1.Znaczenie kory mózgowej w czynnościach życiowych.
Kora mózgu odbiera i analizuje informacje z narządów zmysłów. Odbywają się w niej także procesy skojarzenia, stąd też wysyłane instrukcje określające reakcje ruchowe. Odpowiada za czucie somatyczne, widzenie, słyszenie, czucie, uczenie się oraz planowanie i polecenie ruchów. W przedniej części płata ciemieniowego znajdują się ośrodki czuciowe (tzw. kora czuciowa lub kora somatoczuciowa) i dochodzą tu impulsy z receptorów. W tylnej części płata czołowego znajdują się ośrodki ruchowe (tzw. kora ruchowa lub kora somatoruchowa). Odpowiada za bardzo precyzyjne ruchy. Prawa półkula mózgu kontroluje lewą stronę ciała, natomiast lewa półkula prawą stronę ciała. W płatach potylicznych zlokalizowane są ośrodki wzrokowe, które interpretują obrazy. W płatach skroniowych umieszczone są ośrodki słuchowe.
2.Dojrzałość płciowa.
Zwierzęta gospodarskie osiągają dojrzałość płciową, kiedy są zdolne do wytwarzania i uwalniania pełnowartościowych gamet. Samce są zdolne do ejakulacji nasienia, a u samic występuje pierwsza w życiu ruja i jajeczkowanie. Samice wszystkich gatunków zwierząt osiągają dojrzałość płciową wcześniej niż samce. Wiek dojrzałości płciowej jest uwarunkowany genetycznie. Wiele czynników jednak może ją opóźniać lub przyspieszać, np. warunki środowiskowe ( nieprawidłowe żywienie), klimatyczne, temperatura. Samice mające okresowy kontakt z samcami jak np. loszki, osiągają dojrzałość płciową wcześniej niż wychowywane w odosobnieniu. Jeśli ruja występuje sezonowo jak np. u owiec, osiągnięcie dojrzałości płciowej zależy od terminu porodu. Maciorki urodzone w styczniu osiągają dojrzałość płciową później ( w wieku ok. 8 miesięcy ) niż maciorki tej samej rasy urodzone w kwietniu ( w wieku 6 m-cy ). Neurohormonalna regulacja osiągnięcia dojrzałości płciowej jest procesem złożonym związanym ze zmianami na wszystkich poziomach osi regulacyjnej: podwzgórze-przysadka-gonady, jak również w wyższych partiach ośrodkowego ukł.nerwowego. U samic i samców brak wyraźnych różnic w poziomach LH, FSH, PRL podczas dojrzewania płciowego.
Osiągnięcie dojrzałości płciowej przez zwierzęta nie jest równoznaczne z osiągnięciem dojrzałości hodowlanej. Ta ostatnia występuje później i jest związana z odpowiednim rozwojem całego organizmu.
3.Mechanizm działania hormonów.
Ogólnie działanie hormonów polega na aktywacji lub dezaktywacji pewnych mechanizmów komórkowych w tkankach docelowych (narządach docelowych). Na przykład insulina tak wpływa na komórki, że aktywuje mechanizmy pobierania glukozy, co powoduje spadek stężenia glukozy we krwi. Aktywacja lub dezaktywacja odbywa się przez łączenie ze specyficznymi błonowymi lub wewnątrzkomórkowymi receptorami.
Wiele hormonów ma działanie wzajemnie antagonistyczne - np. insulina i glukagon. Insulina powoduje spadek stężenia glukozy we krwi, a glukagon wzrost jej stężenia.
Hormony tropowe - Wśród hormonów można wyróżnić grupę, której zadaniem jest regulacja czynności innych hormonów. Na przykład hormon tyreotropowy (TSH) wydzielany przez przedni płat przysadki mózgowej wpływa na zwiększenie wydzielania hormonów tarczycowych - tyroksyny , a także trójjodotyroniny. Poza tym TSH wpływa na zwiększenie ukrwienia gruczołu tarczowego, a także taką przebudowę strukturalną pęcherzyków tarczycowych, która pozwala sprostać wymogom zwiększonej czynności hormonalnej. Tę grupę nadrzędnych "nadzorców" nad innymi gruczołami dokrewnymi nazywamy hormonami tropowymi.
Nad hormonami tropowymi kontrolę sprawuje wyższe piętro nadzoru. Znajduje się ono w części mózgu nazywanej podwzgórzem. Podwzgórze produkuje hormony uwalniające i hamujące, które wpływają na wzrost lub spadek wydzielania hormonów tropowych produkowanych przez przysadkę. I tak, istnieje tyreoliberyna powodująca uwalnianie (zwiększenie wydzielania) hormonu tropowego - TSH (hormon tyreotropowy), a także somatostatyna, która zmniejsza wydzielanie hormonu wzrostu przez komórki przysadki mózgowej.
Mechanizm uwalniania - Ogólnym mechanizmem działającym w obrębie układu hormonalnego jest ujemne sprzężenie zwrotne. Produkt wydzielany przez dany gruczoł dokrewny np. tarczycę - czyli tyroksyna (T4) (a także bezpośrednio trójjodotyronina), wpływa hamująco na gruczoł dokrewny nadzorczy czyli przysadkę mózgową. Powoduje to spadek wydzielania TSH przez przysadkę i z kolei hormonów tarczycy. Spadek nie może przekroczyć pewnej określonej granicy, gdyż wówczas ujemny wpływ maleje, co pozwala na ponowne produkowanie większych ilości TSH. Jest to element homeostazy i system ten działając we wzajemnym sprzężeniu, utrzymuje równowagę hormonalną organizmu.
4.Proces krzepnięcia krwi.
Krzepnięcie krwi - naturalny, fizjologiczny proces zapobiegający utracie krwi w wyniku uszkodzeń naczyń krwionośnych. Istotą krzepnięcia krwi jest przejście rozpuszczonego w osoczu fibrynogenu w sieć przestrzenną skrzepu (fibryny) pod wpływem trombiny. Krzepnięcie krwi jest jednym z mechanizmów obronnych organizmu w wypadku przerwania ciągłości tkanek.
Kaskada krzepnięcia krwi
W warunkach fizjologicznych proces krzepnięcia krwi zostaje zapoczątkowany przez przerwanie ciągłości łożyska naczyniowego i ma za zadanie zapobiec wynaczynieniu i utracie krwi. W miejscu przerwania ciągłości naczynia dochodzi do odsłonięcia macierzy podśródbłonkowej na której płytki krwi (trombocyty) natychmiast przylegają, a następnie na skutek zlepiania się trombocytów ze sobą tworzą tzw. czop płytkowy. Płytki krwi ulegają aktywacji i uwalniają szereg substancji czynnych z ziarnistości α i β, które dodatkowo nasilają ich aktywację. Dochodzi do zmian morfologicznych płytek, a także ekspresji wielu białkowych cząsteczek na ich powierzchni. Uwolniona serotonina powoduje zwężenie naczyń krwionośnych w obrębie zranienia. Chociaż czop płytkowy czasowo doprowadza do zahamowania krwawienia, właściwy skrzep powstaje poprzez utworzenie sieci włóknika, która powoduje jego wzmocnienie i stabilizację. Dochodzi do tego przez aktywację kaskady krzepnięcia - krążące w osoczu krwi nieaktywne czynniki krzepnięcia zaczynają się wzajemnie aktywować. Płytki pod wpływem uszkodzeń mechanicznych wydzielają trombokinazę, która uruchamia szereg procesów prowadzących do powstania właściwego czynnika inicjującego krzepnięcie krwi - w procesie tym ważne są jony wapnia oraz białkowe czynniki osocza. W rezultacie kaskady krzepnięcia aktywna forma czynnika X, przy współdziałaniu nieenzymatycznego kofaktora - czynnika Va oraz fosfolipidów powierzchniowych tworzy kompleks (protrombinaza), który w sposób proteolityczny przekształca protrombinę do trombiny. Trombina z kolei powoduje przekształcenie fibrynogenu (osoczowego białka krążącego we krwi) w fibrynę (białko nierozpuszczalne w wodzie), która tworzy sieć włókien, będących szkieletem skrzepu.
Dla prawidłowego zachodzenia procesu krzepnięcia krwi niezbędna jest witamina K, która umożliwia syntezę protrombiny w wątrobie.
5.Zjawisko snu i czuwania.
Kontrolę nad stanami czuwania i snu ma układ siatkowaty pnia mózgu.
Czuwanie opiera się na procesie fizjologicznym, polegającym na aktywacji ośrodków korowych, dzięki którym organizm odbiera bodźce ze środowiska zewnętrznego, reaguje na nie i utrzymuje pełny kontakt z otoczeniem. Czuwanie jest wynikiem wzmożonej aktywności aktywującego układu siatkowatego śródmózgowia. Mechanizmy czuwania pobudzane są przez impulsy czuciowe dopływające z obwodu do ośrodkowego UN. Wyniki badań wykazały, że w przedniej części mostu jest ośrodek nerwowy, który - mimo braku impulsacji z obwodu ciała - wysyła toniczną impulsację do układu siatkowatego utrzymując stan czuwania.
- CZUWANIE: aktywacja układu siatkowatego zachodzi przez impulsy dośrodkowe wzmagające aktywność kory mózgowej , a ta na zasadzie sprzężenia dodatniego aktywuje ukł. siatkowaty. Wzmożona aktywność ukł. siatkowatego wpływa na ośrodki obwodowe, zwiększając np. napięcie mięśniowe, co na zasadzie sprzężenia zwrotnego dodatniego aktywuje ukł. siatkowaty. Aktywacja ukł. siatkowatego powoduje wzrost aktywności ukł. współczulnego i wydzielanie przez niego noradrenaliny, która aktywuje ukł. siatkowaty.
Sen stanowi przejściową utratę łączności czuciowej z otoczeniem, która może być natychmiast przywrócona przez działanie dostatecznie silnych bodźców zewnętrznych.
Podczas snu zachodzi:
zmniejszenie pobudzenia w wielu ośrodkach UN - spadek ich pobudliwości
ustanie aktywności ruchowej zwierzęcia - obniżenie napięcia mięśniowego
przewaga części przywspółczulnej ukł. autonomicznego nad kontrolą organizmu
zwalnia się częstość skurczów serca
ciśnienie krwi się obniża
oddechy stają się wolniejsze i bardziej regularne
temperatura ciała i produkcja moczu spadają
źrenice się zwężają
ruchy żwacza ulegają znacznemu zwolnieniu
W przebiegu snu wyróżniamy dwa różne poziomy:
Poziom snu powierzchownego (snu powolnego), któremu towarzyszy powolna elektryczna aktywność kory mózgowej.
Poziom snu głębokiego (snu szybkiego = paradoksalnego), któremu towarzyszy przyspieszona aktywność elektryczna kory mózgowej podobna jak w czasie czuwania, a różni się od tego stanu występowaniem:
szybkich, skojarzonych ruchów gałek ocznych
bardzo dużym spadkiem, a nawet zanikiem napięcia w mięśniach szyjnych podtrzymujących głowę
Występują wtedy marzenia senne.
Zasadnicze mechanizmy snu znajdują się w obrębie siatkowatego układu pnia mózgu i strukturach podkorowych.
Miejscem powstawania snu mogą być jądra szwu wytwarzające serotoninę i miejsce sinawe (układ hamujący mostu) wytwarzający noradrenalinę. Obie aminy biogenne mają na przemian działać na układ siatkowaty doprowadzając do snu. Jądra szwu mogą być odpowiedzialne za sen powierzchowny, a miejsce sinawe za sen głęboki.
Według innych poglądów czuwanie jest oparte na sprzężeniu zwrotnym dodatnim, a sen na sprzężeniu zwrotnym ujemnym.
Po dłuższym czuwaniu następuje zmęczenie i stopniowy spadek aktywności neuronów poszczególnych pętli sprzężenia zwrotnego dodatniego, a szczególnie układu siatkowatego a przez to jego aktywującego wpływu na korę mózgową. Sen nie jest aktem biernym ośrodkowego UN, ale wypadkową aktywności układu siatkowatego i hamującego wpływu mostu, którego przewaga nad aktywującym ukłądem śródmózgowia decyduje o zapadaniu w sen.
U zwierząt domowych sen występuje wielokrotnie w ciągu doby.
Eq i Ru - sen powtarza się wielokrotnie w ciągu doby, w sumie ok. 6h;
U Ru występuje 85% snu powolnego i 15% snu paradoksalnego
U Fe występuje 77% snu powolnego i 23% snu paradoksalnego
Częściowym niepełnym snem, wywoływanym w określonych warunkach u ludzi, jest sen hipnotyczny (hipnoza). Sprowadzić go można przez koncentrację uwagi osoby usypianej na określonych bodźcach działających na narządy wzroku i słuchu, receptory skóry itp. oraz na treści monotonnych słów wypowiadanych przez hipnotyzera. Stanu hipnotycznego, identycznego z hipnozą występującą u ludzi, nie można wywołać u zwierząt. Można natomiast spowodować u nich odruchową reakcję bezruchu (odruchowy bezruch - IR), niezupełnie słusznie nazywaną hipnozą zwierzęcą. Można ją wywołać zwróceniem całej uwagi wzrokowej zwierzęcia na jakiś szczegół w otoczeniu lub szybkim, energicznym, pokonującym wszelki opór ułożeniem, zwierzęcia na grzbiecie lub boku, przytrzymaniem go przez 20-30s i następnym ostrożnym usunięciem rąk.
Bodźce wywołujące opisany odruch aktywują szereg wstawkowych neuronach zstępującego hamującego układu siatkowatego znajdującego się w moście i rdzeniu przedłużonym. Wysyłane przezeń impulsy zstępujące wywołują powstanie postsynaptycznych potencjałów hamujących na motoneuronach rdzenia kręgowego i przez to głębokie, ogólne hamowanie mięśni szkieletowych. Kora mózgowa hamuje natomiast układ limbiczny i wzmaga aktywność kontrolnego ośrodka odruchowego bezruchu. Bezruch różni się od snu naturalnego, gdyż jest wynikiem określonych manipulacji i coraz rzadziej daje się wywołać u gatunków z lepiej rozwiniętą korą mózgową. Brak przy nim szybkich ruchów gałek ocznych oraz spadku napięcia w mięśniach szyjnych.
1.Potencjały spoczynkowe i czynnościowe.
Potencjał spoczynkowy to różnica potencjałów (napięcie) między obiema stronami błony plazmatycznej niepobudzonej komórki pobudliwej.
Wszystkie napięcia na błonie wyraża się jako różnice potencjału wnętrza komórki do potencjału po stronie zewnętrznej. Potencjały spoczynkowe mają wartości ujemne, które w komórkach nerwowych wahają się między -65 mV a -90 mV. Powstanie potencjału spoczynkowego jest spowodowane przede wszystkim tendencją jonów potasu do przepływania zgodnie z gradientem stężenia tych jonów z wnętrza na zewnątrz błony komórkowej. Powoduje to pozostanie niewielkiego nadmiaru ładunków ujemnych po wewnętrznej stronie błony. Inne jony (np. sodu) jedynie w niewielkim stopniu wpływają na wartości potencjału spoczynkowego. Siła elektrochemiczna, powodująca ruch jonu poprzez błonę komórkową, jest różnicą między potencjałem spoczynkowym a potencjałem równowagi dla danego jonu. Potencjał równowagi jonu jest to taki potencjał, przy którym wpływ tego rodzaju jonów z komórki jest równy ich wpływowi do jej wnętrza.
Potencjał równowagi dla poszczególnych jonów można obliczyć stosując równanie Nernsta. Potencjały spoczynkowe można obliczyć stosując równanie Goldmana, które uwzględnia wszystkie zaangażowanie rodzaje jonów.
Potencjał czynnościowy przejściowa zmiana potencjału błonowego komórki, związana z przekazywaniem informacji. Bodźcem do powstania potencjału czynnościowego jest zmiana potencjału elektrycznego w środowisku zewnętrznym komórki. Wędrujący potencjał czynnościowy nazywany jest impulsem nerwowym. Potencjał czynnościowy trwa od 1 ms do kilku sekund i osiąga maksymalnie wartości około +40 mV. Hiperpolaryzacja następcza trwa kilka milisekund.
Potencjały czynnościowe powstają na wzgórku aksonowym neuronu i rozprzestrzeniają się po błonie aksonu. Zachowują się one zgodnie z zasadą "wszystko albo nic": do zapoczątkowania potencjału czynnościowego niezbędny jest bodziec o intensywności wystarczającej do zdepolaryzowania neuronu powyżej określonej wartości progowej; wszystkie potencjały czynnościowe w danej komórce mają tę samą wielkość. Między początkiem bodźca a początkiem potencjału czynnościowego występuje krótkie opóźnienie, tzw. czas utajenia (latencja). W czasie trwania potencjału czynnościowego neurony stają się niepobudliwe, zaś w czasie występowania hiperpolaryzującego potencjału następczego ich pobudliwość jest zmniejszona. Zjawiska te określa się odpowiednio jako refrakcję bezwzględną i względną. Zjawiska refrakcji stanowią ograniczenie dla maksymalnej częstotliwości, z jaką neuron może wytwarzać potencjały czynnościowe. Zapobiega to sumowaniu potencjałów czynnościowych i zapewnia przewodzenie potencjałów czynnościowych w aksonie tylko w jednym kierunku.
2.Specyfika krążenia płucnego.
Krążenie płucne spełnia trzy główne zadania:
zapewnia wymianę gazową między powietrzem pęcherzykowym a krwią
stanowi znaczny i o zmiennej objętości zbiornik krwi
uczestniczy w ogólnoustrojowych, metabolicznych regulacjach
Część układu krążenia obejmująca: prawy przedsionek serca, prawą komorę serca, pień płucny, tętnice płucne i żyły płucne (prowadzące krew do lewego przedsionka serca). W krążeniu płucnym (małym), odwrotnie niż w krążeniu dużym, tętnice prowadzą krew odtlenowaną, a żyły krew utlenowaną.
Ciśnienie napędowe, utrzymujące krew w ruchu i użyte na pokonywanie wszystkich oporów, wynosi zaledwie 8-10 mm Hg. Krążenie płucne przy tak małej różnicy ciśnień jest możliwe dzięki bardzo małym oporom, brak tu naczyń oporowych, a wszystkie naczynia tętnicze są łatwo rozciągliwe i elastyczne. Największy opór występuje w naczyniach włosowatych, lecz dzięki wpływom oddechów jest on również nieznaczny. Wpływ oddechów polega na tym, że spadek ciśnienia w jamie opłucnej powodujący wdech, czyli rozszerzenie pęcherzyków płucnych, zwiększa również światło naczyń włosowatych oplatających pęcherzyki. W czasie wdechu w ciągu dwóch kolejnych skurczów serca, w płucnych naczyniach włosowatych i naczyniach żylnych gromadzi się dodatkowa objętość krwi. Dopiero podczas wydechu, wskutek zmian ciśnienia w jamie opłucnej i elastyczności tkanki płucnej oraz naczyń krwionośnych, dodatkowa objętość krwi opuszcza płuca żyłami płucnymi, docierając do przedsionka i lewej komory. Dzięki temu w czasie wydechu następuje zawsze wzrost objętości wyrzutowej lewej komory i wzrost ciśnienia krwi w tętnicach krążenia dużego.
Inną swoistą cechą krążenia płucnego jest prawie całkowity brak filtracji w obszarze naczyń włosowatych.
