1
TRANSMISJA IMPULSÓW NERWOWYCH – materiały dodatkowe
Neuroprzekaźnik
(Przekierowano z
Neurotransmiter
)
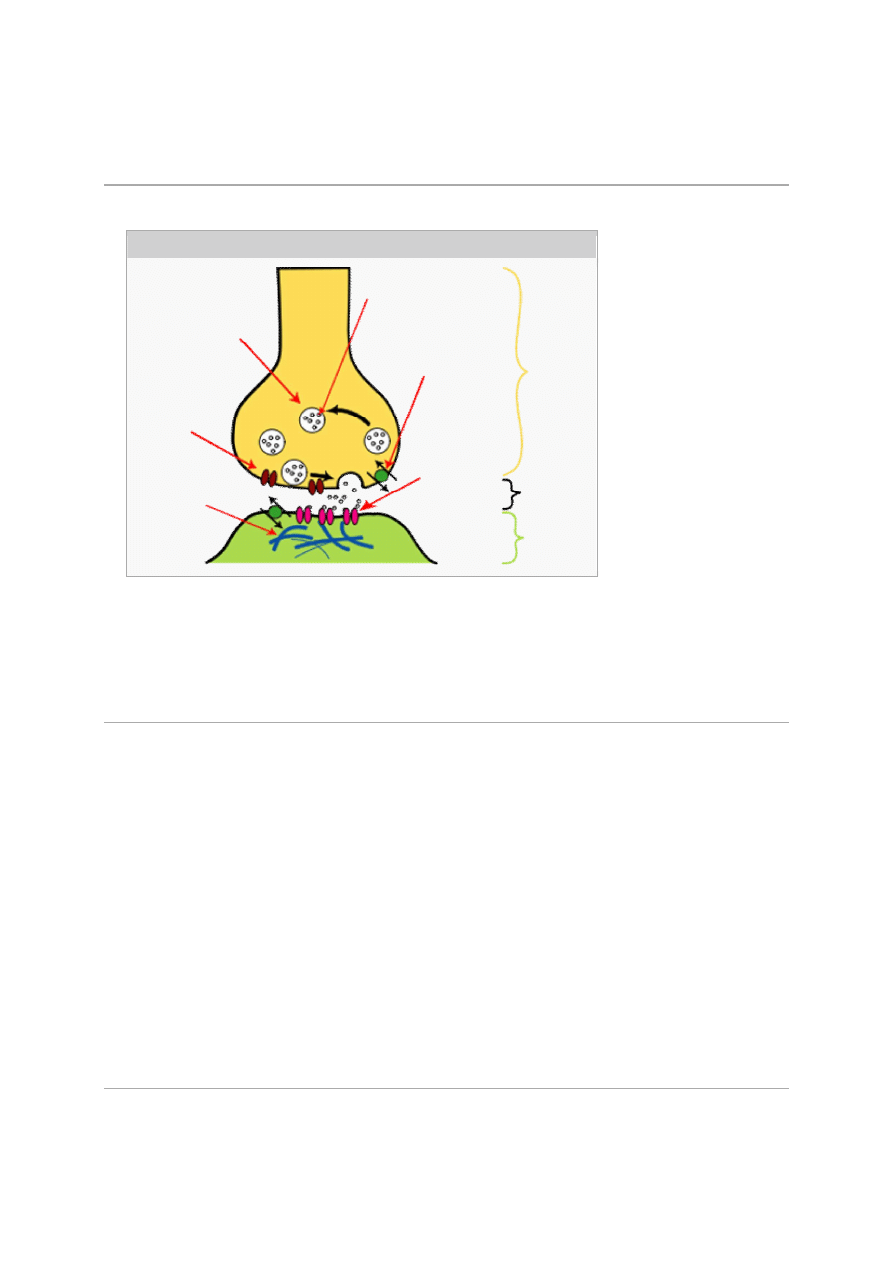
Struktura typowej
synapsy
chemicznej
Neuroprzekaźnik, neurotransmiter, neuromediator –
związek chemiczny
,
którego
cząsteczki
przenoszą
sygnały
pomiędzy
neuronami
(komórkami nerwowymi)
poprzez
synapsy
, a także z komórek nerwowych do mięśniowych lub gruczołowych. Najbardziej
rozpowszechnionymi neuroprzekaźnikami
są:
glutaminian
,
GABA
,
acetylocholina
,
noradrenalina
,
dopamina
i
serotonina
.
Działanie neuroprzekaźnika
Neuroprzekaźnik służy do zamiany sygnału elektrycznego na sygnał chemiczny w
synapsie
i do
przekazywania tego sygnału z jednej komórki (zwanej presynaptyczną) do innej (zwanej
postsynaptyczną). W klasycznym przypadku neuroprzekaźnik jest zgromadzony w pęcherzykach
synaptycznych znajdujących się w komórce presynaptycznej blisko błony presynaptycznej. W
rezultacie depolaryzacji błony presynaptycznej pęcherzyki te przyłączają się do błony
presynaptycznej, następuje fuzja ich błony z błoną presynaptyczną i
egzocytoza
czyli uwolnienie
zawartego w nich neuroprzekaźnika do szczeliny synaptycznej – zamiana sygnału elektrycznego
na chemiczny. Na błonie postsynaptycznej występują
receptory
danego neuroprzekaźnika.
Przyłączenie neuroprzekaźnika do błony postsynaptycznej powoduje zmianę jej polaryzacji (tzn.
ujemnego
potencjału elektrycznego
wnętrza komórki postsynaptycznej mierzonego względem
przestrzeni zewnątrzkomórkowej). W przypadku synapsy pobudzającej jest to zmiana dodatnia,
zwana depolaryzacją. W przypadku synapsy hamującej jest to zmiana ujemna, zwana
hyperpolaryzacją. Tak więc następuje tu zamiana sygnału chemicznego na elektryczny. W obu
przypadkach ta zmiana polaryzacji jest następnie przenoszona wzdłuż błony komórki
postsynaptycznej i w pewnych przypadkach, jeżeli jest wystarczająco silna, może być
propagowana wzdłuż
aksonu
.
