Genetyka
4/5. OGÓLNA CHEMICZNA I FIZYCZNA CHARAKTERYSTYKA DNA
- DNA jest to nosnik informacji genetycznej we wszystkich org.
- jest podwojna nicia, skrecona prawoskretnie ( lewoskretna uzyskujemy w war.labolat.)
- wlasciwosci dzieki ktorym pelni funkcje matrycy inf genetycznej :
1. prosta budowa chemiczna
2. nukleotydy jako podstawowe cegielki budowlane , które zbudowane sa z trzech czlonow : zasad azotowych ktore sa na zewnatrz spirali; deoksyryboz i gr.fosforanowych ktore sa umieszczone na zewnatrz
3. lancuchowy charakter
-zasady dzielimy na pirymidynowe : C i T ; purynowe A i G
-istotne cechy modelu Watsona i Cricka :
1. DNA jest 2-niciowa czast. o ksztalcie komplementarnie skreconej spirali;
2. lancuchy lacza sie ze soba za pom. wiazan wodorowych miedzy zasadami w parze : A laczy sie dwoma z T, G trzema z C
3. srednica helisy wynosi 2 nm a na jeden pelny skret przypada 10 par nukleotydow
-kopiowanie DNA czyli replikacja ma charakter semikonserwtywny
-w czasie replikacji powst. widelki replikacyjne a do powielania DNA sa potrzebne zlozone ukl. enzymatyczne
-replikacja prowadzi do powielania DNA czyli z kazdej dwuniciowej czast. powst. dwie potomne o identycznej sekwencji nukleotydowej. Zapewnia to ciaglosc trwania DNA.
-DNA posiada temp. topnienia ,dochodzi do rozdzielenia nici
-DNA mozemy zwazyc ,mozemy wyrazic il. par zasad ,dlugosc mierzymy w nm
-DNA moze wyst. w jadrze kom ,plastydach ,mitochondriach
6. KOD GENETYCZNY I JEGO CHARAKTERYSTYKA
KOD GENETYCZNY- proste reguly wg. ktorych zaszyfrowana jest inf. genetyczna. Do jego najwazniejszych cech naleza :
-trojkowy charakter - podst. jedn. inf. o stalej wielkosci jest zawsze trojka nukleotydow (kodon) oznaczajaca aminokwas
-nienakladanie sie trojek czyli kodonow
-bezprzecinkowosc - pomiedzy trojkami nie ma dod. znakow
- jednoznacznosc - konstrukcja kodu i spos. jego realizacji pozwalaja na precyzyjne i zawsze takie same odczytanie inf. i zbudowac na tej podst. okreslone bialka. Dana trojka zawsze ozn. tylko JEDEN rodzaj aminokwasu.
-degeneracja - jeden aminokw. moze byc zakodowany przez kilka trojek
-uniwersalnosc - we wszystkoch organizmach reguly kodu sa takie same. Nie oznacza to wszakze ze wszystkie org. maja taka sama inf. genetyczna ,wrecz przeciwnie!
-kolinearnosc - danej kolejnosci kodonow zawsze podlega dana kolejnosc aminokw. w bialku
-posredni charakter - matryce DNA nigdy nie sa bezposrednio wykorzystane do ukladania aminokw.
AUG - trojka kodujaca metionine u eucariota i formylometionine w procariota. Znaczenie tego kodonu jest ogromne poniewaz synteza kazdego bialka zaczyna sie od znalezienia trojki startowej czyli AUG i wstawienia aminokwasu startowego.
UAA, UAG, UGA trojki te nie koduja zadnego aminokwasu i okresla sie je jako kodony nonsensowne (terminalne, STOP). Ich zadaniem jest wyznaczenie miejsca zakonczenia syntezy bialka.
7. GENOM - CHARAKTERYSTYKA I ORGANIZACJA
GENOM - podstawowy komplet informacji genetycznej. Jest pewna funkcjonalna caloscia bo zawiera niezbedny komplet inf. gen. potrzebnej do funkcjonowania organizmu. Im bardziej zaawansowany ewolucyjnie organizm tym wiekszy jego genom. U eucariota genom jest pojedynczy - haploidalny ,u procariota - diploidalny czyli podwojny. W tym przypadku instrukcja genetyczna wyst. w dwoch kopiach. W duzym uproszczeniu mozna wyroznic trzy poziomy organizacyjne ukladow biol. :
1. w prostych formach genomy sa male i nie zawiaraja informacji niezbedznej do samodzielnego funkcjonowania
2. w organizmach prokariotycznych ilosc inf. genetycznej i jej organizacja pozwala juz na funkcjonowanie ukl. o komorkowym poziomie zlozonosci ,zdolnych do samodzielnego zycia.
3. pojemnosc informacyjna genomow eukariotycznych i co wazne sposob jej organizacji umozliwiaja funkcjonowanie na b. Wysokom poziomie zlozonosci takim ze u wiekszosci zachodzi proces roznicowania sie komorek w tkanki i organy. Nie oznacza to jednak ze kazdy org. Eukariotyczny jest wielokomorkowcem!!!
W czasie podzialow kom. material jadrowy organizuje sie w oddzielne twory - chromosomy. Sa to bardzo skomplikowane „paczki” zawierajace matrial genetyczny. Ilosc takich „paczek” jest cecha gatunkowa, np. czlowiek posiada 46, muszka owocowa 8. Graficzna prezentacje garnituru chromosowego danego organizmu nazywamy kariotypem (scislej - idiogramem).
Poniewaz chromosomy zawiraja zarowno bialka jak i kwasy nukleinowe ( glownie DNA) oba rodzaje makroczasteczek wspoluczestnicza w organizacji genomu tworzac chromatyne.
Chromatyne dzielimy na euchromatyne : gdzie chromosomy sa aktywne genetycznie, i heterochromatyne : jest nieaktywna. Stan chromosomalny chromatyny oraz histony wyst. tylko w DNA jadrowym.
9. RODZAJE SEKWENCJI DNA
Sekwencje - ulozenie nukleotydiow w DNA.
Organizmy najprostsze maja DNA w postaci unikalnej i niepowtarzalnej
-bakterie - 99,7% unikalnek sekwencji
-slimak - 40%
-ropucha - 55%
-mysz - 60%
-czlowiek - 70%
sekwencje : 1. unikalne, 2. srednio czesto powtarzalne, 3.czesto powtarzalne
AD.1. sekwencje unikalne : w matrycach genetycznych wirusow i bakterii , powtarzaja sie tylko raz w calym genomie.
Występowanie licznych kopii poszczególnych genów nie jest cechą wyłącznie eukariontów; E. coli, na przykład, ma siedem genów dla rRNA, prawdopodobnie doprzeprowadzania szybkiej syntezy dużej liczby rybosomów. Transkrypcja jednego tylko genu dostarczałaby rRNA zbyt wolno, by zaspokoić potrzeby gwałtownie rosnącej komórki. Eukarionty również mają liczne kopie genów do rozwiązywania podobnych problemów zaopatrzeniowych; na przykład w typowym genomie ssaka jest kilkaset genów dla rRNA. Jednak eukarionty wykorzystują liczne kopie genów do innych celów.
Kolejną część nadmiarowego DNA stanowią liczne niefunkcjonalne kopie genów, tak zwane pseudogeny. Pseudogeny zazwyczaj nie mogą ulegać ekspresji albo ze względu na liczne delecje (ubytki fragmentów sekwencji kodującej), albo z powodu mutacji uniemożliwiających syntezę białka, lub też brak odcinków promotorowych bądź sekwencji regulatorowych. Liczba pseudogenów jest różna dla różnych genów (od 1 do 1000) i zazwyczaj zmienia się w zależności od gatunku - wyjątkowo wysoka jest u ssaków.
Okazuje się zatem, że w genomach eukariotycznych sekwencja genu może występować tylko raz lub też należeć do całej rodziny sekwencji powtórzonych. Członkami tej rodziny może być kilka funkcjonalnych genów, które są ze sobą blisko spokrewnione, ale ulegają ekspresji w różnych okresach rozwoju lub w różnych tkankach bądź komórkach, albo kilka pseudogenów, albo jednych i drugich. Bywa, że są zebrane razem w jednym regionie genomu albo rozrzucone nawet po wielu różnych chromosomach.
Inną przyczyną wielkości niektórych genomów eukariotycznych jest występowanie rodzin sekwencji, które powtarzają się wielokrotnie, ale o ich znaczeniu ciągle niewiele wiemy. Klonowanie molekularne i sekwencjonowanie potwierdziło istnienie takich licznie występujących sekwencji DNA, odkrytych wcześniej dzięki klasycznym technikom biochemicznym. Rodziny powtarzających się sekwencji mogą zawierać setki tysięcy, a nawet miliony powtórzeń, i zwykle stanowią 10%, a często nawet niemal 50% genomu. Długość powtórzonej jednostki wynosi od dwóch do wielu tysięcy par zasad. Podobnie jak powtórzone geny, powtarzające się sekwencje albo występują bezpośrednio po sobie (na przykład: 5'-AGGAGGAGGAGG-... i tak dalej), albo są rozrzucone po różnych miejscach chromosomu.
