
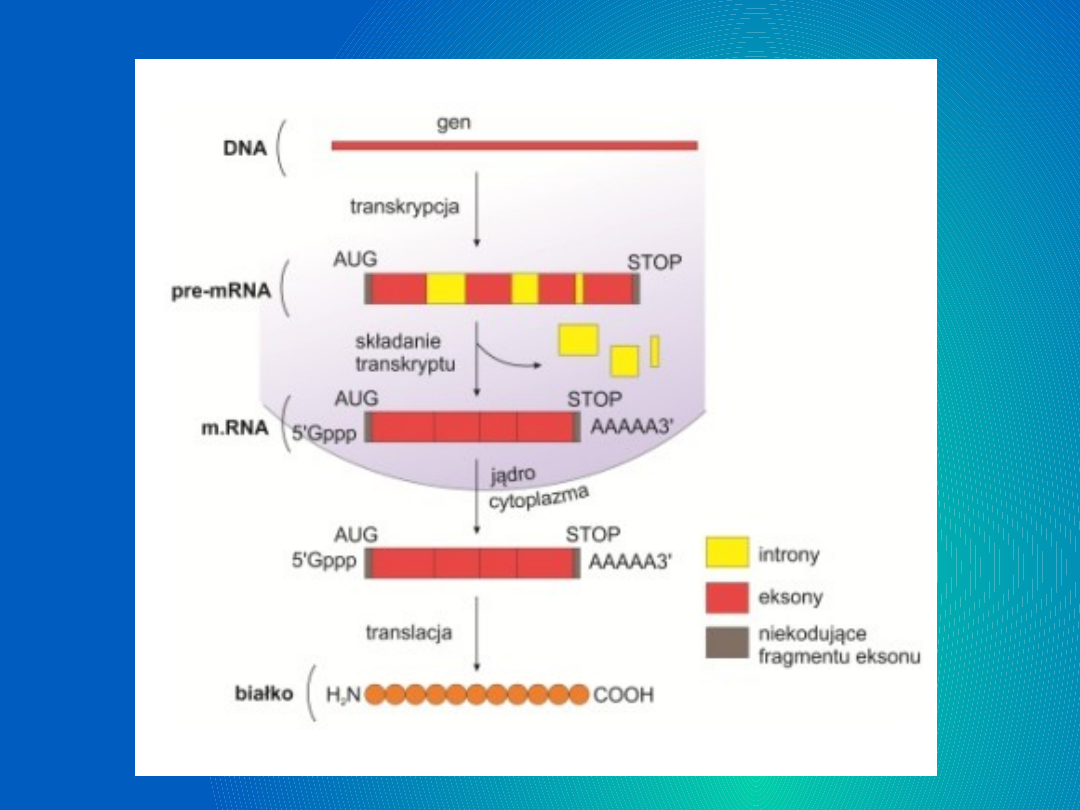
Dojrzewanie
pre-mRNA

pre-mRNA
RNA będący bezpośrednim produktem
transkrypcji, najczęściej zawierający
niekodujące sekwencje nukleotydów.
Elementy niekodujące,
introny
, są
usuwane w procesie dojrzewania
(splicingu). Po wycięciu niekodujących
sekwencji, dołączeniu
guanylowej czapki na końcu
5' łańcucha i sekwencji poli-A nakońcu
3' łańcucha powstanie
właściwy mRNA biorący udział
w translacji.

Dojrzewanie
Pre-mRNA nim stanie się matrycą
pełnowartościową do syntezy cząsteczki
białka musi przejść przez proces jakim
jest dojrzewanie. Dojrzewanie pre-mRNA,
polega na modyfikowaniu obu końców
pre-mRNA. Na końcu 5' zostaje dodana
czapeczka a do końca 3' zostaje dodana
sekwencja 30-200 nukleotydów
adeninowych. To tzw. koniec poli (A).

Dojrzewanie składa się z 3 etapów:
-capping ( czyli dodawanie czapeczki )
-poliadenylacja ,która podnosi stabilność
transkryptu;
-splicing -w nim są eliminowane niekodujące
wewnętrzne rejony RNA informacyjnego.
Alternatywna poliadenylacja i splicing generować
mogą różne produkty ( mRNA ) które wykrywane są
w tkankach różnych, co oznacza, iż jeden gen
kodować może więcej niż jedno białko.
Dojrzewanie pre-mRNA stanowi warunek eksportu z
jądra transkryptu.
Dojrzały mRNA ulega związaniu z białkami
wykazującymi wpływ na translację, stabilność i
transport tej cząsteczki.

