<Wstecz Powstanie życia
Jak powstało życie?
Autor tekstu: Marcin Klapczynski
Słowo wstępne
Zachęcony przez Pana Mariusza Agnosiewicza postanowiłem napisać cykl trzech artykułów o powstaniu życia na Ziemi. Dostępny mi materiał podzieliłem na trzy części:
Pierwsza z nich to opis doświadczenia Millera-Ureya, które rozpoczęło rozkwit dziedziny biologii opisującej powstanie życia. Starałem się przekazać posiadaną mi wiedzę w sposób jasny i czytelny. Ponieważ zagadnienie to często jest wykorzystywane w nieuczciwy sposób przez kreacjonistów poprzez zaciemnianie faktów i ukrywanie sukcesów (i jak zwykle zawalanie czytelnika terminami naukowymi, krasomówczą propagandą i rysunkami spod rąk artystów plastyków), opisałem je dość szczegółowo i w większości ze znanych mi wariantów. Wynik doświadczenia i opis powstałych cząsteczek są nawiązaniem do
Części drugiej, która przedstawia szeroko akceptowaną teorię świata RNA, jako pierwszego świata biochemicznego. Od powstania, poprzez ewolucje in vitro, po wielorakość rodzajów. Świat pre-RNA. W artykule znajdzie się również informacja o konkurencyjnych teoriach: świat DNA, świat białek. Do kompletu potrzebna byłaby jedynie
Część trzecia, która opisuje powstanie pierwszych błon - prekursorów komórek, możliwości uwięzienia w nich systemów biochemicznych, co najwyraźniej prowadzi nas do powstania życia pełną, bakteryjną gębą.
Mam nadzieję, że moje artykuły spotkają się z pozytywnym, ale także i krytycznym odbiorem, co może zaowocuje ciekawymi dyskusjami na Forum Racjonalisty. Jestem gotów na odparcie wszelkich zarzutów i rozpatrzenie komentarzy.
Jak powstało życie?
Około trzy i pół miliarda lat temu młoda Ziemia nie wydawała się zbyt przyjazna życiu. Bezkresne oceany prebiotycznego roztworu, brak tlenu, powierzchnia bezlitośnie chłostana promieniami ultrafioletowymi jałowiącymi wszystko, czego dosięgły. Amoniak, metan, wodór i woda to prawdopodobnie główne składniki redukcyjnej atmo- i hydrosfery - nijak obiecujące powstanie tak niesamowitego tworu jakim jest życie. Jednak dzięki wyładowaniom elektrycznym w prabulionie zaczęły powstawać specyficzne cząsteczki organiczne - najważniejsze z nich to aminokwasy będące podstawowym składnikiem białek budujących nasze tkanki oraz pirymidyny i puryny - zasady azotowe wchodzące w skład łańcuchów RNA i DNA - najważniejszych nośników informacji w świecie ożywionym. Wyładowania elektryczne wciąż powodowały syntezę związków na całej powierzchni Ziemi, najwyraźniej z dobrym skutkiem - trzy i pół miliarda lat później młody student przyglądał się podobnym - sztucznie indukowanym wyładowaniom w kolbie wypełnionej metanem, amoniakiem i wodorem.
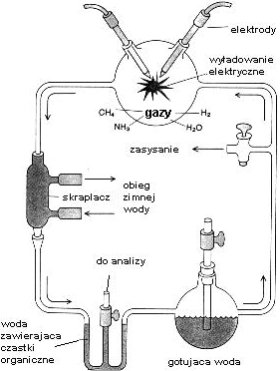
Tym studentem był Stanley L. Miller, pracujący w laboratorium Harolda C. Ureya na Uniwersytecie Chicagowskim. Dzięki determinacji młodego człowieka, doświadczenia te doszły do skutku, gdyż Urey miał wiele wątpliwości co do ich wyniku. Miller po raz pierwszy przeprowadził syntezę w wczesnych latach 50-tych dwudziestego wieku. (Patrz rysunek) Dolna kolba symulowała podgrzewany "ocean" parujący poprzez obieg i tworzący "atmosferę" zawierającą metan, amoniak i wodór w górnej kolbie. Tam też mieszanina gazów była poddawana działaniu prądu elektrycznego umożliwiając zachodzenie reakcji. Rozpuszczalne w wodzie produkty symulowanych procesów prebiotycznych były następnie skraplane i poddane analizie. Eksperyment okazał się zaskakująco wydajny - naukowcy spodziewali się jedynie śladów prymitywnego świata biochemicznego - uzyskali zaś 13 aminokwasów (z dwudziestu budujących białka w żywych organizmach) oraz pirymidyny i puryny (składniki łańcuchów RNA i DNA). Dwa ostatnie składniki okazały się niestabilne w pierwszych eksperymentach, co wzbudziło później wiele wątpliwości co do wysuniętej teorii o pionierskiej roli kwasów nukleinowych. Wartym wspomnienia jest fakt, że ten sam zestaw molekuł został znaleziony w meteorycie z Murchison, życie więc mogło powstawać w podobny sposób w dalekich nam częściach Wszechświata. Prawdopodobnie Ziemia to jedno z bardzo rzadkich w nim miejsc sprzyjających ekspansji życia.
Doświadczenie Millera-Ureya
Wyniki doświadczenia okazały się bardzo obiecujące, część świata naukowego podważała wiarygodność doświadczenia sugerując zanieczyszczenie bakteryjne, jednak wkrótce rozwiano wszelkie wątpliwości - aparatura użyta do doświadczenia była sterylizowana znacznie dłużej niż to jest wymagane.
Należy pamiętać, że skład praatmosfery jest oparty na pewnych założeniach. Spekuluje się również, iż duży odsetek stanowił CO2, produkowany przez wulkany, w którego obecność (zamieniając z metanem) rezultaty eksperymentów prebiotycznych nie są już tak imponujące. Nikt jednak nie wykluczy nieobecności metanu w atmosferze młodej Ziemi. Idea redukującej atmosfery ukazała się po raz pierwszy w latach dwudziestych ubiegłego wieku w opracowaniu rosyjskiego naukowca Oparina. Zakładał on, że pierwsze prymitywne organizmy musiały być heterotroficzne (czerpiące źródło energii z zewnątrz, lecz jej nie wytwarzające), podążając za znaną zasadą: im mniej biosyntezy, tym mniejszy stopień komplikacji. Zakładamy oczywiście, że najpierw powstały organizmy mniej skomplikowane. Dlatego najbardziej prawdopodobna jest atmosfera redukująca, jako warunki wyjściowe do powstania składników organicznych.
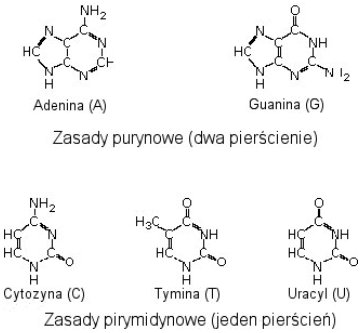
Zupełnie niezrozumiałe jest uporczywe wskazywanie na labilność powstałych cząsteczek w prawdziwych warunkach prebiotycznych przez krytyków teorii samorzutnego powstania życia. Zdaję sobie sprawę, że mieszanina pobierana do analizy w aparacie Millera była schładzana, lecz łatwo również wyobrazić sobie naturalne warunki - ocean to nie statyczna zupa, nowopowstałe cząsteczki mogły być np. wmywane pod skały, które chroniły je przed niszczycielskim promieniowaniem UV. Poza tym, jeśli synteza przebiegała gwałtownie, przy specyficznych warunkach część cząsteczek mogła wytrącić się z roztworu (tak jak kożuch na mleku) tworząc przypadkowo warstwę ochronną dla cząsteczek znajdujących się poniżej. Ponieważ uzyskanie dużych stężeń cząsteczek, zwłaszcza pirymidyn, okazało się bardzo trudne, zespół badaczy wyszedł z tak zwaną 'teorią lagun'. Młoda Ziemia to nie tylko oceany, lecz również plaże, zagłębienia skalne. Wystarczyło więc zagęścić mieszaninę i podsuszyć - otrzymano imponujące ilości zasad azotowych [_1_]. Okazało się więc, że teoria kwasów nukleinowych jako cząsteczek pionierskich ma bardzo solidne podstawy.
Co ciekawe, używając otrzymane mieszaniny, poddawano je dalszym eksperymentom symulującym warunki panujące na "lagunach" - z prostych związków powstał kwas pantotenowy, związek ściśle skorelowany z koenzymem A, kluczowym elementem systemu formowania białek i cykli metabolicznych [_2_].