Następną swoistą cechą krążenia płucnego jest odmienna reaktywność mięśniówki naczyń krwionośnych. Hipoksja, która w dużym układzie krążenia jest głównym czynnikiem powiększającym przepływ przez naczynia włosowate, w krążeniu płucnym działa odwrotnie, to znaczy kurczy mięśniówkę gładką naczyń.
3.Procesy wchłaniania w jelicie grubym.
Jelito grube stanowi odcinek przewodu pokarmowego, w którym nie zachodzi wchłanianie produktów trawienia. Natomiast poza gromadzeniem i wydalaniem kału jelito grube spełnia istotną rolę w procesie wchłaniania wody i elektrolitów, nie jest więc wyłącznie zbiornikiem "na resztki", ale elementem układu pokarmowego aktywnie uczestniczącym w zachowaniu homeostazy organizmu.
Trawienie w wyższych partiach przewodu pokarmowego wymaga środowiska wodnego. W jelicie cienkim znajduje się więc, bez względu na to, ile wody zostaje przyjęte z pokarmem, pewna ilość wody wraz z rozpuszczonymi w niej solami mineralnymi przyjętymi oraz wydzielonymi z sokami trawiennymi. W jelicie grubym dochodzi do odzyskiwania tych związków. I tak do światła jelita grubego dostaje się około półtora litra płynu na dobę, wraz z 200 mmol sodu, 100 mmol chlorków oraz 10 mmol potasu. Z tej puli 90% jest wchłanianych zwrotnie i wraca do krwioobiegu. Sód, najsilniej absorbowany ze światła jelita grubego, wchłaniany jest przeciwko gradientowi stężeń (jego stężenie w kale jest niższe aniżeli w surowicy krwi) i jest to proces aktywny. Wchłanianiu sodu towarzyszy aktywna eliminacja jonów potasu, których zawartość w kale jest wyższa niż w surowicy krwi. Wiąże się to z przeciwnym do ruchu potasu kierunkiem przenikania sodu - te dwa procesy są ze sobą powiązane.
Wchłanianie wody jest procesem biernym, towarzyszy ruchowi sodu. Większość wody i sodu wchłaniana jest w proksymalnym odcinku okrężnicy.
5.Wydalanie moczu.
Przechodzenie moczu do pęcherza moczowego.
Drogami wyprowadzającymi mocz są kanaliki zbiorcze, miedniczka nerkowa oraz moczowody, pęcherz moczowy i cewka moczowa. W zależności od gatunku zwierząt miedniczka obejmuje brodawkę bezpośrednio (pies kot owca koza koń-nerka jednobrodawkowa ; bydło brak miedniczki a kielichy są uzupełnieniami moczowodu (nerka wielobrodawkowa). Przechodzenie moczu w kierunku moczowodu jest dzięki skurczom mm. gładkich kielicha nerkowego. Skurcze te powodują zmianę ciś. hydrostatycznego w tzw. przestrzeni martwej (kanaliki zbiorcze, miedniczki, moczowody). Cofaniu się moczu z miedniczki zapobiega obecność mięśnia zwieracza kielicha. Moczowody transportują mocz do pęcherza moczowego. Wykonują one ruchy
perystaltyczne rozchodzące się falami co 5-20s przesuwając mocz z prędkością 2-3cm/s. Bodźcem do perystaltycznego ruchu jest wypełnienie ich płynem. Ciśnienie wew. moczowodu jest niskie (10mmHg). Nerwy regulujące motorykę należą do układu autonomicznego, jednakże istnieje pewien automatyzm , o którym świadczy zdolność do prawidłowej czynności nawet po wyizolowaniu. Mają one również unerwienie czuciowe.
czynności pęcherza moczowego.
Pęcherz moczowy jest zbiornikiem moczu i narządem umożliwiającym jego wydalanie. Główny mięsień, będący mięśniem wypierającym mocz, jest zbudowany z sieci mięśni gładkich. W okolicy dna pęcherza (ujście wewnętrzne cewki moczowej) włókna zbiegają się nad nim łukowato, tworząc pasmo- Mięsień zwieracz wewnętrzny. .Mięsień zwieracz zewnętrzny (zwieracz cewki moczowej) jest mięśniem poprzecznie prążkowanym zamykającym ujście cewki moczowej. Ciśnienie wewnątrz pęcherza pustego jest równe ciś. śródbrzusznemu. W miarę napływania do niego moczu ściany ulegają rozszerzeniu (nabłonek przejściowy). Mocz nie cofa się ponieważ moczowody wnikają do niego skośnie a ich ujścia osłania fałd błony śluzowej. Zdolność pęcherza do adaptacji do narastającej ilości płynu bez zmiany ciśnienia wewnątrz umożliwia prawidłowe przesączanie kłębowe oraz przepływ moczu z moczowodów. Ta adaptacja łączy się z rozluźnieniem mięśni i nie zależy ona od układu nerwowego. Fizjologiczna pojemność pęcherza moczowego (znoszona bez dolegliwości) wynosi od 250 do 500 ml. Mocz przebywający w pęcherzu nie ulega większym zmianom (może ulegać niewielka zmiana ilości sodu i wapnia) dzięki nieprzepuszczalności błony śluzowej względem jego składników.
unerwienie pęcherza moczowego.
Unerwienie ruchowe jest i z układu autonomicznego jak i somatycznego. Włókna współczulne pochodzą z lędźwiowego odcinka rdzenia kręgowego (L2-L5) i tworzą nerw podbrzuszny. Powodują one zwolnienia napięcia z mm. czyli ułatwiają zwiotczenie i rozszerzanie pęcherza moczowego. Jednocześnie zwiększają one skurcz m. zwieracza wew. i zewn. włókna ruchowe przywspólczulne pochodzą z segmentu krzyżowego (S3-S4) i tworzą nerw miedniczny. Somatyczne unerwienie m zwieracza zewn.pochodzi od nerwu sromowego. Włókna tych nerwów powodują wzrost napięcia mięsni wypierających mocz, zwalniając przy tym napięcie zwieraczy wew. i zew., ułatwiając wydalanie moczu. Nerwami podbrzusznymi przenoszone jest uczucie bólu i temperatury. Włókna dośrodkowe nerwu miednicznego przewodzą impulsy sygnalizujące o stanie wypełnienia pęcherza moczem. Nerw sromowy przenosi głębokie i powierzchowne bodźce czuciowe z pęcherza.
Ośrodki nerwowe kontrolujące oddawanie moczu.
Wydalanie moczu z pęcherza jest aktem odruchowym. Odruchem tym kierują ośrodki znajdujące się w korze mózgowej, podwzgórzu, pniu mózgu i rdzeniu kręgowym. U zwierząt przyuczonych (np. pies) oddawanie moczu jest odruchem zależnym od woli. U innych odbywa się mimowolnie bez udziału wyższych ośrodków. Główne ośrodki kierujące wydalaniem moczu znajdują się w rdzeniu kręgowym. ponadto dużą role w regulacji czynności odgrywają ośrodki podkorowe : ułatwiające wydalanie moczu z pęcherza oraz utrudniające. Ośrodki ułatwiające mieszczą się głównie w górnej części mostu, a hamujące w śródmózgowiu. Prawidłowe wydalanie moczu jest uwarunkowane równowagą czynności tych ośrodków podkorowych. Ośrodki, od których zależą świadome kontrolowanie tego odruchu leżą w korze mózgowej, w tylnej części płata czołowego. Jest jeszcze korowy ośrodek odczuwania parcia. Wydalanie moczu zależy od regulacji antagonistycznych reakcji mięśni wypierających mocz i mięśni zwieracza.
Mechanizm oddawania moczu
W ścianie pęcherza mieszczą się receptory wrażliwe na rozciąganie. Wartością progową dla nich jest ciśnienie wewn. pęcherza rzędu 14mmHg wywołujące czucie parcia. Ilość moczu jednak nie decyduje gdyż progowe ciśn. może powstać przy 50 jak i 500ml moczu. O czuciu parcia decyduje ciśnienie moczu i rozciąganie ścian pęcherza. W chwili dowolnego oddawania moczu wydalanie może być wspomagane tłocznią brzuszną. Natomiast dowolne przerwanie oddawania moczu spowodowane jest skurczem mięśnia dźwigacza odbytu, który unosząc dno pęcherza wywołuje skurcz mięśni zwieracza oraz zwiotczenie mięśni wypierających mocz. Przytłumienie czucia parcia związane jest z wpływem korowych ośrodków na mięśnie zwieracza zew. cewki moczowej. Jednak przy znacznym rozciągnięciu ścian pęcherza dalsze przytłumienie jest niemożliwe i dochodzi do mimowolnego oddania moczu. Pęcherz moczowy nie opróżnia się całkowicie. Pozostaje niewielka ilość moczu (1-2%) tzw. mocz resztkowy. Jego ilość zależy od sprawności skurczowej mm. wypierających mocz która z wiekiem maleje (obniżenie się napięcia mięśniowego). Ogólnie wydalanie moczu u samic trwa krócej ze względu na budowę cewki moczowej. Częstość oddawania moczu w okresie pourodzeniowym, a także u osobników starszych jest większa.
4.Wymień i scharakteryzuj podstawowe formy zachowania się zwierząt.
Zachowanie się zwierząt to każda uzewnętrzniająca się reakcja organizmu lub ogół reakcji o charakterze odruchu na bodźce zewnętrze lub o charakterze samoistnym, taj zachodząca pod wpływem popędu. Reakcją może być mimika poza , ruch, (które mogą występować razem np. w pozach i popisach zewnętrznych), autotomia (odruchowe , czynne odrzucenie przez zwierzę części ciała) , fonacja ( u wielu zwierząt mająca znaczenie w porozumiewaniu się ) zmiana barwy , zmiana własnego pola elektrycznego (np. u ryb elektr.) , usuwanie kału moczu, wydzielanie subst.chemicznych (np.cieczy , wosku , nici przednich) ,broluminiscencja.
Elementarne rodzaje zachowań się zwierząt mają charakter stosunkowo prostych dziedzicznych reakcji odruchowych jak ; odruchy obronna, kinezy (ruchliwość lokomotoryczna zwierząt spowodowana wewnętrzną motywacją (np. popędem) taka kineza może stanowić początkowe ogniwo zachowania apetencyjnego wielu instynktów ( jak pokarmowego , seksualnego czy snu ), taksje , tropizmy.
Złożone wzorce zachowań zwierząt(neurofizjologiczne podstawy) są zwykle oparte na instynkcie, modyfikowane są przez uczenie a nawet inteligencje i są zależne od popędu.
Wrodzonym mechanizmem zachowanie się zwierząt jest INSTYNKT wrodzony zespół aktów ruchowy i złożonych form zachowania się charakt. Dla danego gat. Zw. Wywołanych przez bodźce zewn. I wew. Oraz zachodzących na tle podwyższonej pobudliwości określonych ośrodków nerwowych. Służą one zaspokajaniu potrzeb organizmu jak odżywianie się (instynkt pokarmowy) , ochrona przed czynnikami szkodliwymi (instynkt samozachowawczy) ,zachowanie gat. ( instynkt płciowy) dążność do swobody środowiska ( instynkt badawczy).
Instynkty decydują o specyficznym zachowaniu się, które nie wymaga wcześniejszej nauki ( zachowanie instynktowne).Zachowanie instynktowne służą zaspokajaniu potrzeb organizmu i zwykle towarzysza im mniejsze lub większe emocje.
MOTYW, POPĘD, EMOCJE: akty zachowania się zwierząt opierają się na czynnościach nabytych podczas przebywania zwierząt w danym środowisku , motywowanym przez określone potrzeby biologiczne. Liczne formy zachowania , służące zaspokajaniu potrzeb pokarmowych , płciowych , kontaktu z nowym otoczeniem ,potrzeby zabawy , są w odróżnieniu od instynktów formami nabytymi.
Mechanizm zaspokajania potrzeb zaczyna się od motywu czyli wew. Stanu org. Uruchamiającego i organizującego zachowanie się zwierzęcia do zaspokajania określonej potrzeby biologicznej.
Zachowanie determinowane motywami nosi nazwę zachowania motywacyjnego. Motyw składa się z 2 komponentów :popędu( przejawia się odpowiednimi czynnościami ruchowymi i jest motorem działania w kierunku osiągnięcia określonego celu) , drugim komponentem jest chęć osiągnięcia określonego celu lub zdobycia nagrody.
Popędy można podzielić na :
-apetytywne (dodatnie) ukierunkowujące działalność organizmu na osiągnięcie kontaktu z bodźcami atrakcyjnymi ( z pokarmem , wodą , partnerem płciowym)
-awersyjne 9ujemne, obronne) ukierunkowują aktywność organizmu na obronę własnego ciała przed działaniem szkodliwych czynników (reakcją jest strach wyzwalający reakcję ucieczki i wściekłość wywołująca reakcję agresji)
Motywom i popędom towarzyszą przeżycia ( uczucia) przykre lub przyjemne. Uczucia o dużym nasileniu są emocjami :
-dodatnie ; radość zadowolenie
-ujemne; gniew , smutek , obawa, przygnębienie.
(Istnieją teorie wg których emocje kompensują deficyt informacji koniecznych do osiągnięcia celu oraz zaspokojenia popędu.
1)Wiliam Jolwes
emocje : postrzegane reakcje organizmu .bodźce emocjonalne pobudzają korę mózgową a ta ruchową część i następuje reakcja fizjologiczna postrzegana jako emocje.
Uszkodzenie kory mózgowej zmniejsza kontrole na emocjami
2)Canon Bard
Za reakcje emocjonalne odpowiedzialne jest podwzgórze.
3)Papez
Powstawanie emocji zachodzi w pętli zaczynającej się od tylnej części wzgórza i przez sygnały zmysłowe docierają do kory i podwzgórza. Kora uczuciowa i podwzgórze wpływa na zakręt obręczy , który przesyła ja przez korę ruchową do hipokampa przez jądro przegrody do ciał suteczkowych i jąder podwzgórza odpowiedzialnego za reak.fizjiol.)
Emocjom towarzyszą subiektywne przeżycia oraz obiektywne zmiany cielesne ; reakcje ruchowe, postawne , głosowe i tzw.ekspresje mimiczne oraz wew.zmiany fizjologiczne (strachowi: przyspieszona akcja serca, oddychanie , pocenie się , rozszeżenie źrenic). Rodzaj emocji można poznać u zwierząt po charakterystycznym ustawieniu uszu ( pies , kot ) ruchach ogona ( pies , kot ), ruchach kończyn ( „grzebanie” u koni) jeżeniu się sierści 9 pies , kot , także konie i bydło)po wydawaniu odpowiednich głosów (szczekaniu, warczeniu u psa miauczeniu i kota).
Nowe formy zachowania mogą być wynikiem uczenia się czyli nabywaniem doświadczenia ,wprawy i umiejętności w zakresie określonych czynności pod wpływem ich wykonywania ich.W procesie uczenia się zachodzi odbieranie informacji w ośrodkowym układzie nerwowym i takie jej gromadzenie w różnych strukturach mózgu by mogła ona później ulegać wybiórczej mobilizacji i reprodukcji..Uczenie się zachodzi na podłożu aktywności różnych struktur i układów OUN zwłaszcza kory mózgowej i wzgórza układu rąbkowego i siatkowego.
Z tymi ośrodkami związane są wszelkie rodzaje zachowań. W podwzgórzu są ośrodki nerwowe pobudzane przez różne wpływy środowisk zewn lub wewn generujących takie lub inne motywy i popędy. Są to ośrodki głodu , sytości , pragnienia , popędu płciowego , ,macierzyństwa , ośrodki ucieczki , ,agresji i termoregulacji. Drażnienie tych emocji powoduje czynności (zachowania) zmierzające do zaspokojenia potrzeb biologicznych.
2.Podział odruchów i znaczenie.
Odruch jest automatyczną stereotypową odpowiedzią ustroju na podrażnienie określonych receptorów zewnętrznych lub wewnętrznych, zachodzącą przy udziale ośrodkowego układu nerwowego(rdzenia kręgowego lub mózgowia).
Odruchem nazywamy reakcję efektora (ruchową, wydzielniczą) występującą przymusowo na bodźce zewnętrzne lub wewnętrzne, zachodzącą przy udziale układu nerwowego.
Anatomicznym podłożem niezbędnym do wystąpienia reakcji odruchowej jest łuk odruchowy.
Każdy łuk odruchowy składa się z pięciu zasadniczych elementów:
1)receptora
2)dośrodkowej drogi doprowadzającej (aferentnej)
3)ośrodka nerwowego
4)odśrodkowej drogi wyprowadzającej (eferentnej)
5)efektora.
Warunkiem wystąpienia reakcji odruchowej jest ciągłość łuku odruchowego i normalna funkcja wszystkich jego części składowych
Odruchy można najogólniej podzielić na: animalne (somatyczne) i autonomiczne (wegetatywne)
Animalne- efektorem jest skurcz mięśni szkieletowych
Autonomiczne- odruchy wydzielnicze ,,naczynio ruchowe, skurcze mięśniówki gładkiej narządów wewnętrznych.
Odruchy można również podzielić na: bezwarunkowe (wrodzone) i warunkowe(nabyte w życiu osobniczym).
Odruchy wrodzone są zawsze tymi samymi reakcjami na ten sam bodziec, niezmiennymi w ciągu życia.
Odruchy nabyte wykazują dużą zmienność w odpowiedzi na bodźce: mogą zanikać i znowu powstawać.
W zależności od efektu, który powoduje pobudzenie lub hamowanie efektora odruchy dzieli się na:
a)odruchy pobudzeniowe
b)odruchy hamulcowe
W zależności od liczby synaps w łuku odruchowym odruchy możemy podzielić na:
a) monosynaptyczne
b) wielosynaptyczne (polisynaptyczne)
W zależności od poziomu lokalizacji ośrodka odruchu w CSN odruchy dzielimy na:
a) odruchy rdzeniowe
b) odruchy mózgowe
Zasadniczymi rdzeniowymi odruchami ruchowymi są jednosynaptyczne odruchy rozciągowe (miotatyczne) i wielosynaptyczne odruchy zginania lub prostowania.
Pierwsze zaliczane są do odruchów własnych, ponieważ receptory ich leżą w tym samym mięśniu co efektor. Drugie nazywane są odruchami obcymi, dlatego że ich receptory znajdują się w skórze lub błonie śluzowej, leżą więc poza reagującymi mięśniami.
Odruchy rozciągowe wywoływane są pobudzeniem wrzecion mięśniowych (zakończenia pierścieniowo-spiralne), z których impulsy wędrują poprzez dwuneuronowy, jednosynaptyczny łuk odruchowy do tych samych mięśni, w których wrzeciona uległy pobudzeniu.
Impulsy z wrzecion mięśniowych aktywują synchronicznie motoneurony jednego segmentu rdzeniowego, stąd przy odruchach rozciągowych nie dochodzi do szerzenia się stanu pobudzenia w rdzeniu (promieniowanie, irradiacja pobudzenia), a reakcją odruchową jest zawsze skurcz pojedynczy mięśnia.
Przykładem odruchu rozciągowego jest odruch kolanowy (rzepkowy). Uderzenie młoteczkiem opukowym w ścięgno mięśnia czworogłowego uda poniżej rzepki powoduje krótkotrwałe rozciągnięcie mięśnia, przez co pobudza receptory jego wrzecion mięśniowych i wywołuje odruchowy jego skurcz (wyprostowanie nogi w stawie kolanowym).
