Wykład 1
Komórka nerwowa (NEURON) - podstawowa jednostka strukturalna i czynnościowa OUN
Akson może być otoczony osłonką mielinową (która warunkuje cechy przekazywania informacji przez tę komórkę).
Początek aksonu stanowi wzgórek aksonu, na którym powstaje impuls.
4 strefy czynnościowe:
strefa wyjścia = receptorowa (dendrytyczna)
inicjacja impulsów( wzgórek aksonu) - wytwarzanie potencjału czynnościowego
strefa przewodzenia (akson)
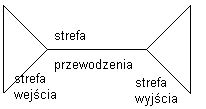
strefa wyjścia informacji o bodźcu = zakończenie neuronu -> uwolnienie transmitera synaptycznego.
Komórki układu nerwowego jako jedyne w naszym organizmie są pobudliwe (mogą odbierać bodźce, przekształcać je w potencjał czynnościowy i przekazywać ten bodziec na inne komórki nerwowe aż do komórek eferentnych)
Właściwości neuronu:
POBUDLIWOŚĆ
- zdolność wytwarzania zmian elektrolitycznychpotencjał spoczynkowy
potencjały postsynaptyczne
potencjały czynnościowe
Miejscem generowania i przewodzenia potencjału jest BŁONA KOMÓRKOWA. Jest podłożem morfologicznym potencjału, posiada różnicę przepuszczalności dla poszczególnych jonów (K+, Na+, Cl-, A-) oraz zdolność do czynnego transportu jonów.
Błona komórkowa posiada dla odpowiednich jonów kanały, przez które są one transportowane
kanały sodowe - DOKOMÓRKOWE (ze środowiska zewnętrznego do wewnętrznego) /zgodnie z gradientem/
kanały potasowe - ODKOMÓRKOWE /zgodnie z gradientem/
pompa sodowo-potasowa - enzym hydrolizujący ATP (dostarcza energii) /transport Na+ i K+ wbrew gradientowi/
Potencjał spoczynkowy - jeżeli na powierzchni komórki nerwowej pozostającej w stanie spoczynku ustawimy 2 elektrody badające to między nimi stwierdzimy różnicę potencjałów elektrycznych - IZOPOTENCJALNA
Jeżeli jedną z wyżej wymienionych elektrod wprowadzimy do środka komórki nerwowej pozostającej w spoczynku to między nimi stwierdzamy różnicę potencjałów: -70mV
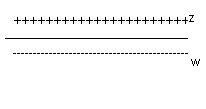
Co decyduje o potencjale spoczynkowym?
Różnice między jonami Na+ i K+ po obu stronach błony (ASYMETRIA JONOWA)
Dyfuzja tych jonów zgodnie z gradientem
Selektywna przepuszczalność błon względem tych jonów
Obecność metabolicznej pompy w Na+ - K+ błonie
PRZEPUSZCZALNOŚĆ błony dla Na+, K+, A-, Cl- zależy od stosunku średnicy jonu do średnicy kanału błonowego przez który jon dyfunduje.
Stosunek przepuszczalności K+:Na+:Cl-:A- -> 10:1:4:0 w stanie spoczynku
Stężenie Na+ w ECF (przestrzenie zewnętrzkom.) jest 10 x > niż w ICF
Stężenie K+ w ECF (przestrzenie zewnętrzkom.) jest 30 x < niż w ICF
g. A- = 0 -> tzn. w ICF > niż w ECF
-> elektrostatyczne oddziaływanie między A- i Cl- -> utrudnia dyfuzja Cl- do wnętrza w ECF > niż w ICF (zj. Donnona)
Potencjał równowagi jonu = wielkość potencjału elektrycznego po jednej stronie błony komórkowej, która równoważy się dyfuzji danego jonu ze strony przeciwnej -> obliczamy ze wzoru Nersta
En = (R*T/z*F) * ln Couni/Cin
Potencjał błonowy - jest to wypadkowa potencjału równowagi dla wielu jonów przy czym wartość potencjału błonowego jest najbliższa potencjałowi równowagi tego jonu, dla którego jest w danej chwili najbardziej przepuszczalna.