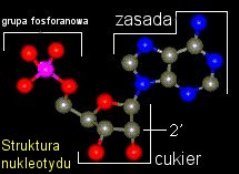
Każdy przyzna, że mieszanina zasad azotowych nadal nie obiecuje większej, spontanicznej złożoności, mimo, iż nawet ryboza (cukier wchodzący w skład nukleotydów tworzących RNA) została uzyskana w eksperymencie prebiotycznym i oczyszczona w obecności związków ołowiu [_3_]. Jak powstały pierwsze nukleotydy? Otóż już w roku 1963 wykazano, iż ze związków obecnych w mieszaninie uzyskanej przez Millera może powstać nukleotyd adeninowy [_10_], nota bene najbardziej wytrzymały na promieniowanie UV. Może właśnie ta cecha naznaczyła go później jako najbardziej "popularne" źródło energii chemicznej - ATP? Grupa naukowców udowodniła, że jeśli w roztworze będzie obecny metafosforan etylu, ryboza i adenina, po naświetlaniu UV powstaną znaczne ilości nukleotydu. Nie możemy być całkowicie pewni, że metafosforan etylu istniał na młodej Ziemi. W roku 1965 wykazano jednak prostą drogę dla formowania się wszystkich nukleotydów (razem z adeninowym) poprzez odpowiednie wystawienie na wysoką temperaturę i zastosowanie różnych źródeł grupy fosforanowej, które mogły istnieć w prebiotycznym środowisku. [_11_]
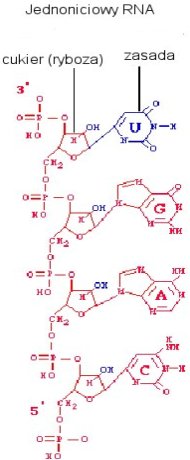
Wiemy, że nukleotydy mogą się spontanicznie formować w losowe łańcuchy na minerałach. Glin i krzem, wypłukane ze skał wulkanicznych, formowały warstwową strukturę, z ujemnym ładunkiem między warstwami. Cecha ta powodowała przyciąganie jonów i tworzenie przez nie dodatniej warstwy, która zaś była idealnym podłożem do formowania się łańcucha RNA. [_4_] Obecnie próbuje się wyjaśnić jak wiązania między nukleotydami są formowane. To odkrycie jest jednym z bardziej spektakularnych w biologii powstania życia. Przypadkowa formacja łańcuchów RNA mogła prowadzić do powstania form katalitycznie aktywnych, czyli cząsteczek o aktywnym i skłonnym do mutacji replikowaniu się. Fascynujący świat pionierskich cząsteczek RNA zostanie przeze mnie opisany w następnym artykule.
Co ciekawe, inny minerał skalny potrafi katalizować łączenie się aminokwasów w łańcuchy peptydowe. [_5_] Nie są to jeszcze aktywne białka, ale coś na podobieństwo krótkich ich fragmentów. Czyż to wyraźnie nie wskazuje na względnie dużą szansę na powstanie zalążków życia z prostych substancji i skał?
Skoro już jesteśmy przy aminokwasach - w doświadczeniach prebiotycznych pojawił się jeszcze jeden problem. Jak okazało się, mieszanina aminokwasów była racemiczna, czyli występowały w niej po równo izoformy D i L (formy te posiadają różną zdolność do skręcania płaszczyzny polaryzacji swiatła). Należy tu wspomnieć, że również racemiczną reprezentację cząstek znalezionych w meteorycie z Murchison. Nota bene, inny wynik tego doświadczenia wskazywałby na zanieczyszczenie bakteryjne.
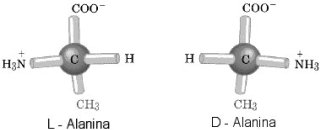
Jak wiemy, świat żywy jest zdecydowanie zdominowany, z bardzo małymi wyjątkami [_6_], [_7_], formami L aminokwasów. Wydaje się, że przewaga ta została wytworzona później, na drodze selekcji - po pierwsze, odporności na promieniowanie UV, po drugie, okazuje się, że formy D znacznie słabiej łączą się do tRNA [_8_] - małych transporterów, które przenoszą aminokwasy do rybosomów, gdzie składane są łańcuchy białkowe. Poza tym, do reliktów aktywnej selekcji zaliczyłbym również specjalny enzym - D-Tyr-tRNA Tyr deacylaza - który usuwa niepoprawny D aminokwas przyłączony przez przypadek do tRNA [_9_]. Fakty z obecnego świata biochemicznego bogatego w relikty pozwalają nam przypuszczać, że tak jak wspomniano wyżej formy L utrwaliły się na drodze selekcji.
Czy świat powstały ze szczęścia i przypadku w świetle powyższych faktów wydaje się niemożliwy? Niepotrzebne jest tu żonglowanie statystyką, wymysły o czarnych i białych kulkach w worku, gdyż tak naprawdę żaden kreacjonista nie jest w stanie wymyślić wzoru na życie. Jeśli uzmysłowimy sobie, że szansa na powstanie aktywnych cząsteczek życia była niemal równa, (dodajmy nawet: niewielka), w każdej kropli na całym obszarze ziemskim, to czyż biadolenie o wygranej na loterii nie wydaje się bezpodstawnym i nieracjonalnym bełkotem? Czy nie lepiej cieszyć się dotychczasowymi sukcesami? O wiele ciekawiej jest zadawać sobie pytania: jak to mogło być; wypełniać te luki w naszej niewiedzy i tworzyć kompletną teorię powstania życia, początków początku, nas samych. Wiem, że dużo łatwiejsze dla wielu będzie wpychanie w tę lukę kreatora z jego magiczną różdżką, ale jeśli tak łatwe miałoby być nasze poznawanie, życie byłoby najnudniejszym tworem, jaki powstał.
Jak powstał pierwszy system biochemiczny?
Autor tekstu: Marcin Klapczyński
Podążając za porównaniem Leslie E. Orgela nauka o początkach życia przypomina bardzo skomplikowaną i dobrze zaplątaną historię kryminalną. Jest w niej wielu podejrzanych, każdy ma jakieś alibi, jednak każdy również coś kręci, czujny nos naukowca-detektywa węszy wciąż jakąś intrygę. Na dodatek nikt nie chce zostać sędzią, ława przysięgłych niczego nie rozumie, a każdy z powołanych rzeczoznawców mówi coś innego. Materiał dowodowy jest delikatnie mówiąc szczątkowy, proces zdecydowanie jest poszlakowy. Jednak mimo wszystko postaram się przedstawić głównych podejrzanych i wpakować chociaż niektórych do aresztu tymczasowego.
Należałby się tutaj krótki wstęp, aby rozjaśnić zagadnienie pierwszych systemów biochemicznych, które byłyby następnym poważnym krokiem w procesie powstawania życia, zaraz po powstaniu podstawowych cząsteczek życia opisywanych przeze mnie w poprzednim artykule (str. 3065). Zachęcam, aby nie rezygnować z powodu czasami skomplikowanie wyglądających ilustracji oraz złożonych opisów, gdyż przy uważnym przeczytaniu nie powinny stanowić problemu.
Zacznijmy od podstawowego przepływu informacji. Jak każdy wie, podstawowym jej nośnikiem w organizmach żywych jest dzisiaj DNA enkryptujący ją za pomocą czterech "liter" A,C,G,T - nukleotydów w których skład wchodzą między innymi zasady azotowe (patrz poprzedni artykuł) od których nazw skrótami są te litery. Co oznacza właściwie 'kod genetyczny'? Otóż litery zawarte w DNA ułożone są w odpowiedniej kolejności, czyli mają zakodowany pewien sens, instrukcję. Instrukcja ta służy do budowania białek - cząsteczek złożonych z łańcuchów aminokwasów (patrz poprzedni artykuł) zwiniętych w "kłębki", które tworzą nasze mięśnie, paznokcie, skórę, przenoszą tlen, czy też bronią nasz organizm przed patogenami. Jednym słowem - to cegiełki budujące najważniejsze części naszego organizmu. Jednak informacja ta nie jest przekazywana bezpośrednio z DNA na łańcuch białkowy - ma pewnego pośrednika - tak zwany informacyjny mRNA (z ang. messenger RNA).
Rysunek 1. (por. przyp. 1.) Procesy zachodzące w komórce bakteryjnej.
Rysunek 2. (por. przyp. 1.) Procesy zachodzące w komórkach organizmów wyższych.