Skurczowi mięśni prostowników podczas odruchów rozciągowych towarzyszy zawsze zwiotczenie mięśni zginaczy w tej samej kończynie. Podczas pobudzania neuronów zaopatrujących jeden mięsień ulegają hamowaniu neurony unerwiające mięsień antagonistyczny. Zjawisko to nazywane jest hamowaniem antagonistycznym, opierającym się na wzajemnym unerwieniu zwrotnym. Jeśli nerw dośrodkowy pobudza ośrodek skurczu mięśni prostowników, to jednocześnie uruchamia połączenie z ośrodkiem mięśni zginaczy tej samej kończyny. Połączenie to zawiera hamujący neuron wstawkowy, kończący się synapsą hamującą na neuronie ruchowym ośrodka mięśni zginaczy. Wskutek tego dochodzi do zwiotczenia mięśni antagonistycznych.
Odruch źreniczny (bezwarunkowy wegetatywnym, wielosynaptyczny, obcym).Odruchowe zwężanie lub rozszerzanie źrenicy reguluje ilość światła wpadającego na siatkówkę oka w zależności od warunków oświetlenia. Zwieracz źrenicy unerwiony jest przez układ przywspółczulny a ośrodek zwężania źrenic leży w śródmózgowiu.Źrenice zwężają się pod wpływem: intensywnego oświetlenia, wpatrywania się w przedmioty blisko leżące, podczas snu, w czasie prawidłowej narkozy, zwiększonego napięcia układu przywspółczulnego oraz pod wpływem niektórych środków farmakologicznych, np. ezeryna, morfina, pilokarpina.
Mięsień rozszerzający źrenicę unerwiony jest współczulnie, a ośrodek rzęskowo-rdzeniowy (ośrodek rozszerzania źrenicy)znajduje się w 2-3 pierwszych odcinkach piersiowego rdzenia kręgowego Słabe oświetlenie oraz patrzenie w dal powoduje rozszerzenie źrenic. Rozszerzają się one również pod wpływem, bólu, emocji, duszności, zbyt głębokiej narkozy. Również podanie atropiny, kokainy powoduje rozszerzenie źrenic.
1.Żółć- rodzaje, skład i znaczenie.
Głównymi organicznymi składnikami żółci są so1e kwasów żółciowych. U zwierząt mięsożernych i wszystkożernych przeważa kwas glikocholowy, u roślinożernych, a szczególnie u bydła, dużo jest kwasu taurocholowego. Znaczna część kwasów żółciowych jest wchłaniana z powrotem z dalszych odcinków jelita cienkiego do krwi i przenoszona do wątroby. Zjawisko to nosi nazwę krążenia soli kwasów żółciowych.
Barwniki żółciowe są produktami rozpadu hemu w układzie siateczkowo-śródbłonkowym. Bilirubina jest wychwytywana przez komórki wątrobowe i wydzielana do żółci, w której stężenie tego barwnika kilkaset razy przewyższa jego stężenie w krwi. Bilirubina i biliwerdyna pod wpływem działania enzymów bakteryjnych przechodzą w jelicie grubym w barwnik kału -sterkobilinę, który częściowo jest wchłaniany do krwi, dając barwnik moczu -urobilinogen.
W żółci występują wolny oraz zestryfikowany cholesterol, lecytyny, tłuszcz obojętny, wolne kwasy tłuszczowe i ślady białka.
Jest to płynna wydzielina wątroby, jedna z substancji wspomagających soki trawienne.
Jest to zielonkawo-brunatny, lepki i kleisty płyn, o odczynie obojętnym. Zawiera barwniki żółciowe, kwasy żółciowe i ich sole, cholesterol, lecytynę, mocznik, sole mineralne, sole kwasów tłuszczowych. Żółć jest niezbędna w procesie emulgowania tłuszczów, czyli rozbijania ich na drobną zawiesinę umożliwiającą trawienie.
Wydzielana przez wątrobę, a magazynowana w pęcherzyku żółciowym, z którego spływa do dwunastnicy po zadziałaniu cholecystokininy, w czasie trawienia pokarmów.
Żółć wpływa na aktywność niektórych enzymow w swietle jelita wspomaga trawienie i wchłanianie tłuszczów oraz witamin rozpuszczalnych w tłuszczach: A, D, E, K i F. regulacja ruchow przewodu pokarmowego, procesow sekrecyjnych bezpośrednio lub poprzez uwalnianie hormonow jelitowych (sekretyna). Razem z żółcią wydalany jest cholesterol, kwasy żółciowe, leki, toksyny, barwniki żółciowe i substancje nieorganiczne, takie jak Cu, Zn i Hg.
Wydzielone do jelita kwasy żółciowe po zakończeniu swego udziału w trawieniu podlegają reabsorpcji w procesie krążenia wątrobowo-jelitowego.
W pewnych stanach zaburzeń chorobowych dochodzi do odkładania jej składników, w wyniku czego wytrąca się rozpuszczalny kamień żółciowy, prowadząc do schorzenia nazywanego kamicą pęcherzyka żółciowego lub kamicą dróg żółciowych.
, Zolc stale wydzielana jest do kanalikow żółciowych przez hepatocyty (kanalikowa frakcja zolci) W okresie miedzy posiłkowygm, zolc gromadzi się w pecherzyku żółciowym, po posilku najintensywniej oproznia swa zawartosc.
Wyrozniamy :
-zolc pecherzykowa : gromadzona i wydalana z pęcherzyka żółciowego
- zolc wątrobowa : plynaca przewodami wątrobowymi, omijajaca pęcherzyk.
W koncowym odcinku jelita biodrowego kwasy żółciowe SA wchłaniane , wędrują do watroby, tworząc: pętle krążenia wątrobowo jelitowego. W regulacji glowna funkcja - wpływy nerwowo humoralne. Uklad parasympatyczny wpływa stymulująco na opróżnianie pęcherzyka, hormony tarczycy, trzustki, sekretyna - reguluja procesy sekrecji zolci
Homeostaza (gr. homoíos - podobny, równy; stásis - trwanie) - zdolność do utrzymania stanu równowagi dynamicznej środowiska, w którym zachodzą procesy biologiczne. Zasadniczo sprowadza się to do równowagi płynów wewnątrz- i zewnątrzkomórkowych. Pojęcie homeostazy wprowadził Walter Cannon w 1939 roku na podstawie założeń Claude Bernarda z 1857 r. dotyczących stabilności środowiska wewnętrznego. Homeostaza jest podstawowym pojęciem w fizjologii. Pojęcie to jest także stosowane w psychologii zdrowia dla określenia mechanizmu adaptacyjnego.
temperatura ciała (u organizmów stałocieplnych),
stężenie związków chemicznych w płynach ustrojowych,
ciśnienie tętnicze krwi,
ciśnienie parcjalne tlenu i dwutlenku węgla we krwi.
Kontrola wymienionych parametrów odbywa się poprzez receptory (głównie chemoreceptory), które informacje o wartości określonego parametru przekazują do interpretatora (np. w przypadku temperatury ciała ssaków do podwzgórza), gdzie dokonuje się porównanie wartości wykrytej ze stałą wartością prawidłową (tzw. punktem nastawczym) lub, co zdarza się częściej, z jej przedziałami akceptowalnymi. Jeśli aktualny stan parametru jest zbyt wysoki lub zbyt niski, centrum integrujące wymusza na efektorach odpowiedź odpowiednią do sytuacji.
Mechanizmy utrzymania homeostazy Mechanizmy wytwarzania odpowiedzi i tym samym regulacji wartości parametru można podzielić na dwie grupy:
fizjologiczne (np. zwiększenie częstotliwości skurczów mięśni w celu podwyższenia temperatury) oraz
behawioralne (np. wyjście z cienia, w tym samym celu).
Fizjologiczne mechanizmy opierają się na sprzężeniu zwrotnym (ang. feedback):
ujemnym
W wyniku tego sprzężenia zwrotnego następuje zmiana wartości parametru na zbliżoną do punktu stałego. Zasadniczo osiągnięcie wartości punktu nastawczego jest niemożliwie, toteż wartości zawsze wymagają regulacji (tzw. ang. hunting about the norm). Możliwe jest modyfikowanie wartości punktu stałego w wyniku adaptacji.
wyprzedzającym (wczesnym)
W wyniku tego sprzężenia zwrotnego występuje reakcja na zmiany parametru kontrolowanego, pomimo że w chwili odpowiedzi wartość parametru pozostaje jeszcze w zakresie wartości akceptowalnych (np. picie wody w czasie jedzenia przez szczury). Wymaga działania celowego - przewidywania.
dodatnim
W wyniku tego sprzężenia zwrotnego występuje reakcja na bodziec, poprzez pogłębienie wartości nieprawidłowej dla innego celu (np. odczuwanie bólu na poziomie neurotransmiterów). W tym sprzężeniu pewne parametry są regulowane, podczas gdy inne mogą przyjmować wartości różne od akceptowalnych i nie podlegają regulacji (ale pozostają pod kontrolą).
4.Regulacja ciśnienia krwi.
REGULACJA CIŚNIENIA KRWI. -zależy od regulacji pracy serca i regulacji światła naczyń (regulacja obu czynników na drodze nerwowej i humoralnej).
REGULACJA CIŚNIENIA NA DRODZE NERWOWEJ
regulacja pracy serca odbywa się poprzez odruchy:
1 z baroreceptorów (presoreceptorów) zatoki tętnic szyjnych łuku aorty, przez włókna czuciowe nerwu IX i X impuls dociera do ośrodka hamującego ciśnienie krwi powodując wzrost impulsacji co powoduje wzrost akcji serca spadek ciśnienia spadek impulsacji spadek akcji serca wzrost impulsacji ośrodka hamującego
2 z chemoreceptorów (łuk aorty, zatoki tętnic szyjnych) wrażliwe na : wzrost prężności CO2, spadek prężności O2, wzrost stężeń jonów H+ powoduje pobudzenie ich i wzrost akcji serca.
3 z końcowych naczyń żylnych i przedsionka: wzrost napływu krwi do serca wzrost akcji serca wzrost ciśnienia krwi
B)regulacja światła naczyń przez odruchy z:
1 baroreceptorów (łuk aorty, zatoki tętnicy szyjnej): rozszerzenie naczyń spadek ciśnienia krwi
2 chemoreceptorów : zwężenie naczyń wzrost ciśnienia krwi (nie zwężają się naczynia mięśni szkieletowych, wieńcowe serca i mózgu )
REGULACJA CIŚNIENIA NA DRODZE HUMORALNEJ
humoralna regulacja pracy serca: adrenalina i insulina: akcja serca-wzrost, ciśnienie krwi- wzrost glukagon tyroksyna
humoralna regulacja światła naczyń krw.: noradrenalina wazopresyna prostaglandyna F2alfa angiotensyna <----> bradykinina histamina acetylocholina kw.mlekowy CO2 OGÓLNIE: zwiększenie częstości i siły skurczów serca powoduje wzrost ciśnienia tętniczego. Podobnie działa zwężenie naczyń. KONTROLA PRZEZ OŚRODEK SERCOWY I NACZYNIORUCHOWY.
5.Omów fazy cyklu płciowego.
Przygotowaniem dorosłej samicy do rozrodu jest ruja = estrus, pojawia się cyklicznie w ciągu roku, z częstotliwością zależna od gatunku zwierzęcia i warunków w jakich on przebywa. Okres między rujami nazywamy cyklem płciowym= cyklem rujowym.
Zwierzęta poliestralne mające ruje cyklicznie, wielokrotnie w ciągu całego roku to krowa (długość cyklu 21 dni), Su-21, mysz-4. Zwierzęta poliestralne sezonowo - cykle powtarzają się u nich kilkakrotnie, ale w określonych porach roku np.: owca (wiosna, 17), kotka (wiosna-lato, 14-21). W pozostałych porach roku jest brak rui = anestrus sezonowy. Zwierzęta monoestralne (jeden cykl w roku) i diestralne (dwa cykle w roku) - u psa zależnie od gatunku. Konie - około 25% wykazuje regularne cykle w ciągu całego roku (cykl 21 dni), inne są sezonowo poliestralne z największą aktywnością jajników od maj do września. U zwierząt wolno żyjących sezonowa aktywność rozrodcza zapewnia najlepsze warunki do rozwoju potomstwa.
Głównym czynnikiem regulującym pojawienie i zanikanie cykli rujowych jest gonadoliberyna - GnRH (FSH-RH, LH-RH), jest wytwarzany w neuronach podwzgórza, następnie uwalniany do krwi, jest wynikiem oddziaływania środowiska, a głownie światła i żywienia (długość dochodzenia impulsów świetlnych). Informacje świetlne zakodowane w salwach impulsów o różnej częstotliwości i kształtach docierają do śródmózgowia, a stamtąd do podkorowych i korowych ośrodków nerwowych i do szyszynki. Szyszynka działa hamująco na wytwarzanie GnRH w podwzgórzu, jest ona najbardziej aktywna w ciemności, przy braku stymulacji świetlnej. Wydłużenie dnia hamuje czynność szyszynki co pobudza czynność wydzielniczą podwzgórza. Odwrotnie jest przy skróconym dniu.
Aktywność rozrodcza samic jest hamowana w czasie laktacji, na skutek dużej częstotliwości drażnienia gruczołów mlecznych i uwalniania w wyniku tego oksytocyny i prolaktyny. U Su, owiec brak cykli laktacji = niepłodność laktacyjna.
Cykl rujowy - okres między jedną a następną rują. Dzieli się on na fazę ciałka żółtego = lutealną i fazę pęcherzykową. Pierwszy dzień rui przyjmuje się jako dzień zerowy cyklu. W tym czasie pęcherzyk jajnikowy jest u szczytu rozwoju, a więc w stadium przedowulacyjnym. W 1-2 dniu cyklu musi nastąpić owulacja, a po niej tworzenie ciałka żółtego. Po rui pierwszą fazą cyklu jest faza ciałka żółtego = lutealna. Trwa ona u Su - do 13 dnia, u Bo - do 16 dnia cyklu. W fazie lutealnej rozróżnia się okres tworzenia i rozwoju ciałka żółtego (u Su, Bo - pierwsze 4 dni cyklu) i okres pełnej aktywności wydzielniczej komórek lutealnych ( u Bo 4- 16 dzień cyklu). Po tym okresie jest 1-2 dniowy okres luteolizy, w czasie którego jest gwałtowny spadek stężenia progesteronu we krwi. Po luteolizie zaczyna się krótka (kilkudniowa) faza pęcherzykowa. U Bo od 18 do 21 dnia cyklu i rozwija się i osiąga dojrzałość jeden z pęcherzyków jajnikowych. Inne, o zaawansowanym rozwoju podlegają atrezji. Wyjątkowo krótka faza pęcherzykowa cyklu nie świadczy o tym, że jedynie w tym okresie dojrzewają i rozwijają się pęcherzyki jajnikowe. Ich rozwój jest długi, pobudzany kolejnymi pulsami wydzielania hormonów gonadotropowych. W tym okresie fazy ciałka żółtego pęcherzyki jajnikowe w nieprzerwanym procesie rosną, dojrzewają i wytwarzają nieduże ilości hormonów steroidowych. Jest to jednak rozwój zwolniony. Luteoliza i gwałtowny spadek stężenia we krwi progesteronu stają się bodźcem dla podwzgórza do wzmożonej pulsacji wydzielania gonadoliberyny. Powoduje to zwiększone pulsacyjna uwalnianie FSH i LH. Gonadotropiny z kolei gwałtownie przyśpieszają rozwój jednego u Bo lub około 20 u Su pęcherzyków jajnikowych, przygotowując je do owulacji. Faza pęcherzykowa w cyklu rujowym jest końcową, jedynie krótka fazą przygotowania pęcherzyka do owulacji po jego długim wcześniejszym rozwoju.
U kobiety cykl płciowy trwa 28 dni i jest wyznaczony krwawieniem menstruacyjnym. Bezpośrednio po krwawieniu, w czasie którego wydalane są wraz z krwią na zewnątrz złuszczone fragmenty wcześniej rozbudowanej (w przewidywaniu ciąży) błony śluzowej macicy, rozpoczyna się 13 - 14 dniowa faza pęcherzykowa = proliferacyjna zakończona owulacją. Druga połowa cyklu odpowiada fazie ciałka żółtego = faza sekrecyjna. Jest to okres rozrostu błony śluzowej macicy i jej gruczołów wydzielniczych pod wpływem progesteronu. Przy braku ciąży fazę tę kończy menstruacja.
1.Mechanizm sprzężenia zwrotnego w czynności odruchowej.
W organizmie istnieją układy sprzężenia zwrotnego obwodu z centrum, odgrywające szczególną rolę przy wykonywaniu złożonych aktów ruchowych. Dzięki sprzężeniom zwrotnym jest osiągana dokładność i płynność ruchów oraz ich każdorazowe przystosowanie do zaistniałej sytuacji zewnętrznej, która wywołała dane ruchy. Odruchowa samoregulacja czynności fizjologicznych w organizmie opiera się również o mechanizm sprzężenia zwrotnego. Najczęściej mamy do czynienia ze sprzężeniem zwrotnym ujemnym, przy którym odchylenie czynności od stałego poziomu staje się bodźcem prowadzącym do restytucji zaburzonego stanu. Wynik działania takiego sprzężenia jest ujemny w stosunku do przyczyny wywołującej zaburzenie, a sam jego mechanizm stabilizuje rozpatrywany układ. O ten mechanizm opiera się utrzymanie homeostazy organizmu. Działanie sprzężenia zwrotnego ujemnego przy odruchach obserwuje się wtedy, gdy dopływające z obwodu impulsy, powstające podczas danej reakcji odruchowej (impulsy zwrotne), doprowadzają do następowego jej osłabienia.
Przy sprzężeniu zwrotnym dodatnim organizm reaguje pogłębieniem zmian zapoczątkowanych działaniem bodźca. Wynik działania tego sprzężenia jest dodatni, prowadzi ono do wzmocnienia i utrwalenia powstałego zakłócenia. Dzięki działaniu tego typu sprzężenia dochodzi przy odruchach do nasilenia reakcji odruchowej pod wpływem impulsów zwrotnych. Przy odruchach czynne jest najczęściej zarówno sprzężenie zwrotne dodatnie jak i ujemne. Impulsy zwrotne, powstające np. podczas odruchów mięśniowych, wywołują lub nasilają stan pobudzenia w jednych ośrodkach, a stan hamowania w drugich.
O działanie sprzężenia zwrotnego ujemnego opierają się m.in. odruchy rozciągowe, odruchowe zmiany napięcia mięśniowego oraz samoregulacja pobudliwości neuronów ruchowych za pośrednictwem wstawkowych neuronów hamujących.
4.Mechanizm wchłaniania w jelicie cienkim.
Wchłanianie w jelicie cienkim polega na przechodzeniu produktów trawienia przez nabłonek jelita do krążenia krwi w celu zaopatrzenia tkanek organizmu w niezbędne do życia składniki.