Potencjał czynnościowy
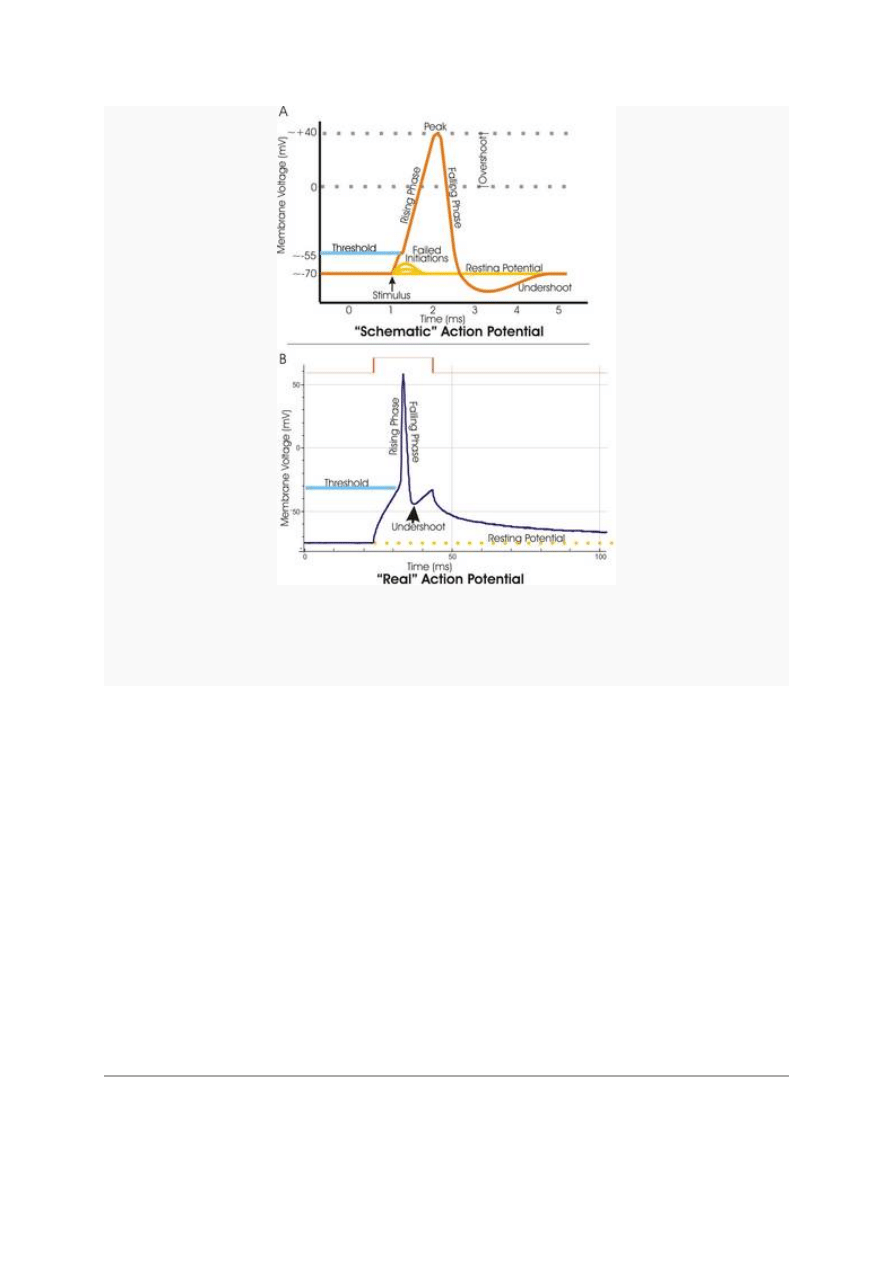
2
Figure 1. A. Wyidealizowany obraz potencjału czynnościowego (iglicowego), pokazujący jego poszczególne
fazy. B. Rejestrowane różnymi technikami rzeczywiste potencjały czynnościowe mogą odbiegać od
idealnego wzorca.
Potencjał czynnościowy (czyli iglicowy) - przejściowa zmiana potencjału błonowego komórki,
związana z przekazywaniem informacji. Bodźcem do powstania potencjału czynnościowego jest
zmiana potencjału elektrycznego w środowisku zewnętrznym komórki. Wędrujący potencjał
czynnościowy nazywany jest impulsem nerwowym. Faza
depolaryzacji
i
repolaryzacji
potencjału
czynnościowego (iglica) trwa nie więcej niż 1 ms i osiąga maksymalnie wartości około
+30
mV
.
Hiperpolaryzacja
następcza może trwać kilkadziesiąt milisekund. W trakcie potencjału
czynnościowego neurony stają się niepobudliwe, zaś później, podczas hiperpolaryzującego
potencjału następczego ich pobudliwość jest zmniejszona. Zjawiska te nazywamy refrakcją
bezwzględną i względną
[1]
. Ze względu na okres refrakcji bezwzględnej oraz refrakcji względnej
komórki nerwowe człowieka nie mogą generować potencjałów czynnościowych z dowolną
częstotliwością. Jednak w najbardziej sprzyjających okolicznościach częstotliwość potencjałów
czynnościowych może dojść do 100 impulsów na sekundę
[2]
.
Miejscem powstawania potencjału czynnościowego w komórce nerwowej jest tzw. wzgórek
aksonowy, skąd potencjał iglicowy rozprzestrzenia się po powierzchni błony komórkowej wypustki
osiowej- aksonu. Generowaniem potencjałów czynnościowych rządzi zasada: "wszystko albo
nic": do zapoczątkowania potencjału czynnościowego niezbędny jest bodziec o intensywności
wystarczającej do zdepolaryzowania neuronu powyżej określonej wartości progowej; wszystkie
potencjały czynnościowe w danej komórce osiągają tę samą amplitudę
[3]
.
Przebieg potencjału czynnościowego w komórce nerwowej
Jeśli dokomórkowe prądy kationów przeważają nad odkomórkowymi, następuje depolaryzacja od
poziomu
potencjału spoczynkowego
(ok. -70 mV) do potencjału krytycznego czyli
progowego
(ok.
-50 mV). Po osiągnięciu potencjału progowego, następuje otwarcie bramkowanych elektrycznie

3
(zależne od napięcia) kanałów przewodzących odkomórkowo
kationy
potasowe oraz kanałów
przewodzących kationy sodowe do wnętrza komórki. Wskutek różnicy stężeń i potencjałów
kationy sodowe szybko napływają do wnętrza komórki, niwelując różnicę potencjałów pomiędzy
środowiskiem zewnątrz- i wewnątrzkomórkowym do zera (depolaryzują błonę komórkową). Jest
to początek tzw. potencjału iglicowego. Ze względu na dużą początkową różnicę stężeń, mimo
wyrównania potencjałów, kationy sodowe napływają w dalszym ciągu do wnętrza komórki,
powodując odwrotną polaryzację błony, tzw. nadstrzał dochodzący do +35 mV. Następuje to
stosunkowo szybko; w ciągu ok. 2 ms następuje już całkowita inaktywacja kanałów sodowych. Po
rozpoczęciu aktywacji sodowej, w odpowiedzi na depolaryzację, następuje aktywacja potasowa,
czyli otwarcie kanałów dla odkomórkowego prądu kationów potasowych. Powoduje to zmianę
potencjału wnętrza komórki z powrotem na ujemny, czyli repolaryzację. W tym czasie nie jest
możliwe ponowne pobudzenie komórki, jest to okres
refrakcji
bezwzględnej
[4]
. W czasie trwania
potencjału iglicowego, a zwłaszcza pod koniec - proporcje kationów po obu stronach błony są
odwrócone (Na
+
wewnątrz, K
+
na zewnątrz komórki). Przez cały czas trwania potencjału
czynnościowego, aktywna jest
pompa sodowo-potasowa
oraz inne układy transportujące jony.