SPOCZYNEK:
błona najbardziej przepuszczalna dla K+
najbliższa potencjałowi równowagi dla K+, a najbardziej odległa od potencjału równowagi dla Na+
słabo przepuszczalna dla jonów Na+
Pompa Na+-K+
elektrogenna
transportuje czynnie 3 jony Na+ odkomórkowo i 2 K+ dokomórkowo -> dlatego, ze wewnątrz komórki jest niedobór elektronów
Cechy:
ATP
Prawidłowe działanie uwarunkowane jest obecnością Mg2+
Aktywność proporcjonalna jest do stężenia Na+ wewnątrz i K+ na zewnątrz
Asymetria działania
Blokują: niedotlenienie, glikozydy naparstnicy, ⇓ temperatura, jony Ca2+
Następstwem zahamowania pompy Na+-K+ jest ⇓ odkomórkowego prądu sodowego i ⇓ elektroujemności wewnątrz komórki = DEPOLARYZACJA
3 czynniki:
różna przepuszczalność błony komórkowej dla jonów
zjawisko Donnana
obecność pompy sodowo-potasowej
Stwarzają warunki:
w stanie spoczynku neuronu rozkład jonów po obu stronach błony kom jest asymetryczny
w stanie spoczynku na skutek asymetrii błony komórkowej jest spolaryzowana -> różnica potencjałów = -70 mV
w stanie spoczynku istnieje gradient stężeń dla Na+ skierowany dokomórkowo, dla K+ odkomórkowo
w stanie spoczynku istnieje gradient elektryczny dla Na+ i K+ skierowany dokomórkowo
BODZIEC - każde zjawisko lub zmiana zjawiska zachodząca w środowisku zewnętrznym i/lub wewnętrznym organizmu powodująca (+) lub (-)
Zadziałał bodziec
(+) (-)
Depolaryzacja Hiperpolaryzacja
Inicjacja/intensyfikacja Zahamowanie/spadek intensywności procesów procesów fizykochemicznych procesów fizykochemicznych
PODZIAŁ BODŹCÓW:
ze względu na naturę
1. fizyczne (mechaniczne, termiczne, elektryczne, świetlne, dźwiękowe)
2. chemiczne (związki nieorganiczne i organiczne)
3. biologiczne (stan pobudzenia układu nerwowego, potencjał czynnościowy, humoralne: hormony i mediatory)ze względu na siłę
1. podprogowe - o sile < niż bodźca progowego -> kom. nerwowe odpowiadają zmianą potencjału błonowego o cechach:
a) nierozprzestrzeniania się
b) odpowiedzi miejscowej
c) jej wielkość - od siły bodźca
- potencjał elektrochemiczny
- katelektronus
- anelektronus
2. progowe - doprowadza komórki do poziomu wyładowań -> pot czynnościowe (maksymalna odpowiedź)
3. nadprogowe - wywołuje taką samą odpowiedź jak bodziec progowy
4. maksymalny - o sile, która powoduje powstanie potencjału czynnościowego we wszystkich aksonach wchodzących w skład nerwu
5. uszkadzające - nocyceptywneze względu na wywoływany odruch
- bezwarunkowe
- warunkowepróg pobudliwości receptora
adekwatne - w stosunku, do którego receptor ma mniejszy próg pobudliwości
nieadekwatne - w stosunku do których próg pobudliwości receptora jest wyższy niż w odniesieniu do bodźca adekwatnego
Próg pobudliwości - najsłabszy bodziec zdolny wywołać w danych warunkach określoną reakcję (odwrotność pobudliwości)
POTENCJAŁ CZYNNOŚCIOWY
Jeżeli na komórkę nerwową zadziała bodziec o sile CO NAJMNIEJ progowym to komórka odpowie potencjałem czynnościowym, który:
jest gwałtownym, krótkotrwałym odwróceniem polaryzacji komórki - DEPOLARYZACJA
rozprzestrzenianie się wzdłuż włókna nerwowego ze stałą prędkością ( zależy od ∅ włókna i osłonki mielinowej) ze stałą amplitudą (uwarunkowaną wielkością elektroujemności wnętrza neuronu) -> „WSZYSTKO ALBO NIC”
FAZY POTENCJAŁU CZYNNOŚCIOWEGO
-70mV - + 40 mV = DEPOLARYZACJA - odwrócenie spoczynkowej polaryzacji neuronu
-70mV - -63mV - katelektronus - odpowiedź miejscowa bierna, jony nie przechodzą przez błonę
-63mV - -55mV - odpowiedź miejscowa czynna
-55mV - +40mV - szybka depolaryzacja
+40mV - -70mV - REPOLARYZACJA - powrót do wartości spoczynkowej
+40mV - -55mV - szybka repolaryzacja
-55mV - -70mV - depolaryzacja następcza
-70mV - ⇓ - hiperpolaryzacja następcza
DEPOLARYZACJA - ⇓ elektroujemności wnętrza neuronu (b. dodatnie)
HIPERPOLARYZACJA - ⇑ elektroujemności wnętrza neuronu (b. ujemne)
Jonowe podłoże potencjału czynnościowego
- Depolaryzacja = nagła krótkotrwała zmiana ⇑ przepuszczalności i przewodności elektrycznej dla jonów Na+ (aktywacja sodowa - otwarcie szybkich dokomórkowych kanałów sodowych i odwrócenie potencjałów w kierunku wartości dodatniej)
- Szczyt potencjału czynnościowego
- Repolaryzacja = wzrost przepuszczalności dla K+ (i przewodności) (aktywacja potasowa - otwarcie odkomórkowych kanałów potasowych, koniec szybkiej repolaryzacji)
W następstwie odkomórkowej dyfuzji K+ wnętrze neuronu staje się ujemne. Stężenie Na+ wewnątrz > niż w spoczynku, K+ na zewnątrz > w spoczynku.
Uruchomienie pompy sodowo-potasowej, która wyrzuca nadmiar Na+, potem następuje hiperpolaryzacja następcza. Jej następstwem jest ⇓ Na+ wewnątrz komórki i przywrócenie potencjału spoczynkowego. Dlatego zahamowana zostaje działalność pompy sodowo-potasowej -> SPOCZYNEK
Gdy komórka znajduje się w stanie odpowiedzi na bodziec to zmienia się wraz z czasem jej pobudliwość.
BEZWZGLĘDNA REFRAKCJA - (-55mV do 1/3 ramienia zstępującego) - otwarte kanały - nic nie jest w stanie pobudzić komórki
WZGLĘDNA REFRAKCJA - (1/3 ramienia zstępującego do początku depolaryzacji następczej) - jeżeli bodziec będzie nadprogowy to komórka odpowie
MIĘŚNIE
tkanka pobudliwa
narządy z nich zbudowane mają w bardzo znacznym stopniu zdolność kurczenia się (⇓ długości)
Skurcz mięśnia odbywa się pod wpływem bodźców (skutecznych):
mechanicznych
elektrycznych
chemicznych
+ SARKOLEMMY
Potencjał czynnościowy rozprzestrzenia się wzdłuż kanalików T lub sarkolemmy
EFEKT: - aktywacja układów kurczliwych = białka kurczliwe: miozyna + aktyna
zmiana napięcia mięśnia
zmiana długości mięśnia
PODZIAŁ MIĘŚNI:
Poprzecznie prążkowane:
a)szkieletowy
b) sercagładkie
CHARAKTERYSTYKA:
Ad 1. Przestrzenne uporządkowanie nitek aktyny i miozyny (efekt prążkowania)
Ad 2. Układ aktyny i miozyny jest bezładny i nie występuje w tym typie mięśni efekt prążkowania
Ad 1a.