W części wierzchołkowej oraz podstawno-bocznej błony komórkowej enterocytu istnieją liczne wyspecjalizowane mechanizmy transportowe odpowiedzialne za przechodzenie głównie jonów i związków organicznych przez błonę komórkową. Mechanizmy te zachodzą dzięki obecności kompleksów białkowych odpowiedzialnych za transport błonowy. Mechanizmy transportu działają wybiórczo na określone cząsteczki, a dla danej substancji występują tylko na jednej z błon komórki, albo wierzchołkowej, albo podstawno-bocznej. Wśród mechanizmów transportu można rozróżnić transporty: aktywny, wtórny aktywny i bierny.
Transport aktywny opiera się na bezpośrednim zużyciu energii metabolicznej podczas transportu przez błonę komórkową. Podczas transportu aktywnego energia przechowywana w ATP jest wydatkowana na ruch jonów lub molekuł przez błonę komórkową zachodzący wbrew gradientowi elektrycznemu lub chemicznemu. W jelitach cienkim i grubym największe znaczenie ma transport aktywny zależny od aktywności pompy Na,K ATPazy. Ten rodzaj transportu jest obecny we wszystkich pobudliwych komórkach.
Wtórny transport aktywny wykorzystuje jako nośnik elektrochemiczny gradient jonów Na+ pomiędzy obszarami wewnątrz- i zewnątrzkomórkowym. Z mechanizmu tego korzysta wiele białek określanych jako białka symportowe, które charakteryzują się obecnością jednego lub więcej miejsc wiążących jon Na oraz dodatkowe miejsce wiążące selektywnie inną cząsteczkę. Białka symportowe występują na błonie wierzchołkowej enterocytu. Przy użyciu transportu wspólnego z sodem wchłanianiu ulega wiele substancji organicznych, w tym: glukozy, aminokwasów, witamin i kwasów żółciowych. Istnieją jeszcze inne rodzaje aktywnego transportu wspólnego przy udziale białek wymieniaczy lub antyportów. Dla enterocytu wymieniacz sodowo-protonowy lokalizuje się w błonie wierzchołkowej komórki. Innym rodzajem transportu jest wspólny system transportu wtórny w stosunku do innego zależnego transportu aktywnego. Zależy on także od białek transportujących, a jest napędzany siłą gradientu elektrochemicznego uzyskanego dzięki mechanizmowi wtórnego transportu aktywnego. Najlepszym przykładem takiego transportu jest transport aktywny chlorków i dwuwęglanów za pomocą wymieniacza Cl/HCO3.
Transport bierny odbywa się poprzez kanały jonowe błony komórkowej albo bezpośrednio poprzez połączenia ścisłe pomiędzy komórkami. Kanały jonowe zbudowane z białek błony komórkowej stanowią drogę transportu biernego na drodze dyfuzji. Jony przemieszczają się zgodnie z gradientem elektrochemicznym i bez żadnego wydatku energii metabolicznej.
5.Luteoliza.
Wytwarzany w ciałku żółtym progesteron przygotowuje drogi rodne do przyjęcia zarodka, jego implantacji i dalszego rozwoju ciąży. W przypadku braku ciąży utrzymanie ciałka żółtego jest nie potrzebne. W takiej sytuacji u Su w 13-14 dniu, u Bo 16-17 dniu od owulacji w kom. ciałka żółtego rozwija się proces zwyrodnienia tłuszczowego nazywany luteolizą. Ilość wytworzonego progesteronu gwałtownie maleje i maleje jego poziom we krwi. Proces ten jest wynikiem oddziaływania na ciałko żółte luteolitycznego hormonu prostaglandyny F2 ( PGF2 ), wytwarzanego w kom. błony śluzowej macicy. Komórki te uwalniają PGF2 do włosowatych naczyń krwionośnych i chłonnych błony śluzowej macicy, skąd- naczyniami żylnymi i chłonnymi przechodzącymi przez więzadło szerokie macicy- PGF2 podąża do krążenia ogólnego. Cząsteczki PGF2 nie mogą trafić do krwi tętniczej i są one rozkładane podczas przepływu krwi żylnej przez płuca. Krew tętnicza zaopatrująca jajniki nie ma czynnej PGF2. Aby PGF2 mogła dotrzeć do jajnika i spełnić swoje luteolityczne lokalne dla układu rozrodczego zadanie, przenika ona (na zasadzie mechanizmu przeciwprądowego) z krwi żylnej i z chłonki obszaru więzadła szerokiego macicy, spotyka się w krezce jajnikowej z drobnymi, pętlącymi tu odgałęzieniami tętnicy jajnikowej, przenika do ich wnętrza i z krwią tętniczą dociera do ciałka żółtego. Luteoliza w ciągu kilku dni powoduje całkowity zanik czynności wydzielniczej ciałka żółtego. Gdy wnikają do niego fibroblasty, cały gruczoł przekształca się w bezstrukturalne ciałko białawe(corpus albicans), pozostaje przez długi czas w jajniku. Gwałtowny spadek poziomu progesteronu we krwi po luteolizie jest sygnałem dla podwzgórza do zwiększenia częstotliwości i wielkości (amplitudy) pulsów wytwarzanej gonadoliberyny (GnRH). W rezultacie wolno rozwijające się dotychczas pęcherzyki jajnikowe są pobudzane przez FSH i LH do rozwoju i dojrzewania. W ciągu kilku dni wykształca się dojrzały pęcherzyk(i), osiągając stadium przedowulacyjne.
Gwałtowny spadek poziomu progesteronu we krwi powoduje również zmiany w ukrwieniu i czynności błony śluzowej macicy. Rozbudowana błona śluzowa, przystosowana do przyjęcia zarodka, staje się zbędna. U większości gatunków zwierząt następuje jej obumieranie i resorbcja. U naczelnych złuszczeniu błony śluzowej towarzysz pękanie drobnych naczyń krwionośnych. Ułatwia to wydalenie na zewnątrz, łącznie z krwią, martwej błony śluzowej. Jest to krwawienie menstruacyjne, które kończy jeden okres aktywności jajnika, a poprzedza rozpoczynający się nowy rozwój pęcherzyka jajnikowego.
5. Instynkt
Instynkty są reakcjami bardziej elastycznymi niż odruchy i dlatego umożliwiają bardziej zróżnicowane, rodzaje zachowania się zwierząt. Instyngtem nazywamy wrodzony zespól aktów ruchowych i zlożonych postaci zachowania się charakterystycznych dla danego gatunku zwierząt, wywolanych przez bodźce zewnętrzne i wewnętrzne oraz zachodzących na tle podwyższonej pobudliwości określonych ośrodków nerwowych.
Przykładowe instynkty: pokarmowy, pobierania wody, pobierania soli mineralnych (np. soli z lizawki), oddechowy (u zwierząt wtórnie wodnych, gdyż u zwierząt lądowych oddychanie jest nieskomplikowanym odruchem), pielęgnacji ciała, rozrodczy (wraz z terytorializmem) i opieki nad potomstwem, snu.
Podwzgórze, hormony, wplyw czynników zewnętrznych, reakcje te są wrodzone i zaprogramowane.
4.Specyfika krążenia wątrobowego.
Całkowita ilość krwi przepływająca przez wątrobę człowieka wynosi ok.1500-2000ml/min. Cecha najbardziej charakterystyczna krążenia wątrobowego jest krążenie utworzone przez siec dziwną żylno-żylną. Krew tętnicza z tętnicy wątrobowej miesza się z krwią żylną z żyły wrotnej we wspólnych dla obu naczyniach- naczyniach włosowatych. Krew żyły wrotnej posiada niskie ciśnienie ok.6-12 mm Hg i obniża się do 3-4mm Hg w wątrobowych nacz. Włosowatych. Natomiast krew z tętnicy wątrobowej płynie tętniczkami miedzyzrazikowymi, miedzypłacikowymi, śródpłacikowymi pod dość znacznym ciśnieniem. Spotkanie się obu strumieni krwi jest możliwe, dzięki różnym objętościom wpływającej tu krwi żylnej i tętniczej. Żylna krew wrotna płynąca z przewodu pokarmowego i śledziony stanowi ok. 70-80%, a tętnicza krew ok. 20-30% ogólnej objętości krwi.
Wspólne naczynie włosow., do którego wpływa krew tętnicza i żylna ma szczególną budowę tzn. tworzy nieregularne zatokowe struktury i zbud. jest ze śródbłonka o dużej przepuszczalności, typu okienkowanego o bud. ciągłej i nieciągłej, z dużą ilością porów i szczelin. Cecha charakt. jest to ze każdy hepatocyt , przylega swoja jedna strona pokryta mikrokosmkami do nacz. Włosowatego. Powierzchnia hepatocytu przylegająca do nacz. Włosowatego jest powierzchnia pobierającą- z krwi docierają tu tlen oraz glukoza, aminokwasy, zw. tłuszczowe itp., i jest jednocześnie powierzchnia wydzielniczą, przez którą hepatocyty przekazują do krwi utworzone w nim peptydy i białka( wszystkie białka osocza z wyjątkiem immunoglobulin), mocznik i inne.
Hepatocyty ułożone są w wątrobie w taki sposób, że kiedy komórka powierzchnia jednej strony bocznej styka się bezpośrednio ze ścianka nacz.włosow., to przeciwległa wykazująca wgłębienie- styka się z taka sama strona drugiego hepatocytu.
Oba wgłębienia hepatocytow tworzą przewodzik żółciowy.
W obrębie syncytium, jakie tworzą kom. Śródbłonka, wyst. Kom. gwiaździste ukł. Siateczkowo-śródnabłonkowego. Są to osiadłe makrofagi, które wychwytują i niszczą antygeny, zużyte krwinki czerwone i inne szkodliwe dla org. cząsteczki podlegające fagocytozie.
Zmieszana krew żylna i tętnicza docierają do żyły ośrodkowej, umieszczonej w środku płacika wątrobowego. Stąd odpływa do żył wątrobowych uchodzących do żyły głównej.
Czynnikami reguł. Przepływ krwi przez wątrobę, poza ukł. Adrenergicznym i cholinergicznym, są liczne związki chemiczne wchłaniane z przewodu pokarmowego i docierające do wątroby z krwią wrotna hormony np. insulina, glikagon i hormony przewodu pokarmowego- cholecystokinina ,VIP,sekretyna, gastryna.
7. Filtracja klębkowa
Jest to proces fizyczny polegający na przechodzeniu przez blonę filtracyjną z osocza do przestrzeni klebka, wody oraz wszystkich rozpuszczalnych w niej substancji. W przesączu klebuszkowym tzw moczu pierwotnym wyst związki o 70 tys mas cząst. Nie przenikają bialka, jedynie male ilosci albuminy. Wielkość filtracji zależy od calkowitej powierzchni filtracyjnej nefronow, przepuszczalności blony filtr oraz ciśnienia filtracyjnego glownie hydrostatycznego.
Pierwszym elementem filtru jest śródblonek naczyń wlosowatych zajmujących elementy morfotyczne krwi. Drugą barierę stanowi blona podstawna komórek torebki (blaszki wewnętrznej), uniemożliwiająca filtrację większości cząsteczek bialkowych. Trzecią warstwę filtru stanowi cienka blona zamykajaca szczeliny miedzy wypustkami stepowatymi komórek nablonkowych blaszki wewnętrznej.
8. Podwzgorzowe ośrodki regulacji temperatury
W podwzgórzu po obydwu stronach trzeciej komory mózgu i w szyjnej części rdzenia kręgowego znajdują się termodetektory - struktury termowrażliwe reagujące na zmianę temperatury docierającej do nich krwi poprzez przyspieszenie lub zwolnienie czynności bioelektrycznej.
Ośrodek termoregulacji znajduje się w podwzgórzu i jest dwuczęściowy. W przedniej części podwzgórza mieści się ośrodek eliminacji ciepła, który kontroluje utratę ciepła. W tylnej części podwzgórza znajduje się ośrodek zachowania ciepła, który reguluje zatrzymywanie ciepła w ustroju i stymulację jego produkcji. Obie te części ośrodka termoregulacji są ze sobą połączone drogami biegnącymi po obu stronach bocznej części podwzgórza. Uszkodzenie tylnej części podwzgórza znosi reakcje termoregulacyjne chroniące organizm przed oziębianiem (np. skurcz naczyń krwionośnych skóry, wzrost tempa przemiany materii), nie zmieniając zdolności eliminacji ciepła z ustroju w gorącym środowisku. W tylnej części podwzgórza odbywa się przetwarzanie informacji termicznych z termoreceptorów obwodowych. W przedniej części podwzgórza nie stwierdzono dotąd obecności neuronów reagujących bezpośrednio na stymulację termoreceptorów skóry. Stwierdzono, że pozapodwzgórzowe struktury OUN, a w szczególności rdzeń kręgowy, współdziałają w regulacji temperatury ciała. Struktury termowrażliwe zlokalizowane są w pobliżu kanału rdzeniowego lub w obrębie jego ścian.
1.Typy układu nerwowego wg Pawłowa.
Pod względem anatomicznym układ autonomiczny dzielimy na:
układ współczulny (sympatyczny), czyli piersiowo - lędźwiowy
układ przywspółczulny (parasympatyczny), czyli czaszkowo - krzyżowy
Obwodowe włókna obu układów ulegają w swym przebiegu przerwie, ponieważ utworzone są przez wypustki osiowe dwóch kolejnych neuronów. Ciała nerwowe pierwszych znajdują się w ośrodkowym układzie nerwowym, a ich aksony po wyjściu na obwód kończą się w zwojach autonomicznych, tworząc synapsy z drugimi neuronami. Włókna pierwszych neuronów to włókna przedzwojowe, a włókna drugich to włókna zazwojowe.
Zwoje współczulne wchodzą w skład pnia współczulnego (zwoje przykręgowe) albo leżą oddzielni (zwoje przedkręgowe). Zwoje współczulne natomiast leżą w pobliżu unerwianego narządu lub w nim samym (zwoje śródścienne).
Ciała komóre współczulnych neuronów przedzwojowych leżą w słupach bocznych substancji szarej rdzenia kręgowego, tworząc jądro pośrednio-boczne (zakres Th - L ). Aksony tych neuronów pokryte otoczką mielinową wchodzą do pnia współczulnego i:
tworzą synapsy w jego zwojach lub
przechodzą bez przerwy przez te zwoje, tworząc synapsy dopiero w zwojach przedkręgowych
Włókna zazwojowe, bezmielinowe biegną do odpowiednich narządów. Jeden akson przedzwojowy tworzy synapsy przeciętnie z 15 neuronami zazwojowymi. Tworzą się duże rozgałęzienia, co zwiększa zakres docierania impulsów.
Część włókien dochodzi do mięśni gładkich przewodu pokarmowego nie kończy się bezpośrednio na nich, ale na przywspółczulnych komórkach zwojowych obecnych w ścianach żołądka i jelit. Dopiero aksony przywspółczulne docierają do komórek mięśniowych.
Komórki wydzielnicze rdzenia nadnerczy same stanowią zmodyfikowany zwój współczulny. Docierają do nich włókna współczulne przedzwojowe, a rolę zazwojowych pełnią wydzielane hormony (głównie adrenalina) i roznoszący je układ krążenia.
Układ współczulny zaopatruje w swe włókna mięśnie gładkie wszystkich narządów, mięsień sercowy oraz gruczoły wewnętrznego i zewnętrznego wydzielania. Włókna współczulne dla głowy odchodzą jako włókna zazwojowe od przedniego zwoju szyjnego i biegną w pniach nerwów czaszkowych (IX,X,XI,XII) oraz tworzą sploty wokół tętnicy szyjnej zewnętrznej i wewnętrznej.
Ciała komórek przywspółczulnych neuronów przedzwojowych leżą w pniu mózgu (śródmózgowie i rdzeń przedłużony) i w rdzeniu krzyżowym. Aksony biegną jako włókna przedzwojowe w pniach nerwów III,VII,IX,X i nerwów rdzeniowych wychodzących z segmentów krzyżowych (S - S). Część czaszkowa układu przywspółczulnego zaopatruje głowę, narządy klatki piersiowej i jamy brzusznej, część krzyżowa - narządy miednicy i narządy moczowo - płciowe. 1 akson przedzwojowy łączy się z ok.2 zazwojowymi, co ogranicza nieco zasięg działania.
Oprócz włókien odśrodkowych są włókna dośrodkowe. Nazywane są trzewnymi włóknami czuciowymi. Przekazują one impulsy od receptorów (chemo, mechano) rozsianych w narządach trzewnych do rdzenia kręgowego. Ciała tych komórek leżą w zwojach międzykręgowych, a duża część ich włókien biegnie w pniu nerwów błędnych.
1.Pierwszy i drugi układ sygnałów.
Pierwszy układ sygnałów stanowią tzw. mediatory. Chodzi tutaj głównie o hormony, które niosą sygnał i za pomocą specyficznych receptorów przekazują go do komórki. Hormony mogą wpływać na jeden lub wiele typów komórek. Każda jednak musi posiadać receptor o dużym powinowactwie do danego hormonu. Ilość, jak i powinowactwo receptorów w kom może ulec zmianie pod wpływem różnych czynników, np. żywienia lub pod wpływem samego hormonu działającego na zasadzie sprzężenia zwrotnego.
Przekazywanie sygnału do kom zachodzi za pośrednictwem swoistych receptorów, które mogą być zlokalizowane:
w jądrze kom lub w cytozolu
Wówczas hormon przenika przez błonę kom, łączy się z receptorem w cytozolu i wnika z nim do jądra, wiążąc się z receptorem na DNA. Powoduje wówczas transkrypcję genów. Takie receptory posiadają hormony steroidowe i hormony tarczycy.
w błonie kom
Hormon wiąże się z takim receptorem, zmieniając jego konformację. To powoduje aktywację cyklazy adenylanowej, która w obecności Mg2+ przekształca ATP w cAMP, który pełni rolę wtórnego przekaźnika (subst. pośredniczącej). On aktywuje kinazy białkowe, które katalizują reakcje fosforylacji z udziałem ATP. Fosforylacja większości enzymów prowadzi do ich aktywacji. Często ten sposób reakcji hormonu z receptorem prowadzi do uaktywnienia kaskady reakcji, które w bardzo krótkim czasie i przy bardzo małej ilości hormonu są w stanie spowodować rozległe i bardzo silne efekty. Przykłądem są reakcje rozpadu glikogenu.
Oprócz cAMP rolę wtórnego przekaźnika sygnałów pełni cGMP, aktywowany przez cyklazę guanylanową. Posobnie jak cAMP inicjuje procesy fosforylacji katalizowane przez fosforylazy i ,tak jak cAMP, cGMP rozkładany jest przez fosfodiesterazę.
Mniej ważnymi subst. pośredniczącymi są również trifosforan inozytolu (IP3) i diacyloglicerol (DAG). Ich powstawanie katalizuje fosfolipaza C. Wtórnym przekaźnikiem są także prostaglandyny aktywujące lub hamujące powstawanie cAMP.
Sam kompleks hormon-receptor nie jest jednak w stanie aktywować cyklaz czy fosfolipazy C. Pośredniczą w tym białko G i kalmodulina. Ta druga aktywowana jest przez jony Ca2+ i wtedy aktywuje cyklazy, rozpoczynając kaskadę fosforylacji. Sama jednak jej ulega, w wyniku czego do cytoplazmy i aktywuje fosfodiesterazę, która rozkłada cAMP do 5'AMP.