Przyjrzyjmy się temu nieco bliżej [_1_]. Zarówno u Prokaryota (prymitywnych organizmów pozbawionych jądra, np. bakterii) jak i Eukaryota (czyli komórek roślinnych czy zwierzęcych) proces przebiega podobnie. [Patrz rysunki.] Informacja z DNA przepisywana (tzw. transkrypcja) jest na mRNA poprzez enzym zwany polimerazą RNA. Przepisywanie polega na dobieraniu odpowiednich liter (w alfabecie RNA nie ma T, zastąpiona jest natomiast poprzez U) przez co informacja jest wierna. Następnie kod genetyczny mRNA jest tłumaczony (tzw. translacja) na łańcuch białkowy.
U Prokaryota oba procesy zachodzą w cytoplazmie i symultanicznie. U Eukaryota są one rozdzielone przestrzennie i czasowo; mRNA ulega często dodatkowej obróbce przed tłumaczeniem. Translacja w obu przypadkach przebiega w rybosomach - "fabryczkach" produkujących białka. Poszczególne aminokwasy aktywowane i doprowadzane są przez małe cząsteczki transportującego - tRNA.
Kiedy czytelnik połapie się w tym wszystkim, może nasunie mu się pewna myśl - skoro mówimy tu o powstaniu życia.....zaraz, zaraz to co z tego było pierwsze?! Białko nie może powstać bez informacji z DNA, zaś DNA nie może powielać się, ani przekazywać informacji bez białka (polimerazy)! I tu dochodzimy do słynnego problemu kury i jajka w biochemii. Co było pierwsze?
|
Najbardziej popularną obecnie, jest teoria świata RNA jako pierwszego systemu biochemicznego. Okazuje się, że niektóre cząsteczki RNA potrafią replikować się w prymitywny sposób, na zasadzie komplementarności nici, zawierając jednocześnie zakodowaną informację, np. ligazy [_2_][_2_]. Jeszcze bardziej ciekawym przykładem jest enzym pochodzenia wirusowego tzw. Q-beta replikaza - zdolny do replikowania RNA, działający na instrukcji zawartej na matrycy. [_12_] Ma on niesamowite zdolności katalityczne niegdyś przypisywane tylko białkom. Potrafi np. samodzielnie produkować RNA w zamkniętych pęcherzykach mając do dyspozycji tylko nukleotydy oraz RNA "na wzór". [_13_] Cząsteczki te zwane są rybozymami. Jedną z głównych przesłanek jest skład mieszaniny prebiotycznej (patrz poprzedni artykuł) - rybozy i zasad azotowych. Poza tym łączenie się nukleotydów w losowe łańcuchy na minerałach jest bardzo dobrze udokumentowane. Samoreplikujące RNA posiadałoby bardzo ważną cechę - brak perfekcyjnej dokładności w kopiowaniu siebie. W przypadku polimerazy w naszych komórkach istnieje skomplikowany i wysublimowany system sprawdzania i korekty źle wstawionych literek. Brak systemu edytującego w przypadku pierwszych RNA byłby źródłem wybuchu ich różnorodności i przypadkowego nabycia przez nie nowych cech. Czyli obrazowo przedstawiając: na warstwie mineralnej powstaje łańcuszek ...ACAGAUUAAAC... i pływa sobie w pierwotnym roztworze bezczynnie, ale zaś przypadkowo złożony ...GGGAGGUUCCUCCC... wygina się i przyjmuje taką pozycję, która umożliwia mu samoprzecięcie się, czyli zyskuje zdolność katalityczną. Pierwsze kopiowanie zachodziło zapewne na spontanicznym "doklejaniu" się swobodnie pływających "literek". Te kopie tworzą dalsze kopie, te zaś dalsze, itd.
Tutaj chciałbym wspomnieć o spektakularnym doświadczeniu. Naukowcy z Kalifornii przeprowadzili eksperyment ewolucji in vitro z udziałem około 500 trylionow losowo wygenerowanych cząsteczek RNA [1]. Poddano je pewnej selekcji: wiązania do pewnej substancji oraz aktywności. Rybozymy umieszczono w warunkach korzystnych do swobodnej, tudzież błędnej replikacji. Wynik był niesamowity - produktem końcowym puli rybozymów była struktura bliźniaczo podobna do tRNA - elementu, który rozpoznaje i "nosi" aminokwasy do rybosomu umożliwiając budowę łańcucha białkowego.
W swoim poprzednim artykule wspominałem o abiotycznej drodze powstawania nukleotydów, która nie wydaje się zbyt wydajna na potrzeby rozwijającego się życia. Otóż sytuacja wygląda zupełnie inaczej już w obecności RNA - grupa naukowców z Cambridge udowodniła, że może ono wspomagać i przeprowadzać reakcję tworzenia nukleotydów [2]. Pierwotna synteza tych cegiełek była prawdopodobnie mało wydajna, jednak w momencie powstania odpowiednio aktywnych cząsteczek RNA została znakomicie udoskonalona. Udowodniono również katalizę powstawania odcinków RNA, czyli dodatkowe wspomaganie dla prostego mechanizmu tworzenia łańcuchów na minerałach wspomnianego wyżej [2].
Istnieje jeszcze bardzo ważna przesłanka - przyjrzyjmy się jeszcze raz tym małym fabryczkom produkującym białka - okazuje się, że zbudowane są nie tylko z białek, lecz również RNA o bardzo konserwatywnej sekwencji (są prawie identyczne w całym świecie ożywionym). Okazuje się, że te RNA bynajmniej nie stanowią rusztowania, ale są enzymatycznie aktywne, co oznacza, że biorą czynny udział w łączeniu aminokwasów w łańcuch białkowy. Wzbudza to w nas optymistyczne podejrzenie, iż w dalekiej przeszłości samo RNA, samodzielnie bez "łupiny" białkowej, zaczęło nieśmiale tłumaczyć instrukcję na pierwsze białka [3]. Te ostatnie jako bardziej elastyczne i znacznie bardziej aktywne najprawdopodobniej zaczęły przejmować funkcje enzymatyczne pozostawiając RNA przede wszystkim magazynowanie informacji. Wkrótce pojawił się inny bohater - DNA, który już tylko przez swoją dwuniciowość jest znacznie bardziej wiarygodnym magazynierem - obie nici muszą do siebie "pasować" - A pasuje tylko z T i odwrotnie, G pasuje tylko do C i odwrotnie (nazywane to jest komplementarnością par zasad). To dlatego (stabilność) w nowoczesnym świecie biochemicznym RNA został zdegradowany głównie do chłopca na posyłki między DNA a białkami. Przy wybuchu różnorodności aktywnych białek, powstał również wyrafinowany system sprawdzania wstawionych liter, który służy nam w każdej naszej komórce.
Pozwolę sobie tutaj na małą dygresję. Chciałbym odpowiedzieć na jeden z poważniejszych zarzutów ze strony bojówkarzy Intelligent Design o nieredukowalności systemów biochemicznych. Chociażby powyższy przykład rybosomów daje jasno do zrozumienia, iż "niepełna" maszyneria komórkowa dalej będzie pełnić swoją funkcję. Transkrypcję można zasymulować in vitro bez obecności wszystkich czynników, które występują w komórce, z nadal korzystnym skutkiem. W wielu przypadkach usunięcie wydawałoby się niezbędnych elementów ze szlaków metabolicznych ukazuje, że istnieje droga "na skróty" - system prostszy, może i będący reliktem. Wszystko wskazuje na to, iż systemy biochemiczne również podlegały i podlegają procesom ewolucji.
Powracając do tematu - krótkie odcinki RNA są niezbędne do powielania się DNA (np. podczas podziału komórki, kiedy materiał genetyczny musi być zdublowany), może to również świadczyć na korzyść RNA, które poprzedzało DNA.
To główne zarzuty kierowane w kierunku RNA, popatrzmy teraz na białka, mimo iż raczej nie są akceptowane jako pionierzy pierwszego systemu biochemicznego. Główna przesłanka, to powstanie większości z aminokwasów w zupie Millera (patrz poprzedni artykuł) - 13 z dwudziestu oraz innych substancji, które mogą prowadzić do powstania reszty. Mimo, iż pewien rodzaj minerału może katalizować powstawanie krótkich odcinków białkowych, jednak dobranie w ten sposób przypadkowo sekwencji, która utworzyłaby chociażby niewielkie, funkcjonalne białko zajęłoby kosmiczną liczbę kombinacji. Poza tym takowe, nawet funkcjonalne białko, nie miałoby możliwości przekazania informacji, utworzenia kolejnego, "lepszego" lub podobnego pokolenia. Dlaczego zajmujemy się więc białkami? Pamiętajmy, że poruszamy się trochę po omacku, gdyż pierwszy system biochemiczny mógł nie pozostawić żadnych śladów w systemach dzisiejszych. Istnieje teoria, że świat genetyczny (czyli ten który niesie informacje z pokolenia na pokolenie) poprzedzony był światem metabolicznym [4] (czyli "przerabiającym" cząsteczki) w którego skład mogły wchodzić białka. Daje to kilka innych możliwości, gdyż aparatura takiego systemu metabolicznego mogła ułatwić powstanie RNA. Jednak wydaje się, że użyteczne białka powstały raczej na matrycy samoreplikujących się RNA.