komórki długie, wielojądrzaste, poprzecznie prążkowane włókna mięśniowe
poszczególne komórki nie posiadają połączenia autonomicznego i czynnościowego
unerwione są przez somatyczny układ nerwowy ( są zależne od naszej woli)
warunkiem skurczu jest (+) nerwowe: układ somatyczny pełni rolę inicjującą skurcz
depolaryzacja sarkolemmy spowodowana jest aktywacją sodową (otwarcie dokomórkowych kanałów Na+ )
Ad 1b. syncylium = masa wielojądrzasta
obecne poprzeczne prążkowanie
między komórkami występują złącza o niskiej oporności (zastawki) -> m. sercowy jest syncylium czynnościowym - masa o tej samej czynności
unerwiony prze AUN (jest niezależny od naszej woli)
posiada własny autonomiczny mechanizm rozrusznikowy (tzn. pewne struktury generują potencjał czynnościowy bez ingerencji z zewnątrz
AUN -> modeluje czynność skurczową (zmiany HR i siły skurczu) HR - częstość skurczów serca
za depolaryzację odpowiadają jony Na+ i Ca+
Ad 2.
brak poprzecznego prążkowania
zawierają znacznie mniej białek kurczliwych -> siła osiągana przez komórki mięśni gładkich w skurczu jest < niż w mięśniu szkieletowym
unerwione przez AUN (niezależne od naszej woli)
brak tropiny C i skąpe ER
Ca2+, odpowiedzialny za depolaryzację, dyfunduje za środowiska zewnątrzkomórkowego
-> następuje fosforylacja lekkich łańcuchów miozyny
-> aktywacja kalmoduliny
-> rozkład ATP => SKURCZMogą być (+) bodźcami elektrycznymi (unerwienie), bodźcami mechanicznymi (szybkie rozciąganie) -> dokomórkowy ruch Ca2+ => SKURCZ
Podział i charakterystyka mięśni gładkich:
Różnice - stosunek przestrzenny unerwiających je zakończeń AUN do komórek mięśniowych
Typy:
I wielojednostkowe
II jednostkowe (trzewne)
III pośrednie
Ad I
obfite unerwienie
współczulny splot podstawny z licznymi żylakowatościami, gęsty
odległość między żylakowatościami (NA, ATP, NPY) a komórką mięśniową wynosi 10-30 nm (jak szczelina synaptyczna) - do nich przylegają komórki połączone niskooporowymi złączami; pobudzenie obejmuje szybko cały mięsień i jest krótkie
skurcz ustępuje po ustaniu impulsów nerwowych
obecność receptorów α 2 hamuje uwalnianie NA (noradrenalina)
brak własnego automatyzmu
silna kontrola nerwowa
mało wrażliwe na KA (katecholaniny) krążące
np. ściana nasieniowodów, pęcherzy nasiennych, zwieracz i rozwieracz źrenicy
Ad II
skąpe unerwienie
duża odległość między komórkami mięśniowymi a żylakowatościami (.100 nm)
słaby wpływ współczulny - z dużym opóźnieniem - długo utrzymuje się pobudzenie (wolny wychwyt zwrotny, typ I)
bardzo wrażliwe na KA krążące
słaba kontrola AUN
obecność komórek rozrusznikowych (słaby automatyzm)
np. mięśnie gładkie ścian jelita (poza zwieraczami), głównie warstwa podłużna, moczowód, duże naczynia tętnicze
Ad III
np. średnie i małe tętnice (warstwa środkowa)
Podział i charakterystyka miocytów mięśni poprzecznie prążkowanych szkieletowych
włókna wolno kurczące się = czerwone = tlenowe = typ I = SO
włókna szybko kurczące się = białe
1. odporne na zmęczenie = tlenowo-glikolityczne = typ II A = FOG (fast twitch oxidative-glycolitic)
2. podatne na zmęczenie = glikolityczne = typ II x = FG (fast twitch fatigable)
JEDNOSTKA RUCHOWA - kompleks, w skład którego wchodzi jeden motoneuron i włókna mięśniowe unerwione wyłącznie przez ten motoneuron. Jest najmniejszą jednostką czynnościową mięśnia.
TYPY
S - slow twitch = wolna
F - fast = szybkie
FR - fast resistant = odporne na zmęczenie
FF - fast fatigable = meczące się
Morfologia mięśnia szkieletowego
Włókna mięśniowe -> pierwotne pęczki mięśniowe połączone śródmięsną -> otoczone omięsną wewnętrzną i zewnętrzną -> cały mięsień otoczony jest namięsną -> a ta: powięzią
MIĘSIEŃ
brzusiec (włókno mięśniowe)
ścięgno -> na jednym końcu lub na obu końcach łączy mięsień ze szkieletem
włókno mięśniowe -> ułożone równolegle między końcami mięśnia
Budowa włókna mięśniowego
Miofibryle dzielą się na miofilamenty = białka kurczliwe: miozyna, aktyn, tropomiozyna
Poprzeczne prążkowanie = różne załamanie światła poszczególnych części włókien mięśniowych.