Białko G natomiast sąγ i zbudowane jest z 3 podjednostek, przy czym podjednostki są różne. Normalnie białkojednakowe dla wszystkich białek, a G jest związane z GDP. Powstały kompleks hormon+receptor (H+R) łączy się z kompleksem G+GDP, co powoduje zwiększenie powinowactwa białka G do GTP. Dochodzi więc do zamiany GDP z GTP. Kompleks H+R+G+GTP rozpada się szybko na H+R rozpada się szybko na +GTP. Ten ostatni jako forma aktywna aktywuje i GγH+R, G- cyklazę lub fosfolipazę C. Kiedy kaskada zostaje zapoczątkowana -GTP, a GTP zostaje przekształcone w GDP.enzym dysocjuje do G i staje się nieaktywne. Przykłady:γ-GDP łączy się z G-G str:153.
5.Aparat przykłębkowy i jego rola.
Aparat przykłębkowy znajduje się w obrębie bieguna naczyniowego. Wyróżnia się tu 3 bardzo ważne elementy morfologiczne:
komórki ziarniste (jg, mioepitelialne, przykłębuszkowe)- wystepują w obrębie błony środkowej tętniczki doprowadzającej. Pełnią funkcje wydzielnicze (wydzielają reninę).
komórki siatki wypełniające przestrzeń między tętniczką doprowadzającą a tętniczką odprowadzającą- tworzą mezangium zewnętrzne
komórki plamki gęstej- część komórek nabłonkowych kanalika II rzędu, stykająca się ściśle z biegunem naczyniowym ciałka nerkowego. Pełnią funkcję narządu osmoreceptorowego.
Aparat przykłębkowy odgrywa bardzo ważną rolę w regulacji czynności nefronu.
Jest on zbudowany tak że jedne jego komórki są detektorami stężenia chlorku sodu w przylegającej pętli kanalika krętego nefronu, zaś inne komórki w odpowiedzi na sygnały detektora działają w sposób dwojaki. Komórki kłębków nerkowych są głównym miejscem wytwarzania hormonu krwiotwórczego - erytropoetyny.
.Budowa i funkcje neuronów.
Neurony stanowią podstawowe jednostki strukturalne, czynnościowe i troficzne układu nerwowego. Wyspecjalizowane są one zarówno morfologicznie, jak i fizjologicznie do przekazywania i integracji otrzymywanej informacji. Komórki nerwowe mają zewnętrzną błonę zwaną neurolemą i prócz ciała (soma, perikarion) -dwa typy wypustek: jedną długą, dając odgałęzienia, zwaną aksonem (neurytem), i parę wypustek krótszych, drzewkowato rozgałęziających się w pobliżu ciała komórki, zwanych dendrytami.
Akson i jego odgałęzienia łączy komórkę nerwową~ z innymi komórkami poprzez stykanie się z nimi. Mogą to być inne komórki nerwowe albo komórki mięśniowe lub gruczołowe. Liczne aksony wiążą obwód ciała z ośrodkowym układem nerwowym. Na dendrytach i na błonie otaczającej ciało neuronu kończą się synapsami aksony innych neuronów, doprowadzające przewodzone informacje. Ciało neuronu integruje odebrane sygnały, przekazując je następnie aksonowi, który doprowadza je do innych komórek. Miejsce maksymalnego zbliżenia (styku) zakończenia aksonu z inną komórką nosi nazwę synapsy. Synapsa ma charakterystyczną budowę i przenosi informacje poprzez wydzielane na zakończeniu aksonu substancje chemiczne zwane mediatorami.
Synapsa, wydzielając inne substancje -n e u r o m o d u l a t o r y -może modulować natężenie przekazywanych informacji. Neuron otrzymuje impulsy od tysięcy innych neuronów i sam może tworzyć od 1000 do 10000 synaps na innych komórkach. Synapsy zajmują około 1/3 powierzchni ciała komórki nerwowej i dendrytów. Na pojedynczym neuronie ruchowym (motoneuronie) rdzenia kręgowego może znajdować się około 10000 kontaktów synaptycznych -około 2000 na ciele neuronu i 8000 na dendrytach.
Charakterystyczną właściwością neuronu -podobnie jak komórki mięśniowej -jest POBUDLIWOŚĆ, a podstawowymi jego funkcjami -generowanie i przewodzenie impulsów nerwowych.
Pobudliwością nazywamy zdolność reagowania Żywych komórek na działanie rożnych bodźców zmianą swej struktury lub wszczęciem, nasileniem albo osłabieniem charakterystycznej dla tych komórek funkcji. Wymienione reakcje są następstwem powstającego w komórkach procesu zwanego pobudzeniem (stanem czynnym). Jedną z charakterystycznych funkcji neuronu jest inicjacja impulsów nerwowych, które towarzyszą procesowi pobudzenia neuronu i związane są z odpowiednimi elektrochemicznymi zmianami w obrębie zewnętrznej błony, komórki nerwowej, powstającymi pod wpływem działania na nią bodźców, mających dostateczną silę, szybki czas narastania, działających przez czas konieczny do wywołania procesu pobudzenia. Bodźcami takimi w organizmie są najczęściej bodźce elektryczne i chemiczne. Inicjacja impulsów nerwowych i ich przewodzenie najlepiej dają się prześledzić w błonie aksonu (aksolemie).
Powstawanie hormonów
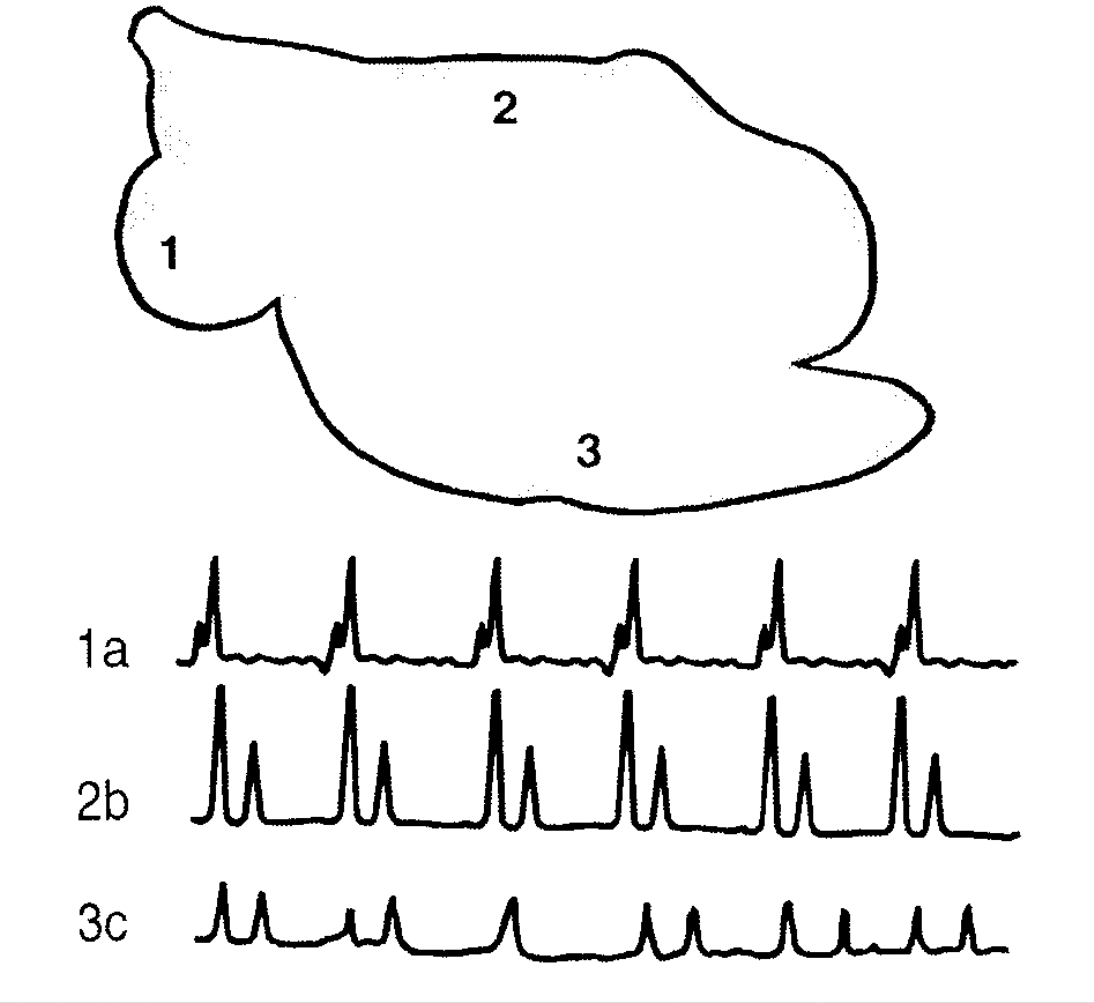
3.Cykle czepcowo-żwaczowy (omówienie z rys).
Cykl ten jest szeregiem następujących po sobie w odpowiedniej kolejności skurczów mięśniówki przedżołądków przeżuwaczy mający na celu odpowiednie wymieszanie i rozdzielenie trawionego pokarmu.
Zadaniem motoryki jest selekcja materiału nadającego Siudo dalszej fermentacji i usunięcie niesfermentowanych pozostałości. Zasadniczo rozróżnia się dwa rodzaje aktywności czepca i żwacza, skurcze mieszające czepca i żwacza oraz skurcze odbijania.
Skurcze mieszające rozpoczyna podwójny skurcz czepca. Pierwszy słabszy skurcz zmniejsza objętość czepca o około połowę, a drugi silniejszy polega na całkowitym skurczu mięśniówki czepca. Następnym etapem jest skurcz worka grzbietowego żwacza polegający na przesuwaniu doogonowo fali silnego skurczu perystaltycznego. Po jego zakończeniu pojawia się skurcz worka brzusznego żwacza, o podobnej sile i kierunku przebiegu, a następnie kolejny skurcz worka grzbietowego żwacza, a o przebiegu dogłowowym. Cykl skurczów mieszających zamyka skurcz worka brzusznego żwacza o przebiegu dogłowowym. Dzięki takiej cyklicznie powtarzającej się sekwencji skurczów treść przedżołądków ulega intensywnemu mieszaniu, ponadto dochodzi do oddzielania małych cząsteczek pokarmu od dużych. Skurcze odbijania następują zaraz po skurczach mieszających i polegają na pojawieniu się w tylnej części żwacza, w worku grzbietowym ślepym żwacza, intensywnej fali skurczowej rozchodzącej się na worek brzuszny żwacza i przebiegającej w kierunku dogłowowym.
Ruchy czepca i żwacza zachodzą cyklicznie (cykl czepcowo-żwaczowy). Zapoczątkowuje je czepiec, który kurczy się dwufazowo. W czasie drugiej fazy skurczu czepca, rozpoczyna się w żwaczu skurcz mieszający, a po nim może wystąpić skurcz odbijania -konieczny przy wydalaniu gazów powstających w żwaczu. Skurcze czepca występują tylko przed skurczami mieszającymi.
W żwaczu owcy można wyróżnić trzy typy ruchów:Typ dwójkowyutworzony jest z powtarzających się regularnie skurczów: mieszającego i odbijania. Występuje on wówczas, gdy przeżuwacz jest dobrze najedzony. Typ skurczów pojedynczychskłada się tylko ze skurczów mieszających. Typ ten występuje u zwierząt głodnych, ponieważ w tym czasie wytwarzanie gazów w żwaczu jest bardzo małe. Typ mieszany ruchów, w którym skurcz odbijania występują nieregularnie, pojawia się u zwierząt średnio najedzonych.
W regulacji motoryki przedżołądków bierze udzial układ nerwowy i czynniki humoralne. Draznienie receptorow w jamie ustnej, gardzieli przelyku powoduje wzrost częstości skurczow zoladka. Pobudzenie receptorow trawienca - hamowanie. Podraznienie włókien parasympatycznych -przyspiesza, sympatycznych -zwalnia. Ośrodki: rdzen przedłużony, układ siatkowaty i rabkowy, kora Mozgowa. Częstosc skurczow uzalezniona jest również od ilości tresci pokarmowej w przedżołądkach, składu, stopnia rozdrobnienia. Czynniki humoralne: przyspieszajace np. acetylocholina, histamina, serotonina, spadek poziomu cukru we krwi, wzrost LKT
Cykl czepcowo-żwaczowy. Schematyczne przedstawienie miejsca zapisu motoryki czepca (1) i żwacza (2, 3): 1 a -krzywa skurczów czepca, 2 b krzywa skurczów worka grzbietowego żwacza, 3 c krzywa skurczów worka brzusznego żwacza.
4.Hormony trzustki.
Trzustka poza funkcją trawienną wytwarza dwa hormony : insulinę i glukagon. Trzustka zbudowana jest z pęcherzyków wytwarzających sok trzustkowy i wsyp trzustki (Langerhansa) wydzielających hormony do krwi.
Wyspy trzustki zawierają trzy rodzaje komórek : alfa, beta i delta.
Komórki BETA - 60% całości leżą w środku każdej wyspy i wydzielają insulinę.
Komórki ALFA - 25% całości i wytwarzają glukagon.
Komórki DELTA - 10% całości, syntetyzują somatostatynę.
Oprócz tych komórek niektóre wyspy posiadają komórki PP w których odkryto polipeptyd trzustkowy. Bliskie położenie tych trzech rodzajów komórek jest niezmiernie ważne dla kontroli wydzielania hormonów. Przykładem może być hamowanie wydzielania glukagonu przez insulinę, oraz silne działanie somatostatyny jako inhibitora wydzielania insuliny i glukagonu.
Insulina, bierze udział w metabolizmie węglowodanów, tłuszczów i białek, odgrywa główną rolę w magazynowaniu nadmiernej ilości substancji energetycznych pobieranych z pokarmem.
Budowa insuliny: Jest małą cząsteczką białka o masie cząsteczkowej 5800, zbudowaną z dwóch łańcuchów aminokwasowych połączonych siarką. W przypadku rozerwania łańcuchów insulina traci swoje właściwości. Prekursorem insuliny wytworzonym w rybosomach jest preprohormon o masie cząsteczkowej około 11500, który w retikulum endoplazmatycznym przechodzi w lżejszą proinsulinę o masie cząsteczkowej 9000. Proinsulina jest biologicznie nieczynna i dopiero w aparacie Golgiego ulega przemianom w 85% do insuliny a w około 15% pozostaje nieczynna, okres półtrwania insuliny wynosi 6 minut, z wyjątkiem cząsteczek związanych z receptorami w tkankach docelowych, w ciągu 10-15 minut hormon ulega degradacji w wątrobie i nerkach pod wpływem enzymu insulinazy.
Działanie insuliny
Aby insulina zaczęła działać konieczne jest przyłączenie białka receptora błonowego o masie 300,000. Receptor insulinowy składa się z czterech oddzielnych podjednostek połączonych mostkami dwusiarczkowymi. Podjednostki ALFA leżą na zewnątrz błony, a beta wewnątrz cytoplazmy. Insulina przyłącza się do podjednostek ALFA powodując autofosforylację podjednostek BETA. Stają się one lokalnymi kinazami białkowymi pobudzającymi fosforylację wielu enzymów cytozolu. Końcowymi efektami działania insuliny na komórki są:
natychmiastowe zwiększenie przepuszczalności błony komórkowej dla glukozy (otwarcie kanału w białku błonowym o masie cząsteczkowej 55,000)
szybkie zwiększenie przepuszczalności błony komórkowej dla jonów potasowych, magnezowych i fosforowych.
Wolniejsze działanie insuliny zachodzące w ciągu następnych 10-15 minut polegające na zmianie aktywności wielu wewnątrz komórkowych enzymów.
Długotrwale zmiany (kilka dni) w tempie translacji mRNA w rybosomach i transkrypcji DNA w jądrze komórkowym.
Insulina zwiększa przepuszczalność błon komórek mięśniowych powodując niknięcie do ich wnętrza dużej ilości glukozy, gdzie może być zużyta w czasie wysiłku fizycznego lub odłożona w formie glikogenu. Jednym z najważniejszych działań insuliny po spożyciu pokarmu jest niemal całkowite zmagazynowanie glukozy w formie glikogenu wątrobowego.
Mechanizm tego procesu
Hamowanie fosforylazy wątrobowej powodującej glikogenolizę.
Przyśpieszenie wychwytu glukozy przez hepatocyty przy udziale glikokinazy.
Zwiększenie aktywności fosfofruktokinazy i syntetazy glikonegowej.
W wyniku działania insuliny na wątrobę, ilość glikogenu może wzrosnąć do 6% całkowitej masy wątroby. Spadek poziomu glukozy we krwi hamuje wydzielanie insuliny a pobudza wydzielanie glukagonu, co powoduje zahamowanie wychwytu glukozy i syntezy glikogenu w wątrobie. Glikogen ulega glikegenolizie i wolna glukoza wraca do krwi. Insulina wpływa na konwersje nadmiernej ilości glukozy do kwasów tłuszczowych które są odkładane w tkance tłuszczowej.
W odróżnieniu od innych tkanek komórki mózgu są przepuszczalne dla glukozy bez udziału insuliny, ale podstawowym warunkiem sprawnego funkcjonowania tkanki nerwowej jest odpowiedni poziom glukozy we krwi.
Insulina powoduje zwiększenie zapasów tłuszczu w tkance tłuszczowej. Zwiększa też transport glukozy do komórek wątroby i w przypadku zwiększenia do 6% koncentracji glikogenu glukoza przechodzi w formę acetylokoenzymuA, który jest substratem w syntezie kwasów tłuszczowych. Ważnych elementem udziału insuliny w przemianach tłuszczowych jest obniżenie aktywności lipolazy.
W ciągu kilku godzin po nakarmieniu zwierzęta insulina przyczynia się do magazynowania białek przez przyśpieszenie transportu aminokwasów do komórek. W Tym procesie działa podobnie do hormonu wzrostu. Insulina bezpośrednio wpływa na przyśpieszenie translacji mRNA a w dłuższym okresie zwiększa transkrypcje wybranych sekwencji DNA i w efekcie stymuluje syntezę białek. Dodatkowo hamuje katabolizm białek oraz glukoneogenezy. Insulina współdziała z somatotropiną w procesie wzrostu, brak jednego z tych hormonów jest przyczyna zahamowania wzrostu.
Regulacja wydzielania insuliny
Główną rolę pełni tu poziom glukozy we krwi, aminokwasy hormony przewodu pokarmowego i układ współczulny. Odpowiedź insuliny na wzrost poziomu glukozy we krwi jest dwufazowa, pierwszy okres występujący w ciągu 3-5 minut to dziesięciokrotny wzrost poziomu insuliny we krwi; jest on krótkotrwały i po 10 minutach obserwuje się spadek poziomu insuliny. Drugi etap działania insuliny rozpoczyna się po 15-20 minutach i trwa 2-3 godziny. Insulina uwalniana jest wtedy z zapasów w wyspach trzustki.
Mechanizm uwalniania insuliny działa na zasadzie sprzężenia zwrotnego - wysoki poziom glukozy stymuluje wydzielanie insuliny która szybko usuwa nadmiar glukozy, transportując ją do wątroby, mięsni i innych tkanek, i przestaje być wydzielana z trzustki.
