
SPALANIE KWASÓW
TŁUSZCZOWYCH
NIEPARZYSTYCH I
ROZGAŁĘZIONYCH
Autor : Bartłomiej Stanula

Utlenianie kwasów tłuszczowych w
mitochondriach prowadzi do wytworzenia dużej
ilości ATP - w procesie nazywanym Beta-
oksydacją kwasy tłuszczowe są rozkładane do
wielu cząsteczek acetylo-CoA, który jest
następnie utleniany w cyklu kwasu cytrynowego
dostarczając kolejnych ATP .
Każda runda utleniania kwasów tłuszczowych
przebiega z udziałem pochodnych acylo-CoA ,
jest katalizowana przez kilka enzymów, zachodzi
z wykorzystaniem NAD+ i FAD jako koenzymów.
Jest to proces aerobowy (wymaga obecności
tlenu)

Kwasy tłuszczowe są transportowane we krwi w postaci wolnych kwasów
tłuszczowych .
Wolne kwasy tłuszczowe (WKT ; FFA, free fatty acids) , określane również jako NEFA
(nonesterified fatty acids) to kwasy tłuszczowe występujące w postaci
niezestryfikowanej .
Kwasy tłuszczowe o dłuższych łańcuchach tworzą kompleksy z albuminą, w
komórce natomiast związane są z białkiem wiążącym kwasy tłuszczowe .
Kwasy tłuszczowe o krótkich łańcuchach są lepiej rozpuszczalne w wodzie i mogą
występować w formie niezjonizowanej lub w postaci anionów .
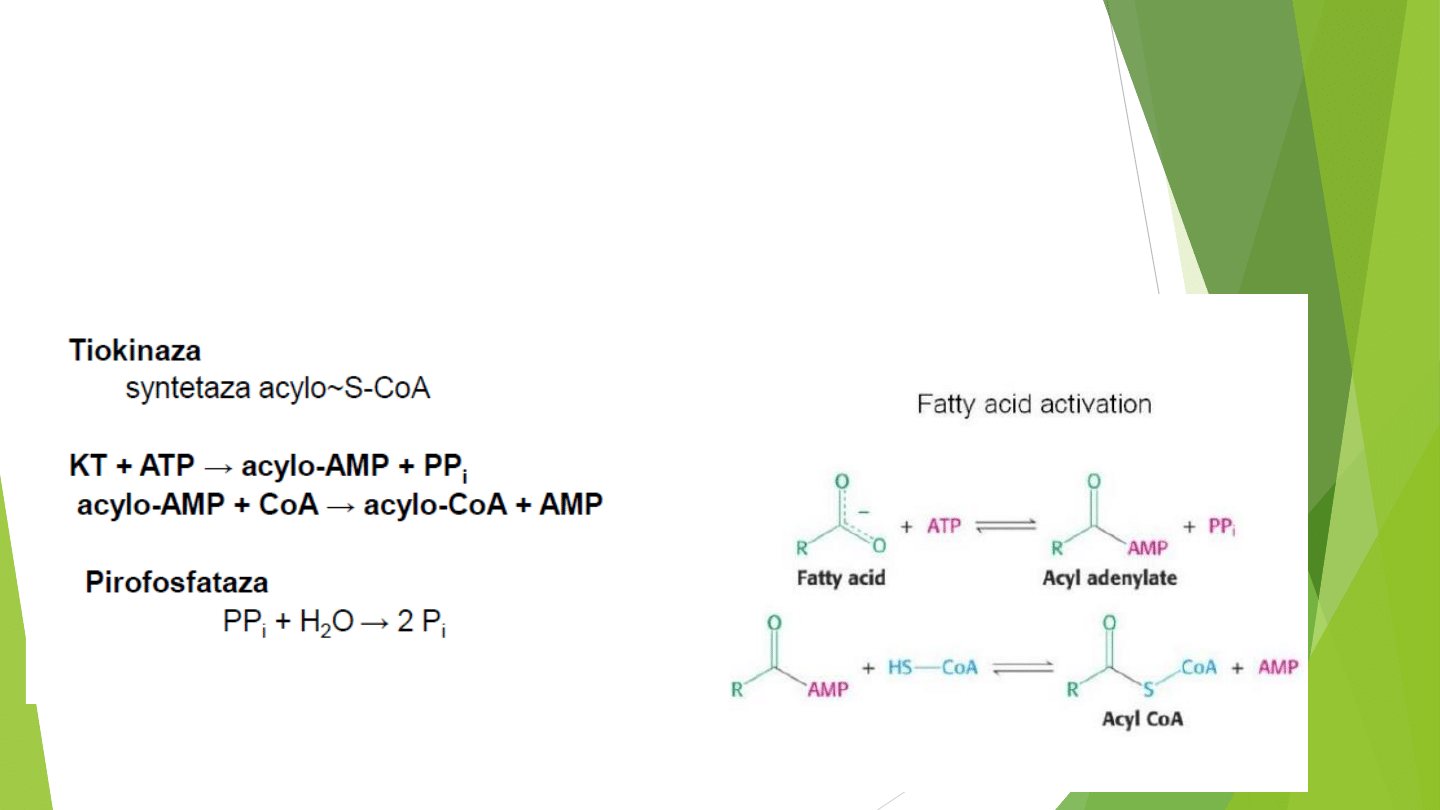
AKTYWACJA KWASÓW TŁUSZCZOWYCH
Kwasy tłuszczowe przed wprowadzeniem w szlaki przemian katabolicznych muszą
zostać przekształcone do inermediatów.