Poziom aktywności pompy sodowo-potasowej jest bowiem uzależniony m. in. od stężenia
kationów sodowych we wnętrzu komórki. Zatem w czasie trwania potencjału iglicowego pompa
działa z najwyższą możliwą prędkością. W ciągu kolejnych kilkudziesięciu milisekund przywraca
proporcje kationów sodowych i potasowych po obu stronach błony komórkowej. Jednak
początkowo błona jest jeszcze w stanie hiperpolaryzacji - różnica potencjałów przekracza wartość
spoczynkową. Jest to okres refrakcji względnej, kiedy komórkę da się pobudzić, ale dużo
większym bodźcem - ze względu na większą "odległość" od poziomu potencjału krytycznego. W
pewnym stopniu, w różnych tkankach, w opisanych procesach biorą udział również inne jony,
głównie chlorkowe (Cl
-
) oraz wapniowe (Ca
2+
).
Potencjał spoczynkowy
Potencjał spoczynkowy to różnica
potencjałów
(
napięcie
) między obiema stronami
błony
plazmatycznej
niepobudzonej komórki pobudliwej.
Wszystkie napięcia na błonie wyraża się jako różnice potencjału wnętrza komórki do potencjału
po stronie zewnętrznej. Potencjały spoczynkowe mają wartości ujemne, które w komórkach
nerwowych wahają się między –65
mV
a –90 mV. Powstanie potencjału spoczynkowego jest
spowodowane przede wszystkim tendencją
jonów
potasu
do przepływania zgodnie
z
gradientem
stężenia
tych jonów z wnętrza na zewnątrz
błony komórkowej
. Powoduje to
pozostanie niewielkiego nadmiaru
ładunków
ujemnych po wewnętrznej stronie błony. Inne jony
(np. sodu) jedynie w niewielkim stopniu wpływają na wartości potencjału spoczynkowego. Siła
elektrochemiczna, powodująca ruch jonu poprzez błonę komórkową, jest różnicą między
potencjałem spoczynkowym a potencjałem równowagi dla danego jonu. Potencjał równowagi
jonu jest to taki potencjał, przy którym wypływ tego rodzaju jonów z komórki jest równy ich
wpływowi do jej wnętrza.
Potencjał równowagi dla poszczególnych jonów można obliczyć stosując
równanie Nernsta
.
Potencjały spoczynkowe można obliczyć stosując
równanie Goldmana
, które uwzględnia
wszystkie zaangażowane rodzaje jonów.
Synapsa
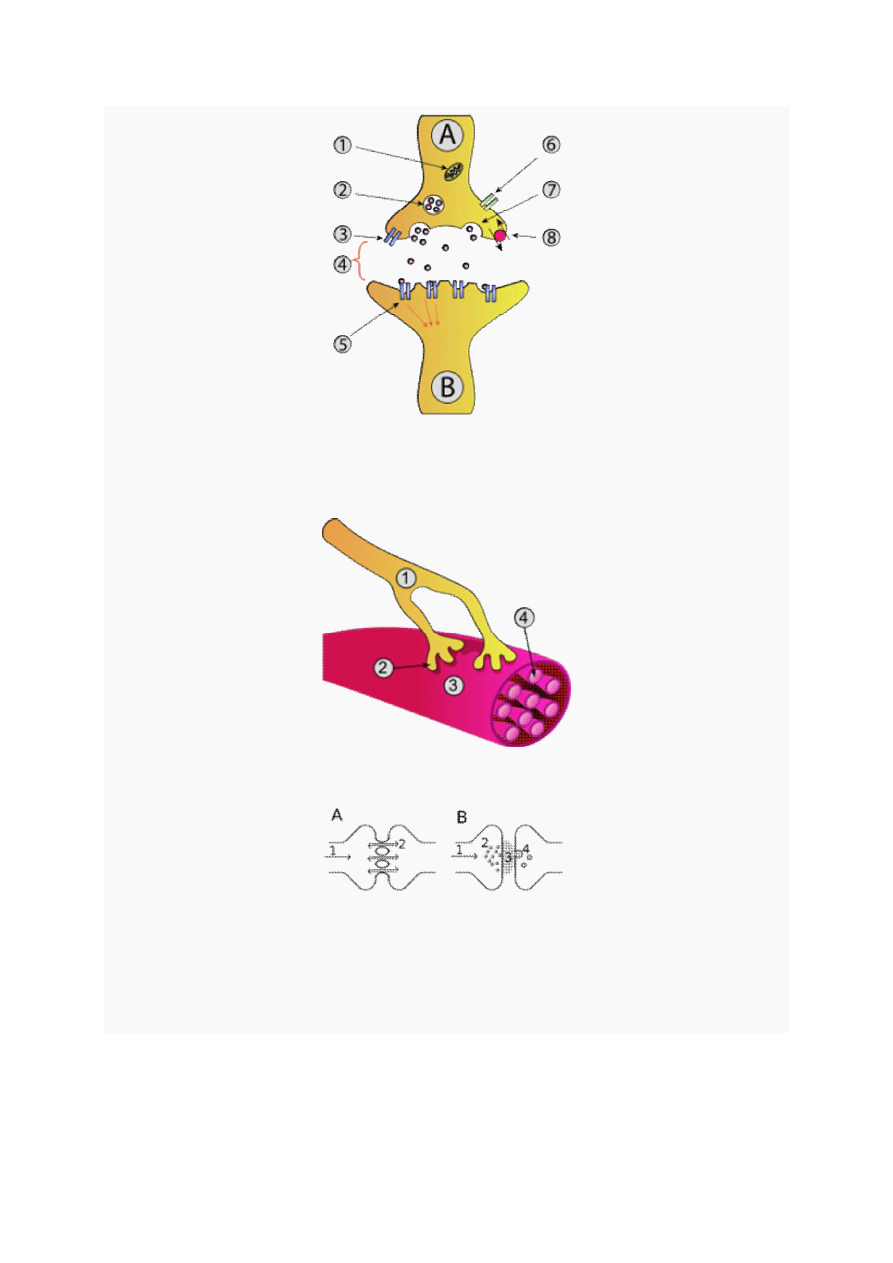
4
Połączenie synaptyczne: 1-
mitochondrium
2-
pęcherzyki
presynaptyczne z
neurotransmiterem
3-
autoreceptor 4-
szczelina synaptyczna
5-neuroreceptor 6-
kanał wapniowy
7-pęcherzyk
uwalniający
neurotransmitery
8-receptor zwrotnego wychwytu mediatora
Połączenie nerwowo-mięśniowe
: 1-
akson
2-połączenie synaptyczne 3-
mięsień szkieletowy
4-
miofibryla
Porównanie synapsy elektrycznej (A) i chemicznej (B); A) pobudzenie (1) jest przekazywane między
komórkami za pomocą transportu jonów (2) przez bezpośrednie
kanały
; B) pobudzenie (1) skutkuje
wydzielaniem pęcherzyków i
egzocytotycznym
wydzielaniem (2) substancji przekaźnikowej (3)
wchłanianej
endocytotycznie
(4) przez komórkę – odbiornik.