Przyjrzyjmy się teraz sławetnemu, prezentowanemu nieomal z czcią w prasie - DNA, które w końcu składuje informację między innymi o nas samych. Raczej nikt nie traktuje DNA jako cząsteczki pionierskiej w świecie prebiotycznym. Jego spontaniczne powstanie jest praktycznie niemożliwe, mimo iż nawet deoksyryboza (cukier w nukleotydzie DNA - patrz poprzedni artykuł) jest bardziej stabilna od rybozy i bardziej reaktywna. Na dodatek, dowiedziono, że deoksyryboza rozpada się 2.6 raza wolniej w 100 st.C niż ryboza [5]. Najprawdopodobniej DNA powstało na zasadzie odwrotnej transkrypcji - kierunek jest tutaj odwrócony - już prawdopodobnie w obecności białek. DNA potrafiło przyjąć formę dwuniciową, nukleotyd U został podmieniony przez metylowany T - końcowym efektem była cząsteczka bardzo stabilna, wytrzymała i wiarygodna przy przekazywaniu i przechowywaniu informacji.
Ponieważ całkiem możliwym wydaje się, iż świat RNA wcale nie musiał być pierwszym systemem biochemicznym, naukowcy z pasją rozpoczęli poszukiwania tajemniczej cząsteczki, która zatarła po sobie wszelki ślad z miejsca zbrodni. Należałoby więc poszukać podejrzanego, któryby łatwo tworzył spontanicznie łańcuch, mógł nieść informacje genetyczną, najlepiej korzystając z języka zasad azotowych i w prosty sposób mógł przekazać pałeczkę dla RNA. Pozwolę sobie przedstawić teraz bardziej egzotycznych podejrzanych.
Jednym z ciekawszych kandydatów jest PNA [6] (z ang. peptide nucleic acid), który może zawierać informację, tworzyć komplementarne pary zasad, mając jednocześnie bardzo prosty, amidowy "kręgosłup" - spójrz na rysunek powyżej - tutaj porównanie do pojedynczej nici DNA. Co więcej, zostało dowiedzione, iż PNA może przekazać informację na RNA służąc jako matryca [7], przekazując funkcję temu ostatniemu.
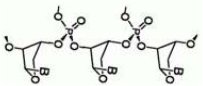
Zespół badaczy z Bostonu sugeruje, że jedną z pierwszych genetycznych molekuł mogła być również cząsteczka TNA [8] bardzo podobna do RNA, tylko oparta na triozie, cukrze prostszym niż ryboza. Spójrz na rysunek obok (B oznacza base, czyli dowolną zasadę wchodzącą w skład łańcucha). Prace nad właściwościami trwają.
Również wciąż analizuje się właściwości p-RNA (z ang. pyranosyl-RNA), który bardzo łatwo formuje polimery, cukier zaś występuje w innej formie [9].
W mowie końcowej, szanowna ławo przysięgłych, trudno jest nie wskazać RNA jako wyraźnego faworyta, jednak przypomnijmy sobie, że formowanie się życia zachodziło około czterech miliardów lat temu. To co dziś mamy do dyspozycji, to bardziej poszlaki, niż mocne dowody. Zaznaczam jeszcze raz, iż pierwszy system biochemiczny mógł nie pozostawić zupełnie żadnych śladów w obecnym zestawie komórkowej maszynerii. Dlatego tajemnica może drzemie w innych, nietypowych cząsteczkach zdolnych do niesienia informacji? Obserwujmy więc uważnie najnowsze odkrycia biologii powstania życia, zanim główni podejrzani wyjdą za kaucją.
Proces w sprawie pierwszego systemu biochemicznego mamy za sobą, jednak coś tu nie pasuje nam w całości.....jakim cudem ta biochemiczna kakofonia zorganizowała się w funkcjonalne komórki? Ów cud postaram się przedstawić w następnym artykule. Serdecznie zapraszam.
Kiepska sprawa z UFOludkami
Autor tekstu: Roman Zaroff
Od zarania dziejów ludzkość fascynowała możliwość istnienia innych światów i innych istot rozumnych. Do pewnego stopnia rezultatem tej fascynacji są przeróżne mitologie i religie oraz mity i legendy w których występują antropomorficzne monstra, stwory i bóstwa. W dzisiejszym świecie rolę mitów i legend, a często też religii przejęła wiara w UFO-ludków. Raporty na temat UFO, porwań i innych bzdur wypełniają komercyjne wiadomości w TV, radiu i brukowej prasie. Pojawili się UFO-lodzy, kluby porwanych, setki bezwartościowych, pseudo-naukowych, publikacji, filmów etc. Ten margines intelektualny oparty na ignorancji lub szwindlu wytworzył wrażenie, że kwestią życia pozaziemskiego i pozaziemskich istot inteligentnych zajmują się jedynie nawiedzeni i oszuści. W rzeczywistości tak wcale nie jest. Istnieje np. przy NASA ośrodek exobiologii poszukujący życia poza Ziemią. Od lat, choć często borykając się z trudnościami finansowymi, funkcjonuje program SETI (Search for Extraterrestial Inteligence - Poszukiwania Pozaziemskiej Inteligencji).
Niestety naukowe poszukiwania ET jak dotychczas nie dają rezultatów. Coraz więcej naukowców skłania się do opinii, że cywilizacje technologiczne nie muszą być wcale powszechne we Wszechświecie. Co oczywiście nie oznacza, że ich być nie może, że kiedyś nie istniały lub, że istnieją w odległych rejonach Drogi Mlecznej lub w innych galaktykach. Do takich wniosków skłaniają naukowców wyniki trwającego z małymi przerwami od lat 30 programu SETI, i ostatnio ogłoszony przez nich raport.
Trzeba się zgodzić z osobami związanymi z SETI, że jeśli jakaś pozaziemska cywilizacja wysyłałaby sygnały w kosmos użyłaby logicznie najprostszej metody mającej charakter uniwersalny. Takim medium do wysyłania sygnałów są fale radiowe o częstotliwości emisji podstawowego składnika Wszechświata to jest wodoru. Wynosi ona 1,42 gigaherców. Nasłuch SETI był prowadzony w częstotliwościach od 1 do 3 gigaherca.
Do analizy stworzono klasyfikacje cywilizacji technologicznych. Według tzw. skali Arecibo cywilizacja typu I to taka, która jest w stanie wyemitować sygnał o mocy 10 do potęgi 16 watów, czyli tyle ile wynosi cała energia słoneczna otrzymywana w danym momencie przez naszą planetę. Typ II to takie cywilizacje, które są w stanie wyemitować sygnał mocy 10 do 27 potęgi watów, czyli energii gwiazdy wielkości Słońca. Hipotetyczna klasa III, byłaby w stanie użyć energii emitowanej przez całą galaktykę, czyli 10 do potęgi 38 watów. Nasza cywilizacja jest typu 0,7; ale nie oznacza to, że jesteśmy już w stanie wysłać sygnał o mocy otrzymywanej przez Ziemię energii słonecznej. Bowiem typy cywilizacji nie są funkcją liniową a logarytmiczną. Dotychczasowe wyniki nasłuchu przez SETI wskazują, że w promieniu 50 lat świetlnych nie ma cywilizacji typu I, ani nie jest zbyt prawdopodobne, aby istniały cywilizacje o trochę wyższym niż naszym poziomie, które "obecnie" [_1_] próbują nawiązać kontakt z innymi cywilizacjami. Ponadto, nasłuch SETI objął jak dotychczas częściowo gwiazdy odległe o 4000 lat świetlnych (cywilizacje na zbliżonym do naszego poziomie) oraz częściowe poszukiwania cywilizacji typu I do dystansu 40 tysięcy lat świetlnych. Cywilizacje typu II i III prawie na pewno w ogóle w naszej galaktyce nie istnieją.
Te niedawno ogłoszone wyniki badań SETI sugerują, że cywilizacje pozaziemskie na wyższym niż nasz poziomie nie są tak częste jak jeszcze niedawno sądzono. Rzadkie występowanie zaawansowanych cywilizacji technologicznych nie oznacza oczywiście, że życia we Wszechświecie nie ma. Oznacza to jedynie, że najprawdopodobniej nie istnieją w "najbliższym" naszym sąsiedztwie, to jest w promieniu około 50 lat świetlnych.