Jest to jedyny etap w rozkładu kwasów tłuszczowych który wymaga dostarczenia
energii w postaci ATP .

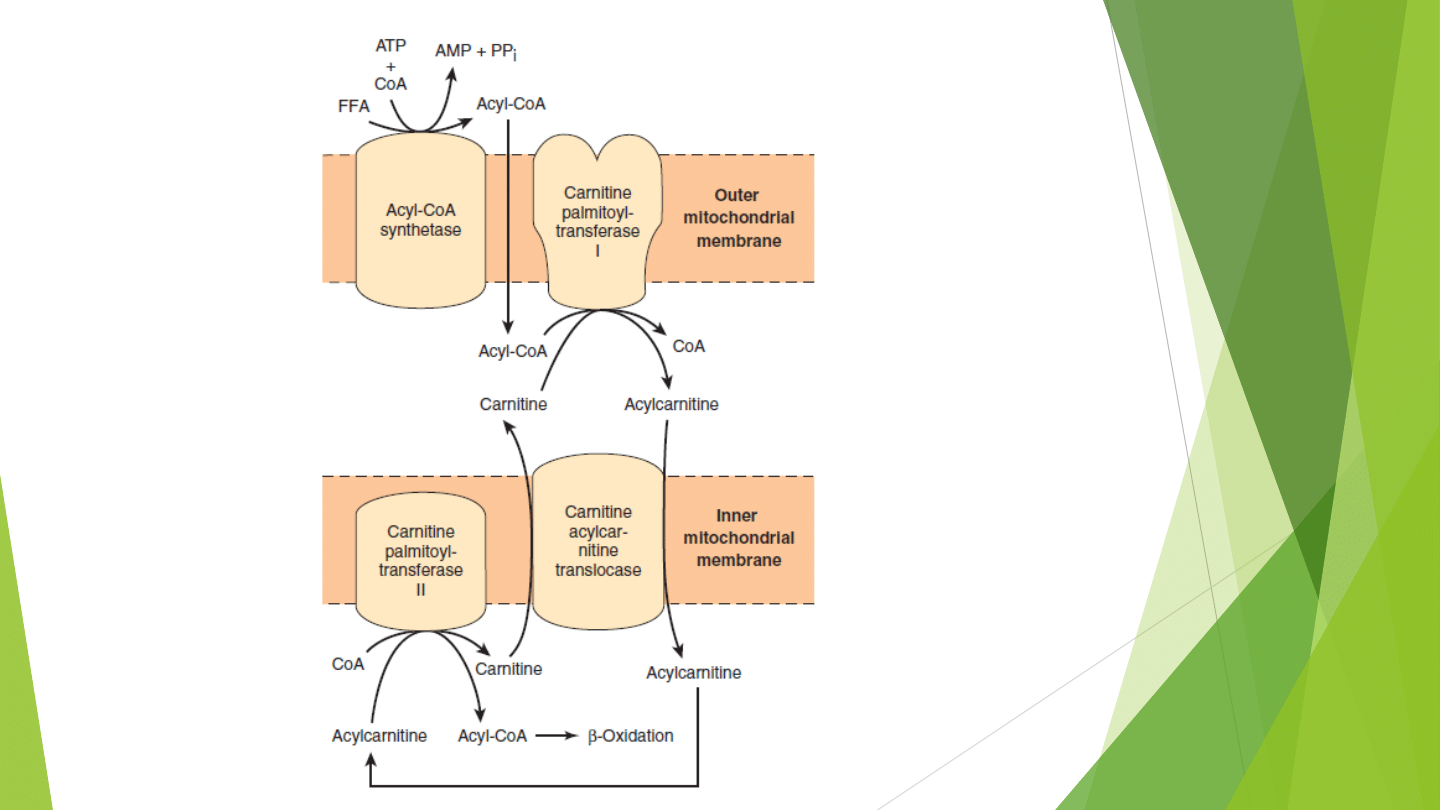
ROLA KARNITYNY W TRANSPORCIE
DŁUGOŁAŃCUCHOWYCH KWASÓW TŁUSZCZOWYCH PRZEZ
WEWNĘTRZNĄ BŁONĘ MITOCHONDRIALNĄ
W systemie transportu aktywnych acyli przez karnitynę
uczestniczą:
o
Palmitoilo-transferaza karnitynowa I (CPT I)
o
Palmitoilo-transferaza karnitynowa II (CPT II)
o
Translokaza karnityna-acylo-karnityna
CPT I występują w zewnętrznej błonie mitochondrialnej .