Capping
wpływa na zwiększenie stabilności pre-mRNA.
Zaopatrzony w czapeczkę transkrypt jest
mniej wrażliwy na działanie fosfataz i nukleaz
tj. enzymów które odpowiedzialne są za
degradację kwasu nukleinowego. Rozpoznanie
czapeczki jest również ważne w czasie inicjacji
translacji.
Tworzenie czapeczki zachodzi z transkrypcją
równolegle, przed zsyntezowaniem całości nici
mRNA pierwotnego.
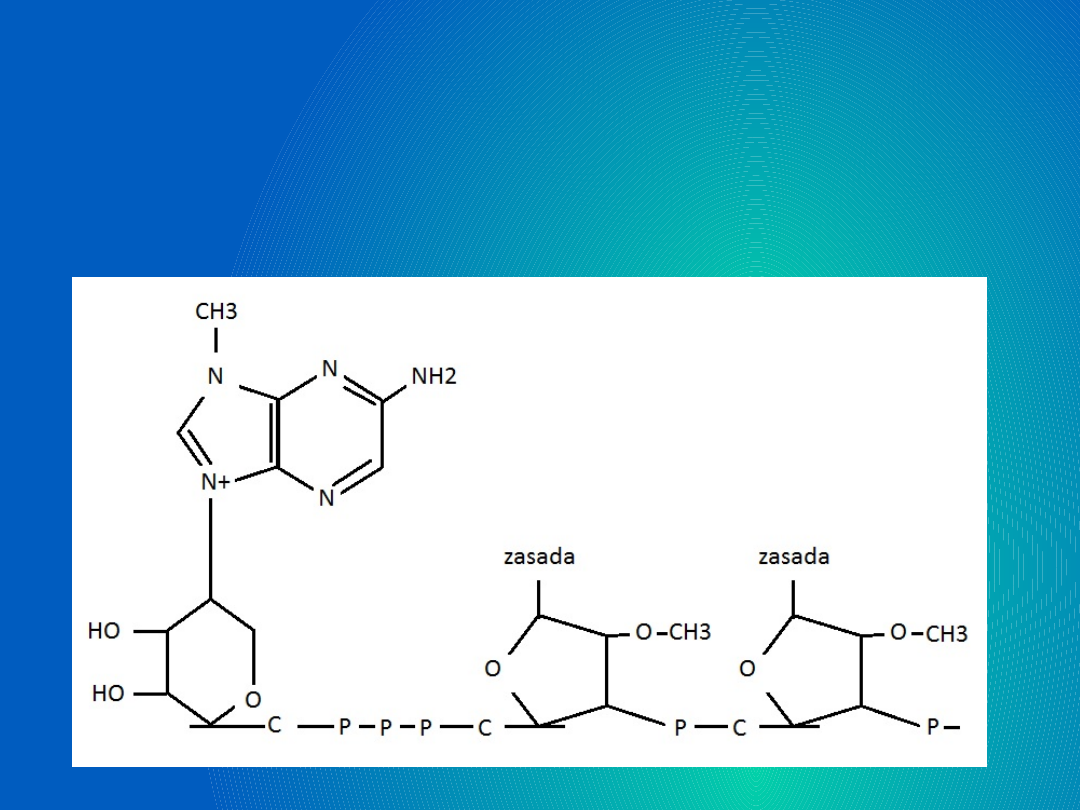
Trifosforan 7-metyloguanozynowy
przyłączony do
2’-O-metylorybonukleozydu, do jego
grupy
5’-hydroksylowej przez 3 reszty
fosforanowe.
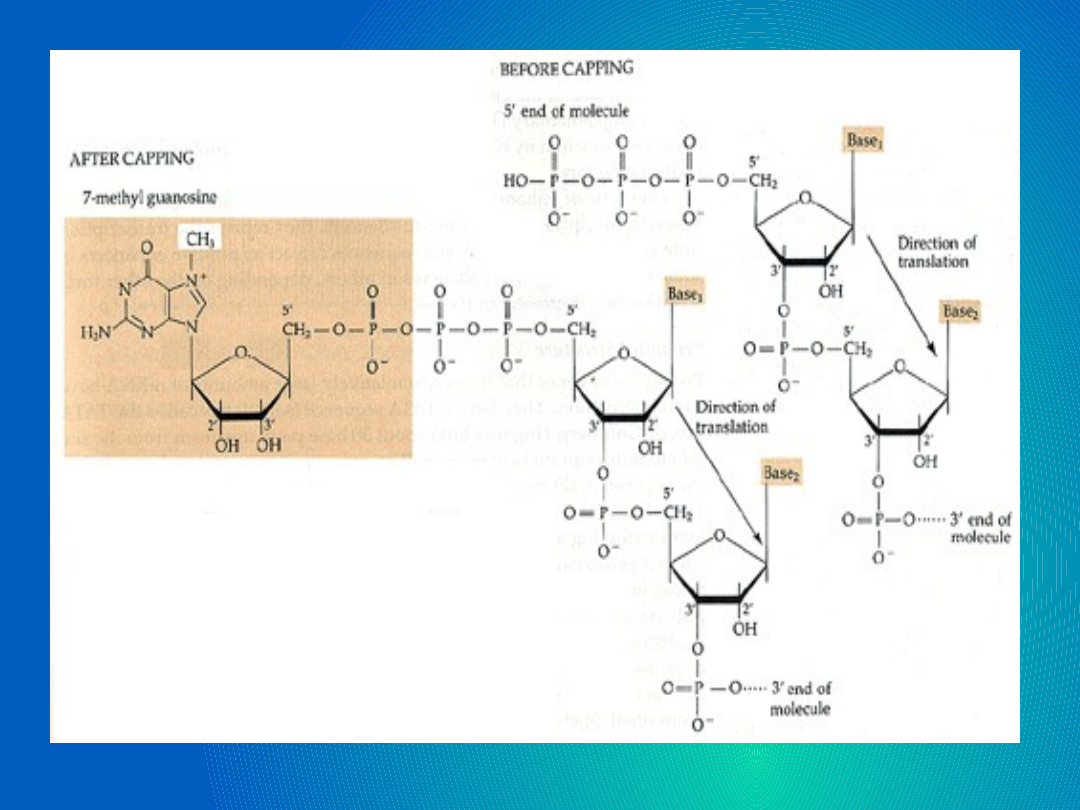