Nie oznacza to też, że życie na Ziemi jest zjawiskiem unikalnym. Większość naukowców sądzi obecnie, że życie nie jest unikalne we Wszechświecie. Sugeruje to ewolucja organizmów żywych na Ziemi. Ziemia liczy sobie 4,6 miliarda lat, ale warunki umożliwiające życie istnieją na niej od około 4 miliardów lat. Najstarsze skamieliny pochodzą z Zachodniej Australii i są datowane na 3,5 miliarda lat. Organizmy te jednak były stosunkowo zaawansowane i musiały wyewoluować znacznie wcześniej. Niektóre geologiczne formacje pochodzące sprzed 3,8 miliarda lat są interpretowane jako pozostałości organizmów żywych. Wynikało by z tego, że życie powstało na Ziemi w stosunkowo krótkim czasie to jest w ciągu około 200 milionów lat. Tak więc przy sprzyjających warunkach powstanie życia opartego na węglu nie może być rzeczą unikalną. Nie oznacza to jednak, że złożone formy organiczne czy istoty inteligentne oraz cywilizacje technologiczne są automatyczną konsekwencją powstania życia. Warto tu zwrócić uwagę na fakt, że organizmy wielokomórkowe wyewoluowały "dopiero" 700 milionów lat temu, a więc jednokomórkowce egzystowały na Ziemi przez ponad 2 miliardy lat. Od powstania wielokomórkowców do istot inteligentnych i cywilizacji technologicznych daleka jest droga a ilość klimatycznych, geologicznych i kosmicznych czynników mających na ten proces wpływ jest przeogromna. Dlatego powstanie życia nie musi koniecznie gwarantować powstania inteligencji czy cywilizacji technologicznej.
Exobiologia jest w dużym stopniu nauką teoretyczną i jej największym problemem jest to, że badania nad pozaziemskim życiem z przyczyn oczywistych muszą opierać się na analogiach ziemskich. To znaczy na jednej formie życia. Niemniej nie jest to czcze gdybanie. Opinia ta jest zapewne zabarwiona naszym antropocentryzmem, albo raczej ziemio-centryzmem. Za życiem opartym na węglu przemawia jednak wiele innych czynników. Podstawowe pierwiastki z których jesteśmy zbudowani występują w dużych ilościach we Wszechświecie. Wodór jest najobfitszy, tlen zajmuje trzecie miejsce. A więc w odpowiednich warunkach nie powinno brakować "budulca" dla wody. Węgiel jest również dość powszechny. Ostatnie badania astronomiczne, astrofizyczne i przy pomocy sond kosmicznych potwierdzają, że skomplikowane związki organiczne (oparte na węglu) są dość powszechne we Wszechświecie. Aminokwasy, nukleotydy, bazy łączące DNA, itp. wykryto w pyle kosmicznym, asteroidach, meteorach i kometach, jak również na Tytanie (księżyc Saturna) i Europie (księżyc Jowisza). Są to jednak tylko "budulce", które nie musiały, i zapewne nie powstały, w efekcie procesów biologicznych. Niemniej odpowiedni "materiał" nie jest rzadkością i potrzebne są tylko sprzyjające warunki żeby życie mogło powstać. Podstawowym budulcem żywych organizmów na Ziemi są białka oparte na skomplikowanych molekułach węglowych. Właściwości węgla umożliwią mu łączenie się w długie skomplikowane cząsteczki aminokwasy, nukleotydy, itp. podstawowe "budulce" żywych organizmów. Niezbędna jest również woda w procesach przemiany materii, jako "nośnik" substancji odżywczych, enzymów, hormonów itp. Oraz tlen jako składnik wody i biochemicznych reakcji energetycznych.
Z innych pierwiastków praktycznie tylko krzem (Si) posiada zbliżone właściwości do węgla. Niestety życie oparte na krzemie jest mniej prawdopodobne niż oparte na węglu. Teoretyczne molekuły krzemowe ze względu na jego chemiczne właściwości byłyby znacznie mniej skomplikowane niż węglowe. Co oznacza znacznie mniejszą potencjalną różnorodność "budulca", a w konsekwencji w najlepszym teoretycznym przypadku nader prymitywne formy życia. Pozostaje jeszcze wiele innych problemów. Nawet prymitywne organizmy żywe cechują się ogromną liczbą skomplikowanych procesów fizjologicznych. Najlepszym dla nich medium są ciecze ze względu na ich właściwości fizyczne. W przypadku życia opartego na węglu rolę taką spełnia woda, składająca się z najliczniejszego we Wszechświecie wodoru i stosunkowo powszechnego tlenu. W przypadku teoretycznego życia opartego na krzemie i środowiska z wysokimi temperaturami nie ma odpowiedniego medium, które w takich warunkach spełniłoby rolę jaką w naszych organizmach spełnia woda. Ponadto wydalanym przez organizm produktem spalania byłby nie gazowy dwutlenek węgla (CO2) a dwutlenek krzemu (SiO2), czyli zwykły piasek. Wszystko to bardzo komplikuje potencjalny metabolizm gdzie pojawia się tu zasadniczy problem zastąpienia wody jakąś cieczą, która w wysokich temperaturach spełniałaby role nośnika w procesach metabolicznych. Dlatego też życie oparte na krzemie jest praktycznie rzecz biorąc nieprawdopodobne. Niemniej życie oparte na węglu może być, jak uczą ziemskie przykłady, bardzo różnorodne.
Odrzucając wąski antropocentryzm najpowszechniejszą i dominującą formą życia na Ziemi są bakterie. Występują one w najbardziej ekstremalnych warunkach i najprzeróżniejszych postaciach. Istnieją bakterie żyjące w wodach gejzerów lub na dużych głębokościach w ziemi, gdzie temperatury przekraczają 100 stopni Celsjusza. W radioaktywnej wodzie, zabójczej dla człowieka, w systemach chłodzenia reaktorów również odkryto spokojnie żyjące bakterie. Nie tak dawno ogromną sensację wzbudził marsjański meteoryt ALH84001 gdzie odkryto odciski struktur przypominających bakterie o rozmiarach od 20 to 250 mikronów. Głównie ze względu na ich bardzo mały rozmiar odrzucono możliwość, że są to odciski marsjańskich bakterii. Gdy miano przejść do porządku dziennego, naukowcy z Uniwersytetu Queenslandzkiego w Brisbane w Australii odkryli żyjące w głębi ziemi organizmy, których rozmiary są takie jak odciski w meteorycie. Dla odróżnienia ich od mikrobów nazwano je nanobami, gdyż mierzą one od 20 do 150 nanometrów. Dla porównania najmniejsze bakterie Mycoplazmy mierzą 150 nanometrów a największe do 50 mikronów. Spór czym są odciski w meteorycie rozgorzał więc od nowa a sprawa życia na Marsie jest wciąż nierozstrzygnięta.
Jednym z kluczowych założeń exobiologów, jest że życie oparte jest na węglu. Obecnie badania nad życiem w kosmosie idą w kierunku poszukiwania odpowiednich warunków. To znaczy wody, odpowiednich temperatur i związków organicznych opartych na węglu. Jednocześnie zwraca się uwagę na potencjalną znaczną obecność wolnego tlenu. Bowiem byłoby to "sygnaturą" życia opartego na białku węglowym. Warunki do powstania życia odkryto w Układzie Słonecznym w kilku miejscach. Takie sprzyjające warunki mogą zaistnieć na planetach lub na dużych księżycach. W naszym Układzie Słonecznym mamy jak na razie trzech potencjalnych kandydatów: Marsa, Tytana i Europę. Nie wykluczone, że może również dołączyć do tego grona Ganimed.
Amerykańska marsjańska misja "Pathfinder i Sojourner" potwierdziła, że w zamierzchłej przeszłości na Marsie istniały spore zasoby wodne. Jednocześnie w 1996 roku odkryto dziwne struktury w meteorycie marsjańskim znalezionym na Antarktydzie. Struktury, które mogą być skamielinami marsjańskimi bakterio-podobnych organizmów, sprzed 3.6 miliarda lat. Czy są to skamieliny czy też struktury mineralne trudno jeszcze w tej chwili powiedzieć. Intensywne badania trwają w wielu instytutach naukowych świata. Jak wykazują badania woda w stanie wolnym istniała najprawdopodobniej na Marsie i pokrywała znaczną część tej planety jakieś 3 do 4 miliardów lat temu. Jeśli istniała tam woda mogło powstać na Marsie życie. Obecnie na Marsie, prawie całkowity brak wody oraz atmosfera składająca się przede wszystkim z dwutlenku węgla nie sugeruje istnienia życia. Niemniej niewykluczone, że w przeszłości życie na Marsie istniało. Niemniej biorąc pod uwagę przykłady ziemskie i ogromną adaptowalność życia do ekstremalnych warunków niewykluczone, że życie mogło tam przetrwać np. pod powierzchnią Marsa. Istnieją przecież na Ziemi beztlenowe bakterie, jak też bakterie i archae głęboko w skorupie ziemskiej w temperaturach przekraczających +100 stopni Celsjusza. W ciągu najbliższych 10 lat NASA planuje kilka misji marsjańskich włącznie z przywiezieniem na Ziemię próbek gruntu marsjańskiego. Musimy więc uzbroić się w cierpliwość.
