3. Mikrofilamenty (filamenty aktynowe)
Osiągają grubość około 7 nm
Zbudowane z aktyny, które morze występować w dwóch postaciach (monomer lub polimer); aktyna stanowi 15-20% całości białek każdej komórki.
Aktyna G (monomer) jest białkiem globularnym występującym w cytoplazmie; jest to pojedynczy łańcuch polipeptydowy; białko idealnie sferyczne, poznano sekwencję 375 aminokwasów. Rozróżniamy cztery izomeryczne formy G-aktyny:
α - występuje w mięśniach poprzecznie prążkowanych
β i γ - występują w mięśniach gładkich i komórkach niemięśniowych
δ - występuje tylko u Acanthoamoeba
Aktyna F (polimer) jest białkiem włóknistym; włókno aktynowe widoczne w komórkach to dwa śrubowato skręcone łańcuchy F-aktyny. Rozróżniamy:
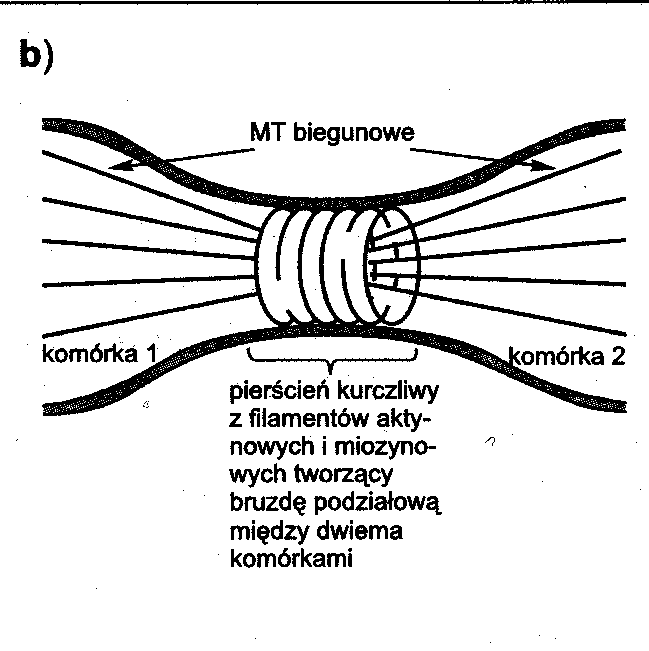
F-aktynę labilną, która ulega depolimeryzacji w temperaturze 4°C, jest bogata w gelsolinę, odpowiada za kształt komórki, buduje lameliopodia i filopodia, buduje włókna naprężeniowe, pozwala na ruch bakterii w komórce gospodarza oraz odpowiada za podział cytoplazmy (tworzy pierścień kurczliwy w cytokinezie).
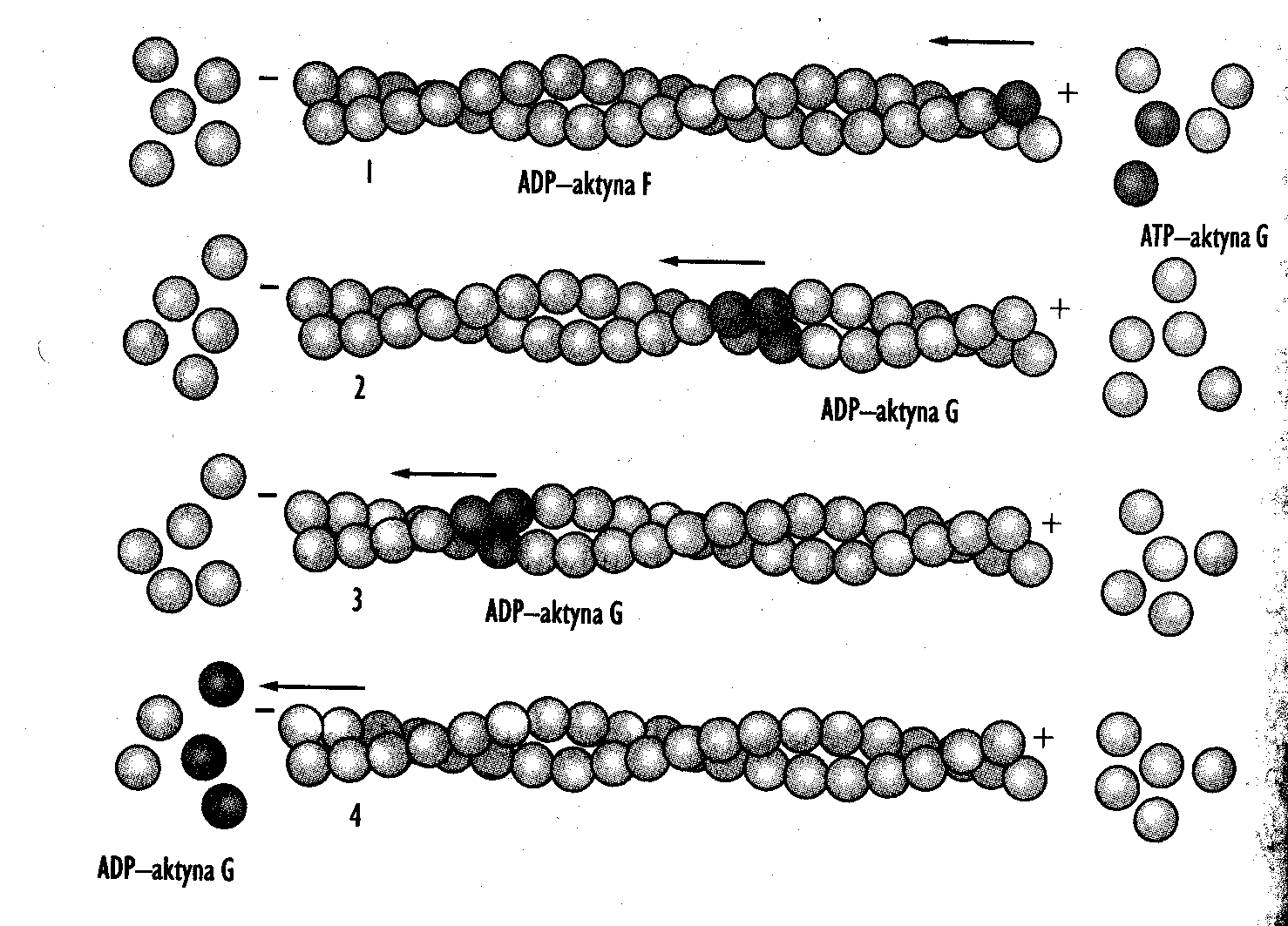
F-aktynę stabilną, która buduje stałe struktury (np. mikrokosmki), jest odporna na depolimeryzację w temperaturze 4°C, uboga w gelsolinę, buduje aparat kurczliwy komórek mięśniowych.