Proce wytworzenia czapeczki składa się z
3 części:
-modyfikacji trifosforanu z końca 5'
poprzez uwolnienie pojedynczej
fosforanowej reszty drogą hydrolizy;
-przyłączenia GTP 5'-5'-trifosforanowym
wiązaniem;
-metylacji guaniny zachodzącej w pozycji
N7 czyli przyłączenie metylowej reszty (-
CH3 ) do 7-go atomu azotu w danej
cząsteczce guaniny.

Poliadenylacja
W pobliżu końca 3’ cząsteczki występuje
sekwencja zasad, która jest sygnałem do
dodania fragmentu złożonego z szeregu
nukleotydów adeninowych, czyli tzw.
fragmentu poli(A). Enzymy znajdujące się w
jądrze rozpoznają ten sygnał i przecinają w
tym miejscu cząsteczkę mRNA. Następnie do
końca 3’ zostaje przyłączone od 30 do 200
nukleotydów adeninowych, tworząc ogon
Poli(A).
Syntezę ogona poli(A) zablokować może
kordycepina.
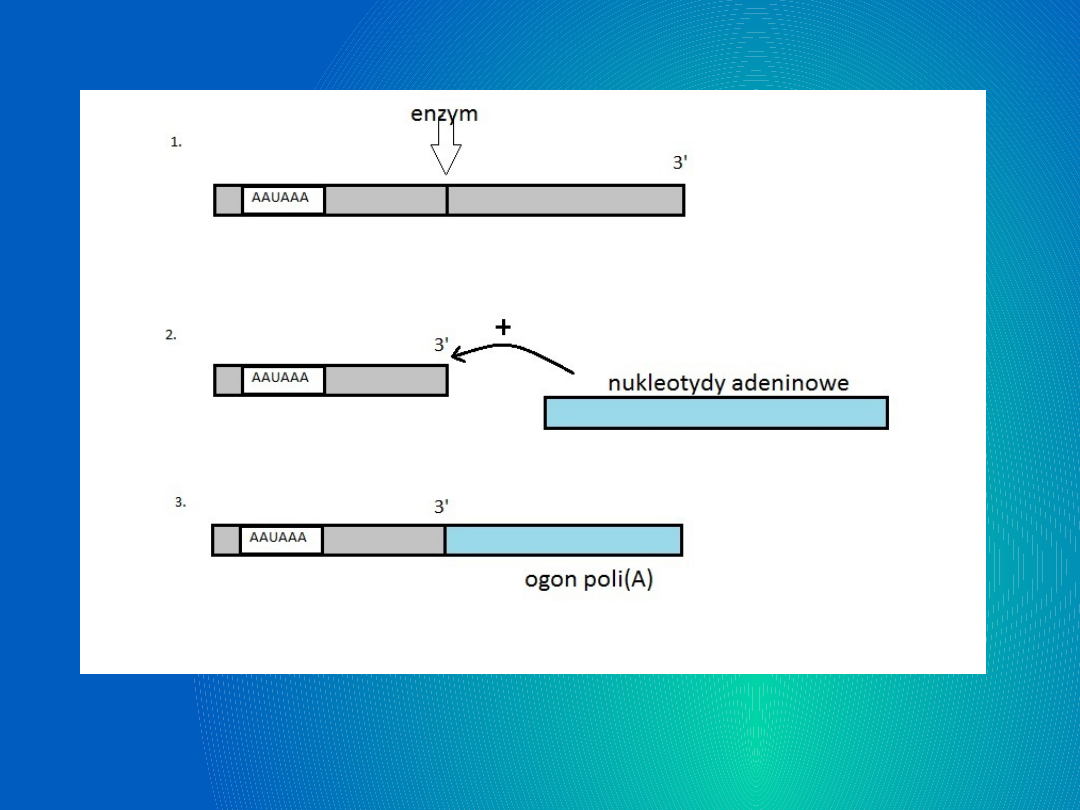