CPT II oraz translokaza karnityna-acylo-karnityna są
umieszczone w wewnętrznej błonie mitochondrialnej (CPT
II po jej wewnętrznej stronie)
Syntetaza acylo-Coa (tiokinazy) występują w siateczce
śródplazmatycznej, peroksysomach, a także wewnątrz
mitochondriów i na zewnętrznej błonie mitochondrialnej .
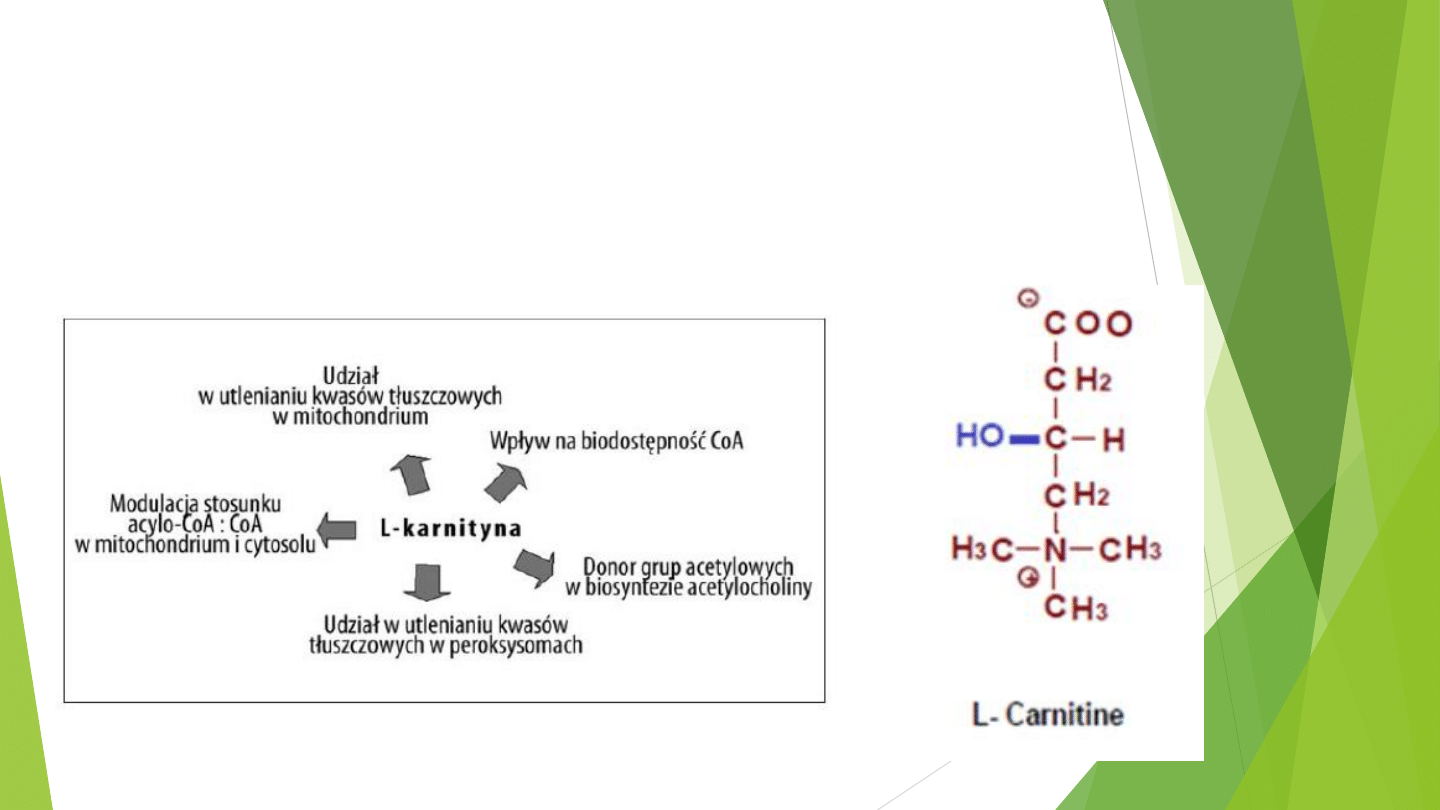
KARNITYNA
Beta-hydroksy-gamma-trimetyloaminomaślan
(CH3)3N+-CH2-CH(OH)-CH2-COO-
Wykazuje szeroką dystrybucję tkankową, lecz w szczególnie dużym stężeniu występuje w
mięśniach szkieletowych.

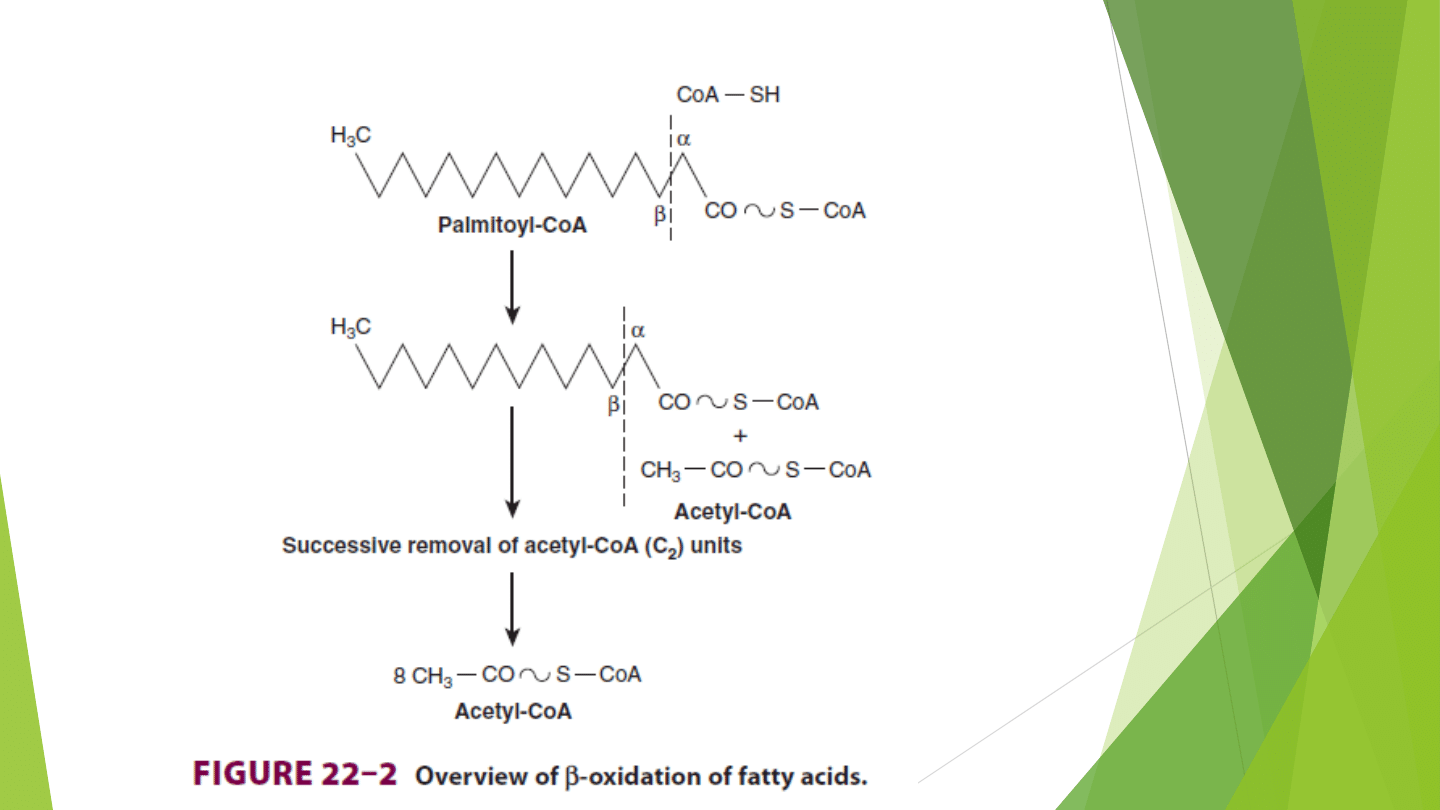
Beta-oksydacja
W każdej rundzie Beta-oksydacji następuję odcięcie dwuwęglowej
reszty od końca karboksylowego acylo-CoA
Rozerwaniu ulega wiązanie między atomami węgla alfa(2)-beta(3) ;
stąd nazwa procesu