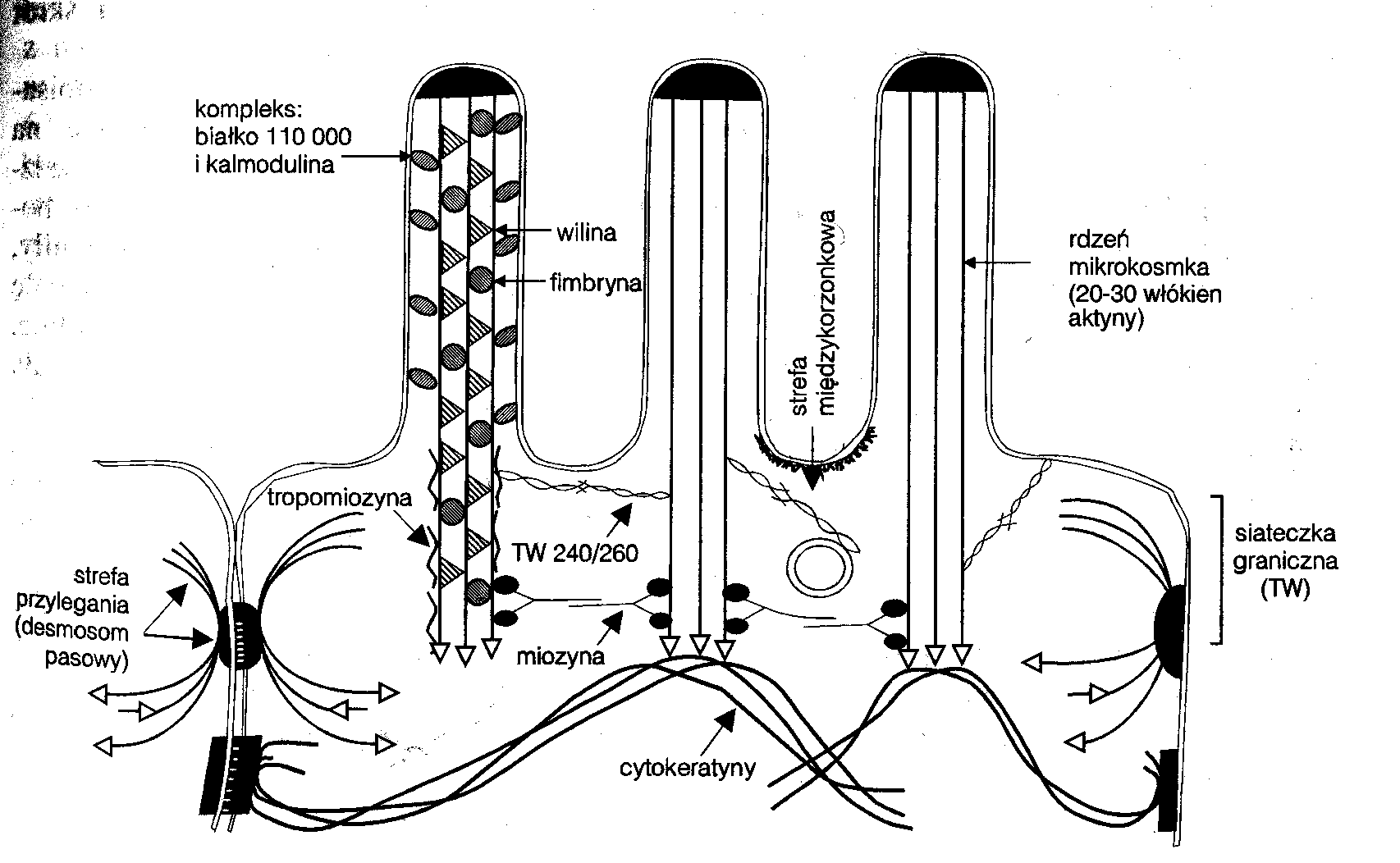
Mikrokosmki
Na powierzchni komórek nabłonka jelitowego występuje od 1300 do 1500 mikrokosmków, które ogromnie powiększają powierzchnię chłonną komórki.
Mikrokosmki są palczastymi wypustkami cytoplazmatycznymi o średnicy 50 - 100 nm.
Rdzeń mikrokosmka stanowi wiązka 20 - 30 filamentów aktynowych zachowująca stabilność dzięki wilinie i fimbrynie ulokowanej pomiędzy filamentami.
Podstawę mikrokosmka mogą budować również inne białka.
Ruch pełzakowaty
Tego typu sposób poruszania się występuje min. u ameb oraz białych krwinek (granulocyty obojętnochłonne, które migrują z krwi do tkanek).
Za ruch pełzakowaty odpowiadają wypustki powstające poprzez wypchnięcie powierzchni komórki w danym kierunku. Jest to możliwe dzięki polimeryzacji aktyny (mikrofilamenty są spolaryzowane; na biegunie + proces polimeryzacji biegnie szybciej, więc mikrofialment ulega wydłużeniu).
