNOŚĆ F-V. MOC MIĘŚNIA.
Mięśnie szkieletowe w układzie ruchu człowieka
pełnią rolę siłowników służących do napędu jego
części biernych – segmentów ciała. Funkcja ta jest
realizowana dzięki temu, że mięśnie wykazują
zdolność aktywnej zmiany długości, jednocześnie
generując siłę, która – przenoszona przez
dźwignie kostne – może poruszać segmenty ciała
względem siebie oraz oddziaływać na zewnątrz
układu ruchu wywołując siły reakcji. Obydwie te
funkcje – zmiana długości i wyzwalanie siły –
oznaczają, że mięśnie szkieletowe są zdolne do
wykonywania pracy mechanicznej.

Prędkość przemian energetycznych
zachodzących w mięśniu
szkieletowym ma wartość
skończoną, zatem i moc
maksymalna, rozwijana przez
mięsień, jest skończona i nie może
przekroczyć pewnej wartości
granicznej.

Wartość siły rozwijanej przez
mięsień jest zależna od prędkości
jego skracania się; zależność ta
w przybliżeniu jest odwrotnie
proporcjonalna.

Zarówno siła maksymalna, jak
i maksymalna prędkość skracania
się mięśnia są ograniczone pewnymi
wartościami maksymalnymi,
charakterystycznymi dla danego
mięśnia, zależnymi od jego budowy
(np. składu włókien), temperatury
itp.
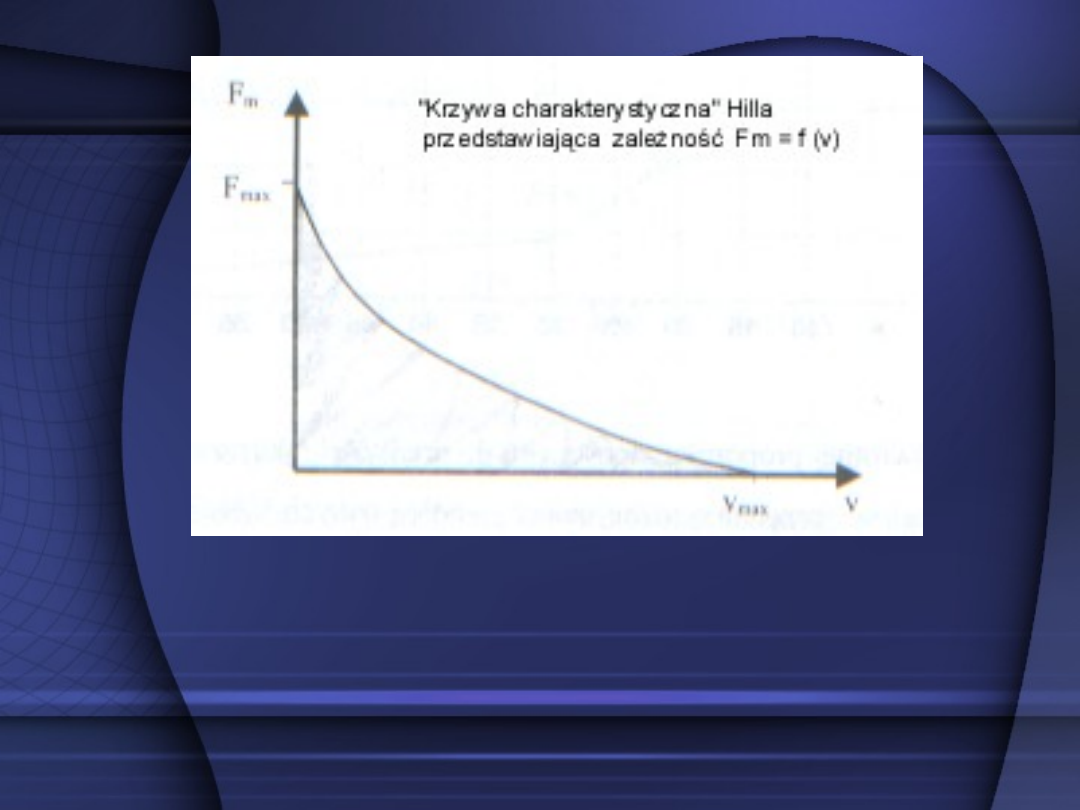
Przebieg hiperboli tłumaczy zależność między siłą
a prędkością : największą siłę (maksymalną) rozwija
mięsień przy prędkości V=0. Przy maksymalnej zaś
prędkości skracania się mięśnia przejawia on na
zewnątrz zerową siłę.