Czynniki stymulujące wydzielanie insuliny:
hormony przewodu pokarmowego : gastryna, sekretyna, cholecystokinina, oraz żołądkowy peptyd hamujący GIP
Glukagon
Hormon wzrostu
Kortyzol
Progesterol i estrogeny
Stres
Chorobą powstającą w wyniku braki insuliny jest cukrzyca (diabetes melitus) a najczęstszą przyczyną niedostatecznego wydzielenia insuliny jest degeneracja komórek BETA trzustki. Główne efekty braku insuliny to zmniejszenie zużycia glukozy przez komórki i wzrost jej poziomu we krwi od trzech do dziesięciu razy powyżej poziomu (hiperglikemia), zwiększony metabolizm tłuszczów. W wyniku cukrzycy nadmiar cukrów osocza jest wydalany z moczem (w ciągu doby nawet 100g glukozy).
Symptomy braku insuliny to:
poliuria
pragnienie
nadmierny apetyt
spadek masy ciała
Glukagon
Jest syntetyzowany w komórkach ALFA trzustki, ma budowę polipeptydu (29 aminokwasów) o masie cząsteczkowej 3500. Jego działanie jest przeciwstawne do insuliny, powoduje wzrost poziomu glukozy we krwi, stymuluje glikogenolizę i glikogenogenezę.
W procesie glukoneogenezy glukagon aktywuje enzymy niezbędne w konwersji pirogonianu do fosfoenolopirogronianu oraz zwiększa ilość aminokwasów przeznaczonych do syntezy glukozy. Dodatkowo aktywuje lipazol. W dużych ilościach glukagon może wzmagać prace serca, sekrecje żółci i hamować wydzielanie soków trawiennych. Bodźcami wywołującymi glikoneogenezę są spadek poziomu glukozy we krwi, duża koncentracja alaniny i argininy i wysiłek fizyczny.
Somatostatyna
Wydzielana przez komórki DELTA, to 14-aminokwasowy polipeptyd o okresie półtrwania około 3 minuty we krwi. Wydzielanie somatostatyny stymulują czynniki związane z pobieraniem pokarmu:
wzrost poziomu glukozy
wzrost poziomu aminokwasów
wzrost poziomu kwasów tłuszczowych
wzrost poziomu hormonów przewodu pokarmowego
Somatostatyna hamuje wydzielanie zarówno insuliny jak i glukagonu. Zmniejsza skurcze żołądka, dwunastnicy, pęcherzyka żółciowego oraz obniża wydzielanie i absorpcję w przewodzie pokarmowym. Wynika z tego ze głównym celem somatostatyny jest zmniejszenie metabolizmu składników pokarmowych czyniąc je dłużej dostępnymi.
3.Mechanizm oddychania u ptaków.
WDECH
W czasie wdechu powietrze przechodzi w następujący sposób : jama nosowa krtań górna tchawica ( w jej rozwidleniu narząd głosu - krtań dolna ) 2 oskrzela główne oskrzeliki przedsionka płuca ( wychodzą stąd : 6-8 oskrzeli dogrzbietowych ( te łączą się poprzez oskrzela zewn. z tylnymi workami powietrznymi - 2 piersiowe tylne i 2 brzuszne ) oraz 4-6 oskrzeli brzusznych ( komunikują się z workami powietrznymi przednimi ( 2 piersiowe przednie, 2 szyjne, 1 międzyobojczykowy ; od ww oskrzeli odchodzą oskrzeliki oddechowe - w nich następuje wymiana gazowa z krwią - u ptaków brak pęcherzyków płucnych ) oskrzela dogrzbietowe tylne worki powietrzne
Pewna część powietrza przechodzi do worków tylnych przez bezpośrednie połączenia i nie podlega wymianie gazowej dlatego w workach przednich jest niższe ciśnienie cząsteczkowe tlenu a wyższe CO2 niż w workach tylnych.
WYDECH
Z worków przednich ( powietrze z poprzedniego wdechu ) tchawica na zewnątrz
W tym czasie także :
Z worków tylnych ( bogate w tlen ) oskrzela zwrotne płuco ( tu : wymiana gazowa )
W czasie następnego wdechu zużyte powietrze z płuc worki przednie ( tu przebywa do wydechu ) na zewn. z powietrzem z płuc
W czasie wdechu zwiększona pojemność klatki piersiowej i jamy brzusznej pozwala na wciągnięcie powietrza do płuc i worków tylnych.Mostek ( duży u ptaków ) oddala się od kręgosłupa przednio-dolnie, żebra prostują się, mm.międzyżebrowe zewn. odgrywają tu istotną rolę, przepony brak.
W czasie wydechu żebra powracają do pozycji wyjściowych dzięki mm. międzyżebrowym wewn.i brzusznym.
W czasie lotu kl. piersiowa jest nieruchoma - ruch skrzydeł powoduje wciskanie i wyciskanie powietrza z i do płuc poprzez wpływ na zachyłki worka międzyobojczykowego.
Inne zadania worków powietrznych :
Rola w termoregulacji
Pneumatyzacja kości
Pośrednio uczestniczą ( przez ucisk ) na wydalanie kału i jaja
Ułatwiają nurkowanie ptakom wodnym
4.Omów regulację procesów rozrodczych.
Proces kompleksowy z uczesnictwem wielu czynników. Biorą w niej udzial uklady hormonalny nerwowy oraz czynniki środowiskowe. W przedniej części przysadki produkowane są hormony takie jak: LH FSH PRL. Nadrzędny wobec nich jest GnRH
Podwzgórze wydziela swoje neurohormony w sposób pulsacyjny
2.Regulacja czynności nerek.
1. Autoregulacja - polega ona na tym że ukrwienie nerek nie ulega większym zmianom przy szerokich wahaniach ciśnienia tętniczego krwi. Zdolność do autoregulacji dotyczy tylko kłębków korowych i nie zależy od czynników pozanerkowych. Autoregulacja jest możliwa dzięki postępującemu skurczowi mięśniówki tętniczek doprowadzających, w miarę zwiększania ciśnienia tętniczego oraz relaksacji naczyń doprowadzających, w miarę obniżania ciśnienia krwi.
2. Regulacja nerwowa czynności nerek - wpływ układu nerwowego na czynność nerek może być bezpośredni lub pośredni. Decydujący wpływ wywiera układ nerwowy autonomiczny, zwłaszcza część współczulna. Pobudzenie układu współczulnego powoduje znaczne obkurczenie naczyń nerkowych (tętniczych), co prowadzi do obniżenia przepływu krwi oraz spadku filtracji kłębkowej osocza.
Nerwowa (odruchowa) regulacja czynności nerek wyzwalana jest przez receptory z różnych układów, głównie z baroreceptorów sercowo-płucnych i naczyniowych, z mechano i chemoreceptorów zlokalizowanych w nerkach, a także z receptorów trzewnych (w jelitach, mięśniach gładkich, układzie wrotnym wątroby).
3. Regulacja hormonalna czynności nerek - funkcja nerek zostaje pod kontrola hormonów, które można podzielić tu na 2 grupy: o działaniu antynatriuretycznym i antidiuretycznym oraz natriuretycznym i diuretycznym.
Hormonem antydiuretycznym jest wazopresyna.
4. Układ renina-angiotensyna-aldosteron.
5. Przedsionkowy peptyd natriuretyczny.
6. Mózgowy peptyd natriuretyczny.
7. Korowy peptyd natriuretyczny.
8. Urodylatyna.
9. Adrenomedulina.
10. Glikokortykoidy.
11. Hormony tarczycy i gruczołów płciowych.
3.Krwinki płytkowe.
Najmniejsze różnokształtne krwinki bezjądrzaste
Dostrzegalne w rozmazie jako skupiska małych romboidalnych komórek bez barwnika w cytoplazmie
Jest ich 30-100 razy więcej niż krwinek białych
Są to małe fragmenty cytoplazmy oderwane od olbrzymich komórek szpiku - megakariocytów
Biorą udział w procesie krzepnięcia krwi
Zawierają one dużą ilość substancji obkurczającej naczynia krwionośne - serotoniny
Gromadzą się w miejscach uszkodzenia naczyń, gdzie przylegają do uszkodzonej ściany naczyniowej i uwalniają serotoninę, która powoduje miejscowy skurcz ściany naczyniowej, zmieniając krwawienie
Są szeroko stosowane w leczeniu nowotworów
4.Trawienie białek w jelitach.
Trawienie białek zaczyna się dopiero w żołądku, gdzie komórki główne komórek gruczołowych żołądka wydzielają nieczynny enzym pepsynogen. Komórki okładzinowe wydzielają kwas solny, w obecności którego pepsynogen przekształca się w postać czynną - pepsynę. Pepsynogen może być także aktywowany przez aktywną pepsynę.
Trawienie białek jest kontynuowane w świetle jelita dzięki enzymom soku trzustkowego. Nieaktywne enzymy soku trzustkowego ulegają aktywacji na zasadzie reakcji przebiegającej kaskadowo. Czynnikiem aktywującym trypsynogen do trypsyny jest enterokinaza wytwarzana przez enterocyty. Aktywna trypsyna działa autokatalitycznie oraz prowadzi do aktywacji wszystkich pozostałych proenzymów soku trzustkowego, proteaz, nukleaz i enzymów lipolitycznych. Enzymy trzustki ulegają uszkodzeniom w świetle przewodu pokarmowego, tylko niewielka ich część ulega wydaleniu z kałem.
Trawienie przyścienne białek (oraz polisacharydów) w jelicie cienkim opiera się, podobnie jak w świetle jelita, na hydrolizie enzymatycznej z tą różnicą, że enzymy rąbka szczoteczkowego są przytwierdzone wiązaniami chemicznymi do powierzchni błony komórkowej enterocytu. Enzymy trawienia przyściennego są wytwarzane w enterocytach i transportowane na zewnętrzną powierzchnię błony komórkowej wierzchołkowej części komórki, gdzie ulegają zakotwiczeniu fragmentem łańcucha. Takie umocowanie enzymu ogranicza pole jego działania do strefy dyfuzyjnej, leżącej bezpośrednio nad komórką. Powstałe w wyniku pierwszego etapu trawienia peptydy w celu dalszego rozpadu muszą się przemieścić do strefy dyfuzyjnej. Peptydazy obecne w strefie dyfuzyjnej są przytwierdzone do błony komórkowej enterocytu. Zadaniem ich jest hydroliza peptydów powstałych w wyniku trawienia w świetle jelita. Efektem trawienia przyściennego jest powstanie wolnych aminokwasów oraz dipeptydów i tripeptydów, które mogą ulec wchłonięciu. Zatem końcowe trawienie peptydów odbywa się dwuetapowo - w rąbku szczoteczkowym i wewnątrz enterocytu.
5.Motywy, popędy i emocje.
Ukierunkowane akty zachowania się spowodowane są ( oprócz zachowań wrodzonych ) określonymi potrzebami biologicznymi, które wywołują zawsze pewne zachwianie wewn. Równowagi organizmu. Na tym podłożu powstają motywy - jest to wewn. stan organizmu uruchamiający i organizujący zachowanie, zmierzające do zaspokojenia określonej potrzeby biol.
Motywy :
homeostatyczne - działające na rzecz przywrócenia homeostazy organizmu
płciowe
macierzyńskie
poznawcze
socjalne
uczuciowe itp
Zachowania motywacyjne mogą trwać wiele godzin a nawet dni.
Składają się z 2 komponentów : popędów ( wewn. mechanizmy mobilizujące osobnika do działania ) oraz chęci osiągnięcia określonego celu lub zdobycie wystarczającej nagrody. Zaspokojenie popędu powoduje jego wygaszenie lub zahamowanie.
Popędy :
- apetytywne ( dodatnie, zachowawcze ) - ukierunkowujące działalność organizmu na osiągnięcie kontaktu z bodźcami atrakcyjnymi ( pokarm, woda, partner płciowy, potomstwo )
- awersyjne ( ujemne, obronne) - ukierunkowują działalność organizmu na obronę własnego ciała przed działaniem szkodliwych czynników ( strach ucieczka, wściekłość agresja itd. )
W związku z działaniem motywów i popędów zwierzęta doznają przeżyć o przyjemnych lub przykrych dla nich cechach, te z nich które towarzyszą popędom nazywane są uczuciami ( np. głodu, pragnienia, lęku, gniewu, bólu, satysfakcji płciowej ). Uczucia o szczególnie silnym nasileniu ( gdy popęd jest dostatecznie silny ) to emocje. Są to psychiczne napięcia związane z silnymi przeżyciami, wyróżniają się reakcjami, podczas których nagromadzone w organizmie zapasy energetyczne ulegają gwałtownemu wyzwoleniu ( występują one na tle odp. nasilonych potrzeb np. pokarmowych ).
Bodźce oraz wywołane przez nie emocje można podzielić na :
dodatnie - radość , zadowolenie itd.
ujemne - gniew, smutek ,obawa, przygnębienie, niezadowolenie
Emocje towarzyszą dążeniom do osiągnięcia określonego celu i zaspokojenia np popędu pokarmowego, płciowego, samozachowawczego zwłaszcza gdy zaspokojenie to natrafia a przeszkodę ( emocje dynamizując organizm dodają mu niejako siły do pokonania trudności ). Towarzyszy im pobudzenie somatycznego i autonomicznego ukł. nerw. oraz ukł. wewn. wydzielania ( gł. adrenalina - hormon lęku i noradrenalina - hormon gniewu ), oraz zmiany cielesne tj odp. reakcje ruchowe, postawne, głosowe i ekspresje mimiczne - np. strachowi towarzyszy - przyspieszenie akcji serca, oddychania, zwężenie naczyń skórnych ,zimne poty, stroszenie włosów, rozszerzenie źrenicy, suchość w jamie ustnej, drżenie mięśni ( jest to tzw stan emocjonalny ). Rodzaj emocji u zwierząt można poznać po określonych zachowaniach - np. char. ustawienie uszu czy ogona , stroszeniu sierści, tupaniu, szczerzeniu zębów, wydawaniu określ. odgłosów ( warczenie, mruczenie itd. ).
1.Termogeneza bezdrżeniowa i drżeniowa.
TERMOGENEZA- to zespół zjawisk ( przemian ) w organizmie których celem jest,
produkcja ciepła tak aby przywrócić lub utrzymać zrównoważony bilans cieplny organizmu.
ZRÓWNOWAŻONY BILANS CIEPLNY- to zasada na której opiera się utrzymanie niezmiennej temperatury wewnętrznej u zwierząt stałocieplnych. Temperatura ta jest stała gdy ciepło wytwarzane w drodze przemian metabolicznych równoważy ilość ciepła oddanego do otoczenia.
Organizm zwierzęcy narażony na stres zimna może wyprodukować ciepło na dwa podstawowe sposoby, przez:
termogenezę drżeniową- opartą na mechanizmie kurczenia się mięśni szkieletowych, niezależnie od woli zwierzęcia
termogenezę bezdrżeniową- opartą na uzyskiwaniu ciepła ze wzrostu przemian metabolicznych
Te dwa sposoby produkcji ciepła najczęściej działają jednocześnie wzajemnie się uzupełniając.
TERMOGENEZA BEZDRŻENIOWA
(metaboliczna)
Ten rodzaj termogenezy jest efektem reakcji chemicznych zachodzących głównie w narządach trzewnych, np.: wątrobie, nerkach, sercu, przewodzie pokarmowym; ale także w mięśniach czy układzie nerwowym, a u noworodków i zwierząt które ulegają hibernacji w tkance tłuszczowej brunatnej. Sam proces termogenezy przebiega w mitochondriach komórek wymienionych narządów i tkanek. Polega on na ustalaniu się gradientu protonowego w poprzek błony mitochondrium i następującym po nim wydostawaniu się protonów na zewnątrz mitochondrium, lecz bez wytworzenia ATP, gdyż nie przechodzą one przez kompleks ATP-zy tylko przez białko kanałowe zwane termogeniną.
Proces ten regulowany jest na drodze neurohumoralnej, w której główną rolę spełniają hormony rdzenia nadnerczy( adrenalina i noradrenalina), a także tarczycy( tyroksyna i trijodotyronina).
Inicjacja tak jak i utrzymanie tego procesu odbywa się za pomocą układu nerwowego.
Ośrodki odpowiedzialne za regulację temperatury znajdują się w podwzgórzu.
Na wydzielanie tarczycy wpływ mają ośrodki temoregulacyjne przedniej części podwzgórza powodujące wydzielanie TRH który pobudza przednią część przyssadki do wydzielenia TSH, który to hormon wywołuje wzrost wydzielania hormonów tarczycy T3 i T4.
Na wydzielanie hormonów rdzenia nadnerczy ADRENALINA i NORADRENALINA ma wpływ układ autonomiczny współczulny powodujący wzrost ich stężenia, a także wydzielanie noradrenaliny na zakończeniach nerwowych(adrenergicznych) tego układu.
Za pomocą tych hormonów następuje wzmożony metabolizm w tkankach docelowych czego skutkiem jest wydzielanie się ciepła.
TERMOGENEZA DRŻENIOWA
(mięśniowa)
Ten rodzaj termogenezy polega na synchronicznych skurczach mięśni zginaczy i prostowników w obrębie mięśni głowy(mięśnie żwaczowe), kończyn, tułowia.
Mimo iż przy słabej intensywności drżenia jest ono możliwe do zahamowania siłą woli to w istocie jest to proces mimowolny niezależny od kory mózgowej.
Drżenie mięśni jest wywołane drażnieniem niską temperaturą na receptory obwodowe a także rdzeń kręgowy czy podwzgórze które wyzwalają oprócz drżenia tonus mięśniowy(również uznany za jedną z reakcji termoregulacyjnych).
Istnieje założenia że ośrodek drżenia mięśniowego znajduje się w podwzgórzu prawdopodobnie to ten sam który jest ośrodkiem termoregulacyjnym zimna. Wpływy z tego ośrodka przenoszone są drogami zstępującymi rdzenia kręgowego do motoneuronów alfa, a ich aksonami do mięśni szkieletowych. Rytmiczność ich wyładowań jest uwarunkowana czynnością samych motoneuronów jest ona pod wpływem impulsacji z ośrodka drżenia mięśniowego oraz obwodowej recepcji termicznej. Dochodzi tu także wpływ impulsacji z receptorów ścięgnowych i mięśniowych.
2.Specyfika krążenia mózgowego.
Mózg charakteryzuje się ogromnym zużyciem tlenu. Nie może być tam zaciągany dług tlenowy. Mózg nie może działać przy ograniczonym dopływie tlenu. Kilkusekundowa przerwa w dostarczaniu tlenu powoduje zauważalne zaburzenia w pracy mózgu. Ustanie krążenia powoduje utratę przytomności po kilku sekundach i nieodwracalne zmiany w neuronach po kilku minutach.
Największe zużycie tlenu i najgęstsza sieć naczyń włosowatych występuje w częściach mózgu gdzie jest przewaga perykarionów czyli w istocie szarej. Przepływ krwi wynosi tu 100ml/min/100g tkanki. Jest to przepływ większy niż w mięśniu sercowym. W obszarach mózgu gdzie przeważa istota biała- dendryty, neuryty przepływ krwi jest pięciokrotnie mniejszy. Przez mózg przepływa około 10-15% całej krwi krążącej. Głównym mechanizmem regulacji przepływu krwi jest autoregulacja.
Autoregulacja jest modyfikowana przez miejscowo uwalniane czynniki metaboliczne, głównie jony wodorowe. Rozszerzają one naczynia, umożliwiając usuwanie CO2 . Regulacja ta oparta jest na sprzężeniu zwrotnym, gdyż usunięcie CO2 ze środowiska obniża koncentrację jonów wodorowych.