Synapsa – miejsce komunikacji błony kończącej
akson
z
błoną komórkową
drugiej komórki -
nerwowej
lub komórki efektora (narządu wykonawczego), np. mięśni lub gruczołu.
Impuls nerwowy
zostaje przeniesiony z jednej komórki na drugą przy udziale
substancji
o
charakterze
neuroprzekaźnika
(zwanego czasem neurohormonem) – mediatora synaptycznego
(synapsy chemiczne) lub na drodze impulsu elektrycznego (synapsy elektryczne). Synapsy

5
otoczone są przez
astrocyty
, które uczestniczą w przekazywaniu sygnału między neuronami.
Powtarzające się przekazywanie impulsu
długotrwale wzmacnia przewodnictwo synaptyczne
.
Powstawanie sieci neuronów o zwiększonym przewodnictwie jest podstawą uczenia się i
zapamiętywania
[1]
.
Nowe synapsy powstają w procesie zwanym
synaptogenezą
.
Połączenia synaptyczne
Ze względu na rodzaj komórek, między którymi przekazywany jest sygnał, wyróżnia się synapsy:
nerwowo-nerwowe – połączenie między dwiema komórkami nerwowymi
nerwowo-mięśniowe – połączenie między komórką nerwową i mięśniową
nerwowo-gruczołowe – połączenie między komórką nerwową i gruczołową
Typy synaps
Ze względu na sposób przekazywania impulsu wyróżnia się synapsy elektryczne i chemiczne.
Elektryczne
W tych synapsach neurony prawie się stykają (gł. połączenia typu „neksus”). Kolbka
presynaptyczna oddalona jest od kolbki postsynaptycznej o 3,5
nm
. Możliwa jest
wędrówka
jonów
z jednej komórki do drugiej – przekazywanie dwukierunkowe. Impuls jest bardzo
szybko przekazywany. Występują w mięśniach, siatkówce oka, części korowej
mózgu
oraz
niektórych częściach serca.
Chemiczne
W tych synapsach komórki są od siebie oddalone o ok. 20 nm, między nimi powstaje
szczelina
synaptyczna
. Zakończenie neuronu presynaptycznego tworzy kolbkę synaptyczną, w której są
wytwarzane neuroprzekaźniki (mediatory – przekazywane w pęcherzykach synaptycznych), które
łączą się z receptorem, powodując depolaryzację błony postsynaptycznej. Występują tam, gdzie
niepotrzebne jest szybkie przekazywanie impulsu, np. w narządach wewnętrznych.
Obecnie znanych jest ok. 60 związków, które pełnią funkcję mediatorów. Wyróżnia się wśród nich
neuroprzekaźniki pobudzające lub hamujące wzbudzanie potencjału czynnościowego. Głównym
neuroprzekaźnikiem pobudzającym w korze mózgowej ssaków jest
kwas glutaminowy
. Inne
znane mediatory pobudzające to
np.:
acetylocholina
,
noradrenalina
,
adrenalina
,
dopamina
,
serotonina
,
histamina
. Głównym
neuroprzekaźnikiem hamującym jest
kwas γ-aminomasłowy
(GABA). Pozostałe mediatory
hamujące to:
glicyna
i peptydy
opioidowe
.
Synapsa nerwowo-mięśniowa
Przez synapsę nerwowo-mięśniową następuje przekazanie sygnału z
motoneuronu
do
mięśnia
szkieletowego
. W pobliżu komórki mięśniowej neuron traci
osłonkę mielinową
i rozdziela się na
wiele cienkich odgałęzień, które kontaktują się z błoną komórki mięśniowej (błoną
postsynaptyczną). W miejscach styczności na końcówkach nerwu (błonie presynaptycznej)
znajdują się
kolbki synaptyczne
zawierające pęcherzyki z
neurotransmiterem
acetylocholiną
(ACh). Przestrzeń między błoną pre- i postsynaptyczną to
przestrzeń synaptyczna.
W błonie presynaptycznej są strefy aktywne, w których zachodzi
egzocytoza
pęcherzyków z
neurotransmiterem i uwolnienie ACh do przestrzeni synaptycznej. W tej błonie mieszczą się
także
kanały wapniowe
typu N.
W błonie postsynaptycznej, naprzeciw stref aktywnych, tworzą się pofałdowania synaptyczne. Na
ich krawędziach znajdują się receptory acetylocholiny typu synaptycznego, które są kanałami
jonowymi otwierającymi się w wyniku przyłączenia ACh.

6
Schemat działania synapsy chemicznej
Gdy impuls nerwowy dotrze do zakończenia aksonu, powoduje otwarcie kanałów jonowych
selektywnie wpuszczających jony wapnia. Jony te uaktywniają
migrację
pęcherzyków
presynaptycznych (zawierających
mediator
– substancję chemiczną,
np.
adrenalinę
,
noradrenalinę
,
acetylocholinę
). Na drodze
egzocytozy
pęcherzyki te uwalniają
zawartość do
szczeliny synaptycznej
.
Mediator wypełnia szczelinę synaptyczną i część z jego cząsteczek łączy się z receptorami na
błonie postsynaptycznej. Powoduje to otworzenie się kanałów dla jonów sodu, a w efekcie
depolaryzację błony postsynaptycznej. Jeżeli depolaryzacja ta osiągnie wartość progową,
otwierają się kolejne kanały sodowe wrażliwe na napięcie, skutkiem czego pojawia się
potencjał
czynnościowy
i indukuje falę przechodzącą przez cały neuron.
Cząsteczki mediatora działają w szczelinie synaptycznej jedynie przez określony czas. Jest to
spowodowane istnieniem receptorów na błonie presynaptycznej, które zajmują się zwrotnym
wychwytem (re-uptake) mediatora.
Narządy ścięgniste Golgiego
Narządy ścięgniste Golgiego – rodzaj
receptorów
miotatycznych zlokalizowanych w ścięgnie w
pobliżu jego przejścia w tkankę mięśniową. Odpowiadają za
objaw scyzoryka
w spastyczności.
Narządy ścięgniste są niewrażliwe na pasywne rozciąganie, a ich aktywację powoduje skurcz
sąsiadujących
komórek mięśniowych
. Gdy włókna mięśnia szarpią, organy Golgiego
emitują
impulsy nerwowe
do
rdzenia kręgowego
. To powoduje rozluźnienie mięśnia i umożliwia
utrzymanie określonej pozycji ciała. Narządy ścięgniste pełnią także funkcję ochronną,
zapobiegając oderwaniu się mięśni od miejsca przyczepu (wskutek np. zbyt silnego napięcia).
Budowa
Pojedynczy narząd składa się z wiązki włókien ścięgnowych zamkniętych w torebce włóknistej.
Do narządu wnika jedno lub dwa włókna dośrodkowe należące do grupy Ib, o średnicy ok. 16
μm
[
Odruch bezwarunkowy
Odruch bezwarunkowy – reakcja wrodzona (
odruch
), automatyczna, zachodzi poprzez
pobudzenie odpowiednich
receptorów
, zakończeń nerwowych, nerwów czuciowych oraz
pobudzenie organów efektorowych (głównie mięśni) poprzez nerwy ruchowe lub autonomiczne.