Atakowane poprzez nukleazę miejsce nie jest
miejscem dowolnym, jest wyznaczone przez
sekwencję: AAUAAA zlokalizowanej w
zróżnicowanej odległości od miejsca
przeznaczonego działania enzymu.
Specyficzna nukleaza tnie kwas nukleinowy nici
pre-mRNA na cząsteczki. Do ostatniego
nukleotydu (końca 3`) polimeraza II RNA dodaje
kolejne adeninowe nukleotydy. Powstaje ogon
poliA.
Podobnie jak czapeczka ogon poli A chroni
cząsteczkę transkryptu pierwotnego przed
działaniem różnych nukleaz. Ponadto może mieć
znaczenie w procesie translacji, bowiem okazuje
się, iż transkrypt bez ogona poli A stanowi
matrycę mniej wydajną przy syntezie różnych
białek.

Polimeraza II RNA dodaje na końcu 3'
transkryptu pierwotnego kolejne adeninowe
nukleotydy dopiero po tym jak zadziała
specyficzna nukleaza czyli enzym tnący
nukleinowy kwas w ramach jego cząsteczki.
Odcinek poli A nie jest jak się okazuje
dołączany do nukleotydu ostatniego
wbudowanego w drodze transkrypcji, lecz do
tego właśnie, który okazał się ostatnim
nukleotydem po rozcięciu danej nici pre-mRNA

Wycinanie intronów – splicing
Nim powstanie dojrzały, ostateczny mRNA mogący
podlegać translacji, pre-mRNA musi zostać
pozbawiony intronów. Proces ten wykorzystuje jeden
aparat enzymatyczny wspólny dla pozbywania się
wszystkich intronów.
To oznacza, iż introny muszą posiadać pewne wspólne
elementy które rozpoznawane są przez wspomniany
splicingowy aparat. Z porównywania różnych
-sekwencji intronów wynika, iż łączą je podobieństwa
następujące:
-na końcu 5' posiadają zawsze kolejno: guaninową
oraz uracylową resztę (5'- GU)
-na ich końcu 3' występuje guaninowa reszta
poprzedzona adeninową resztą (AG - 3')
-zawierają wewnątrz tzw. miejsce rozgałęzienia;
kluczową rolę dla splicingu w tym miejscu odgrywa
adeninowy nukleotyd.