Techniki analizy molekularnej umożliwiły stwierdzenie, że pewne rodzaje sekwencji DNA występują w okolicy charakterystycznych struktur chromosomów, takich jak centromery, telomery czy organizatory jąderka. Na przykład region chromosomu nazywany organizatorem jąderka zawiera ciąg powtórzeń genów dla rRNA.
Telomery znajdują się na końcach długich liniowych dwuniciowych cząsteczek chromosomalnego DNA. U wszystkich eukariontów sekwencje telomerowe są bardzo podobne. Zawierają różną liczbę krótkich, następujących po sobie powtórzeń, takich jak: 5'-CCCTAACCCTAACCCTAA... 3'-GGGATTGGGATTGGGATT... itp.u ludzi lub: 5'-CCCCAACCCCAACCCCAA... 3'-GGGGTTGGGGTTGGGGTT... itp.u kilku pierwotniaków. Telomerowe powtórzenia są dodawane na końcach nowo powstałego DNA dzięki specjalnemu enzymowi. Te szczególne sekwencje terminalne pozwalają komórce odróżnić prawdziwe telomery od pękniętych chromosomów. Pęknięte chromosomy są często łączone w przypadkowy sposób, telomerowe zakończenia stanowią jednak pewne zabezpieczenie przed takimi niepożądanymi zdarzeniami.
AD.2. sekwencje srednio czesto powtarzalne-wystepuja w calym genomie i sa rozdzielone sekwencjami unikalnymi
AD.3. sekwencje powtarzalne z wysoka czestotliwoscia wystepuja w :
· heterochromatynie - zgrupowane kolo centromerow
· koncach chromosomow
· satelitarnym DNA
sekwencje powtorzeniowe
1. wystepuja w tandemowych blokach lub sa rozproszone
2. w zaleznosci od miejsca wystepujaw chromosomie:
· sekwencje tworzace duze tandemowe bloki w DNA satelitarnym, 4 frakcje, w poszczegolnych frakcjach wyst.homologoiczne sekwencje dziki ktorym identyfikujemy chromosomy, wystepuja u czlowieka, zwierzat roslin
· sekw.tworzace male tandemy zgrupowane lub rozproszone w minisatelity stosowane do diagnostyki molekularnej ( np. ustalanie ojcostwa )
· sekwencje mikrosatelitarne tworzace proste tandemy rozproszone rownomiernie po genie. Te sekewncje moga byc 1-nukleotydowe,2,3,4,5, wykorzystywane do diagnostyki molekularnej i mapowania genow.
10. POZIOMY REGULACJI AKTYWNOSCI GENET. REPLIKACJA DNA
Replikacja prowadzi do podwojenia ilosci DNA. Dzis wiemy ze replikacja DNA ma charakter semikonserwatywny - w takim spos. kopiowania dochodzi do rozplecenia podwojnej spirali cząst. macierzystej, jednak bez rozkladu jednoniciowych lancuchow na mniejsze fragmenty. Nastepnie do kazdej ze „starych” nici dosyntezowywane sa nowe.
Replikacja katalizowana jest enzymatycznie przez enzym „odczytujacy” matryce jednak wkrotce po jego odkryciu okazalo sie ze polimeraza DNA spelnia jedynie funkcje pomocnicze, np uzupelnia brakujace albo naprawia uszkodzone fragmenty DNA.
1. Poczatek procesu - miejsce inicjacji replikacji : w kazdej czast.DNA jest jeden specjalny odcinek ktory ma specyficzna sekwencje nukleotydow i jego sekwencja to cos w rodzaju zapisu „ tu zaczynac replikacje”. Nazywa sie z ang. Origin, skrot - ORI
2. Rozdzielenie nici bez zniszczenia struktury 1 rzedowej. U bakterii e-coli bialko inicjatorowe Dna-a wykorzystuje energie z ATP i rozrywa wiazania wodorowe w miejscu replikacji. W to miejsce wchodzi bialko Dna-b ktore wypiera bialko inicjatorowe i rozplata nic w obie strony. W ten sposob rozwiera sie oczko i powstaja widelki replikacyjne. Teraz na „teren budowy” wchodzi bialko primazy, ktore syntezuje primery - odcinki startowego RNA. Utworzenie primerow konczy etap inicjacji.
3. wstawienie nowych nukleotydow i korekta popelnionych bledow, etap elongacji. Wiernosc kopiowania wymaga aby synteza przebiegala w jednym kierunku. Poniewaz nici w DNA sa anyrownoległe to na jednej zachodzi ciagla syteza a na drugiej nici (tzw. opoznionej) tworzone sa tzw fragmenty okazaki, każdy fragment okazaki składa się z primera RNA i krótkiego odcinka DNA o długości około 200 nukleotydów.
4. Zakonczenie czyli terminacja replikacji. Po zakończeniu replikacji całej cząsteczki primery są wycinane z nowych nici polinukleotydowych, powstałe "dziury" są wypełniane przez polimerazę DNA, a końce tych fragmentów są łączone w jeden ciagly łańcuch przez ligazę DNA.
11. PODSTWY MOLEKULARNE MUTACJI
wyrozniamy 3 rodzaje zmiennosci ktore zasadniczo sie roznia od siebie sposobem w jaki powstaja, jednak zawsze prowadza do zmian w orgaznizmie:
1. zmienność fluktuacyjna:
· nie jest dziedziczna
· jest wynikiem interakcji genotypu i środowiska
2. zmienność rekombinacyjna
· jest dziedziczna
· jest wynikiem kombinacji genow
· np. crosing-over
3. zmienność mutacyjna
· jest to zmiana dziedziczna powstajaca nagle, skokowo
Mutacje DNA moga powstawac przez bledy w proc.replikacji a takze w wyniku dzialana czynnikow fizycznych i chemicznych. Powstale zmiany w strukturze DNA moga wywolac rozne mutacje, badz zaburzyc replikacje DNA i spowodowac smierc komorki. W organizmach wystepuja setki genow i kodonow ktore poprawiaja bledy i regeneruja uszkodzenia w DNA dzieki czemu jest utrzymywana wzgledna stalosc sekwencji nukleotydowej.
Powstawanie i utrwalanie sie mutacji jest koniecznym warunkiem utrzymania sie zmiennosci dziedzicznej organizmow jako motoru ewolucji.
Istotna role w wytwarzaniu duzych zmian w strukturze DNA moga odgrywac ruchome elementy genetyczne, zmieniające swą lokalizację w obrębie genomu, ktore odkryto w latach 50 i nazwano TRANSPOZONAMI.
Z przemieszczaniem się transpozonów mogą być związane bardzo poważne zmiany w DNA: delecje, inwersje i duplikacje obejmujące bardzo nieraz obszerne rejony i powodujące nieraz utratę funkcji niektórych genów, a wzmożoną ekspresję innych. Zmiany w strukturze DNA stanowia zmiany premutacyjne ktore jesli nie zostana usuniete w wyniku reperacji, moga doprpwadzic do zmian mutacyjnych w nast. cyklach replikacyjnych DNA. Mutacjami dziedzicznie przekazywanymi beda te zmiany w DNA ktore maja inna wyjsciowa sekwencje nukleotydow :
- zmiany w helisie jednej pary zasad w druga (najczestsze punktowe)
- wypadniecie (delecja) pary zasad lub odwrotnie - wstawienie dod. pary zasad (insercja). Powstaja znacznie rzadziej.
Mutacje punktowe moga miec rozne efekty - zmiane jednego kodonu w drugi, ktory wyznacza inny aminokwas lub ten sam - wtedy brak wplywu na bialka. Duze zmainy w sekwencji nukleotydowej calkowicie eliminuja ekspresje roznych genow (np. dluga delecja).
12. PRAWA MUTACJI WG. DE VRIESA
Termin mutacja wprowadził do genetyki 1901 H. de Vries. Mozemy uwazac de Vriesa za pierwszego badacza, ktory postawil zagadnienie powstawania nowych form na gruncie eksperymentalnym. Na podstawie bogatego materialu doswiadczalnego de Vries opracowal tezy dotyczace powstawania nowych drobnych gat. , ktore mozna nazwac prawami mutacji :
1. Nowe drobne gat. powstaja nagle, bez posrednictwa form przejsciowych;
2. Nowe drobne gat. sa przewaznie ustalone od chwili ich powsatnia;
3. Wiekszosc nowo powstalych form ma cechy drobnych gat. a nie odmian (formy rozniace sie jedna cecha od formy wyjsciowej);
4. Drobne gat. pojawiaja sie w pewnej liczbie osobnikow rownoczesnie lub w tym samym okresie czasu;
5. Mutacje sa bezkierunkowe. Zmiany mutacyjne obejmuja rozne organy i zachodzic moga prawie we wszytskich kierunkach.
13. RODZAJE MUTACJI
Mutacja - nagła, skokowa zmiana mat. genetycznego, ktora moze sie dzidziczyc. Mutacje maja charakter losowy i bezkierunkowy (zmiany powst. bez zadnego planu i z cecha „wszystko sie moze zdarzyc”.