Reakcja odruchowa przebiega bez uświadomienia, to znaczy, że nerwy wywołują odruch
(pobudzają mięśnie) przed powiadomieniem mózgu.
Przykłady odruchów bezwarunkowych
odruch akomodacji oka
odruch krztuśny
(
ang.
gag reflex)
odruch nurkowania
(ang. Mammalian diving reflex)
odruch przedsionkowo-oczny
(ang. Vestibulo-ocular reflex)
odruch ścięgna Achillesa
odruch rzepkowy
, inaczej
odruch kolanowy
(ang. Patellar reflex)
odruch źreniczny
odruch wymiotny

7
Odruchy monosynaptyczne
Są przykładem odruchów bezwarunkowych, których realizacja odbywa się na poziomie rdzenia
kręgowego, z wykorzystaniem tylko dwóch neuronów.
Odruch rozciągania mięśnia
Każdy mięsień poprzecznie prążkowany posiada oprócz włókien mięśnia roboczego, również tzw.
wrzeciona nerwowo-mięśniowe. Te wyspecjalizowane receptory odruchu rozciągania mięśnia,
ledwie widoczne gołym okiem, zlokalizowane są w brzuścu mięśnia.
Wrzecionka nerwowo-
mięśniowe
posiadają budowę podobną do włókna mięśnia roboczego, są jednak od nich cieńsze.
W obrębie wrzecionka znajduje się sieć cienkich dośrodkowych włókien nerwowych, tzw.
zakończeń pierścieniowo-spiralnych, które bardzo łatwo reagują na rozciąganie. W odpowiedzi
na bodziec, generowany jest potencjał czynnościowy, który przy pomocy grubego włókna typu Ia
wędruje przez korzeń tylny do istoty szarej rdzenia. Tam sygnał przełącza się przez synapsę na
komórkę ruchową przedniego rogu rdzenia zaczynając impuls nerwowy w neuronie eferentnym
(grube włókno nerwowe typu α). Komórka nerwowa eferentna ma bogate rozgałęzienia do
włókien roboczych mięśnia. Impuls wywołuje reakcję skurczową całego mięśnia (wszystkich
włókien), w efekcie zmienia się stan napięcia wrzecionowych komórek mięśniowych i do układu
nerwowego kierowana jest informacja zwrotna. Monosynaptyczny odruch rozciągania mięśnia
stanowi podstawę m.in. odruchu rzepkowego, ze ścięgnem Achillesa oraz odruchu ze ścięgna
mięśnia dwugłowego ramienia.
Odruch rzepkowy działa następująco: uderzenie w ścięgno rzepkowe rozciąga wrzeciona
mięśniowe w mięśniu czworogłowym z przodu uda. To rozciągnięcie jest szybkie jak mgnienie
oka, lecz mimo to stymuluje konkretne neurony czuciowe, których dendryty mają zakończenia we
wrzecionach mięśnia i których aksony mają zakończenia bezpośrednio w neuronach ruchowych
w rdzeniu kręgowym. Nerwy eferentne stymulują mięsień czworogłowy uda, powodując jego
skracanie się i podskakiwanie stopy. Odruch ścięgnisty jest specyficzny w tym, iż jego sprzężenie
zwrotne dotyczy jedynie mięśnia, w którym wrzeciono jest zlokalizowane. "Podobnie jak
wszystkie inne odruchy, ten również zachodzi na ułamek sekundy zanim zdacie sobie z niego
świadomie sprawę. Czujecie, że zachodzi, już po fakcie, po tym jak odruch zakończył swój cykl.
Odnotowujecie też to wrażenie świadomie wyłącznie dlatego, iż oddzielne receptory modalności
dotyku przesyłają wiadomość do kory mózgowej i tą drogą do świadomego umysłu."
Objaw scyzoryka w spastyczności
Objaw scyzoryka w spastyczności jest kolejnym odruch rozciągania, lecz powoduje on raczej
rozluźnianie niż skurcz mięśni docelowych. Bodźcem dla objawu nie jest dynamiczne rozciąganie
wrzeciona mięśnia, lecz ściskanie kurczliwy nacisk na receptory czuciowe ścięgna. Nacisk ten
odruchowo powoduje rozluźnienie mięśnia przyczepionego do ścięgna, oraz zapięcie stawu.
Receptory czuciowe odbjawu scyzoryka w spastyczności znajdują się w organie ścięgnistym
Golgiego. Większość receptorów jest tak naprawdę zlokalizowanych blisko połączeń mięśniowo-
ścięgnistych, gdzie łączą one niewielkie kawałeczki tkanki łącznej, z odpowiadającymi im
włóknami mięśniowymi. Organ ścięgnisty Golgiego jest więc aktywowany przez kurczenie
komórek mięśni, które ułożone są w jednej linii (w szeregu) z receptorami. Najnowsze badania
wyjaśniły, że organ ścięgnisty Golgiego jest stosunkowo niewrażliwy na pasywne rozciąganie,
lecz zaczyna wystrzeliwywać impulsy nerwowe z powrotem do rdzenia kręgowego, kiedy tylko
włókna mięśnia zaczynają go szarpać. Inaczej niż w przypadku odruchu ścięgnistego, tutaj
przychodzące aksony czuciowe nie mają zakończeń bezpośrednio w neuronach ruchowych, (co
mogłoby zwiększyć ich aktywność i stymulować skurcz mięśniowy), lecz w hamujących
neuronach wstawkowych, które zmniejszają aktywność neuronów ruchowych i tym samym
powodują odprężanie się mięśnia. Jeśli stymulujecie receptor, odruch rozluźnia mięsień.
Odruch polisynaptyczny
Jest to odruch, którego szlak przebiega przez wiele neuronów przełącznikowych. Jego tor ulega
rozgałęzieniu, a na jego przebieg i efekt końcowy mogą mieć wpływ różne ośrodki mózgowia i
rdzenia kręgowego.

8
Odruchy zginania
Inaczej nazywany odruchem ucieczki lub odruchem obronnym. Powstaje on w przypadku
zaistnienia bodźca bólowego (np. dotknięcie gorącej patelni, nastąpienie na ostry kamień, ukłucie
kolcem). Podrażniony receptor bólowy wysyła sygnał do rdzenia kręgowego, przez korzenie
tylne. Tam zostaje on przełączony na neurony różnych dróg aferentnych i eferentnych.
Docierające do mięśni impulsy powodują gwałtowny skurcz tych grup mięśniowych, których
działanie jest konieczne do oddalenia się od niebezpiecznego czynnika. Równocześnie
rozluźnieniu ulegają mięśnie antagonistyczne. W przypadku bodźca pochodzącego z kończyny
dolnej, ciężar ciała przenoszony jest automatycznie na drugą kończynę. Nad zachowaniem
równowagi czuwają ośrodki mózgowia,
móżdżku
i
pnia mózgu
. Odruchy zginania są pochodzenia
rdzeniowego, a nie mózgowego. Tak więc nawet gdyby rdzeń kręgowy został odcięty od mózgu,
odruch zginania nadal cofnąłby rękę lub stopę przed zagrażającym bodźcem.