Zależność siły mięśnia od prędkości
jego skurczu wiąże się z mocą
mięśnia. Moc rozwijana przez
mięsień pracujący izometrycznie
(tzn. wyzwalający maksymlaną siłę)
jest równa zeru. Podobnie jest z
mocą mięśnia kurczącego się z
prędkością maksymalną. Oznacza
to, że w obydwu przypadkach moc
użyteczna mięśnia jest równa zeru.

Dla pośrednich wartości prędkość
skurczu mięśnia, iloczyn siły
mięśnia
i prędkości jest większy od zera.
Oznacza to, że dla pewnej wartości
V moc rozwijana przez mięsień
osiąga maksimum.

Moc użyteczna mięśnia zależy od
prędkości jego skracania, a jej
wartość maksymalna jest
rozwijana przy pewnej określonej
prędkości skurczu.

ARCHITEKTURA MIĘŚNI
– BIOMECHANICZNY
MODEL BUDOWY.

Zachowanie mięśni szkieletowych,
nazywanych też poprzecznie
prążkowanymi zależy –
w przeciwieństwie do mięśni
gładkich trzewi – od woli człowieka.
Należą one do czynnego układu
ruchu i są zdolne do wyzwolenia siły
oraz wykonania pracy mechanicznej.

Brzusiec mięśnia jest zbudowany
z włókien mięśniowych otoczonych
powięzią przechodzącą w ścięgna,
które przyczepiają się do kości.
Ścięgna rozpostarte między kością a
brzuścem przenoszą siłę pobudzonych
włókien mięśniowych na kości. Ze
względu na szeregowe położenie
ścięgien wobec włókien mięśniowych
przenoszą one taką siłę, jaką wyzwala
mięsień. Mięsień jest pokryty dwiema
warstwami tkanki łącznej – omięsnymi.

Obie warstwy omięsnej zawierają
włókna kolagenowe oraz sprężyste,
które przechodzą w warstwę
śródmięsną otaczającą pojedyncze
włókna mięśniowe. Cytoplazmatyczna
błona komórkowa, otaczająca zespół
włókien składających się na włókno
mięśniowe, nazywa się
sarkolemą.
Specyficzną rolą sarkolemy jest
przenoszenie pobudzenia.

Tkanki łączne, stanowiące otoczki
poszczególnych elementów czynnych
mięśni wraz ze ścięgnem stanowią
jego elementy bierne i mają znaczenie
w przenoszeniu siły mięśnia. Włókienko
składające się z segmentów zwanych
sarkomerami – odcinki przedzielone
liniami Z – jest zbudowane
z nitek
aktyny
i
miozyny
, zwanych
łącznie nitkami mięśniowymi –
miofilamentami
.

Schemat układu mostków miozynowych względem
nitek aktyny a) w stanie spoczynku, b) pod
wpływem jonów wapnia Ca++ mostki tworzą
połączenia
z aktyną, przy czym następuje aktywacja ATP,
c) energia rozpadu ATP zostaje zużyta do zmiany
kąta ustawienia głowy mostka, co powoduje
przesunięcie aktyny względem miozyny.

Dziękuję za uwagę.
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
- Slide 41
- Slide 42
- Slide 43
- Slide 44
- Slide 45
- Slide 46
- Slide 47
- Slide 48
- Slide 49
- Slide 50
- Slide 51
- Slide 52
- Slide 53
- Slide 54
- Slide 55
- Slide 56
- Slide 57
- Slide 58
- Slide 59
- Slide 60
- Slide 61
- Slide 62
- Slide 63
- Slide 64
- Slide 65
- Slide 66
- Slide 67
- Slide 68
Wyszukiwarka
Podobne podstrony:
UAKTYWNIANIE JEDNOSTAWOWEGO APARATU RUCHU, FIZJOTERAPIA, NEUROLOGIA
Budowa i rozwój aparatu ruchu
DIAGNOSTYKA APARATU RUCHU--8 nowe(1), Fizjoterapia, fizjoterapia
biologiczne podstawy, uklad miesniowy, Budowa i rozw?j aparatu ruchu
biologiczne podstawy, Stawy, Budowa i rozw?j aparatu ruchu
biologiczne podstawy, Stawy, Budowa i rozw?j aparatu ruchu
DIAGNOSTYKA APARATU RUCHU
Aparat ruchu zasady funkcjonowania
Dochody budżetowe jednostek samorządu terytorialnego ćw
2 Aparat ruchu nmg
01 ĆW. APARATU ARTYKULACYJNEGO , ĆW
biomechanika - aparat ruchu, Praca własna studenta
Aparat ruchu i skóra - PYT, Sprawdziany i Odpowiedzi gimnazjum, Sprawdziany z odpowiedziami z biolog
[14.10.2014] Aparat trójosiowy, ćw.11, Politechnika Koszalińska
więcej podobnych podstron