Mutacje roznia sie skala zmiany, zmainie moze ulegac niewielki fragment DNA jak i caly genom. Najprostsza klasyfikacja zaklada wielkosc zmiany :
1. Mutacje genowe (punktowe) - w krorych zmiana sekwencji nukleotydowej odbywa sie na odcinku DNA mniejszym niz jeden gen. najczesciej jest to tylko (?) zmiana pojedynczej pary nukleotydow lub sekwencji niewiele dluzszej. Nie mozna ich diagnozowac przy uzyciu mikroskopu.
2. Chromosomowe (aberracje chromosomowe) - takie w ktorych zmianie ulega struktura jednego chromosomu. Mutacje o zasiegu wiekszym niz jeden gen a mniejszym niz jeden chromosom.
3. Genomowe - polegajace na zmianie liczby kompletnych chromosomow. Tego rodzaju mutacje mozna stwierdzic analizujac kariotypy pod mikroskopem.
AD.1. Mutacje punktowe mozna podzieli wg. bardzo prostego klucza :
A. Wynikajace z podstawienia wlasciwej zasady przez inna - nzw.to substytucja.Zastapienie jednej puryny druga lub pirymidyny pirymidyna np.G przez A nazywamy tranzycja. Jesli Zasieg mutacji ogranicza sie do samej zmiany , poniewaz w substytucji nie ulega przesunieciu ramka odczytu.
· Mutacje milczace (ciche) - zmiana zasady na inna moze wypasc akurat w trzeciel literze danej trojki. W ten spos. sekwencja nie ulegnie zmianie.
· Mutacje nonsensowne - zmiana jednaj zasady w trojce moze zmienic ja w trojke nonsensowna czyli znak STOP i moze zostac zahamowana translacja DNA. Czesto maja woplyw na aktywnosc kodowanego bialka lub prowadza do powstawania zmutowanego fenotypu.
· Mutacje zmiany sensu - zmiana jednej zasady w trojce kodujacej moze moze zmienic ja w inna trojke sensowna przez wbudowanie innego aminokwasu. Efekt moze byc bardzo rozny. Wystepuje zwykle w jednaj z dwoch pierwszych zasad kodonu. Podobne aminokwasy maja podobne kodony , np w trojke AUC podstawi sie G zamiast U i jesli taka zmiana zajdziew czesci lancucha polipeptydowego o malej roli , moze nieznacznie wplynac na funkcje tego bialka. Jesli jednak bylo inaczej skutkiem moze byc np.anemia sierpowata. W ten spos. Powstaja tez nowe bialka co ma znaczenie w ewolucji.
B. mutacje wynikajace z utraty (delecja) badz wstawienia nukleotydu (insercja). Z uwagi na sposob odczytu inf. genetycznej opuszczenie lub dodanie zmienia ramke odczytu az do konca sekwencji kodujacej. Inaczej mowiac poczynajac od miejsca defektu wszystkie trojki zmieniaja swoj uklad. Skutek jest tak , ze :
· polipeptyd jest syntezowany ale sa przylaczane inne aminokwasy i ostatecznie powstaje inne bialko - najczesciej bezuzyteczne.
· polipeptyd jest syntezowany ale sa przylaczane inne aminokwasy i ktoras z kolejnych „nowo powstaluych' trojek jest sygnales STOP (trojka nonsensowna). Synteza zostanie przerwana a krotki lancuch nie bedzie spelnial zadnej funkcji biologicznej.
AD.2. Mutacje chromosomowe dzielimy na :
A. Wewnatrzchromosomowe - gdy zmiany ograniczaja sie do jednego chromosomu :
· Dficjencje - utrata fragmentu chromosomu. Dochodzi do tego gdy peknie chromosom, a jedna z powstalych czesci ulega degradacji i eliminacji.
· Duplikacje - zdublowanie fragmentu chromosomu. Odgrywaja niebagatelna role w proc. Ewolucji.
· Inwersje - obrocenie odc. chromosomu o 180 `
B. Miedzychromosomowe - translokacja - przeniesienie odc. z jednego chromosomu na drugi niehomologiczny.
AD.3. Mutacje genomowe zmieniaja liczbe chromosomow.
Ta kategoria obejmuje najwieksze zmiany mat. genetycznego. Powstale w ich wyniku mutanty dzielimy na :
A. Autoploidy - org. powstajace na skutek mutacji osobn. jednego gat. dzielimy je na :
· Aneuploidy - org. o zmienionej l.pojedynczych chromosomow, np u muszek owocowych, mutacje wynikaja z nierozchodzenia sie chromosomow homologicznych w czasie mejozy - nondysjunkcja
· Monosomiki (2n-1) - zamiast pary chromosomow homolog. jest tylko jeden w zygocie.
· Trisomiki (2n+1) - zamiast dwoch sa trzy.
B. Alloploidy - powstale z polaczenia dwoch roznych genomow - z odmiennych gat. najczesciej nie wytwarzaja zywotnych gamet bo nie maja chromos.homologicznych ktore moglyby ze soba koniugowac.
14. PRZYCZYNY POWSTAWANIA MUTACJI
Mutacje moge powstawac:
1. Samorzutnie (spontanicznie)bez udzialu czynn. Fiz. I chem. Wtedy powst. mutacje i najmniejszej skali zmiany. Czestosc zachodzenia tych mutacji jest b.mala i moga nastepowac dwojako :
A. Na skutek przypadkowego oddzialywania trudno uchwytnych czynnikow zewnatrz- i wewnatrz-komorkowych na konformacje przestrzenna aparatu replikacyjnego.
B. Jako konstrukcja plynaca z samej natury. Kazdy proc.biologiczny daje jakis odsetek bledow wynikajacej z samej zlozonosci tego przebiegu..( np. bledne funkcjonowanie polimerazy DNA, usuniete enzymy reperujace )
2. W sposob indukowany przy udziale czynnika fizycznego lub chemicznego i zachodza w naturalnych jak i labolatoryjnych warunkach. Do czynnikow mutagennych zaliczamy :
A. fizyczne :
· Promienie jonizujace, rentgenowskie i gamma niosa duza porcje energii ktora jest wchlaniana przez skladniki DNAi czasteczki te ulegaja uszkodzeniu - najczesciej rozerwaniu.
· Promieniowanie ultrafioletowe (UV) sa szczegolnie niebezpieczne dla malych organizmow i dla wierzchnich warstw ciala czlowieka. Promieniowanie tego rodzaje wplywa na wzrost miedzy pirymidynami lezacymi obok siebie. Szczegolnie czestko kolo tyminy - powstaja dimery tymidynowe. Przeszkadzaja one w odczycie DNA
· Wysoka temperatura - ma wplyw na jakosc reakcji i tempo pracy enzymow.
B. chemiczne :
· kwas azotawy powoduje oksydacyjna dezaminacje. W ten spos. Adenina zamienia sie w hipoksantyne ktora zachowuje sie jak guanina. Ostatecznie zamiast pary AT powstaje GC
· Ipertyt ten gaz bojowy modyfikuje zasady azotowe
· Barwniki akrydynowe - ich czasteczki wciskaja sie miedzy pary nukleotydow DNA, czego skutkiem jest wstawienie nowych lub utrata nukleotydow.
· Analogi zasad azotowych, np.bromouracyl - ich wlaczenie powoduje bledne odcztyt przez polimeraze DNA a przez tonieprawidlowe wstawianie nukleotydow.
· Nadtlenek wodoru i amoniak zw.o duzej aktywnosci chemicznej i moga np. reagowac z czasteczkami DNA
· Kolchicyna - alkaloid blokujacy wytw.wrzeciona kariokinetycznego i dochodzi do zaburzen w rozchodzeniu sie chromosomow
· Niektore weglowodany i ich pochodne, np. beznopiren - wystepujace np. w dymie papierosowym
Promieniowanie jaonizujace moze zwiekszyc czestosc mutacji nawet 1000-krotnie
15. CZESTOTLIWOSC MUTACJI
Normalnie to zjawisko jest bardzo rzadkie, szczegolnie u organizmow nizszysch. U wyzszych zachodzi czesciej. Szybkosc mutacji wzrasta gdy komorki sa wystawione na dzialanie mutagenow fizycznych lub chemicznych. Obserwowana w komorkach czestosc pojawien mutacji jest wyraznie nizsza niz ilosc powstajacych uszkodzen. Wiekszosc drobnych uszkodzen jest usuwanych w procesach reperacji, dotyczy to golwnie bakterii. U tych organizmow wiekszosc bledow powstajacych w czasie replikacji jest usuwana natychmiast przez polimeraze kornberga. Istnieje tez mozliwosc reperacji uszkodzonego DNA w innych sytuacjach.