Wzajemne hamowanie
Odruchy zginania nie tylko powodują aktywizację mięśni zginacza, do przyciągnięcia dłoni lub
stopy w stronę tułowia, lecz również rozluźniają one prostownik, który następnie pozwala na
swobodne zgięcie. Dzieje się tak za sprawą neuronów wstawkowych hamujących. Kiedy neurony
wstawkowe ułatwiające oddziałują na neurony ruchowe unerwiające zginacze, w ten sposób
powodując ich skurcz, neurony wstawkowe hamujące oddziałują na neurony ruchowe
unerwiające prostowniki, powodując ich rozluźnianie. Ten fenomen jest znany jako wzajemne
hamowanie i jest on integralną częścią odruchu zginania. Inaczej niż odruchy rozciągania,
odruchy zginania powodują efekty znacznie wykraczające poza strefę oddziaływania bodźca.
Można zaobserwować to, kiedy pielęgniarka nakłuwa palec wskazujący dziecka, aby pobrać
krew. Reaguje cała górna kończyna dziecka, a nie tylko zginacze podrażnionego palca.
Gwałtowne szarpnięcie do tyłu wskazuje, iż odruch zginania ułatwia neurony ruchowe zginacza i
powstrzymuje neurony ruchowe całej górnej kończyny.
Odruch skrzyżowanego wyprostu
Odruch skrzyżowanego wyprostu jest kolejnym składnikiem odruchów zginania wspomaga
przeciwną stronę ciała. Za pomocą tego odruchu, podczas gdy członek z podrażnionej strony
zgina się, członek po drugiej stronie się rozciąga. Dzieje się tak, kiedy człowiek staje jedną stopą
na żarze w pobliżu ognia. Nie myśląc o tym, podnosi podrażnioną stopę w pośpiechu, skracając
zginacze i rozluźniając prostowniki po tej samej stronie ciała – wszystko w jednym momencie –
paluch, kostka, kolano, biodro, a nawet tułów. Podczas gdy podrażniona stopa unosi się do góry,
odruch skrzyżowanego wyprostu skraca prostowniki i rozluźnia zginacze po przeciwnej stronie
ciała, wzmacniając jego postawę i powstrzymując od przewrócenia się w ogień. Odruch
skrzyżowanego wyprostu jest realizowany przez neurony wstawkowe, których aksony krzyżują na
przeciwległą stronę rdzenia kręgowego i unerwiają neurony ruchowe w odwrotny sposób niż ten
po stronie podrażnionej – neurony ruchowe mięśni prostownika są ułatwiane, zaś neurony
ruchowe mięśni zginacza są powstrzymywane. Odruchy zginania służą również wielu innym
funkcjom ochronnym. Na przykład podczas poważnego skręcenia kostki zakończenia bólowe
stają się bardziej wrażliwe. Przy kolejnym skręceniu kostki, wyższe centra w mózgu skojarzone z
wolą i świadomością pozwolą odruchom kręgosłupowego zginania na niepowstrzymane działanie
i ciało upadnie, zanim jego waga całkiem przygniecie narażony na niebezpieczeństwo staw.
Odruchy u noworodków

9
Odruch chwytny u noworodka
odruch płaczu.
odruchy: szukania i ssania, najlepiej rozwinięte odruchy noworodka. Po pogłaskaniu w
policzek dziecko natychmiast odwróci główkę w jego stronę i otworzy usta w poszukiwaniu
jedzenia; nawet jeśli odnajdzie tylko palec, zacznie go mocno ssać.
odruch Moro (nazwa pochodzi od nazwiska badacza, który go opisał), inaczej zwany
odruchem obejmowania. Gdy gwałtownie zmienimy położenie ciała noworodka, albo też w
pomieszczeniu, w którym się znajduje, rozlegnie się nagły hałas lub ostry dźwięk, dziecko
zareaguje energicznym wyprostowaniem rąk i nóg. Plecy wygnie w łuk, a głowę odchyli do
tyłu; następnie zaciśnie pięści, a odrzuconymi wcześniej na boki rękami wykona powolny
ruch objęcia klatki piersiowej.
odruch chwytny - można zaobserwować go, wkładając palec w dłoń noworodka; zaciska
on wówczas palce tak mocno, że można go nawet unieść. Dotyczy to każdego przedmiotu
przyłożonego do wewnętrznej strony dłoni noworodka.
odruch pełzania - jeśli ułożonemu na brzuchu noworodkowi delikatnie podrażnić stopy,
zacznie on nieznacznie czołgać się do przodu.
odruch podparcia i stąpania, inaczej zwany odruchem chodu automatycznego. Jeśli
chwycimy nowo narodzone dziecko pod pachy, tak aby jego stopy dotykały podłoża, zacznie
ono przebierać nogami, tak jakby chciało chodzić. Odruch ten nie ma jednak nic wspólnego z
właściwym chodzeniem.
Powyższe odruchy charakterystyczne są tylko dla okresu noworodkowego i zanikają w miarę
upływu czasu - odruch Moro po około pięciu miesiącach życia, chwytny po ok. trzech, a chodu
automatycznego po ok. dwóch miesiącach.
Wrzecionko nerwowo-mięśniowe
Wrzecionko nerwowo-mięśniowe

10
Wrzecionko nerwowo-mięśniowe składa się z 2-10 cienkich komórek mięśniowych -
miocytów
.
Miocyty te nazwano
miocytami intrafuzalnymi
i podzielono na dwie grupy, które różnią się
ułożeniem
jąder
, średnicą i długością komórek mięśniowych (miocytów). Są to receptory typu
tonicznego, oznacza to, iż przy stałym obciążeniu danego
mięśnia
częstotliwość wyładowań jest
stała. Wzrost rozciągnięcia tego mięśnia zwiększa częstotliwość wyładowań, natomiast
zwiększenie obciążenia zwiększa liczbę
receptorów
wyładowujących się. Wrzecionko posiada
bardzo niski
próg pobudliwości
, ponieważ już obciążenie mięśnia masą 1-2 g wywołuje
utrzymujące się wyładowanie wrzecionek. Wrzecionka ułożone są równolegle do
miocytów
ekstrafuzalnych
. Wrzecionko otacza torebka łącznotkankowa, która po obu stronach przyczepia
się do
śródmięsnej
miocytów ekstrafuzalnych. Obwodowe części miocytów w przeciwieństwie do
tych położonych w środkowej części wrzecionka są poprzecznie prążkowane i mają zdolność
kurczenia się.