Powstające w wyniku Beta-oksydacji jednostki dwuwęglowe to
acetylo-CoA
Enzymy które tworzą system utleniania kwasów tłuszczowych,
występują w macierzy mitochondrialnej jak i zewnętrznej błonie
tych organelli ( w sąsiedztwie łańcucha oddechowego)
Enzymy te katalizują proces utleniania acylo-CoA do acetylo-CoA
Proces cykliczny
Powstają duże ilości równoważników redukcyjnych – FADH2 i NADH
, które są następnie wykorzystywane w procesie fosforylacji
oksydacyjnej ( synteza ATP)


Kwasy tłuszczowe o nieparzystej liczbie atomów są utleniane w
procesie Beta-oksydacji, dostarczając acetylo-CoA do momentu
powstania reszty trójwęglowej (prioponylo-CoA).
Propionylo-CoA jest przekształcany w bursztynylo-CoA (jeden z
metabolitów cyklu kwasu cytrynowego)
Reszta propionylowa (powstająca w wyniku utl. Kwasów
tłuszczowych o nieparzystej liczbie atomów węgla ) jest zatem
jedyną częścią kwasu tłuszczowego o właściwościach
glukogennych.
Utlenienie :
•
FADH2 = 1,5 MOLA ATP
•
NADH = 2,5 MOLA ATP

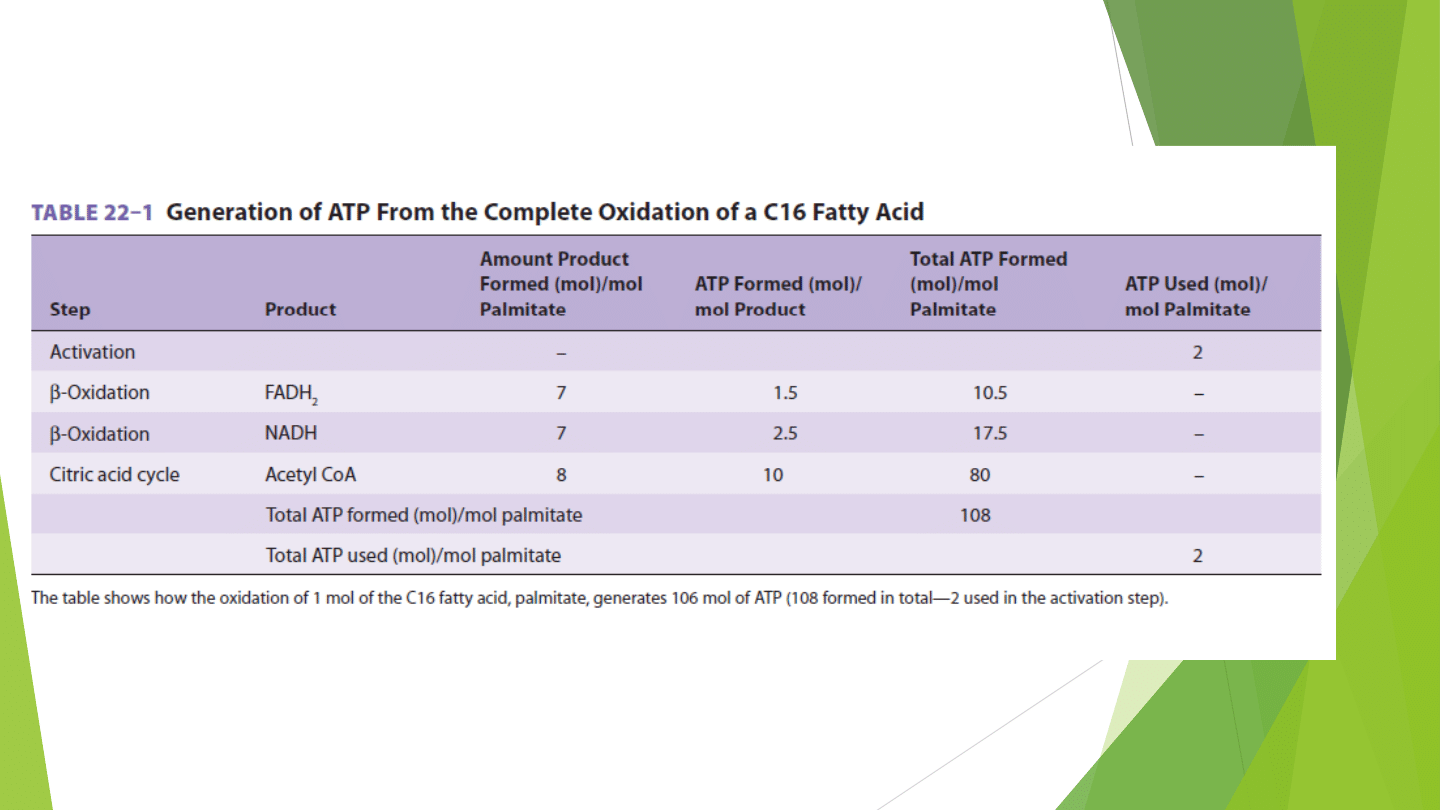

UTLENIANIE KWASÓW TŁUSZCZOWYCH O BARDZO DŁUGICH
ŁAŃCUCHACH (VLCFA – very long chain fatty acid)
Zachodzi w peroksysomach i prowadzi do wytworzenia acetylo-CoA i
nadtlenku wodoru (H202)
Nadtlenek wodoru powstaje w reakcji katalizowanej przez FAD-
zależną dehydrogenazę i jest następnie rozkładany przez katalazę
Funkcjonujący w peroksysomach system enzymów ułatwia utlenianie
kwasów tłuszczowych o bardzo długim łańcuchu (C20,C22)
Enzymy szlaku Beta-oksydacji znajdujące się w peroksysomach nie