Zasadniczo rozróżniamy dwa typy wypustek: cienkie i ostre filopodia oraz płytkowate lameliopodia.
ABP (Actin Binding Proteins) - białka towarzyszące aktynie
Jest to grupa białek mających powinowactwo do mikrofilamentów.
Dzięki tym białkom mikrofilamenty mogą być tworzyć różne układy przestrzenne:
Gelsolina - powoduje depolimeryzację mikrofilamentów (cięcie na kawałki).
Tropomiozyna - stabilizuje strukturę mikrofilamentów.
Miozyna - nadaje własność tworzenia sieci.
Filamina - powoduje formowanie sieci.
Fimbryna - powoduje utrzymywanie mikrofilamentów w pęczkach.
Winkulina - odpowiada za powiązanie mikrofilamentów z mikrotubulami i błoną komórkową.
α-aktynina - powoduje powstawanie luźnych wiązek.
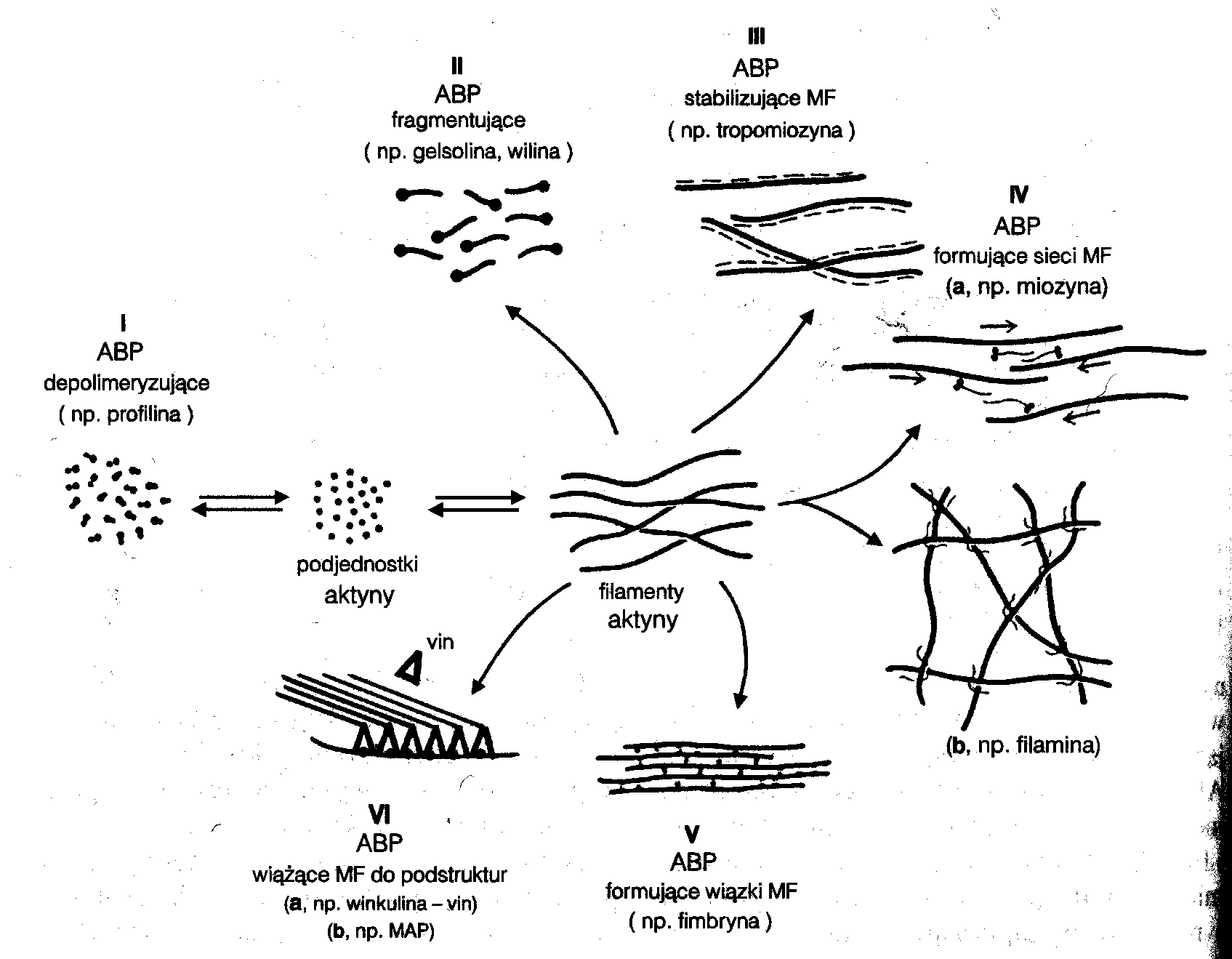
Spektryna - formowanie sieci i wiązanie mikrofilamentów z błoną komórkową.