Podział miocytów intrafuzalnych
miocyty z woreczkiem jąder
miocyty z łańcuszkiem jąder
Unerwienie wrzecionka nerwowo-mięśniowego
Grube włókna nerwowe dośrodkowe, należące do grupy Ia, które po przejściu przez
torebkę tracą osłonkę mielinową i kończą się w środkowej części wrzecionka w postaci
zakończeń pierścieniowo-spiralnych – pierwotnych, występują w obydwu typach miocytów.
Cieńsze włókna dośrodkowe, należące do grupy II, kończą się zakończeniami w postaci
„wiązanki kwiatów” – wtórnymi. Umiejscowione są obwodowo, występują jedynie w miocytach
z łańcuszkiem jąder.
Cienkie włókna nerwowe odśrodkowe, należące do grupy A-gamma, kończą się płytkami
ruchowymi na miocytach intrafuzalnych w częściach obwodowych z poprzecznym
prążkowaniem. Dzielimy je na dwa typy:
włókna odśrodkowe gamma-dynamiczne – włókna te znajdują się
w
miocytach
z torebką jąder,
włókna odśrodkowe gamma-statyczne - natomiast te włókna występują w
miocytach z łańcuszkiem jąder.
Pamięć
Pamięć – zdolność do rejestrowania i ponownego przywoływania
wrażeń zmysłowych
,
skojarzeń,
informacji
, występująca u ludzi, niektórych
zwierząt
i w
komputerach
. W każdym z tych
przypadków proces zapamiętywania ma całkowicie inne podłoże
fizyczne
oraz podlega badaniom
naukowym w oparciu o różne zestawy pojęć.
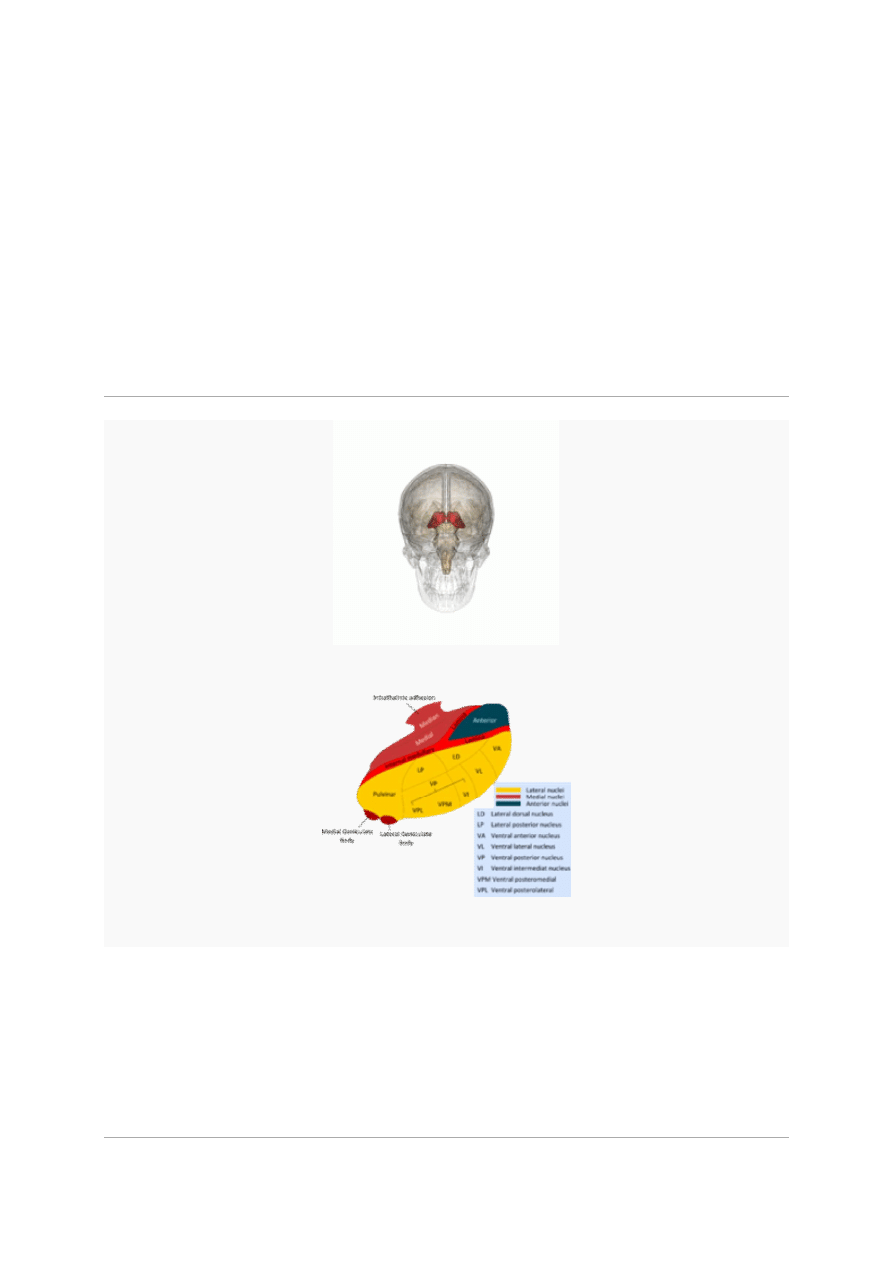
Podwzgórze
Podwzgórze
hypothalamus
hypothalamus

11
lokalizacja podwzgórza w mózgowiu
Narządy
Mózgowie
Podwzgórze zaznaczone w dwóch projekcjach mózgu
Przekrój w płaszczyźnie strzałkowej mózgowia płodu w wieku 3 miesięcy, zaznaczono podwzgórze
Podwzgórze (
łac.
hypothalamus, z
gr.
ὑπó = „pod” θάλαμος „izba, sypialnia”) – część
podkorowa
mózgowia
zaliczana do
międzymózgowia
. Podwzgórze składa się z wielu jąder
spełniających szereg istotnych funkcji.
Podwzgórze produkuje i wydziela neurohormony i w ten sposób funkcjonalne łączy
ośrodkowy
układ nerwowy
z
układem wewnątrzwydzielniczym
, poprzez
przysadkę mózgową
.

12
Podwzgórze reguluje liczne procesy metaboliczne oraz wiele funkcji
autonomicznego
i
ośrodkowego układu nerwowego.
Podwzgórze kontroluje
głód
pragnienie
rytm dobowy
sen
temperaturę ciała
zachowania rodzicielskie.
Podwzgórze jest częścią mózgu wszystkich
kręgowców
[1]
.
Anatomia
Podwzgórze znajduje się między podstawną częścią
kresomózgowia
i brzuszną
częścią
śródmózgowia
. Podwzgórze jest ściśle połączone z przysadką mózgową. Podwzgórze i
przysadka stanowią część
osi podwzgórze-przysadka-nadnercza
.
W przedniej części podwzgórza znajdują się jądra:
płciowodwupostaciowe
nadwzrokowe
przykomorowe
nadskrzyżowaniowe
W części środkowej (guzowej) znajdują się jądra:
brzuszno-przyśrodkowe
grzbietowo-przyśrodkowe
guzowo-suteczkowe
łukowate
W części bocznej znajduje się jądro:
boczne
W części tylnej (sutkowatej) znajdują się jądra:
suteczkowe przyśrodkowe
suteczkowe boczne
przedsuteczkowe
tylne
Neurotransmitery podwzgórza
Neurony syntetyzują około 20 ważnych związków o
właściwościach
hormonów
lub
neuroprzekaźników
.