16. MUTACJE SPONTANICZNE I INDUKOWANE
Spontaniczne powstaja na skutek dzialania czynn. wewnatrznych a indukowane na skutek mutagenow zewnetrznych.
17. CZYNNIKI MUTAGENNE. ICH ZNACZENIE I WYKRYWANIE
Skutecznosc dzialania mutagenow zalezy od : wilgotnosci, dostepnego tlenu, temp, stanu fizjologicznego (wiek,faza komorki ), dostepnosci (stezenia) tla genetycznego, skutecznosci selekcji.
Naprawa uszkodzen powstalych na skutek dzialania promieni UV ( np.u E.coli)
1. fotoreaktywacja - polega na uzyciu specjalnego enzymu - fotoliazy. Czasteczka ta wykorzystuje pochlaniana energie swietlna do rozrywania wiazan kowalencyjnych w dimerach np.tymidynowych. odtwarza wiec strukture DNA znoszac skutek mutacji
2. reperacja przez wycinanie - dimery i inne uszkodzenia moga byc wycinane na odcinku 20-30 nukleotydow. Warunkiem powodzenia jest brak uszkodzen na drugim pasmie DNA. Naprawa polega na tym ze specyficzny enzym endonukleaza nacina nic w poblizu miejsca uszkodzenia. Inny enzym usuwa caly rejon uszkodzenia i wstawia brakujace nukleotydy. Korzysta przy tym z matrycy jaka jest nic nieuszkodzona. Potem pozostaje polaczyc naprawiona nic w calosc przy uzyciu ligazy.
Naprawa uszkodzenpowodowanych przez czynniki chemiczne
1. endonukleazy
2. reperacja przez wycinanie i resynteze
18. PROMUTAGENY I ICH ZNACZENIE
Promutageny to bledy w replikacji. Powstaja one na skutek blednego dzialania polimerazy DNA. Moga byc naprawiane przez specyficzne kompleksy enzymatyczne lub prowadzic do zmian mutacyjnych ( glownie punktowych ).
Skutek mutacji mozna sklasyfikowac:
1. niekorzystne ( najczestsze)
· ograniczajace zdolnosc przezycia w kazdych warunkach - letalne
· zmniejszajaca wartosc adaptacyjna, tak ze w wawrunkach mniej korzystnych osobnik nie ma szans na przezycie - warunkowo letalne
2. neutralne - nie wywolujace zmian wartosci adaptacyjnej u osobnika. Nie sa takie rzadkie i zmiany takie maja istotne znaczenie w procesach ewolucyjnych
3. korzystne - zwiekszajace zdolnosci adaptacyjne osobnika. Zdecydowanie najrzadsze
19. MUTAGENNE CZYNNIKI SRODOWISKOWE
· naturalne ( pekniecia chromosomów)
· powstajace przez dzialalnosc przemyslowa
wykrywa sie je za pomoca roznych testow na mutacje indukowane:
· mikrobiologiczny test Amesa, stosowany powszechnie do oceny mutagenności lub kancerogenności danej substancji lub czynnika (salmonella)
· testy mutagennosci i kancerogennosci zwiazkow chemicznych przeprowadza sie tez na komorkach zwierzecych i na calych organizmach
21. MUTACJE AUKSOTROFICZNE
Inaczej mutacje pokarmowe
Przyczyna jest mutacja genow odpowiedzialnych za synteze enzymow ktroe biora udzial w procecach metabolicznych organizmu. Taka mutacja powoduje ze duzy organizm nie jest w stanie zsyntezowac np. jednago aminokwasu. W praktyce przejawia sie to tym ze organizm nie moze rowijac sie na pozywce mineralnej. Doswiadczenia wskazuja jednak na to ze uzupelnienie pozywki o element ktorego organizm nie moze zsyntezowac umozliwia normalny wzrost i rozwoj organizmu.
23. FAG LAMBDA - CHARAKTERYSTYKA
· jest to fag lagodny 9umiarkowany)
· nalezy do fagow przeprowadzajacych transdukcje ( przeniesienie inf.genetycznej z jednej baktrii do drugiej za pomoca baktreriofaga)
· to ze jest fagiem lagodnym oznacza ze nie musi koniecznie doprowadzic do smierci komorki bakteryjnej po uprzednim namnozeniu (cykl lityczny), moze natomiast wlaczyc sie do chromosomu bakteryjnego i byc razem z nim replikowany
· fag zintegrowany w chromosomie bakterii nazywamy PROFAGIEM
· profag jednak moze ulec oddzieleniu od DNA bakerii (np.pod wplywem czynnikow zewn.tj. promieni UV lub X) wtedy rozpoczyna sie cykl lityczny ktory konczy sie smiercia bakterii
24. GENOM FAGA LAMBDA - CHARAKTERYSTYKA
· DNA faga moze przybrac dwie formy : albo kulista albo liniowa
· Gdy DNA jest w wirusie wtedy jesy liniowe, gdy w bakterii - kuliste lub liniowe
· Zmiana formy liniowej w kolsta jest mozliwa dzieki enzymom, ktore powoduja pekniecie zarowno jednej nici (+) jak i drugiej (-). Pekniecia nie powstaja w tym samym miejscu, ale b.blisko siebie. To powoduje ze liniowe DNA faga na 2 jednoniciowe konce, ktore sa w stosunku do siebie komplementarne. Nazywamy je lepkimi koncami i dzieki temu froma liniowa moze zmienic sie w kolistai odwrotnie. Wymaga to tylko wytworzenia wiazan wodorowych pomiedzy komplementarnymi koncami.
· Koliste DNA faga wystepuje u bakterii w momencie gdy jest ona w fazie litycznej, w fazie lizogenicznej jest forma liniowa wbudowana w chromosom bakterii (profag)
· „lepkie” konce sa wolne o dlugosci do 12 nukleotydow
25. REGULACJA LIZOGENICZNOSCI NA POZIOME MOLEKULARNYM
etapy cyklu lizogenicznego :
1. etap integracji
· polega na wbudowaniu DNA faga w konkretnym miejscu od DNA gospodarza
· powstaje profag - powoduje to brak cyklu litycznego
2. etap trwania :
· jedynym genem ulegajacym ekspresji jest gen C I
· gen C I odpowiada za kodowanie represora bialkowego
· represor z kolei blokuje ekspresje pozostalych genow procz genu C I to powoduje trwanie w fazie lizogenicznej
· ]jedyne inaktywacje represora umozliwia ekspresja innych genow co z kolei powoduje wyciecie profaga z DNA bakterii i rozpoczyna cykl lityczny
3. etap uwalniania :
· represor wykazuje duze powinowactwo do operonow O2 i O12
· represor laczac sie z nimi warunkuje zahamowanie ekspresji innych genow
· taki stan jest dosc slaby i umozliwia i umozliwia autoryzacje represora oraz cyklu litzogenicznego
· jesli profag przechodzi z kom.lizogenicznej do nielizogenicznej okazuje sie ze brak tam bialek represorowych i nie mam co juz utrzymywac fazy lizogenicznej inaktywacje moga powodowac rowiniez np.promieniowania
· pod wplywem enzymow profag zostaje wyciety z DNA bakterii i rozpoczyna sie cykl lityczny
26. CYKL LITYCZNY I JEGO CHARAKTERYSTYKA
Glownym ceklem tego cyklu jest namnazanie sie fagow. W przypadku faga litycznego cykl ten powodowany jest inaktywacja represora bialkowego.
Cykl życiowy bakteriofaga polegający na zakażeniu bakterii, produkcji nowych cząstek fagowych, rozpadzie bakterii i uwolnieniu nowych bakteriofagów.
W pewnych warunkach (promieniowanie jonizujące, promienie ultrafioletowe) następują zmiany w strukturze kwasów nukleinowych (fragmentacja, rozerwanie wiązań) oraz białek genomowych, co powoduje uwolnienie profaga (provirusa) z genomu. Profag namnaża się niezależnie od genomu gospodarza, następuje synteza komponentów białkowych wirusa i składanie ich w dojrzałe formy faga. Dochodzi wówczas do lizy, czyli rozpadu komórki bakteryjnej. Przykładem bakteriofaga łagodnego jest fag lambda. Fag lambda może jednak od razu wejść w cykl rozwojowy lityczny, z pominięciem cyklu lizogenicznego.