Trucizny dla mikrofilamentów:
Cytochalazyna - blokuje polimeryzację filamentów aktynowych.
Falloidyna - uniemożliwia depolimeryzację, a w połączeniu z rodaminą wykorzystywana jest do barwienia mikrofilamentów (na czerwono mikroskop fluorescencyjny).
W cytoplazmie występują czynniki warunkujące występowanie obok siebie dwóch rodzajów aktyny (F i G); zapobiega to niepotrzebnej polimeryzacji i depolimeryzacji; tymi związkami są profilina i tymozyna.
III Cykl komórkowy
Jest to czas od momentu zakończenia jednego podziału do momentu zakończenia podziału następnego.
Na cykl komórkowy składają się zasadniczo dwie fazy:
Interfaza (którą dzielimy na fazę G1, S i G2 oraz czasem G0)
faza podziału.
Często fazy G1 i G2 są zredukowane (np. podczas rozwoju zarodkowego).
Faza spoczynkowa G0 może nastąpić po fazie G1 lub po fazie G2 (dochodzi do podwojenia ilości materiału genetycznego). Długość trwania fazy G0 jest zróżnicowana; może trwać wiele lat; często komórka po ostatecznym zróżnicowaniu przechodzi w fazę G0 i pozostaje w niej aż do śmierci (ponowne wejście w fazę S może być oznaką zmian nowotworowych). W czasie fazy spoczynkowej komórka nie wykazuje zdolności do syntezy DNA i podziału.
Faza G1 to okres czasu od zakończenia podziału do rozpoczęcia syntezy DNA. Czas trwania jest zmienny (przeciętnie kilka - kilkanaście godzin). W fazie tej obserwowany jest wzrost aktywności metabolicznej, zwiększenie masy i objętości oraz wzrost ilości białek i RNA.
Punkt restrykcyjny R to punkt, po osiągnięciu którego komórka na pewno wejdzie w fazę S (jeśli go nie osiągnie, przejdzie w fazę G0).
Faza S charakteryzuje się zróżnicowanym czasem trwania (u człowieka 6 - 8 godzin potrzeba na replikację 1,8 metra nici DNA). Następuje tu podwojenie ilości informacji genetycznej zawartej w DNA.
Faza G2 to faza bezpośrednio poprzedzająca podział komórki. Zachodzi tu wzmożona synteza białek wrzeciona podziałowego (głownie tubuliny), składników do odbudowy otoczki jądrowej oraz błony komórkowej. W fazie tej dochodzi również do semokonserwatywnego podziału centrioli.
Wrzeciono podziałowe
Zbudowane jest z mikrotubul, białek towarzyszących mikrotubulom (dyneina, miozyna, aktyna) oraz zbiorników i pęcherzyków siateczki endoplazmatycznej gładkiej (SER).
W ramach wrzeciona wyróżniamy:
Mikrotubule astralne, które rozpościerają się we wszystkie strony, jednak nie kontaktują się z chromosomami ani z mikrotubulami przeciwnego bieguna.