Budowa i funkcje sarkomeru
Sarkomer - jednostka czynnościowa komórki mięśniowej mięśnia poprzecznie prążkowanego. Jest to część komórki mięśniowej: 1. ograniczona 2 liniami Z
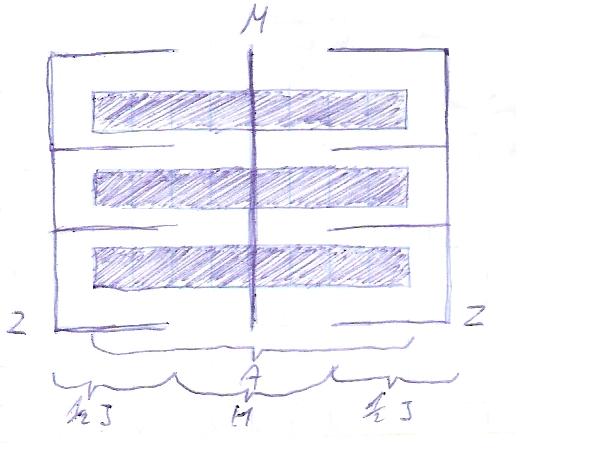
2. w części centralnej mamy prążki:
prążek A (ciemny = anizotropowy) = miozyna
prążek I (jasny = izotropowy) = aktyna
prążek H - ta część miozyny, która nie zawiera głów i nie kontaktuje się z aktyną
prążek M - centrum sarkomeru, zgrubienie miozyny
Sarkomer w spoczynku

MIOZYNA
2 - częściowa główka zawiera ATP-azę i przez szyjkę jest ona wiązana ruchomo z trzonem (ogonem) = meromiozyna lekka
Ruchomość odcinka główka-szyjka pozwala na tworzenie odwracalnego wiązania miozyny z aktyną /KOMPLEKS AKTYNO-MIOZYNOWY/ i wzajemnie przesuwanie się aktyny z miozyną. Na głowie jest miejsce wiązania dla: aktyny, ATP, ATP-azy
AKTYNA
Białko globularne: 2 łańcuchy splecione ze sobą = FILAMENT AKTYNOWY - wokół jest nitka TROPOMIOZYNY (TRM) - co 40 nm cząsteczka TROPONINY (TN) = 3 podjednostki
TN-C - wiąże jony Ca2+
TN-T - wiąże się z tropomiozyną
TN-I - zapobiega tworzeniu się mostków między aktyną a miozyną
Przy [Ca2+] >105 mol/litr - powoduje to aktywację białek i tropomiozyna odblokowuje miejsce przyczepienia miozyny do aktyny.
Po połączeniu głów uwalniany jest fosforan i powoduje to zgięcie głowy miozyny i wciągnięcie do wnętrza sarkomeru włókna aktyny. Gdy do głowy miozyny przyczepi się kolejna cząsteczka ATP zachodzi rozkurcz (odłączenie głów od siebie)
UNERWIENIE CZUCIOWE I RUCHOWE MIĘŚNIA SZKIELETOWEGO
Unerwienie czuciowe = włókna rdzenne (zmielinizowane) czuciowe - do wrzeciona mięśniowego (2-10 włókien mięśniowych śródwrzecionowych = INTRAFUZALNYCH otoczonych łącznotkankową torebką) - ułożone równolegle do reszty włókien mięśniowych = POZAWRZECIONOWYCH (ekstrafuzalnych) roboczych = WŁÓKNA DOŚRODKOWE = CZUCIOWE Ia i II
Unerwienie ruchowe
Agamma = „mały ruchowy układ nerwowy” = neurony ośrodkowe = ruchowe = motoneurony z układu siatkowatego zstępującego pobudzającego
Alfa = z przednich rogów rdzenia kręgowego lub odpowiednich jąder nerwów czaszkowych.