Na rysunkach przedstawiono cykl lityczny (normalny) trzech różnych wirusów. Kolejne etapy rozwoju wirusów oznaczono literami. Po przeanalizowaniu rysunków rozwiąż kolejno zadania 4a i 4b.
27. CHARAKTERYSTYKA I DZIALANIE OPERONU LAKTOZOWEGO
Schemat przedstawia operon laktozowy.
Geny struktury
regulator promotor operator X Y Z
Pojedyncza komorka bakteryjna jest zmuszona do czestych zmian substratow pokarmowych, a ze wzgledu na koszty energetyczne, nie moze sobie pozwolic na stala synteze wszystkich enzymow potrzebnych do ich rozkladu. Zatem w komorkach prokariotycznych musza istniec bardzo precyzyjne mechanizmy regulujace biosynteze tylko tych bialek, ktore sa w danej chwili niezbedne.
Jednym z najlepiej poznanych ukladow regulacyjnych u bakterii jest operon. Jego mechanizm jest oparty na oddzialywaniu bialka regulatorowego z okreslona sekwencja nukleotydowa DNA - operatorem . Operon mozna sobie wyobrazic jako grupe genow kontrolowanych przez wspolny system regulacyjny, majacych jeden promotor i transkrybowanych w postaci pojedynczego mRNA.
Dzialanie operonu mozna przedstawic na przykladzie operonu laktozowego, wystepujacego u paleczki okraznicy (Escherichia coli). Umozliwia on utrzymanie niskiego stezenia enzymow warunkujacych rozklad laktozy w warunkach jej braku oraz szybki wzrost stezenia przy pojawieniu sie substratu. Operon laktozowy sklada sie z trzech lezacych obok siebie genow kodujacych enzymy (lacZ, lacY, lacA) - tzw. genow struktury. W kierunku 5' do tych genow znajduje sie sekwencja DNA, zwana operatorem, z ktora wybiorczo moze sie laczyc bialko zwane represorem. Dodatkowo region operatorowy (operator) naklada sie piecioma nukleotydami na promotor genow struktury, znaczy to, ze dolaczenie represora uniemozliwia transkrypcje tych genow.
Istota mechanizmu regulacyjnego operonu jest to, ze zdolnosc laczenia sie represora z odcinkiem operatorowym jest zmieniana przez drobnoczasteczkowe zwiazki chemiczne. W przypadku operonu laktozowego, polaczenie sie laktozy do bialka regulatorowego (represora) powoduje obnizenie jego zdolnosci do wiazania sie z operatorem. W rezultacie zmieniony w ten sposob represor przestaje blokowac sekwencje operatorowa i umozliwia przylaczenie sie polimerazy RNA do promotora.
Represor jest wytwarzany w komorce bakteryjnej w sposob ciagly, nie zaleznie od tego, czy laktoza jest obecna w srodowisku, czy tez nie. Przy braku laktozy represor uniemozliwia przylaczenie sie polimerazy RNA i rozpoczecie transkrypcji. Pojawienie sie laktozy w srodowisku zmienia strukture czasteczek represora, umozliwiajac tym samym synteze mRNA z zapisana informacja o enzymach rozkladajacych laktoze. Po wyczerpaniu substratu, nie modyfikowane juz czasteczki represora znow lacza sie z operatorem, przywracajac wyjsciowa sytuacje.
Na uwage zasluguje fakt, ze operon laktozowy nie moze byc zaindukowany, kiedy w srodowisku srodowisku obok laktozy znajduje sie latwiej przyswajalne zrodlo wegla - glukoza. Musi wiec istniec mechanizm, dzieki ktoremu glukoza nie dopuszcza do indukcji operonu laktozowego. Zjawisko to nazwano represja glukozowa (ogolnie represja kataboliczna).
A TERAZ PO MOJEMU J
· O.L wystepuje u org. Prokariotycznych
· Umozliwia wykorzystanie konkretnej inf.genetycznej gdy jest ona potrzebna, w przypadku O.L. dotyczy to syntezy enzywmow potrzebnych do rozlozenia laktozy w taki sposob by mogla byc wykorzystana przez bakterie (E.coli)
· E.coli wykorzystuje laktoze jakos zrodlo wegla
· Jednak lepszym subrtratem dla bakterii jest glukoza i jesli glukoza i latkoza sa w srodowisku to glukoze wykorzystuja jako pierwsza
· FRANCOIS JACOB i JAQUES MOROD - naukowcy ktorzy opracowali teorie operonu laktozowego (Nobel)
Bakterie (Ecoli) syntezuja ok 800 roznych bialek. Niektore z nich syntezowane sa w sposob konstytuywny, np bialka szlaku glikolitycznego, podobnie bialka budulcowe, lecz wiekszosc genow bedzie potrzeban tylko przy specjalnych okazjach. Geny kodujace enzymy jednego szlaku metabolicznego czesto zorganizowane sa w operony ktore sa zdolne do jednoczesnego sterowania wieksza liczba genów. Laktoza jest wykorzystywana jako zapasowe zrodlo energii a poniewaz jest dwucukrem jej uzycie wymaga:
· Wprowadzenia czastek laktozy do wnetrza kom. Operacje ta przyspiesza enzym permeaza
· Rozlozenia czastki 2-cukrowej do monosacharydowych skladnikow : glukozy i galaktozy. Reakcje katalizuje beta-galakotzydaza
· Przeksztalcenia galaktozy w glukoze. Byc moze bierze w tym udzial transacetylaza galaktozydowa
Wazne jest to ze w przecietnych war. Ilosc tych enzymow jest b.niska, gdy komorke przeniesiemy z pozywki glukozowej na laktozowa to ilosc ich czasteczek wzrasta do prawie 5% masy wszystkich bialek kom. Jesli il.laktozy w pozywce jest niewielka to po wyczerpaniu zapasow tego zrodla energii stezenie w/w enzymow dosc szybko spada
.
Operon - odcinek ktory jest genem kodujacym bialka kontrolujace ekspresje genow enzymow przerabiajacych laktoze.
Geny trzech enzymow ( geny strukturalne ) polozone sa obok siebie a powyzej sekwencji 1 z nich kodujacego beta-galaktozydaze znajduje sie miejsce inicjacji transkrypcji ( promotor ). Do niego przylacza sie polimeraza RNA przepisujaca wszystkie trzy geny na jedna czasteczke mRNA. Pomiedzy promotorem a a genem pierwszgo enzymu znajduje sie jeszcze inny odc.DNA zwany operatorem. Odgrywa on zasadnicza role w ekspresji genow lezacych ponizej niego ( genow strukturalnych ). W ten sposob tworzy sie sekwencja DNA :
PROMOTOR-OPERATOR-GEN I-GEN II-GEN III
Pozostaje ona pod kontrola innego dosc odlegle polozonego genu ktory transkrybowany jest ciagle a powstajacy w mRNA ulega translacji. Jej produktem jest bialko ktore rozpoznaje przestrzennie sekwencje zasad operatora i oplaszcza sie wokolniego. Represor poniewaz blokuje polimerazie DNA dostep do sekwencji polozonych ponizej - blokuje transkrypcje genow strukturalnych. Gdy zachodzi potrzeba pobrania laktozy element blokujacy zostaje zablokowany :) . Bialko represora moze ulec allsoterycznej modyfikacji tak ze przestaje odczytywac sekwencje operatora i uwalnia go. W ten spos.polimeraza moze zaczac przesuwanie sie wzdluz odcinaka DNA.Czynnikiem ktory moze spowodowac dezaktywacje represora jest laktoza.. Dlatego w tej sytuacji nazywa sie derepresorem .
FUNKCJONOWANIE UKLADU ^ v
· W momencie gdy w srodowisku nie ma laktozy represor wyprodukowany przez gen regulatorowy (i) przylacza sie do operatora ( powinowactwo operatora do represora)
· To powoduje ze pomimo istnienia miejsca promotorowego transkrypcja nie zachodzi - uniemozliwia ja represor
· Jesli w srodowisku pojawia sie laktoza za jej stezenie jest duze wzrasta powinowactwo repsesora do laktozy (represor jest bialkiem allosterycznym ma 2 centra i przylacza sie jednym do operatora a drugim do laktozy)
· Przylaczenie laktozy do operatora powoduje odksztalcenie drugiego centrum aktwynego. Dziala to na zasadzie inhibicji odwracalnej w zal. Od tego ktorego substratu jest wiecej z tym sie laczy.