Mikrotubule kinetochorowe, które kontaktują się z chromosomem w miejscu zwanym kinetochorem; są zaangażowane w rozchodzenie się połówek chromosomów w anafazie.
Mikrotubule biegunowe, które biegną od bieguna do bieguna, zachodzą na siebie; nie łączą się z chromosomami; odpowiedzialne są za oddalanie się połówek chromosomów w anafazie.
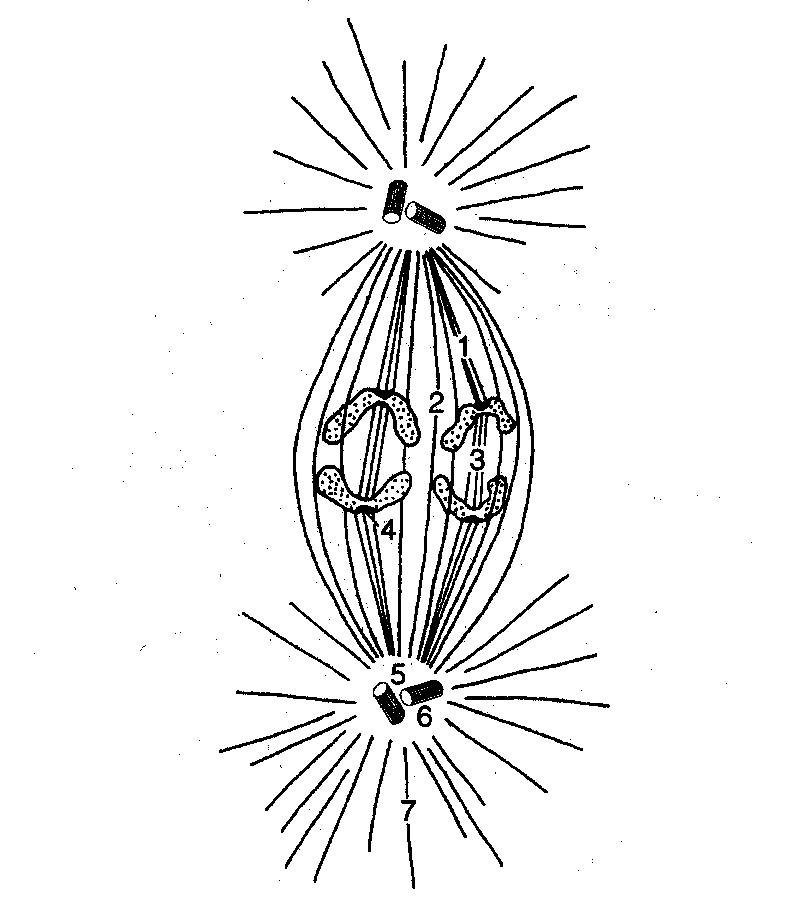
Kompleks synaptomenalny (synaptomalny) to morfologiczny wyznacznik profazy mejozy; odpowiedzialny jest za łączenie się chromosomów homologicznych w biwalenty w czasie zygotenu. Kompleks ten zanika w momencie rozczepienia się chromosomów w diplotenie.
Wykład # 4
1
Wyszukiwarka
Podobne podstrony:
biologia komorki zwierzecej apoptoza
Fizjologia zwierząt wszystkie opracowania, chemia organiczna, biologia ewolucyjna-wykłady, genetyka,
budowa komorki i neuronu, Biology, III rok, Fizjologia zwierząt, Dodatkowe materiały
KOMÓRKI ZWIERZĘCE I ROŚLINNE, Edukacja xD, Biologia, Referaty
Biologia komórki zwierzęcej #3, Biologia komórki zwierzęcej
Budowa komórki zwierzęcej, matura, biologia r
Cytokineza zwierząt i roślin, Biologia komórki
Biologia komórki zwierzęcej
KOMORKA ZWIERZECA, Studia, Biologia
Różnice w budowie między komórką roślinną a zwierzęcą, BIOLOGIA MOJE PRYWATNE
BIOLOGIA, komorki rosline i zwierzece
więcej podobnych podstron