Innymi kandydatami są księżyc Jowisza Europa i Saturna, Tytan. Z przeprowadzonych badań astrofizycznych i z pokładów sond kosmicznych wynika, że na obu tych księżycach znajdują się ogromne zasoby wody pod ich powierzchnią. Europa jest księżycem Jowisza o promieniu 1460 kilometrów (Ziemia 6350 kilometrów). Na jej powierzchni panuje temperatura około -200 stopni Celsjusza. A więc raczej nieprzychylne warunki. Ostatnio jednak amerykańska sonda kosmiczna "Galileo" dostarczyła rewelacyjnych informacji. Na Europie znajdują się ogromne zasoby wody a na jej powierzchni zaobserwowano "lodowe wulkany". Pod powierzchnią woda najprawdopodobniej znajduje się w stanie ciekłym. Utrzymanie się wysokiej temperatury wewnątrz księżyców jest skutkiem działania ogromnej grawitacji Jowisza na różnorodne warstwy tego księżyca. Siła grawitacji wywołuje tu naprężenia i "przesunięcia" różnorodnych warstw (np. jądro księżyców, skorupa) wyzwalające znaczne ilości energii. Nie wykluczone więc, że pod powierzchnią tych planet mogło powstać życie. Oczywiście ich forma byłaby dla nas niewyobrażalna. Nie ma tam przecież słonecznego światła i dwutlenku węgla, więc organizmy żyjące tam musiały by czerpać energię z reakcji chemicznych jak niektóre ziemskie bakterie beztlenowe.
Tytan jest księżycem Saturna zbliżonym wielkością do Merkurego, o promieniu 2550 kilometrów. Średnia temperatura powierzchni wynosi -179 stopni Celsjusza. Jego gęsta atmosfera składa się głównie z azotu i metanu. Laboratoryjna symulacja warunków atmosferycznych Tytana wykazała, że pod wpływem promieniowania kosmicznego i ultrafioletu powstają w niej bardziej skomplikowane związki organiczne, które opadają na powierzchnie księżyca. Uczeni podejrzewają obecność wody, oczywiście w formie lodu, na powierzchni i pod powierzchnią Tytana. Nie jest też wykluczone, że pod powierzchnią księżyca może istnieć woda w stanie płynnym. Podobnie jak na Europie ogrzana wewnętrznymi ruchami księżyca na skutek działania grawitacji Saturna. Mamy więc i tutaj wszystkie potrzebne składniki i w miarę sprzyjające warunki, choć diametralnie różne od ziemskich.
Nie wykluczone, że życie istnieje również na planetach krążących wokół innych słońc. Nasza wiedza o układach planetarnych poza układem słonecznym jest bardzo skąpa. Dopiero w roku 1994 polski astronom pracujący w USA, Aleksander Wolszczan, odkrył trzy planety okrążające pulsara B 1257+12 oddalonego od nas o 1400 lat świetlnych. Jak dotychczas (koniec roku 2000) odkryto blisko 50 planet okrążających inne niż nasze słońca. Niestety, żadna z tych gwiazd nie jest tego samego typu co Słońce, a planety są dużych rozmiarów (najmniejsza 2/3 Jowisza). Tak więc na odkrycie "bliźniaczki" Ziemi musimy jeszcze trochę poczekać. Nie zmienia to jednak postaci rzeczy, że w świetle obecnej wiedzy możemy stwierdzić, że planety krążące wokół innych gwiazd są zjawiskiem dość powszechnym.
Na zakończenie chciałbym raz jeszcze wyrazić przekonanie, że osobiście uważam, że jeszcze za naszego życia znajdziemy dowody na istnienie życia we Wszechświecie. Niewykluczone, że już wkrótce na Marsie. Niestety ku rozczarowaniu UFOlogów będą to najprawdopodobniej tak zwane niższe formy, bakterio-, archeo- lub algo-podobne. Kontakt z obcą cywilizacją, znacznie mniej prawdopodobny, również może kiedyś nastąpić, ale na pewno nie według scenariusza z filmów takich jak "ET" czy "Alien". Muszę kończyć, bo właśnie zaczyna się X-Files ("Z archiwum X")...
Pierwsze błony komórkowe
Autor tekstu: Marcin Klapczyński
Jeśli przebrnąłeś, Drogi Czytelniku, przez moje poprzednie dwa artykuły (str. 3065 i 3150) i masz wrażenie jakbyś przedzierał się przez gęsto zarośniętą przez prebiotyczne stwory dżunglę - masz rację. Nauka o początkach życia to teren wciąż na nowo poznawany, niczym nietknięte ludzką stopą skrawki ziemi, na których botanik z zaskoczeniem odnajduje dziwnego rodzaju kwiaty. Trzymając się dalej tego porównania, wędrówka przez zagadnienie cząsteczek prebiotycznych i pierwszych systemów biochemicznych to łatwiejsza część naszej podróży. I nie chodzi tu o stopień skomplikowania czy też zaawansowania w posiadanej wiedzy - chcąc nie chcąc pchamy się w głębokie błoto z ruchomymi piaskami. Brak nam dobrego przewodnika, jest ich kilku, ale ich nabazgrane na owczej skórze mapy mówią zupełnie coś innego. Najgorsze jest to, że jak już wdepniemy w bagno, konsekwentnie musimy się przez nie przedrzeć, aby uniknąć wyśmiania przez prymitywne plemię Kreacjonusów (które od pokoleń jest zdania, że przez bagna przedzierać się nie warto). Zadanie to ciężkie, ale spróbuję nakreślić mały przewodnik dla odkrywców pierwszych zaczątków już jak najbardziej współczesnego nam życia.
Mimo, iż najbardziej podstawowa definicja życia twierdzi, że tworem żywym jest układ biochemiczny podlegający ewolucji darwinowskiej, jesteśmy raczej skłonni akceptować życie w formie komórkowej. To już jakby namacalny dowód - możemy takie "coś" obejrzeć pod mikroskopem i być świadkiem jak zawzięcie macha rzęską w poszukiwaniu pokarmu. Żywa komórka nie jest jednak niczym innym niż imponująco skoordynowanym i skomplikowanym układem biochemicznym uwięzionym w błonie komórkowej. Właściwie należałoby użyć słowa "chronionym", gdyż błona komórkowa nie jest bynajmniej nieprzenikalną ścianą, a bardzo złożonym, selektywnie przepuszczającym tworem. Rycina 1 to bardzo duże uproszczenie tego niesamowitego, dynamicznego tworu.
Rycina 1. Mozaikowy model błony współczesnych komórek.
Oprócz lipidów, składa się ona z wszelakiej maści białek transbłonowych biorących udział w transporcie cząsteczek do i z wnętrza komórki. Błona komórkowa jest przepuszczalna jedynie dla niektórych. Pośród nich znajdują się cząsteczki wody oraz gazów takich jak tlen, azot i dwutlenek węgla oraz dla lipidów takich jak cholesterol. Ograniczona przepuszczalność dotyczy niektórych jonów, cukrów i innych małych molekuł. Przepływ wody może być zwiększony poprzez ostatnio intensywnie badane kanały wodne. Przez miliardy lat błony wykształcały system kanałów do biernego transportu np. jonów, co jest podstawą wrażliwości komórek. Oprócz tego istnieje również wyrafinowany system transportu aktywnego (zużywającego energię chemiczną) - pozwalający na przenoszenie "pod prąd". Powrócimy do tego zagadnienia trochę później. Błona komórkowa wystawia swoje "czułki" w postaci łańcuchów cukrowych, co jest podstawą wszelakiej interakcji ze środowiskiem. Cała struktura jest elastyczna i znajduje się w ciągłym ruchu.
Jeśli chcemy być konsekwentni, musimy założyć kilka rzeczy, które pozwoliłyby na powstanie pierwszych tworów komórkopodobnych.
Po pierwsze: pierwotna błona musiała uformować się spontanicznie z materiałów dostępnych na młodej Ziemi, bądź/oraz zawleczonych z kosmosu. Mało prawdopodobne byłoby założenie, że pływające swobodnie cząsteczki utworzyły miejscowe zagęszczenie prowadząc do skomplikowanego systemu produkcji lipidów i ich dystrybucji wokół zagęszczenia.