· Represor laczy sie z laktoza i odlacza od operatora dzieki temu mozliwa jest transkrypcja genow strukturalnych Z,Y,A
· Geny strukturalne odpowiedzialne sa za synteze odpowiednich enzymow
Gen Z ŕ beta - galaktordaza? - enzym ten powoduje rozpad laktozy na glukoze i galaktoze
Gen Y ŕ parmeaza? - beta-galaktorydoza? - umozliwia wprowadzenie laktozy ze środowiska zwewnetrznego do wewn. Na drodze transportu aktywnego
Gen A ŕ Ttransacetylacja galaktozydowa - akceptuje gestniejace galaktozydazy, rozklada galaktoze na glukoze
- do momentu gdy w kom.znajduje sie laktoza powtarza sie
- gdy laktozy nie ma represor przylacza sie do operatora i dzialalnosc genow Z,Y,A zostaje zahamowana, nie sa rowniez syntezowane niepotrzebne enzymy rozkladajace laktoze. NIC NIE ROZUMIEM :/
Operon laktozowy nalezy do operonow indukowalnych czyli uruchamialnych, ktore pierwotnie sa blokowane i ulegaja uaktywnieniu jedynie w specyficznych sytuacjach.
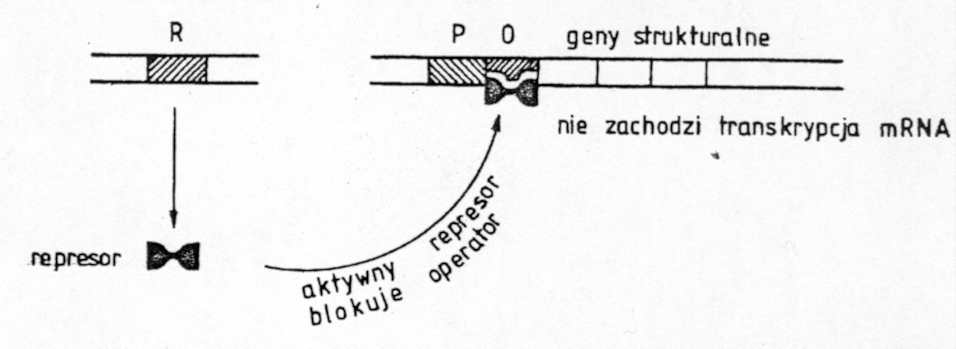
28. MUTACJE REPRESORA W OPERONIE LAKTOZOWYM LAC
1. Powstajace mutacje zwiazane sa z uszkodzeniami w DNA (zmiana sekwencji nukleotydow )
2. moga wynikac rowniez z tego ze polimeraza lubi sie mylic
3. poniewaz represor jest tworem genu regulujacego to jego mutacje beda zwiazane bezposrednio
4. z tym genem (i)
5. gen regulatorowy (i) lezy po lewej stronie nici DNA bakterii i jest oddalony od promotora, operatora i genow strukturalnych
6. wyrozniamy 4 rodzaje mutacji genu regulatorowego :
i - represor nie jest syntezowany. Powoduje to konstytuywna synteze enzymow nie ma bowiem represora ktory moze polaczyc sie z operatorem
is - mutacja superrepresjonujaca - powoduje ze represor ma szczegolnie duze powinowactwo do operatora i wynikiem tego jest sytuacja gdy represor nie oddziela sie od opertora nawet gdy w srod.jest laktoza, nie ma wiec syntezy enzymow. Jest to mutacja dominujaca nad normalnym stanem (i+ )
i-d - gen regulacyjny wytwarza represor ale jest on nieaktywny. Wynikiem tego jest konstytuywna synteza enzymow
i-q - na skutek mutacji promotora genu regulacyjnego nadmiar represorow. Wynikiem jest induktywna ( powodowana induktorem ) synteza
29. MUTACJE GENÓW STRUKTURALNYCH W OPERONIE LAC
Kazdy z tych genow odpowiedzialny jest za powstawanie innego enzymu i przeprowadzanie innej reakcji
· jesli Z -Y+A+ - pomimo aktywnosci Y i A brak podzialu na galaktoze i glukoze, z to w kom.jest duzo laktozy - dzialanie Y
· jesli Z-Y-A+ - brak podzialu brak laktozy w kom
· jesli Z-Y-A- - nic sie nie dzieje
· jesli Z+Y-A+ - jest podzial ale nie ma sprowadzonej laktozy do kom (?)
· jesli Z+Y-A- -jest podzial ale ani nie jest pobrana ani nie ma jej kto obrobic-galaktozy
Wszystkie geny musza funkcjonowac prawidlowo zeby proces byl pelny. Najgorzej wplywa brak lac2 bo aktywnosc YiA nie maja pozniej znaczenia. Przy braku Y po prostu mniej laktozy i szybciej sie wyczerpuje. Przy braku A galaktoza nie moze byc rozlozona.
Z - permeaza
A - transacetylaza galaktozydowa
Y - beta-galaktozydaza
30. MUTACJE CZESCI REGULATOROWYCH OPERONU LAC
Mutacje nastepuja rowniez w promotorze i operatorze pomimo ze nie sa one transkrybowane i reguluja ten proces.
m.o c - mutacja operatora polega natym ze represor nie moze dolaczyc sie do niego. Wynikiem jest synteza enzymow ciagla
m. p. - rozne mutacje promotora powodujaca synteze enzymow ale z rozna wydajnoscia, rozna wydajnosc transprypcji
31. ROLA CAMP W REGULACJI OPERONU LAC
Mamy tu doczyniena z nadrzednym systemem regulacji. Jesli w srod.znajduzje sie glukoza i laktozanajpierw bakt.wykorzystuje glukoze bo do wykorzyst.laktozy potrzeba energii. Zwiekszenie poziomu glukozy powoduje obnizenie zawartosci CAMP ( cykliczny adenozynomonofosforan ) i odwrotnie. Zwiekszenie poziomu CAMP powoduje aktywacje operonu laktozowego. AKTYWACJA operonu jest mozliwa dzieki temu ze CAMP laczy sie z bialkiem regulatorowym CAP (CRP) ,ktore w innym ukladzie jest nieaktywne. CAMP i CAP tworza kompleks, ktory laczy sie ze specyficznym odcinkiem DNA na obwodzie promotorowym. To polaczenie powoduje zmiane struktury promotora, represor odlacza sie od operatora i rozpoczyna sie transkrypcja genow. Samo bialko CAP nie wykazuje powinowactwa do promotara operonu lac, dopiero po polaczeniu z CAMP.
35. REKOMBINACJE SPECYFICZNE I NIESPECYFICZNE
NIE WIEM CZY DOBRZE :
Dzieki rekombinacjom zostaje wymieniony material genetyczny, powstaja nowe kombinacje
Istnieja 2 sposoby rekombinacji mat.genet. :
· crossing-over - Wymiana takich samych fragmentów chromosomów homologicznych. Zachodzi podczas profazy pierwszego podziału mejotycznego. Dzięki crossing-over zwiększa się liczba możliwych kombinacji genów znajdujących się w komórkach płciowych: geny położone na jednym chromosomie mogą być dziedziczone osobno.
· konwersja - rodzaj rekombinacji między dwoma blisko leżącymi homologicznymi regionami DNA, podczas której jedna z sekwencji DNA jest zmieniana tak, by pasować do drugiej; proces ten, nazywany też korektą sekwencji, może zachodzić wewnątrz jednego chromosomu lub między dwoma chromosomami
36. PODSTAWY CYTOPLAZMATYCZNEGO DZIEDZICZENIA
Dziedziczenie cytoplazmatyczne - (pozajądrowe) - dotyczy cech, których geny zlokalizowane są w mitochondriach i chloroplastach.
Dziedziczeniem pozajadrowym nazywac bedziemy te wszystkie zjawiska dziedzicznosci u organizmow euk., ktore uwarunkowane sa czynnikami zlokalizowanymi w DNA wystepujacym poza jadrem kom. Cechy wywolane czynnikami dziedzicznosci pozajadrowymi nie podlegaja mendlowskim prawom dziedziczenia wynikajacym z regularnej segregacji chromosomow w podzialam mitotycznych i mejot. DNA w kom.eukariot. wyst.poza jadrem glownie w chloroplastach i mitochondriach. Wiekszosc zbadanych zjawisk dziedziczenia pozajadrowego dotyczy genow wyst. w DNA mitochondrialnym (mitDNA ) badz chloroplastowym (chlDNA ).
Wykrywanie dziedziczenia pozajadrowego opiera sie na kilku prostych metodach. Jesli mieszance powstale w wyn. Krzyzowania odwrotnego miedzy dwiema f.rodzicielskimi posiadaja odrebne fenotypy to to czest moze to byc wskazowka efektow fenotypowych czynnikow pozajadrowych. W ten sposob mozna wykazac u szeregu roslin dziedziczenie pozajadrowe cech uwarunkowanych przez geny mieszczace sie w chlDNA.