Specyfiką krążenia mózgowego jest obecność bariery krew - mózg Jest ona uwarunkowana budową morfologiczną oraz procesami chemicznymi.
Podstawową morfologiczną strukturą bariery krew - mózg jest odmienna budowa śródbłonka naczyń włosowatych. Komórki śródbłonka łączą się ze sobą poprzez ścisłe złączenia(strefy zamykające) bądź też przez zanik oddzielających je błon komórkowych. Naczynia włosowate mózgu uczestniczące w barierze nie posiadają porów, szczelin ani okienek przez które mogłoby się odbywać przenikanie cząsteczek z osocza i do osocza. Komórki śródbłonka wykazują dużą aktywność metaboliczną, zawierają liczne mitochondria, ale nie występuje w nich proces pinocytozy. Jedyną drogą przenikania i wymiany jest transport przezkomórkowy (transcelularny) związany z koniecznością pokonania obu błon komórkowych i cytoplazmy. Transport przezkomórkowy oparty jest na dwóch występujących tu systemach: dyfuzji przez błony komórkowe i czynnego transportu z udziałem nośników białkowych. Z pierwszego systemu korzystają głównie drobnocząsteczkowe substancje odżywcze i inne rozpuszczalne w tłuszczach. Z drugiego systemu korzystają przede wszystkim peptydy i białka o małej masie, uczestniczące w procesach regulacyjnych (oksytocyna, wasopresyna, opiaty, substancja P, LH-RH, insulina, glukagon, somatostatyna i inne). Wydobywające się z naczyń włosowatych substancje które przenikają przez ścianę kapilaru, nie osiągają bezpośrednio neuronów. To komórki glejowe - astrocyty są łącznikiem między ścianką naczynia włosowatego a neuronami. Jedną z wypustek wykształconą w postaci stopki dotykają do powierzchni naczynia włosowatego, innymi wypustkami nawiązują kontakt z sąsiednimi neuronami. Transport substancji przez astrocyty odbywa się w obu kierunkach: z naczynia do neuronu i z neuronu do naczynia.
Bariera biochemiczna jest drugim etapem bariery krew - mózg. Polega ona na możliwości metabolizowania pewnych związków, które przenikają do cytoplazmy komórek śródbłonkowych lub cytoplazmy astrocytów. Np. endogenne katecholaminy(adrenalina, dopamina, serotonina) są inaktywne w cytoplaźmie przez enzym - monoaminooksydazę. Chroni to neurony przed chwilowymi wzrostami koncentracji katecholamin w osoczu, jakie pojawiają się w wyniku pobudzenia układu współczulnego, strachu, stresu. Wolne kwasy tłuszczowe ulegają szybkiej estryfikacji.
Drugą specyfiką krążenia mózgowego jest brak naczyń układu chłonnego, a obecność płynu mózgowo rdzeniowego. Płyn mózgowo - rdzeniowy wypełnia komory mózgowe, a więc jamy wewnętrzne mózgowia oraz przestrzenie podpajęczynówkowe znajdujące się między pajęczynówką a oponą miękką właściwą. Powstaje on w obszarze splotów naczyniówkowych znajdujących się w ścianie komór i w bezpośrednim ich sąsiedztwie, w tzw. narządach okołokomorowych. Do nich należą: wyniosłość pośrodkowa, narząd naczyniowy blaszki krańcowej, szyszynka i pole najdalsze. Obszar ten jest pozbawiony bariery krew - mózg.
Występuje tu śródbłonek naczyniowy typu okienkowego. Do okienek śródbłonka przylegają wypustki komórek glejowych - tanycytów, które odbierają z krwi do swojej cytoplazmy różne substancje, a następnie przekazują je do płynu mózgowo-rdzeniowego, z którym kontaktują się inną wypustką. Kierunek przekazu może być odwrotny, tanycyty odbierają substancje z płynu mózgowo-rdzeniowego i przekazują je poprzez swoją cytoplazmę do naczyń krwionośnych. Płyn mózgowo-rdzeniowy nie jest przesączem krwi, lecz wydzieliną splotu naczyniówkowego i przylegających tu tanycytów. Zawiera on poza wodą małe ilości białka i odmienny niż krew skład jonowy.
Komory boczne połączone są przez parzysty międzykomorowy otwór z komorą trzecią, ta przez wodociąg mózgu z komorą czwartej. Komora czwarta ma otwory skierowane do przestrzeni podpajęczynówkowej położonej wokół rdzenia przedłużonego i zwanej zbiornikiem wielkim. Zbiornik wielki łączy się z podobnymi przestrzeniami podpajęczynówkowymi wokół mózgu i rdzenia kręgowego. Do przestrzeni podpajęczynówkowej wpuklają się w wielu miejscach wypustki(kosmki pajęczynówki), które wnikają również do zatoki żylnej biorą udział w resorpcji płynu do krwi. Znaczna część płynu wchłaniana jest do naczyń żylnych. Zmienna objętość płynu mózgowo-rdzeniowego uczestniczy w tworzeniu ciśnienia śródczaszkowego. Ciśnienie śródczaszkowe wywiera zasadniczy wpływ na przepływ krwi przez mózg.
3.Rola nerek w regulacji kwasowo-zasadowej.
Rola nerek w regulacji równowagi kwasowo-zasadowej jest związana z ich zdolnością do kontrolowanego, zależnego od aktualnego stanu organizmu, wydalania kwasów i zasad.
W procesach metabolicznych powstaje nadmiar jonu wodorowego (kwasów) w stosunku do zasad, dlatego nadmiar kwasów jest systematycznie usuwany z organizmu. Mocz dobowy zawiera cały nadmiar jonu wodorowego.
Jon wodorowy pochodzi w ustroju z:
przemian fosfolipidów i fosfonukleotydów do kwasu fosforowego
przemian aminokwasów zawierających siarkę do kwasu siarkowego
niepełnych przemian związków cukrowych (do kwasów mlekowego, pirogronowego, oksymasłowego, acetooctowego).
Powstałe kwasy przechodzą do płynów ustrojowych i reagują z buforami tych płynów, dzięki czemu zmiany pH są minimalne. Najważniejszym układem buforowym płynów zewnątrzkomórkowych jest układ wodorowęglanów. Kwas, który przejdzie do płynu zewnątrzkomórkowego, reaguje głównie z wodorowęglanami, tworząc kwas węglowy oraz sól danego kwasu. Kwas węglowy jest źródłem CO2 który jest wydalany przez płuca, a powstała sól danego kwas ego może zostać wydalona z moczem. W wyniku tych reakcji zmniejsza się zawartość wodorowęglanów w organizmie, co powoduje obniżenie jego rezerw buforowych i może prowadzić do zakwaszenia ustroju.
W przypadku gromadzenia się w ustroju nadmiaru zasad, nerka jest narządem, przez który ten nadmiar jest usuwany.
Proces wydalania jonów wodorowych i zasad jest regulowany przez ten sam mechanizm, oparty na 3 ściśle związanych ze sobą zjawiskach:
resorpcji wodorowęglanów
wydalaniu w różnych proporcjach fosforanów jedno- i dwuzasadowych
wytwarzaniu i wydalaniu amoniaku
Resorpcja wodorowęglanów bezpośrednio nie prowadzi do eliminacji H+ z organizmu, jednak przyczynia się do oszczędzania zasad.
Wydalanie jonów wodorowych w postaci fosforanów odbywa się w kanalikach bliższych, dalszych i zbiorczych.
Wytwarzanie i wydalanie amoniaku jest drugą drogą usuwania jonów wodorowych przez nerki.
4.Budowa i zadania krwinek czerwonych.
Erytrocyt - jeden z podstawowych morfotycznych składników krwi. Głównym zadaniem erytrocytów jest przenoszenie tlenu i dwutlenku węgla, co jest możliwe dzięki obecności w nim czerwonego barwnika hemoglobiny, który ma zdolność do nietrwałego wiązania tlenu i przechodzenia w oksyhemoglobinę.
Prawidłowy erytrocyt ssaczy jest okrągłą, dwuwklęsłą w środku komórką o średnicy 6-9 μm - wyjątkiem są owalne erytrocyty daniela, wielbłądów i lam (j.w.). Prawidłowy erytrocyt nazywa się normocytem, krwinki większe od prawidłowych to makrocyty, natomiast mniejsze to mikrocyty.
Krwinki czerwone nie dzielą się. Nie mogą pełnić normalnych funkcji komórkowych, nie mają też mechanizmu, który mógłby naprawiać powstające w nich z czasem uszkodzenia i po kilku miesiącach użytecznego życia (ok. 120 dni) ulegają zniszczeniu głównie w śledzionie rzadziej w wątrobie. Organizm musi zatem nieustannie produkować nowe erytrocyty, które stopniowo zastępują te, które uległy rozpadowi.
Wszystkie komórki krwi są wytwarzane w szpiku kostnym (u dzieci do lat 3 wszystkie komórki są wytwarzane przez śledzionę natomiast erytrocyty zaczynają powstawać w szpiku kostnym dopiero od 4 roku życia), gąbczastej tkance znajdującej się wewnątrz kości w procesie erytropoezy z komórek macierzystych erytrocytów, tzw. erytroblastów z prędkością 120 000 000[2] na minutę.
25. Oddychanie plucne
Przystosowania morfologiczne
Wymiana gazów między powietrzem pęcherzykowym w krwią odbywa się na drodze dyfuzji.
Dyfuzja gazów w płucach zależy od:
różnic ciśnień w obu układach
powierzchni czynnej pęcherzyków i wielkości drogi dyfundujących gazów
grubości błon oddzielającej światło pęcherzyka od hemoglobiny w erytrocycie
Maksymalne zbliżenie krwi do powietrza pęcherzykowego osiągane jest przez szczególną budowę pęcherzyków płucnych i oplatających je naczyń włosowatych.
Budowa pęcherzyka - wysłany 3 typami komórek oddechowych (pneumocytów):
Pneumocyty I rzędu
Ubogi metabolizm własny, dzięki czemu zużywają minimalne ilości dyfundującego przez nie tlenu.
Pneumocyty II rzędu
Zdolne do sekrecji surfaktantu (czynnika powierzchniowego), który pokrywa cienką warstwą wszystkie pneumocyty.
Funkcje surfaktantu:
a) zmniejsza opór sprężysty płuc (dzięki czemu pęcherzyki ulegają łatwo rozciągnięciu w czasie spadku ciśnienia w jamie opłucnej (wdech) i nigdy nie dochodzi do ich zapadania się)
b) zabezpiecza przed infekcją bakteryjną, przed przesiąkaniem płynu międzykomórkowego do pęcherzyka.
c) reguluje poziom wilgotności powietrza oddechowego
Sekrecja pnemocytów uzależniona jest od nerwu błędnego
* pełna czynność nerwu błędnego przypada w końcowych tygodniach życia
płodowego - u „wcześniaka” na skutek braku surfaktantu w czasie wdechu nie
dochodzi do pełnego rozciągania i wypełnienia powietrzem pęcherzyków płucnych
Pneumocyty III rzędu - komórki szczoteczkowe - rola komórek receptorowych
Zaopatrzone w liczne mikrokosmki. Występują pojedynczo i nie we wszystkich pęcherzykach.
Leżą przy zakończeniach czuciowych nerwu błędnego-
ich mechaniczne podrażnienie niedostatecznym wypełnieniem przez powietrze pobudza te zakończenia. Efekt odruchu - silny wdech w postaci westchnienia.
Wszystkie pneumocyty leżą na błonie podstawnej, która styka się z błoną podstawną śródbłonka naczyń włosowatych.
Śródbłonek ten jest zwarty, co przeciwdziała filtracji i zbieraniu się płynu
międzykomórkowego (który byłby przeszkodą dla dyfundujących gazów)
Dyfuzja i wiązanie O2 z hemoglobiną
Odtlenowana krew dopływająca do naczyń włos. pęcherzyków łączy się gwałtownie -
wobec dużej różnicy ciśnień parcjalnych - z dyfundującym z pęcherzyków powietrzem.
W krwince powstaje oksyhemoglobina, w czasie odszczepienie wodoru.
Powoduje to pobranie z nagromadzonego w krwince chlorku potasu i przyłączenie do oksyhemoglobiny jonu potasowego.
Powstanie oksyhemoglobiny zależy od:
prężności O2 i CO2 w danym środowisku
stężenia jonów H+
zawartości 2,3-dwufosfoglicerynianu (2,3-DPG)
temperatury (jej spadek ułatwia wiązanie tlenu)
Dyfuzja CO2 z krwi do powietrza pęcherzykowego
Cały obecny CO2 we krwi żylnej przenoszony jest w 70% przez osocze i w 30% przez krwinki (najwięcej w postaci wodorowęglanów, uzupełniająco - połączenia karbaminowe)
Przyłączenie tlenu do hemoglobiny powoduje natychmiastowe przyłączenie do oksyhemoglobiny jonów potasowych. Pociąga to za sobą lawinowo przebiegające procesy, które (na krótkim odcinku naczynia włosowatego i w ułamku sekundy) powodują:
przejęcie przez krwinkę rozpuszczonych w osoczu wodorowęglanów
uwolnienie do osocza, a następnie do powietrza pęcherzykowego całej zawartości obecnego we krwi CO2
znaczne stężenie jonów Cl-
Jony te wędrują do osocza, a z osocza (dla zrównoważenia) przenikają HCO3-,
które zmieniają się na kwas węglowy (H2CO3), który pod wpływem anhydrazy
węglowej rozpada się na wodę i dwutlenek węgla.
Dwutlenek węgla przenika do osocza, a stamtąd do powietrza pęcherzykowego.
4.Zmęczenie mięsni i stężenie pośmiertne.
Zmęczenie mięśni
To stan przejściowego obniżenia zdolności do pracy, będący następstwem dłuższej aktywności ruchowej, a ustępujący po okresie wypoczynku.
Powody zmęczenia to niezdolność elementów kurczliwych do skurczu i zmiany w procesach metabolicznych leżących u podstawy procesu skurczowego. Są one następstwem wyczerpania się zapasów energetycznych w samym mięśniu, a także gromadzeniem w mięśniu produktów metabolizmu (np. kw. fosforowego, kw. mlekowego itp.), które oprócz upośledzania pracy poszczególnych włókien mięśniowych pogarszają zdolność błon pobudliwych do wytwarzania potencjałów.
Zmęczenie następuje znacznie szybciej, jeśli odcięty zostanie dopływ krwi lub tlenu do mięśni.
W izolowanym mięsniu, drażnionym podnietami powtarzającymi się w krótkich odstępach czasu narastające zmęczenie przejawia się:
przedłużeniem czasu utajonego pobudzenia i okresu refrakcji bezwzględnej
dłuższym czasem skurczu i rozkurczu
zmniejszoną amplitudą skurczów
brakiem powrotu do pierwotnej długości i skłonnościami do występowania przykurczów
zmianą sprężystości
zmniejszeniem siły bezwzględnej
Szybciej jednak od aparatu kurczliwego męcza się synapsy. Powoduje to pogorszenie przekazywania stanu pobudzenia na mięsień. Jest to efekt wyczerpania na płytkach ruchowych zasobów acetylocholiny. Chroni to jednak aparat kurczliwy przed całkowitym wyczerpaniem się zapasów.
Wcześniej jeszcze od synaps męczą się ośrodki nerwowe w rdzeniu kręgowym, jądrach podkorowych i korze mózgowej, które kontroluja pracę mięśni całego organizmu.
Przy dłuższym zmęczeniu mięśnia może w nim dojść do utrzymującego się przez kilka minut przykurczu, czyli tzw. kontraktury fizjologicznej. Mięsień nie reaguje wtedy na żadne bodżce i nie można w jego obrębie stwierdzić potencjałów czynnościowych. Jest to efekt wyczerpania wszystkich zsobów energet. (ATP, fosfokreatyny) tak, że brakuje wiązań wysokoenerget. Do odtworzenia ATP z ADP. Dopiero muszą pojawić się substancje odżywcze, które pomogą odtworzyć ATP potrzebne do rozluźnienia kompleksu aktyna-miozyna warunkującego skurcz.
Oprócz zmęczenia kontrakturę mogą wywołać też silne bodźce termiczne, elektryczne, mechaniczne i chemiczne.
Stężenie pośmiertne
Przez krótki czas po śmierci mięśnie wykazują swe normalne właściwości fizjologiczne. Później jednak pojawia się stan przykurczu podobnego do kontraktury fizjologicznej. Stan ten nazywany jest stężeniem pośmiertnym. Występuje on tym szybciej, im większą pracę wykonywały i większe zmęczenie wykazywały mięśnie przed śmiercią. Wyższa temperatura również ten proces przyspiesza. Przy stężeniu pośmiertnym mięśnie ulegają skróceniu, tracą sprężystość, prążkowanie, pobudliwość, stają się sztywne i twarde.
W normalnej temperaturze stężenie następuje po 1h w mięśniach gładkich, potem w sercu, mięśniach przepony, głowy , szyi, tułowia i na końcu kończyn. Ustępuje po 1-3 dniach, gdy białko ulega autolizie.
Procesy chemiczne zachodzące przy stężeniu pośmiertnym:
Kiedy ustaje krążenie krwi, do mięśni nie docierają substance odżywcze i tlen. Dochodzi więc do przekształcania glikogenu i glukozy w kw. mlekowy i zakwaszenia mięśni. Jednocześnie, ponieważ nie zachodzi fosforylacja oksydacyjna, zaczyna brakować związków wysokoenergetycznych. Brak ATP uniemożliwia rozdzielenie aktyny i miozyny, które wiążą się ze sobą podczas normalnego skurczu, jaki następuje jeszcze chwilę po śmierci. Powoduje to niemożliwość rozkurczu, dlatego mięśnie pozostają skurczone.
1.Móżdżek.
Móżdżek człowieka ma kształt elipsoidalnej bryły spłaszczonej grzbietobrzusznie, o najdłuższym wymiarze poprzecznym. Wyróżniamy powierzchnię górną móżdżku (facies superior cerebelli), bardziej płaską, i powierzchnię dolną (facies inferior cerebelli), silnie wypukloną. Parzyste części móżdzku przez podobieństwo do półkul mózgu nazywane są półkulami móżdżku (hemispheria cerebelli), pośrodku znajduje się nieparzysta część, nazwana robakiem móżdżku (vermis cerebelli). Od móżdżku odchodzą konary móżdżku: górny, środkowy i dolny. Dzięki nim móżdżek otrzyjmuje połączenia z sąsiadującymi strukturami:
konar górny móżdżku prowadzi włókna aferentne i eferentne, z przewagą tych drugich, które łączą się z korą mózgu, śródmózgowiem i przez rdzeń przedłużony z rdzeniem kręgowym
konar środkowy móżdżku prowadzi wyłącznie włókna aferentne, które dochodzą do jąder mostu
konar dolny móżdżku prowadzi włókna aferentne i eferentne, które łączą się przez rdzeń przedłużony z rdzeniem kręgowym
Zarówno półkule jak i robak móżdżku złożone są z istoty szarej i istoty białej. Powierzchnia półkul jest bardzo silnie pofałdowana (po rozprostowaniu zajmuje powierzchnię około 1130 cm² i składa się z istoty szarej, która tworzy korę móżdżku. Pofałdowania kory móżdżku mają postać szczelin móżdżku (fissurae cerebelli), przebiegających poprzecznie na obu powierzchniach móżdzku, dzielących je na zakręty móżdżku (folia cerebelli). Niektóre z tych szczelin sa głębsze i dzielą móżdżek na szereg płacików. Kora składa się z trzech warstw: drobinowej, zwojowej i ziarnistej. Wewnątrz, oddzielone od kory białą warstwą włókien, położone są parzyste jądra móżdżkowe: jądro zębate (nucleus dentatus), jądro czopowate (nucleus emboliformis), jądro kulkowate (nucleus globosus) i jądrowierzchu (nucleus fastigii). Histologicznie charakterystyczne dla móżdżku są komórki Purkinjego (duże, gruszkowate komórki warstwy zwojowej).