Po drugie: błona ta oprócz oczywistej ochrony systemu biochemicznego od środowiska będzie zapewniała chociażby najbardziej podstawowy rodzaj transportu. Miałoby to służyć do utrzymania odpowiedniego stężenia jonów po obu stronach, dostarczania substratów do syntezy wewnątrz oraz usuwanie niepotrzebnych produktów.
Po trzecie: powstała błona musi umożliwić "rozmnażanie" pierwotnej komórce. Czy byłoby to prymitywne pączkowanie, czy bardziej lub mniej zaawansowany podział - sprawny układ biochemiczny musi przekazać korzystne cechy następnemu "pokoleniu".
Powstanie. Witamy w tłustym towarzystwie
Wydaje się całkiem prawdopodobne, że nie tylko cząsteczki dostępne na młodej Ziemi, ale również składniki pozaziemskich obiektów mogły być źródłem powstania pierwszych błon. Szacuje się, że bombardowanie meteorytami wczesnej Ziemi mogło dostarczyć około 107 kg na rok organicznych materiałów [1]. Wydaje się, iż rodzime surowce byłyby wystarczające, trudno jest jednak wykluczyć zawleczenia zarodków życia na Ziemię.
Wróćmy do budowy błony komórkowej, a właściwie do podstawowych "cegiełek" ją budujących. Ograniczę się jednak do podstawowych informacji, niezbędnych do dalszej lektury artykułu. Głównym składnikiem obecnych błon komórkowych są fosfolipidy (Rycina 4A) - cząsteczki zaliczane do grupy związków amfifilowych. Oznacza to, że posiadają obydwie cechy z reguły pojedynczo przypisane do jednej cząsteczki. W tym przypadku jest ona podzielona na dwa regiony - umowna "głowa" jest hydrofilowa, czyli "lubi" środowisko wodne, zaś "ogon" jest hydrofobowy, czyli będzie zawsze dążył od ukrycia się przed cząsteczkami wody.
Wyobraźmy sobie mieszaninę takich cząsteczek - otóż mają one skłonność do spontanicznego łączenia się w tzw. micele - drobne pęcherzyki oraz w dwuwarstwy (Rycina 2 i 3 - widać, że twory te powstają na skutek "uciekania" części hydrofobowych cząsteczek od środowiska wodnego - próbują się one upakować jak najciaśniej, wystawiając na zewnątrz części hydrofilowe). W specyficznych warunkach mogą również tworzyć zamknięte dwuwarstwy, na kilka różnych sposobów opisanych dalej.
Rycina 2. Formowanie uporządkowanych struktur przez cząsteczki amfifilowe. [2]
Rycina 3. Szczegółowy model dwuwarstwy utworzonej przez fosfolipidy.
Zadajmy więc podstawowe pytanie. Czy tak skomplikowane fosfolipidy były dostępne na prebiotycznej Ziemi? Najwyraźniej nie. Były jednak dostępne inne cząsteczki, znacznie prostsze lecz nadal zachowujące właściwości amfifilowe (spójrz na porównanie - Rycina 4). Wśród najpowszechniejszych znajdują się proste kwasy i alkohole oraz pochodne związków zwanych w skrócie PAH (polycyclic aromatic hydrocarbon - policykliczne węglowodory aromatyczne), proponuje się również pochodne izoprenów.
Rycina 4. Składnikiem współczesnych błon komórkowych są fosfolipidy (A). Składnikami pierwszych prymitywnych błon mogły być np. kwas 5-metylo nonanowy (B) obecny w meteorytach, czy substancja zwana w skrócie POPC (C). [2, 3]
O tym, że cząsteczki takiego rodzaju były zdolne do formowania zamkniętych dwuwarstw (pęcherzyków) świadczy doświadczenie zespołu dr Davida Deamera, który wyekstrahował je z skrawka meteorytu z Murchison. Ten dość spory kawałek gwiezdnej skały okazał się bezcennym znaleziskiem. Nie dosyć, że posiadał substancje uzyskane w eksperymencie Ureya-Millera (co potwierdziło jedynie nieuniknione powstanie życia we Wszechświecie), zawierał również cząsteczki amfifilowe takie jak kwas 5-metylononanowy (Rycina 4B) i przedstawicieli z grupy PAH. Otóż okazuje się, że takie cząsteczki mogą formować twory komórkopodobne w miejscowych zagęszczeniach [2]. Tutaj przychodzi z pomocą teoria lagun Stanleya Millera, o której wspominałem w poprzednich artykułach. Otóż parująca woda z niecek, brzegów mogła powodować zwiększenie stężenia w tym również omawianych przez nas cząsteczek. Tak więc podczas symulacji odpowiednio zagęszczone substancje amfifilowe spontanicznie formowały pęcherzyki (Rycina 5 A). Kalifornijscy naukowcy wskazują właśnie na PAH jako główne ich składniki. Mało tego - podobne doświadczenie przeprowadził dr James Dworkin używając jednak ekstraktu z zamrożonej materii międzygwiezdnej [2]. Tutaj głównym składnikiem była mieszanina kwasu i alkoholu dekanowego. (Rycina 5 B)
Rycina 5. (A) - ekstrakt z meteorytu z Murchison; pęcherzyki są zdolne do uwięzienia fluorescencyjnego barwnika. (B) - ekstrakt z materii międzygwiezdnej. Fluorescencyjna substancja barwi dwuwarstwy. [2, 3]
Spontaniczne formowanie się pęcherzyków nie wymaga zewnętrznej chemicznej energii, jednak zachowanych musi być kilka warunków. Jednym z nich jest, jak widzimy powyżej, ich odpowiednie zagęszczenie.
Proces ten mógł zachodzić na różne sposoby, takie jak wspomniane wyparowywanie w lagunach, krótkotrwałe zamarzanie i topnienie, czy adsorpcję na powierzchniach mineralnych [2]. Ostatni przypadek byłby najbardziej faworyzowany, gdyż, jak pamiętamy, na takich powierzchniach mogą być formowane katalitycznie aktywne RNA. Wnikanie cieczy z substancjami w szczeliny dawałoby bardzo dobrą ochronę przed nieprzyjaznymi warunkami zewnętrznymi. Okazuje się, że dwuwarstwowe pęcherzyki mogą być łatwo tworzone z miceli w cyklach ogrzewania i chłodzenia [4], które mogły być bardzo powszechnym zjawiskiem geologiczno-klimatycznym na młodej Ziemi. Proponuje się również inną możliwość - polimeryzację atmosferycznego metanu pod wpływem promieniowania UV. Substancja ta tworzyłaby swoistego rodzaju "kożuch" hydrofobowy pływający na dużych powierzchniach zbiornika i tworzyła parasol ochronny dla pływających pod nim cząsteczek [5]. Ostatnio ta kwestia stała się bardzo ważna, gdyż ponownie odkryto ślady komórkowych organizmów fotosyntetycznych datowanych na 3,7 miliarda lat, więc znacznie starsze zalążki życia musiały powstawać w wysoce niekorzystnych warunkach [6].
Przełomowe badania M. Hanczyca, S. Fujikawy i J. Szostaka publikowane w zeszłym roku zademonstrowały ewidentnie spontaniczne formowanie się dwuwarstwowych pęcherzyków w obecności ilastego minerału - montmorylonitu [7]. Patrz Rycina 5.1.
Rycina 5.1. Oto seria doświadczeń przeprowadzonych przez Hanczyca, Fujikawę i Szostaka. [7] Przedstawione tu pęcherzyki zostały uwidocznione fluorescencyjnie. Panele I, J przedstawiają czerwono barwione RNA zamknięte w wybarwionych na zielono pęcherzykach. Panele F,H przedstawiają wybarwiony montmorylonit - minerał wspomagający formowanie się pęcherzyków. Panele B,D pokazują pęcherzyki zawierające ciekawe twory aluminowo-krzemowe. [_1_]
Złapanie życia w ryzy
Powyżej wspomniana publikacja, moim zdaniem, jest równie wartościowa i sensacyjna jak pierwsze eksperymenty Stanleya-Millera (opisywany w pierwszym artykule). Naukowcy elegancko udowodnili, że formujące się pęcherzyki mogą zamykać w swoim wnętrzu gotowe RNA oraz fragmenty montmorylonitu, na których wciąż tworzą się nowe cząsteczki kwasu rybonukleinowego. Twory takie na dodatek mają zdolność do biernego, podstawowego transportu [7] - zaobserwowano z czasem przenikanie barwnika do wnętrz tych swoistych "pre-komórek".