Glowna metoda genetyczna wykrywania czynn.pozajadrowych jest analiza ich dziedziczenia w dalszych pokoleniach osobnikow heteroplazmatycznych. Zwykle cechy warunkowane czynn,pozajadr.segreguja sie w czasie podzialow miotycznych a nie wykazuja segregacji w czasie mejozy odwrotnie niz geny jadrowe.
A teraz w praktyce :
ENDOBIONTY BADZ WIRUSY JAKO CZYNNIKI DZIEDZICZENIA POZAJADROWEGO
Liczne gat.bakterii, glonow i wirusow wystepujacych w komorkach organizmow zwierz.czy rosl. Jako typowe organizmy pasozytnicze czy chorobotworcze moga czesto nie wywolywac wyraznych objawow chorobowych i nadawac gospodarzom nowe specyficzne wlasciwosci. Niektore tego rodzaju endosymbionty moga byc przekazywane przez gamety nastepnym pokoleniom i cechy wywolywane przez nie dziedzicza sie pozajadrowo.
37. RODZAJE CYTOPLAZMATYCZNEGO DZIEDZICZENIA
1. Dziedziczenie mitochondrialne - w DNA mitochondrialnymzachodza roznego typu mutacje umozliwiajace badanie dziedziczenia mitochondrialnego. Kalsycznym obiektem tych badan sa drozdze u ktorych dzieki zdolnosci do calkowitego wylaczania oddychania i pobierana energii z fermentacji mozliwe jest badanie mutantow mitochondrialnych niezdolnych do oddychania, badania wykazaly ze czasteczki mitDNA wystepujace w heteroplazmonach moga miedzy soba rekombinowac co stwarza podstawy do mapowania genow w czasteczkach mitDNA. W innych organizmach niezdolnych do fermentacji wykryto analogiczne do drozdzowych mutanty odporne na rozne antybiotyki bakteryjne i wykazano ze dziedzicza sie one pozajadrowo i sa zapewne wynikiem mutacji w mitDNA. O mitDNA kregowcow i czlowieka wiemy jeszcze mniej.
2. Dziedziczenie chloroplasowe - chloroplasty wystepuja w licznych gatunkach glonow i tkankach asymilujacych roslin naczyniowych. Ilosc ich na komorke waha sie od kilkuset u glonow do kilku lub jednego. Wytwarzanie defektywnych chloroplastow moze byc zarowno wynikiem mutacji jadrowych dziedziczacych sie wg zasad mendlowskich lub powstaja one tez w wyniku mutacji w genach w DNA chloroplastowym i wtedy wylazuja typowe dziedziczenie pozajadrowe.
38. WŁASCIWOSCI GENOMOW - MITOCHONDRIALNEGO I CHLOROPLASOWEGO
1. Mitochondrialne - koduje 3 rodzaje rRNA , wieksze czasteczki tRNA 3 podjednostki oksydazy cytochromu i jednostke ATPazy.
2. Chloroplastowem - koliste koduja 3-4 Rrna
39. MECHANIZMY ODPOWIEDZALNE ZA JEDNORODZICIELSKIE DZIEDZICZENIE
1. wykluczanie organelli z komorek generatywnych lub jednej komorki, np.plemniek w czasie I i II mitozy traci niemal cala cytoplazme
2. delecja - wypadanie organelli lub calej cytoplazmy podczas rozwoju mikrospory
3. wykluczenie organelli podczas fuzji genetycznej ( zlanie plemnika z jajem )
4. degeneracja, brak replikacji lub kompartmentacji do komorki ( supresja po utworzeniu zygoty )
41. CYTOPLAZMATYCZNA MESKA STERYLNOSC I JEJ TYPY
Cecha bezplodosci pylku u wielu roslin zwiazana jest z cytoplazma. Jednym z lepiej poznanych jest przyklad przekazywania bezplodnosci meskiej przez cytoplazme mateczna u kukurydzy. Rosliny o bezplodnym pylku zawiazuja normalnie nasiona i nie wykazuja nawet sladow bezplodnosci zenskiej, pewnien nieduzy procent tych roslin wytwarza wyjatkowo pojedyncze pylniki zawierajace zywotny pylek. Sa to rosl.czesciowo bezplodne.
Osobniki wykazujace cytoplazmatyczna st.meska znane sa u wielu roslin, np. u kukurydzy, cebuli pszenicy.
Cecha sterylnosci meskiej jest wynikiem wspoldzialania cytoplazmy z genem recesywnym ulokowanym w jadrze kom.rosliny o normalnej cytoplazmie N wytw.normalny pylek, zas o nienormalnej S wykazuja sterylnosc meska, o ile zawieraja jeszcze do tego gen recesywny. Gen ten nie wykazuje zadnego dzialania w normalnej cytoplazmie. Z tego wynika ze wszystkie rosliny wykazujace meska sterylnosc u cebuli maja sklad genet. Smm. Gen dominujacy M wplywa na wystepowanie normalnej cytoplazmu niezaleznie czy jest w formie MM czy Mm.
Od czasu do czasu w niektorych odmianach cebuli pojawiaja sie osobniki o bezplodnym pylku. Sterylosc meska wykryta byla przez jonesa w 1925 roku na odmianie Red Italian. Jones znalazl mutanta gdzie w pylniku nie wytwarzal pytlku. Poz tym roslina byla normalna i nie roznila sie od formy matecznej. Analiza genetyczna wykazala ze meska sterylnosc pojawia sie tylko w przypadku spotkania cytoplazmy S i z obu recesywnymi genami mm.
W obecnosci normalnej cytoplazmy N powstaja zawsze kwiaty normalne niezaleznie czy wystepuje gen m czy M.
ODMIANY :
-genetyczna - nie wytwarza sie pylek
-funkcjonalna - organy meskie nie funkcjonuja jesli nie zostana spelnione pewne wymagania, np kwiat sie nie otworzy
43. PLAZMIDY - RODZAJE I CHARAKTERYSTYKA
Plazmidy to kwasy nukleinowe ktore maja zdolnosc do samoreplikacji (powielania sie)
- episomy - plazmidy ktore moga rekombinowac z DNA jadrowym i wlaczac sie do genomu (albo calkiem albo fragmentarycznie)
- plazmidy wystepuja w cytoplazmie organizmow prokariotycznych
- ich liczba to zazwyczaj 1-2 plazmidu tego samego rodzaju
- w komorkach moze wystepowac kilka rodzajow plazmidow jesli sa one w stosunku do siebie „przyjzane” (jesli sa jadrowe). Moga wystepowac w jednej kom.razem. Jesli stosunek plazmidow do siebie jest negatywny nie moga one istniec w jednej kom.jednoczesnie
- plazmidy namnazaja sie w komorce niezaleznie od jej podzialow
- plazmidy dzielimy na :
1. naturalne
2. sztuczne (powstaja w wyniku zmiany plazmonu - p.dimeralne)
plazmidy niosa ze soba b.wiele roznych wlasciwosci
- wytwarzaja antybiotyki
R - odporny na antybiotyki Ti - agrobacterium tumefaciens
F - przenoszenie
44. AGROBACTERIUM TUMEFACIENS I JEGO STRATEGIA ZYCIOWA
Jest to bakteria atakujaca rosliny - wprowadza swoje geny ( zawarte z plazmidzie Ti) do komorki roslinnej.
Agrobacterium
· jest glebową bakterią patogenną - powoduje choroby roślin: guzowatość korzeni (A.tumefaciens) lub przerosty korzeni (A.rhizogenes)
· Rodzina: Rhizobiaceae, Rodzaje: Rhizobium i Agrobacterium
· Gatunki z rodzaju Agrobacterium: m.in. A.tumefaciens i A.rhizogenes
· Gram (-)
· zawierają duże plazmidy (megaplazmidy): Ti (A.tumefaciens) i Ri (A.rhizogenes)
· potrafi przenosić fragment tego plazmidu (tzw. T-DNA) do genomu komórek roślinnych
· przenoszony fragment niesie funkcje determinujące syntezę opin - związki potrzebne do wzrostu bakterii (źródło węgla i azotu)
· każdy ze szczepów może utylizować jedną z trzech klas opin:
o oktopinę i jej pochodne (lizopina, kwas oktopinowy, histopina)
o nopalinę i jej pochodne (kwas nopalinowy, agrocypina)
o agropinę
· infekuje rośliny z 331 rodzajów, 643 gatunków
· infekcyjność zależy od gatunku i organu rośliny
· gatunki jednoliścienne są "mniej" wrażliwe na infekcję Agrobacterium
TRANSFORMACJA Agrobacterium
Bakteria glebowa Agrobacterium tumefaciens powoduje powstawanie guzów we wszystkich badanych pod tym względem roślinach dwuliściennych i w niektórych jednoliściennych. Za tworzenie się guzów odpowiedzialny jest plazmid Ti (ang. tumor inducing). Fragment tego plazmidu przenoszony jest do komórki roślinnej i stabilnie integruje w genom. Zawarte w tym fragmencie geny ulegają ekspresji w komórce roślinnej. Integrujący fragment nazwano T-DNA (transferred DNA). Analiza genetyczna wykazała, że dwa regiony plazmidu Ti są niezbędne do powstania guza: T-DNA i region vir (virulence). Mutacje we fragmencie T-DNA zmieniają morfologię guza, natomiast mutacje w regionie vir zaburzają jego powstanie. T-DNA zawiera geny które są związane z produkcją fitohormonów (auksyn, cytokinin, kwasu indolilooctowego). Nadprodukcja tych hormonów podwyższa tempo podziału komórki, zapobiega różnicowaniu i w rezultacie powoduje powstanie guza. Inne geny T-DNA odpowiedzialne są za syntezę aminokwasów i pochodnych cukrowych zwanych opinami, oraz za ich wydzielanie z komórki roślinnej. Opiny zawarte w guzie tkanki roślinnej mogą służyć jako Yródło węgla i azotu dla Agrobacterium.