Etapy splicingu:
1.Przecięcie lewego końca (5’) intronu –
tworzy się pętla między końcem 5’ i
przy końcu 3’
2. Nacięcie drugiego końca powoduje
uwolnienie pętli.
3. Eksony łączą się w jednolitą matrycę.
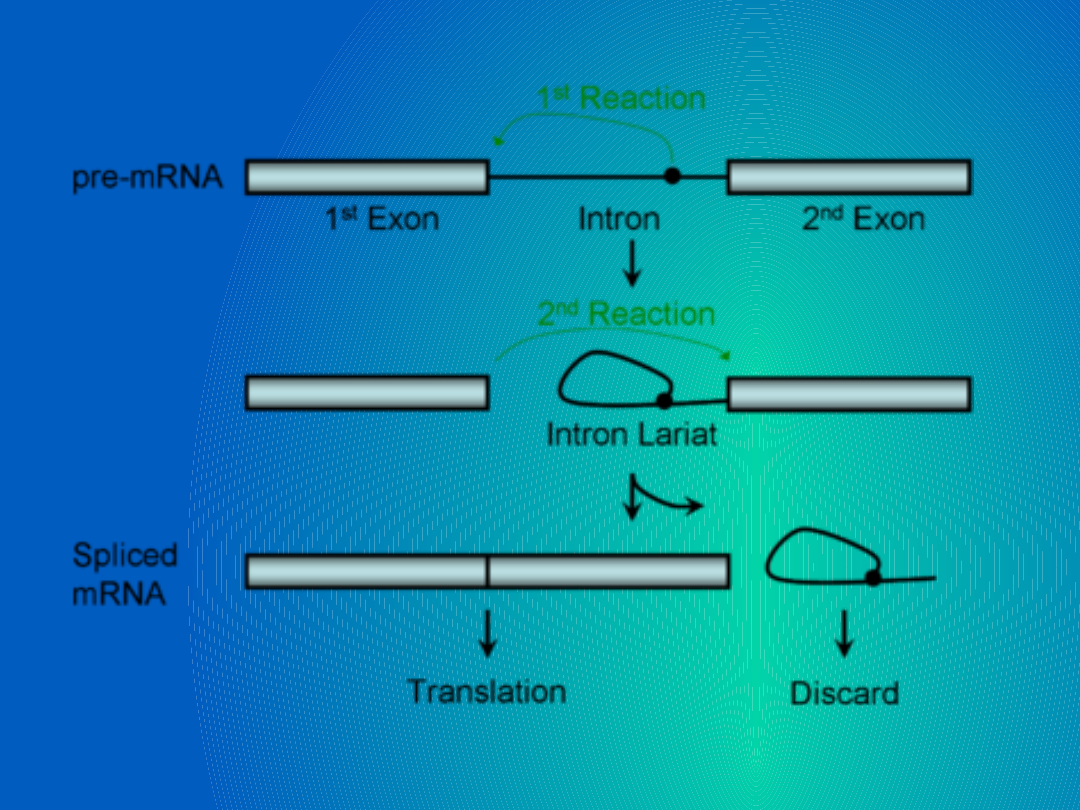

Spliceosom
Struktura katalizująca proces przekształcania
pierwotnego tran skryptu na mRNA. Składa się z
małych, jądrowych cząsteczek
rybonukleoproteinowych (sn-RNP) oraz białek,
towrzących wspólnie kompleksy.
Wyróżnić można 5 rodzajów sn-RNA: U1, U2, U4, U5
oraz U6, a każdy z nich pełni istotną, odrębną
funkcję:
- U1- ulega połączeniu ze splicingowym miejscem 3' i
5' w oparciu o zasadę komplementarności, bowiem
zawiera on w swojej komponencie rybonukleinowej
sekwencję komplementarną względem styków egzon
- intron ;
- U2 - umożliwia wiązanie miejsca rozgałęzienia;
- U6 - obecne w kompleksie w połączeniu z U5 i U4,
katalizuje splicing.

Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
Wyszukiwarka
Podobne podstrony:
Dojrzewanie pre mRNA
Ekspresja genów część 2 Transkrypcja i dojrzewanie pre mRNA 1
DOJRZEWANIE PRE, biochemia
2008 pre mRNA splicing and human did GenDev
TRANSKRYPCJA I DOJRZEWANIE mRNA
Prezentacja 13 Dojrzewanie 2
NS2 lab 4 4 7 en Configure Cisco IOS IPSec using Pre Shared Keys
connections pre intermediate minimock test 0 4 b
Etapy dojrzewania kopalnej materii organicznej
Darmowe Doładowanie telefonu., $$$Darmowe doładowania i kody pre-paid
Kilka przepisów na wędliny dojrzewające
10 dojrzewanie limfoc
Okres dojrzewania
opoznione dojrzewanie plciowe i Nieznany
więcej podobnych podstron