działają na kwasy tłuszczowe o krótszych łańcuchach
Beta-oksydacja zachodząca w tych organellach kończy się na
oktanoilo-CoA
Inną formą Beta-oksydacji peroksysomalnej jest skracanie bocznego
łańcucha cholesterolu podczas biosyntezy kwasów żółciowych
Peroksysomy biorą też udział w syntezie glicerolipidów
eterowych,cholesterolu i dolicholu.
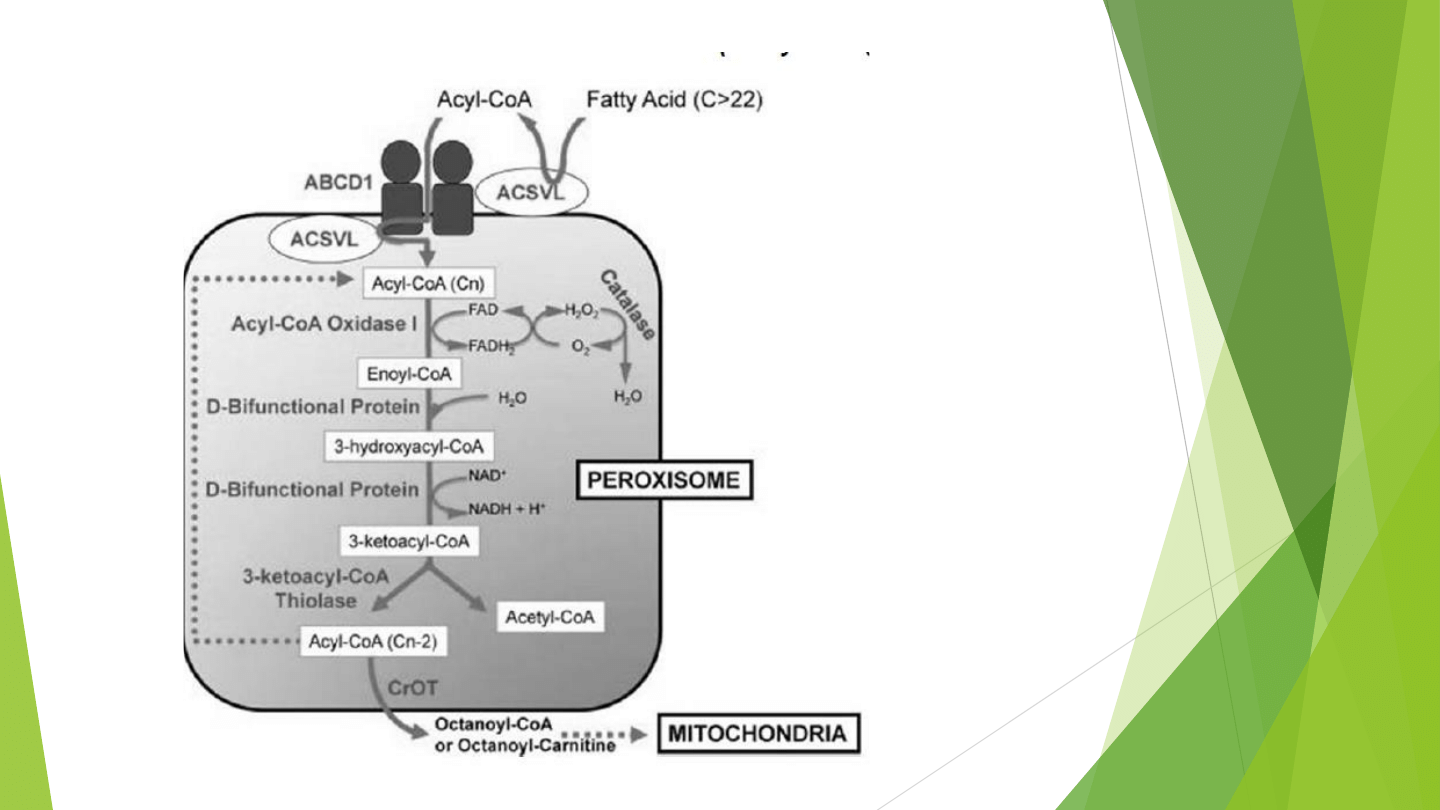
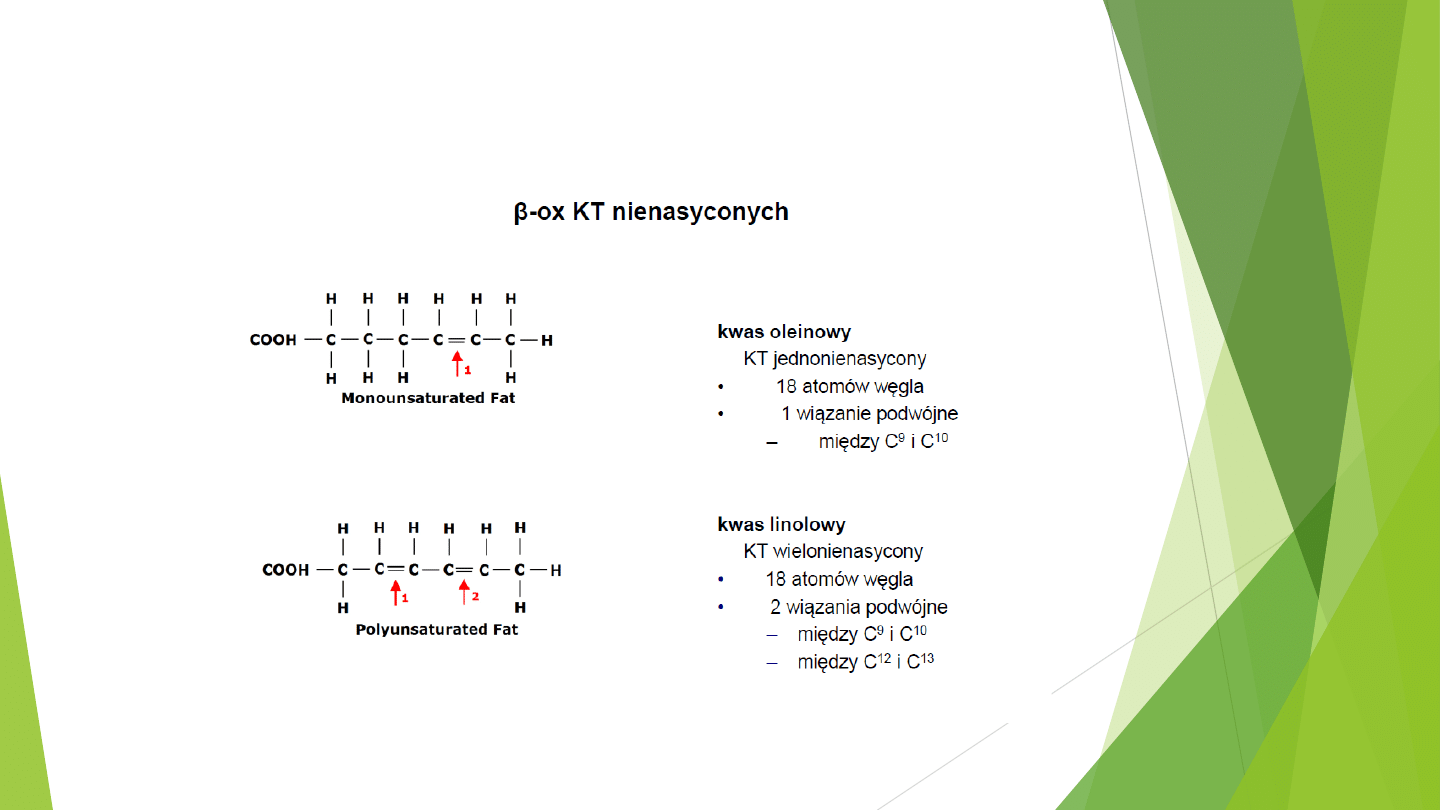
Beta-oksydacja nienasyconych kwasów tłuszczowych
przebiega szlakiem zmodyfikowanej Beta-oksydacji

ASPEKTY KLINICZNE
Niedobór karnityny :
•
Występują głównie u wcześniaków z powodu niewystarczającej
biosyntezy tego związku lub jego nadmiernej utraty drogą
nerkową.
•
Objawy : hipoglikemia, nagromadzenie lipidów i związane z tym
osłabienie mięśni.
•
Leczone poprzez suplementację karnityną.