Wykazano, że obszar T-DNA zawiera sekwencje niezbędne do przeniesienia go do komórki roślinnej. Niezbędny do przeniesienia T-DNA jest region vir, który jest aktywny nawet gdy znajduje się w innym replikonie. Wiadomo, że powtórzenia sekwencji o długości 25 bp tworzące granice T-DNA są niezbędne, ale i wystarczające aby region ten stabilnie integrował w genom roślinny. DNA zawarty między granicznymi sekwencjami przenoszony jest do komórki roślinnej z pomocą białek kodowanych przez geny znajdujące się w regionie vir, a następnie integruje w genom roślinny. Odkrycie mechanizmu tego zjawiska umożliwiło skonstruowanie wektorów używanych obecnie do transformacji komórek roślinnych z wykorzystaniem Agrobacterium.
Proces manipulowania plazmidem wprowadzanym do rośliny, można podzielić na dwa etapy :
1. Klonowani i zmiany w DNA prowadzone w bakteriach E.coli
2. Wprowadzenie gotowego konstruktu do Agrobacterium, które służy do transformacji rośliny.
45. CHARAKTERYSTYKA PLAZMIDU Ti
Charakterystyka plazmidu Ti
· duży kolisty plazmid zawierający min. geny:
- wirulencji
- katabolizmu specyficznych dla szczepu opin
- biosyntezy opin specyficznych dla szczepu
- związane z biosyntezą hormonów roślinnych - specyficzne dla szczepu
· geny wirulencji i katabolizmu opin są zgrupowane razem i znajdują się poza obszarem T-DNA
· geny biosyntezy opin oraz geny związane z syntezą hormonów roślinnych znajdują się w obszarze T-DNA
· obszar T-DNA jest wyznaczony przez specyficzne sekwencje DNA (bezpośrednie powtórzenia o długości 25 pz) zwane granicznymi (LB- Left Border i RB - Right Border)
· tylko prawa sekwencja graniczna (RB) jest wymagana do rozpoznania i przeniesienia T-DNA
· obecność lewej sekwencji granicznej (LB) poprawia efektywność tego procesu
· wielkość plazmidów Ti: 150-800 kb
Typy plazmidów Ti (dzikich)
Istnieją różne typy plazmidów Ti zależnie od tego jaki rodzaj opin potrafią utylizować. Najlepiej poznane są plazmidy Ti nopalinowe i oktopinowe. Obydwa typy posiadają obszar T-DNA ale różniący się strukturą:
· Nopalinowy T-DNA - prosty ciągły segment, około 22 kb
· Oktopinowy T-DNA - złożony z 3 segmentów; lewy segment DNA (TL) = 13kb, centralny segment (TC) = 1.5kb, prawy segment (TR) = 7.8kb; lewy segment zawiera funkcje onkogenne a prawy segment geny syntezy opin
Uogólniony model przeniesienia i integracji T-DNA
· Nić T powstaje i jest przenoszona do komórki roślinnej w postaci kompleksu DNA-białka. Transfer wymaga genów vir zlokalizowanych na plazmidzie Ti i genów chromosomalnych
· Białko, które znajduje się na końcu 5' nici T-DNA wchodzi w interakcję z roślinnym DNA i nacina je
· Jednoniciowe T-DNA jest przyłączane do jednej nici roślinnego DNA a napięcia torsyjne z tym związane powodują rozerwanie drugiej nici roślinnego DNA
· Końce nici T są łączone z końcami roślinnego DNA na drodze ligacji, dosyntetyzowana zostaje nić komplementarna do nici T
· Naprawy (łatanie dziur) i replikacja obszarów integracji nici T prowadzą często do duplikacji i rearanżacji w tym obszarze
46. GENY ODCINKA T-DNA I ICH EFEKTY FENOTYPOWE
Koduje 2 klasy genow warunkujacych transformowany fenotyp :
1. zaangazowanych w synteze opin
2. odpowiedzialnych za biosynteze regulatorow wzrostu roslin
T-DNA
· obszar przenoszony do komórek roślinnych
· zawiera geny kodujące białka zaangażowane w biosyntezę opin i białka zaangażowane w biosyntezę fitohormonów
· pomimo tego, że geny te rezydują na plazmidzie bakteryjnym to są aktywne tylko w komórkach roślinnych
· zawierają typowe eukariotyczne sekwencje regulujące ich transkrypcję: TATA-boxy i CAAT-boxy oraz typowe roślinne sygnały poliadenylacji
· część tych sekwencji wykorzystano do regulacji ekspresji genów markerowych
47. OPINY JAKO MARKERY GENETYCZNE
Opiny tworzone w
tumorowatej tkance moga byc katabolizowane
przez Agrobacterium, ale nie przez inne mikro-
organizmy glebowe, i stanowic jako substancje
odzywcze węgla i azotu. W ten sposób
Agrobacterium tworzy dla siebie nisze ekolo-
giczne poprzez genetyczna modyfikacje komó-
rek roslinnych, proces nazywany równiez genetyczna kolonizacja.
Poniewaz w wektorach ( poch. Plazmidow Ti) usuwa sie geny nadajace po infekcji komorkom roslinnym fenotyp nowotworowy traci sie system slelkcyjny ich niezaleznosc od hormonalnych regulatorow wzrostu. Do selekcji wykorzystuje sie dominujace geny markerowe - fuzja promotora jednej z syntaz opin z genem odpormnosci, odporonosci na antybiotyk.DUPA
48. TRANSFORMACJA DNA JAKO ZJAWISKO OGOLNOBIOLOGICZNE
Transformacja (definicja mikrobiologiczna wg. Griffitha)
zmiana fenotypu organizmu przez wprowadzenie obcego DNA do jego genomu
Transgeniczne rośliny są otrzymywane przez wprowadzenie obcego DNA do tkanki roślinnej i zregenerowanie roślin zawierających obce DNA
Pierwsze próby transformacji roślin - moczenie nasion w DNA, ale to było dawno
Metody transformacji roślin:
- pośrednie (za pomocą „pośrednika”)
· Agrobacterium tumefaciens
· Agrobacterium rhizogenes
- bezpośrednie (wprowadzanie „nagiego” DNA)
· elektroporacja
· PEG
· mikrowstrzeliwanie
· mikroiniekcja
Etapy procesu transformacji
Ţ przyczepienie się Agrobacterium do komórek roślinnych - do 200 bakterii może się przyczepić do 1 komórki roślinnej
Ţ przeniesienie DNA do komórki roślinnej - kilka obszarów T-DNA może być przeniesione do 1 komórki roślinnej
Wyszukiwarka
Podobne podstrony:
opracowania do egz
Politologia religii opracowania do egz Politologia Religii zagadnienia
Politologia religii - opracowania do egz, Bierdiajew, Koszmar złego dobra - Nikołaja Bierdiajewa int
Politologia religii - opracowania do egz, ST PK, 3
Politologia religii - opracowania do egz, RELIGIA I NACJONALIZM wg mnie
Politologia religii opracowania do egz wykłady łętocha
genetyka egz, materiały, genetyka
Prpgram Razem bezpieczniej, Bezpieczeństwo narodowe - UAM Poznań, I rok (2012-2013), Teoria Bezpiecz
OPRACOWANE zagadnienia do EGZ
Opracowane zagadnienia do egz z ped. ogólnej, Pedagogika ogólna
materiały do egz (orto, pulmo, itp), Mukowiscydoza, Mukowiscydoza
Egzamin Opracowane Pytania Wstep do nauki o materialach
TEST POWT do egz rach finansowa czerwiec 2012, PB-materiały, semestr IV, rachunkowość finansowa
więcej podobnych podstron