P. Monnard, współpracujący z D.Deamerem, wykazał, że pęcherzyki formowane z kwasu i alkoholu dodekanowego mogą uwięzić mieszaninę cząsteczek DNA o długości około 600 par zasad (par nukleotydów) [2, 3]. Spójrz na poniższy rycina.
Rycina 6. Uwięzione DNA w pęcherzykach zbudowanych z kwasu i alkoholu dodekanowego. Pęcherzyki z DNA w panelu (A) są czarne, pod wpływem światła UV emitują intensywnie zielone światło. [3]
RNA uwięzione w takim pęcherzyku miało większą szansę na stworzenie funkcjonalnego systemu biochemicznego, niż pływając swobodnie w roztworze. Zwiększa się w tym wypadku możliwość wspólnych interakcji i nabycia nowych funkcji. Zwiększa się stopień uporządkowania całego układu. Co więcej, aktywowane nukleotydy mogły dyfundować w obydwie strony poprzez błonę, dostarczając budulca do nowych łańcuchów RNA. Inną drogą nukleotydów do wnętrza pre-komórki były naturalnie występujące, krótkotrwale tworzące się przerwy w dwuwarstwie. Zagadnienie transportu stanowi poważny problem, ograniczając możliwości pierwszych systemów metabolicznych.
Zakładamy raczej, że energia chemiczna była głównym źródłem zasilania w świecie prebiotycznym. A co z fotosyntezą? Czy ostatnie doniesienia o jej śladach 3,7 miliarda lat temu nie dają do myślenia? Aby wykorzystać energię słoneczną, fotony muszą być najpierw zaabsorbowane przez substancje pigmentowe i przekształcone w niestabilne formy energii chemicznej. Oczywiście systemy porównywalne do dzisiejszych komórek były jeszcze nieobecne, ale istniały wspomniane wcześniej związki z grupy PAH, które mogły zostać inkorporowane w zamknięte struktury dwuwarstwowe. Dotychczas pojawiło się kilka sygnałów o właściwościach PAH jako pigmentu [3]. Badania trwają.
Dzisiejszy, wydajny, nawet bierny (zgodnie z gradientem stężenia przenoszonej substancji) transport oparty jest na białkach przenośnikowych. (Rycina 7). Świat komórkowy dysponuje również fascynującym i wyrafinowanym systemem transportu aktywnego (wbrew gradientowi stężęń - Rycina 8) na wiele sposobów regulowanym.
Rycina 7. Rodzaje transportu biernego współczesnych błon. Transport zachodzi zawsze zgodnie z gradientem stężeń przenoszonej cząsteczki - jeśli substancji jest więcej na zewnątrz, będzie ona transportowana do wnętrza, aby wyrównać stężenia.
Rycina 8. Podstawowe rodzaje transportu aktywnego współczesnych błon. Transport zachodzi niezależnie od gradientu stężeń przenoszonych cząsteczek, ale wymaga dostarczenia energii. Innymi formami transportu aktywnego są również tutaj nie opisywane endo- i egzocytoza.
Możemy spodziewać się więc, że wysoka efektywność przenoszenia rozpoczęła się dopiero po pojawieniu się pierwszych białek, które mogły być "złapane" w błonie. Nie zaobserwowano RNA tworzącego twory transbłonowe. Jak to się stało - jest dla nas na razie nierozwiązaną zagadką, ale trzymajmy kciuki za biologów symulujących powstanie życia...
Czas na dzieci. Czas na lepszą ewolucję
Wreszcie dotarliśmy do kwestii przekazania materiału genetycznego. Wrócimy tutaj do wcześniej wspominanego artykułu trójki naukowców. Postawili oni następującą tezę: prymitywne rozmnażanie najprawdopodobniej polegało na prostym podziale pęcherzyka razem z materiałem zawartym w środku. Jeśli wewnątrz pęcherzyka wykształci się pewien jednolity system, podzielenie go na pół da nam dwa pęcherzyki z systemami zdolnymi do niezależnych zmian i niezależnej ewolucji. Jednak aby utrzymać ten podział takie pra-komórki powinny rosnąć. Okazało się, że pęcherzyki rosną pod wpływem zwiększania się ilości miceli w roztworze. Czyli najpierw powstają micele, które przekształcają się w nowe dwuwarstwowe pęcherzyki, po czym po pewnym wysyceniu ogranicza się powstawanie nowych pęcherzyków a zwiększająca się wciąż ilość miceli. Cześć składników miceli - w na razie szczegółowo nie poznany sposób - łączy się z pęcherzykami zwiększając ich objętość. Proces ten dotyczy 90% puli pęcherzyków w roztworze [7].
Jak zachodzi podział? Otóż bynajmniej nie poprzez reakcje biochemiczne. Naukowcy przepuścili podrośnięte pęcherzyki przez porowaty materiał (o porach rozmiarów 100 nm) i okazało się, że podzieliły się ona na pół z bardzo małą stratą zawartego materiału. Następnie, aby potwierdzić produktywność tego systemu, przeplatali cykle wzrostu (dodając micele) oraz podziału (przepychając przez wspomnianą "gąbkę"). Całe doświadczenie dowiodło, że wzrost pęcherzyków jest możliwy dzięki naturalnemu wzrostowi stężenia miceli, zaś podział może zaistnieć dzięki prostym procesom fizykochemicznym [7].
P. Monnard i D. Deamer poszli o krok dalej - zaproponowali minimalny model komórki oparty na katalitycznym i strukturalnym RNA zobrazowany na Rysunku 9.
Rycina 9. Minimalny model komórki oparty na katalitycznym i strukturalnym RNA. Opis w tekście. [9]
Mimo iż teoria fragmentów RNA o właściwościach polimerazy RNA (produkowania kopii RNA; zob. poprzednie artykuły) jest niekompletna, to jest całkiem prawdopodobna i rozwijana [8]. Naukowcy twierdzą, że niezbędne byłyby trzy systemy [3]. Jeden zawierałby gen polimerazy, na podstawie którego polimeraza produkowałaby swoje bliźniacze kopie. Enzym ten produkowałby również na podstawie innego genu acylotransferazę, która aktywnie brałaby udział we wzroście błony komórkowej. Wreszcie namnażałby cząsteczki RNA odpowiedzialne za "pączkowanie". Model nie ma na razie poparcia doświadczalnego.
Jakimkolwiek torem podążał rozwój komórek - dzisiejsze efekty są imponujące. I pomyśleć, że około czterech, może i więcej miliardów lat temu wystarczyła iskierka. Resztę "roboty" wykonała ewolucja.
Ufam, że pomogłem Czytelnikom Racjonalisty nieco przybliżyć obecny stan wiedzy o początkach życia na Ziemi. Tutaj na razie kończymy naszą wędrówkę, mimo że na horyzoncie widać wiele terenów do eksploracji. Może kiedyś zaproszę jeszcze na podobną eskapadę w nieporównywalnie bardziej skomplikowany i równie fascynujący świat współczesnych komórek i pra-organizmów.
Dzisiejsza nauka potrafi przyprawić nas o zachwyt i coraz częściej zawroty głowy. Cieszę się ogromnie, że jestem świadkiem tak rewolucyjnych odkryć jak zeszłoroczny sukces Hanczyca, Fujikawy i Szostaka. Kilka tygodni temu koreańscy naukowcy, adaptując komórki macierzyste do hodowli, dokonali pierwszego zdecydowanego kroku w kierunku inżynierii tkankowej. Fizycy są coraz bliżej unifikacji, zwieńczenia ludzkiej nauki. Czekamy w napięciu na sygnały życia na Marsie. Mam nadzieję, że udało mi się chociaż trochę zarazić Czytelników moim entuzjazmem i zachęcić ich do własnych poszukiwań i rozmyślań nad początkami życia.
Wyszukiwarka
Podobne podstrony:
Różne teorie dotyczące powstania życia na Ziemi - różne prace, Rozrywka, SZKOŁA
powstanie zycia na ziemi
Komety odpowiedzialne za powstanie życia na Ziemi nowe dowody
Teorie dotyczące powstania życia na Ziemi
Powstanie życia na Ziemi 3
NOTATKI powstanie życia
Kreacjonizm wiara w powstanie życia przez stworzenie
ST o powstaniu życia
Powstanie i ewolucja zycia
Regionalizm jest zjawiskiem powstałym wskutek obrony przed centralizacją życia społeczno
embriopatie, embriopatie (wszystkie uszkodzenia powstałe w okresie życia wewnątrzmacicznego);
Powstanie i ewolucja zycia
Pierwsze formy życia mogły powstać w kraterach poimpaktowych
Weiner Hipotezy o powstaniu i wczesnej ewolucji życia (2009)
więcej podobnych podstron