Funkcja
Podwzgórze zawiera ważne ośrodki kierujące czynnością autonomicznego układu nerwowego,
gospodarką wodną organizmu (regulacją ilości wody i odczuwaniem pragnienia),
termoregulacją
,
czynnością gruczołów wewnątrzwydzielniczych, pobieraniem pokarmu (głód i sytość), przemianą
tłuszczów, przemianą węglowodanów (cukrów), snem i czuwaniem, czynnościami seksualnymi
(cyklami układu rozrodczego, popędem seksualnym) oraz reakcjami emocjonalnymi. Czynność
podwzgórza pozostaje w ścisłym związku z
przysadką mózgową
.
13
Podwzgórze, które jest ośrodkiem preferencji seksualnych w mózgu, pełni odmienne funkcje u
kobiet niż u mężczyzn. U mężczyzn reguluje ono przepływ hormonów w taki sposób, żeby
zapewnić utrzymanie stałego ich poziomu. U kobiet podwzgórze reaguje na wysoki poziom
hormonu powodując dalsze jego wydzielanie
[2]
[
potrzebne źródło
]
.
Podwzgórze stanowi wierzchołek trójkąta łączący bezpośrednio biochemiczny kanał łączności z
kanałami nerwowymi. Tą drogą wszelkiego rodzaju procesy psychiczne mogą wpływać na
zmianę funkcji biochemicznych organizmu żywego. Szczególną pozycję w łączności mózgu z
narządami wewnętrznymi zajmują aminy katecholowe:
adrenalina
i noradrenalina.
Na podstawie doświadczeń na szczurach stwierdzono, że w podwzgórzu zlokalizowany jest
ośrodek przyjemności. Przez podwzgórze do kory nerwowej przepływają bodźce czuciowe,
dlatego podwzgórze nazywane jest podkorowym ośrodkiem czucia.
Wzgórze (anatomia)
Animacja ilustrująca położenie wzgórza w obrębie mózgowia i czaszki człowieka
Schemat budowy wzgórza
Wzgórze (
łac.
thalamus,
ang.
thalamus) - część
międzymózgowia
znajdująca się pod
spoidłem
wielkim
. Przylega do niego
podwzgórze
.
Odpowiada za wstępną ocenę
bodźców
zmysłowych
i przesyłanie ich do kory mózgowej
(oprócz
węchu
). Pełni kluczową rolę w
integracji informacji
czuciowych i ruchowych, w procesach
uwagi i kontrolowania dostępu do danych czuciowych. Stanowi największą ilość
istoty
szarej
międzymózgowia. Wzgórze tworzą skupienia istoty szarej (jądra) pooddzielane przez
pasma
istoty białej
. Stanowi ograniczenie
komór
: komory bocznej oraz komory trzeciej.
Budowa zewnętrzna

14
W opisie anatomicznym wzgórza wyróżnia się dwa bieguny i cztery powierzchnie.
Biegun przedni - jest skierowany w stronę ogona
jądra ogoniastego
i wraz ze
słupem
sklepienia
ogranicza otwór międzykomorowy (foramen interventriculare).
Biegun tylny - tworzy poduszkę (pulvinar).
Powierzchnia górna - stanowi ograniczenie części centralnej komory bocznej. Jest
wypukła, brzegiem bocznym graniczy z
jądrem ogoniastym
. Między nimi przebiega bruzda w
której przebiega prążek krańcowy (stria terminalis). Prążek krańcowy jest zrośnięty z cienką
blaszką, zwana blaszką przytwierdzoną, do której przyczepia się splot naczyniówkowy
komory bocznej. Wzdłuż powierzchni przyśrodkowej przebiega prążek rdzenny wzgórza, do
którego przyczepia się tkanka naczyniówkowa komory trzeciej.
Powierzchnia przyśrodkowa - współtworzy boczną ścianę komory trzeciej. Jest ustawiona
wzdłuż osi strzałkowej. Powierzchnie przyśrodkowe są zrośnięte, stanowiąc zrost
miedzywzgórzowy.
Powierzchnia dolna - przylega bezpośrednio do
podwzgórza
i
niskowzgórza
Powierzchnia boczna - przylega do jądra ogoniastego i
torebki wewnętrznej
Budowa wewnętrzna, obszary wzgórza i funkcje
Wzgórze jest zbudowane głównie z istoty szarej, tworzącej jądra, do której wnikają pasma istoty
białej, zwanymi blaszkami rdzennymi. Wzdłuż bocznej powierzchni wzgórza przebiega
blaszka
rdzenna zewnętrzna
, która oddziela leżące bocznie od niej jądro siatkowate wzgórza od
pozostałych jąder.
Blaszka rdzenna wewnętrzna
przebiega strzałkowo, oddzielając jądro
przyśrodkowe od jąder bocznych. Ku przodowi rozdziela się ograniczając jądra przednie. Z tyłu
blaszka zagina się, oddzielając jądro przyśrodkowe od jąder poduszki. Wewnątrz blaszki znajduje
się jądro śródblaszkowe.
Wyszukiwarka
Podobne podstrony:
Transmisja impulsów nerwowych podsumowanie
Choroby nerwowo-miesniowe, MEDYCYNA - ŚUM Katowice, V ROK, Neurologia, Materiały dodatkowe
materiały dodatkowe leśna
zadania - stężenia, Notatki i materiały dodatkowe, Chemia, materiały od Romka
PRZYSZLOSC KOMPUTEROW, ^v^ UCZELNIA ^v^, ^v^ Pedagogika, promocja zdrowia z arteterapią i socjoterap
Procedura Dopuszczenia Do Obrotu, STUDIA - Kierunek Transport, STOPIEŃ I, MATERIAŁY DODATKOWE
materiały dodatkowe sedymentologia ćw II0001
Cw 5 Struktury Danych Materiały dodatkowe
credo, prezentacje, WSZYSTKIE PREZENTACJE, OAZA, Prezentacje cd, Prezentacje, Prezentacje na OND III
Prpgram Razem bezpieczniej, Bezpieczeństwo narodowe - UAM Poznań, I rok (2012-2013), Teoria Bezpiecz
Wędrowcy - opowiadanie, Materiały dodatkowe
Krasnoludy w Imperium, Materiały dodatkowe
O giełdzie, STUDIA, studia materiały, MATERIAŁY DODATKOWE, dodatkowe INNE
TEMPERAMENT OSOBOWOSC, ^v^ UCZELNIA ^v^, ^v^ Pedagogika, promocja zdrowia z arteterapią i socjoterap
mimika twarzy - opis, STUDIA - Kierunek Transport, STOPIEŃ I, MATERIAŁY DODATKOWE
więcej podobnych podstron