Przekazanie stanu pobudzenia z układu nerwowego na komórkę roboczą mięśnia odbywa się w tzw. PŁYTCE MOTORYCZNEJ = KOŃCOWEJ
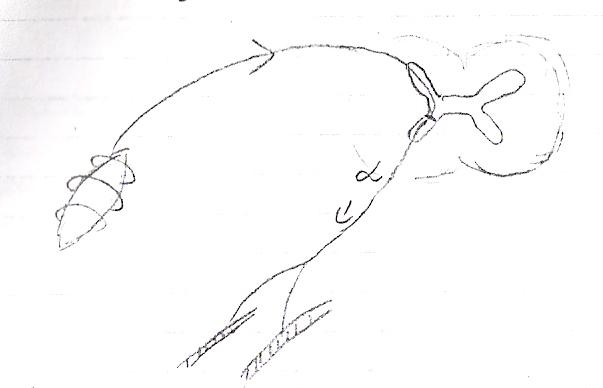
Rozciągnięcie mięśnia szkieletowego powoduje rozciągnięcie woreczkowatej okolicy jąder (WOJ). Powoduje to pobudzenie odkształconych receptorów pierścienno-spiralnych, w wyniku czego następuje generowanie potencjału czynnościowego we włóknie czuciowym I a. Przebiega ona drogą korzeni grzbietowych do rdzenia kręgowego. Następuje przełączenie na włókna ruchowe α, które drogą korzeni brzusznych rdzenia kręgowego trafiają do komórek roboczych mięśnia szkieletowego. Tu w pobliżu swojego zakończenia traci osłonkę mielinową i dzieli się na kilka gałązkę, z których każda unerwia jedną komórkę mięśniową. Zakończenia zawierają pęcherzyki z Ach (mediator). Te pęcherzyki wchodzą w zagłębienia PŁYTKI RUCHOWEJ (zgrubiała część błony komórkowej - sarkolemmy) = ZŁĄCZE NERWOWO MIĘŚNIOWE
Potencjał czynnościowy w motoneuronie α -> bodziec -> uwolnienie Ach z pęcherzyków synaptycznych do przestrzeni synaptycznej -> połączenie cząsteczki Ach z receptorem nikotynowym N w płytce motorycznej -> depolaryzacja (dokomórkowy prąd Na+)
TRIADA MIĘŚNIOWA
cewka poprzeczna T
zbiorniki końcowe
cewki podłużne
1
Wyszukiwarka
Podobne podstrony:
FIZJOLOGIA Układ Nerwowy (wyklady)
Fiziologia -UKŁAD NERWOWY (wykłady), Wykłady, FIZJOLOGIA
patologia ukl nerwowego wyklady, Pedagogika, Wczesne wspomaganie rozwoju, wczesne wspomaganie rozwoj
nerwowy wykład, analityka medyczna, anatomia, ukł nerwowy
w2.Farmakologia ośrodkowego obwodowego ukl nerwowego, wykłady PMWSZ w Opolu - Pielęgniarstwo, Farmak
autonomiczny układ nerwowy wykład
Uszkodzenie rdzenia kręgowego i korzeni nerwowych, Wykłady
Układ nerwowy wykłady prof Woźniaka
Zakazenia ukladu nerwowego wyklad doc
AUTONOMICZNY UKŁAD NERWOWY fizjologia (wyklady)
WYKŁAD- UKŁAD NERWOWY, Medyczne, Studia pielęgniarstwo, Fizjologia
Wykłady fizjologia mięśniowo nerwowy 13 (1)
Wyklad 7 ukl.nerwowy, Dietetyka, Anatomia i fizjologia człowieka, Fizjologia wykłady
Fizjologia układu nerwowego czI WYKŁAD
Dział 2 - Układ nerwowy, dietetyka, 1rok, 2 semestr, fizjologia, wykłady
AUTONOMICZNY UKŁAD NERWOWY fizjologia (wyklady)
Wykład (24 10 2011) Fizjologia ukł nerwowego
WYKLAD fizjologia serca
więcej podobnych podstron