Dziedziczny niedobór CTP I :
•
Wpływa wyłącznie na metabolizm wątroby i wywołuje obniżenie
wydajności zachodzących tu procesów utleniania kwasów
tłuszczowych i ketogenezy, czemu towarzyszy hipoglikemia.
Niedobór CTP II : wpływa głównie na mięśnie szkieletowe
Wrodzone defekty enzymów biorących udział w Beta-oksydacji i
ketogenezie wywołują hipoglikemię nieketotyczną, śpiączkę i
stłuszczenie wątroby

Jamajska choroba wymiotna :
Acyduria dwukarboksylowa
Choroba Refsuma
Zespół Zellwegera (zespół
mózgowo-wątrobowo-
nerkowy)

Źródła :
Biochemia Harpera Ilustrowana , wydanie VII , 2018r.
(rozdział 22)
Wykłady dr. Jadwigii Pietkiewicz
Wyszukiwarka
Podobne podstrony:
ENZYMY prezentacja biochemia
Prezentacja z biochemii - izoenzymy, Lekarski WLK SUM, lekarski, biochemia, enzymy
Prezentacja biochemia
ENZYMY prezentacja biochemia 2
ENZYMY prezentacja biochemia
Aminy biogenne prezentacja biochem
Mikro i Makroelementy pierwiastki życia, Prezentacje Biologiczne PPT, Biochemia
FOSFORYLACJA OKSYDACYJNA - PREZENTACJA, Weterynaria rok 1, Biochemia
Kiedy nasz organizm zużywa tlen powstają produkty pośrednie, Licencjat, Semestr III, Biochemia, Prez
skt-n, Pomoce naukowe, Opracowania, II rok, Biochemia, Prezentacje z seminariów, IV koło
podstawy biotechnologii biochemia prezentacja
biochemia prezentacja
UP Biochemia Biol Prezentacja enzymy
prezentacja finanse ludnosci
prezentacja mikro Kubska 2
Religia Mezopotamii prezentacja
Prezentacja konsument ostateczna
Strategie marketingowe prezentacje wykład
więcej podobnych podstron