Podział anatomiczny
Anatomiczne w móżdżku wyróżnia się płaty: przedni, tylny i kłaczkowo-grudkowy. Płaty przedni i tylny rozdzielone są szczeliną pierwszą.
Podział filogenetyczny i czynnościowy
Ze względu na rozwój móżdżku pełnione oraz przez niego funkcje wyróżniony został dodatkowy podział na:
móżdżek przedsionkowy (móżdżek stary archicerebellum) - odpowiadający za zachowanie równowagi i ruchy gałek ocznych. Poprzez narząd przedsionkowy otrzymuje informacje o pozycji głowy w przestrzeni (w stosunku do otoczenia, jak również w stosunku do samego ciała).
w jego skład wchodzą kłaczek i grudka nazywane wspólnie płatem kłaczkowo - grudkowym - lobus flocconodularis. Uszkodzenie tego płata skutkuje zaburzeniami równowagi z niepewnością stania (astasia) i chodu (abasia). Pacjent chodzi chwiejnie, na szeroko rozstawionych nogach, zataczając się (bezład tułowia). Niepewność ta nie zależy od obecności bodźców wzrokowych (zamknięcie oczu nie pogarsza niezborności).
móżdżek rdzeniowy (móżdżek dawny paleocerebellum) - odpowiadający za koordynację ruchową
móżdżek nowy (neocerebellum) - odpowiadający za planowanie ruchów i napięcie mięśniowe
Funkcje
Móżdżek dostaje informacje z wielu ośrodków mózgu, szybko je analizuje i odpowiednio moduluje, aby ruchy były płynne i dokładne. Decyduje, które mięśnie mają się kurczyć, a których odruch rozciągania ma być zahamowany, z jaką siłą etc. Móżdżek także stale kontroluje przebieg ruchu i wprowadza do niego automatyczne poprawki. Aby to obrazowo wyjaśnić posłużmy się przykładem: widzimy walizkę, którą mamy podnieść. Nie wiemy jednak, że została ona wcześniej przez kogoś opróżniona i teraz jest bardzo lekka. Móżdżek podejmuje więc decyzję, że siła przyłożona ma być dość duża. Efektem jest, że walizka ta "wylatuje w powietrze", a my sami tracimy równowagę. W ułamku sekundy móżdżek redukuje przyłożoną siłę, a nawet każe przeciwstawnemu mięśniowi wyhamować walizkę, aby np. nie został on uszkodzony. Zmienia także napięcie innych mięśni szkieletowych, aby przywrócić równowagę.
koordynacja ruchowa
równowaga
tonus (napięcie) mięśni
uczenie się zachowań motorycznych (np. jazda na rowerze)
decyduje o płynności i precyzji ruchów dowolnych (współdziała z okolicą ruchową kory mózgowej)
Móżdżek otrzymuje informacje z: narządów ruchu - mięśni, stawów i wiązadeł (z proprioreceptorów), ze skóry, narządów wzroku, słuchu, równowagi, rąk, stóp, z okolicy ruchowej kory mózgu, z ośrodków ruchowych rdzenia kręgowego.
Rodzaje informacji docierających do móżdżku:
-o stanie narządów ruchu
-o ruchu aktualnie wykonywanym
- o stanie pobudzenia ośrodków ruchowych
- o zakłóceniach równowagi ciała.
Objawem uszkodzenia móżdżku jest m.in.: ataksja.
2.Grupy krwi i zwierząt.
Podział na grupy uzależniony jest od obecności w otoczce krwinek czerwonych - antygenów, są one glikoproteidami. Osocze poszczególnych grup wykazuje obecność - przeciwciał, skierowanych przeciwko antygenom nieobecnym w krwinkach danej grupy. Do reakcji antygenu z przeciwciałem dochodzi podczas transfuzji niezgodnej grupowo krwi.
U ludzi wyróżnia się 4 grupy krwi: A, B, AB, O
Grupa krwi A-posiada w otoczce erytrocytów antygen A, a w osoczu przeciwciała anty-B.
Grupa krwi B- posiada w otoczce antygen B, w osoczu przeciwciała anty- A
Grupa krwi AB - posiada w otoczce antygeny A i B, w osoczu nie mam przeciwciał.
Grupa krwi 0 - nie posiada antygenów, w osoczu posiada przeciwciała anty- A i anty- B.
Najczęściej występującą grupa krwi jest -A, a najrzadziej AB
Antygen A wyst. Pod postacią @ A1 i A2, dlatego mamy w rzeczywistości 6 grup krwi A1,A2 B,0, A1B, A2B.
Ponadto we krwi człowieka poza głównymi antygenami, wyst. Jeszcze co najmniej 30 antygenów krwinkowych oznaczanych rożnymi literami(M, N, S<,P,G itp.
Niezależnie od układu AB0 istnieje jeszcze układ grupowy Rh. U osobników, u których stwierdza się obecność w otoczce krwinek antygen D, określa się jako Rh+, a u których nie występuje jako Rh-. Najczęściej wyst . odczynnikiem jest Rh+, 85% ludzi , Rh- 15% ludzi.
Cecha charakterystyczna układu Rh jest pojawienie się swoistych przeciwciał anty-D, dopiero po zadziałaniu antygenu D. Przeciwciała anty-D są zawsze wynikiem uczulenia na wprowadzony antygen.
Typowym schorzeniem związanym z ukl. Rh jest choroba hemolityczna noworodków, wyst. W przypadku odziedziczenia przez płód czynnika Rh+, po ojcu, podczas gdy matka ma Rh-. Pod wpływem antygenu D płodu, wytwarzane są przez org. Matki przeciwciała anty-D, których miano wzrasta z każdym nast. Porodem, na skutek przedostania się erytrocytów płodu do organizmu matki. Powstałe przeciwciała przedostają się przez łożysko, zlepiają krwinki płodu, które ulęgają hemolizie i powodują żółtaczkę hemolityczna noworodków.
U zwierząt brak jest prawidłowości w wyst. Przeciwciał i antygenów. Klasyfikacji grup krwi nie jest łatwa na skutek zmniejszonego powinowactwa antygenów i przeciwciał. U bydła wykryto ponad 100 grup krwi, podobnie jak u koni, świń , psów, kotów, owiec. Oznaczanie grupy krwi u zwierząt wykorzystywane w celu ustalenia linii hodowli.
.Wymiana gazów w płucach i tkankach.
Wymiana gazowa w płucach
Przystosowania morfologiczne
Wymiana gazów między powietrzem pęcherzykowym w krwią odbywa się na drodze dyfuzji.
Dyfuzja gazów w płucach zależy od:
różnic ciśnień w obu układach
powierzchni czynnej pęcherzyków i wielkości drogi dyfundujących gazów
grubości błon oddzielającej światło pęcherzyka od hemoglobiny w erytrocycie
Maksymalne zbliżenie krwi do powietrza pęcherzykowego osiągane jest przez szczególną budowę pęcherzyków płucnych i oplatających je naczyń włosowatych.
Budowa pęcherzyka - wysłany 3 typami komórek oddechowych (pneumocytów):
Pneumocyty I rzędu
Ubogi metabolizm własny, dzięki czemu zużywają minimalne ilości dyfundującego przez nie tlenu.
Pneumocyty II rzędu
Zdolne do sekrecji surfaktantu (czynnika powierzchniowego), który pokrywa cienką warstwą wszystkie pneumocyty.
Funkcje surfaktantu:
a) zmniejsza opór sprężysty płuc (dzięki czemu pęcherzyki ulegają łatwo rozciągnięciu w czasie spadku ciśnienia w jamie opłucnej (wdech) i nigdy nie dochodzi do ich zapadania się)
b) zabezpiecza przed infekcją bakteryjną, przed przesiąkaniem płynu międzykomórkowego do pęcherzyka.
c) reguluje poziom wilgotności powietrza oddechowego
Sekrecja pnemocytów uzależniona jest od nerwu błędnego
* pełna czynność nerwu błędnego przypada w końcowych tygodniach życia
płodowego - u „wcześniaka” na skutek braku surfaktantu w czasie wdechu nie
dochodzi do pełnego rozciągania i wypełnienia powietrzem pęcherzyków płucnych
Pneumocyty III rzędu - komórki szczoteczkowe - rola komórek receptorowych
Zaopatrzone w liczne mikrokosmki. Występują pojedynczo i nie we wszystkich pęcherzykach.
Leżą przy zakończeniach czuciowych nerwu błędnego-
ich mechaniczne podrażnienie niedostatecznym wypełnieniem przez powietrze pobudza te zakończenia. Efekt odruchu - silny wdech w postaci westchnienia.
Wszystkie pneumocyty leżą na błonie podstawnej, która styka się z błoną podstawną śródbłonka naczyń włosowatych.
Śródbłonek ten jest zwarty, co przeciwdziała filtracji i zbieraniu się płynu
międzykomórkowego (który byłby przeszkodą dla dyfundujących gazów)
Dyfuzja i wiązanie O2 z hemoglobiną
Odtlenowana krew dopływająca do naczyń włos. pęcherzyków łączy się gwałtownie -
wobec dużej różnicy ciśnień parcjalnych - z dyfundującym z pęcherzyków powietrzem.
W krwince powstaje oksyhemoglobina, w czasie przyłączania tlenu następuje przesunięcie względem siebie łańcuchów białkowych i gwałtowne odszczepienie wodoru.
Powoduje to pobranie z nagromadzonego w krwince chlorku potasu i przyłączenie do oksyhemoglobiny jonu potasowego.
Powstanie oksyhemoglobiny zależy od:
prężności O2 i CO2 w danym środowisku
stężenia jonów H+
zawartości 2,3-dwufosfoglicerynianu (2,3-DPG)
temperatury (jej spadek ułatwia wiązanie tlenu)
Dyfuzja CO2 z krwi do powietrza pęcherzykowego
Cały obecny CO2 we krwi żylnej przenoszony jest w 70% przez osocze i w 30% przez krwinki (najwięcej w postaci wodorowęglanów, uzupełniająco - połączenia karbaminowe)
Przyłączenie tlenu do hemoglobiny powoduje natychmiastowe przyłączenie do oksyhemoglobiny jonów potasowych. Pociąga to za sobą lawinowo przebiegające procesy, które (na krótkim odcinku naczynia włosowatego i w ułamku sekundy) powodują:
przejęcie przez krwinkę rozpuszczonych w osoczu wodorowęglanów
uwolnienie do osocza, a następnie do powietrza pęcherzykowego całej zawartości obecnego we krwi CO2
znaczne stężenie jonów Cl-
Jony te wędrują do osocza, a z osocza (dla zrównoważenia) przenikają HCO3-,
które zmieniają się na kwas węglowy (H2CO3), który pod wpływem anhydrazy
węglowej rozpada się na wodę i dwutlenek węgla.
Dwutlenek węgla przenika do osocza, a stamtąd do powietrza pęcherzykowego.
Wymiana gazowa w tkankach
Odbywa się w czasie przepływu krwi przez naczynia włosowate.
Przekazywaniu tlenu do tkanek sprzyja:
wysoka prężność CO2 w tkankach różnice w prężności O2
zwiększone uwalnianie jonów H+
działanie 2,3-DPG - syntetyzowany w większych ilościach np. w warunkach niedotlenienia tkanek (hipoksji) łączy się z łańcuchami polipeptydowymi w cząsteczce globiny, co powoduje powstanie dodatkowych wiązań między łańcuchami.
Powstaje przez to stabilizacja IV-rzędowej struktury hemoglobiny w jej formie odtlenowanej, utrudniająca łączenie się z tlenem.
Po oddaniu tlenu hemoglobina traci potas.
Prawie równocześnie odbiera CO2 ze środowiska, który w krwince podlega reakcjom:
Zasadnicza część CO2 - łączenia z wodą pod wpływem anhydrazy węglowej (powstaje kwas węglowy)
Kwas węglowy dysocjuje na jon H+ (przyłączony na miejsce potasu) oraz jon HCO3- Wskutek czego w krwince pojawia się znacznie większa koncentracja jonów wodorowęglanowych niż w osoczu, wskutek różnicy stężeń przechodzą one do osocza. Do krwinki zaś przechodzą jony chlorkowe.
Niewielka część - łączenia z globiną (połączenie karbaminowe)
W mięśniach ssaków dyfundujący do komórek tlen jest odwracalnie wiązany przez mioglobinę. Związek tlenu z mioglobiną jest wewnętrznym magazynem tlenu komórki, mioglobina wiążac tlen, a następnie go oddając (w czasie znacznego spadku jego prężności w mięśniach), przez pewien czas skutecznie zapobiega powstawaniu długu tlenowego.
Jej powinowactwo do tlenu jest znacznie większe niż hemoglobiny.
4.Regulacja wydzielania soków trawiennych u ptaków.
U ptaków, podobnie jak u ssaków, układ przywspółczulny pobudza kurczliwość mięśni gładkich przewodu pokarmowego i stymuluje wydzielanie soków trawiennych.
Hormonalna regulacja:
- gastryna - stymulacja wydzielania HCl i pepsyny
- cholecystokinina - stymulacja kurczliwości pęcherzyka żółciowego oraz wydzielania enzymów trzustki (pod wpływem pokarmu dostającego się do dwunastnicy i jelita czczego)
- sekretyna - stymulacja wydzielania wodorowęglanów przez trzustkę
- enteroglukagon - stymulacja glikogenolizy i glukoneogenezy w wątrobie
- polipeptyd trzustkowy - antagonista cholecystokininy, regulacja przemian węglowodanów i tłuszczów
- bombezynopodobne peptydy - stymulacja wydzielania gastryny, enzymów soku trzustkowego
- somatostatyna - hamowanie wydzielania innych hormonów żołądkowo-jelitowych
5.Mechanizm wytwarzania mleka.
Czynność gruczołów mlekowych jest procesem zależnym od współdziałania wielu hormonalnych i nerwowych czynników.
Gruczoły mlekowe są bogato ukrwione. Do gruczołów poprzez tętnice zewnętrzne zaopatrujące napływa krew bogata w tlen oraz składniki niezbędne do biosyntezy mleka. Tętnice po wejściu do środka gruczołu rozgałęziają się na dwie tętnice gruczołu mlekowego: przednią, która zaopatruje przednie ćwiartki i tylną, zaopatrującą tylne ćwiartki. Przenikanie składników krwi do komórek mlekotwórczych może zachodzić na zasadzie dyfuzji oraz aktywnego transportu.
Głównym źródłem kwasów tłuszczowych o krótkich łańcuchach są związki powstające podczas procesów fermentacyjnych w przedżołądkach. Gruczoł mlekowy ma także dużą zdolność wykorzystywania takich substancji lipidowych krwi, jak: octan, beta-hydroksymaślan, lipoproteiny.
Komórki mleko twórcze pęcherzyków wykorzystują substancje pobierane z krwi i syntezują w swej cytoplazmie składniki mleka. Tłuszcz jest syntezowany w retikulum endoplazmatycznym. Kropelki tłuszczu związane z błonami retikulum przesuwane są w kierunku szczytowej części komórki jednocześnie zwiększając swoją objętość.
Biosynteza białek mleka odbywa się w gruczole mlekowym ze składników dostarczonych przez krew. Nie wszystkie aminokwasy wprowadzone do organizmu z pokarmem są wykorzystywane bezpośrednio do syntezy białek mleka. Niektóre są metabolizowane w gruczole mlekowym, np. arginina ulega przemianie do proliny, inne są wykorzystywane do syntezy aminokwasów endogennych. Biosynteza białek mleka zachodzi na rybosomach, które zawierają kwasy rybonukleinowe i około 50 różnych białek, które częściowo współdziałają w tym procesie jako enzymy.
W biosyntezie białek mleka są także wykorzystywane składniki mineralne, jakimi są jony wapniowe, fosforanowe i cytryniany. Fosforylacja kazeiny zachodzi podczas przechodzenia łańcuchów polipeptydowych z retikulum do aparatu Golgiego.
Laktoza podobnie jak niektóre białka mleka tworzy się wewnątrz aparatu Golgiego. Prekursorem laktozy jest glukoza, której poziom we krwi jest regulowany przez dwa przeciwstawne procesy: tempo zużytkowania tego cukru przez komórki oraz jego ilość we krwi. Laktoza należy do tzw. związków osmotycznie czynnych, czyli włączona jest w mechanizm regulacyjny ułatwiający ściąganie wody do pęcherzyków sekrecyjnych.
Przy obniżeniu jej poziomu transport wody jest zmniejszony i powstaje niewielka ilość bardzo skoncentrowanego mleka. Uwalnianie laktozy, podobnie jak białek mleka, zachodzi w procesie egzocytozy. Tą drogą jest wydalany także z komórek mlekotwórczych do pęcherzyków wapń, fosforany i cytryniany.
Wydzielina komórek mleko twórczych podczas przebywania w pęcherzyku może podlegać dalszym zmianom: następuje wymiana wody i składników w niej rozpuszczalnych. Procesy te mogą zachodzić dzięki bogatej sieci naczyń włosowatych otaczających pęcherzyki mleczne i występujących w nich wahaniach ciśnienia hydrostatycznego.
1.Spoczynkowy potencjał błonowy.
Zewnętrzna błona aksonu oddziela jego środowisko wewnętrzne (aksoplazmę) od środowiska zewnątrzkomórkowego (płynu zewnątrzkomórkowego). Ponieważ oddziela ona równocześnie od siebie jony dodatnie i ujemne występujące w tych środowiskach, w warunkach spoczynku istnieje stała różnica elektrycznych potencjałów (różnica napięcia) między zewnętrzną, a wewnętrzną, powierzchnią błony aksonu. Zewnętrzna powierzchnia błony wykazuje potencjał elektryczny dodatni, natomiast wewnętrzna -naładowana jest ujemnie w stosunku do powierzchni zewnętrznej. W związku z tym mówi się o elektrycznej polaryzacji błony aksonu, wywołanej swoistym rozdzielaniem przez tę błonę ładunków dodatnich i ujemnych. Z polaryzacją neurolemy związana jest pobudliwość neuronu i jego gotowość do inicjacji impulsów nerwowych. Opisana różnica potencjałów, warunkująca spoczynkowe napięcie błony, nazywana jest spoczynkowym potencjałem błonowym (membranowym) lub krócej -potencjałem spoczynkowym. Potencjał ten może stać się źródłem prądu elektrycznego, gdy połączymy przewodnikiem elektryczności powierzchnią aksonu z jego wnętrzem. Prąd elektryczny powstający w tych warunkach nosi nazwę prądu spoczynkowego. Potencjał spoczynkowy jest zawsze ujemny, a jego wielkość wewnątrz aksonu wynosi kilkadziesiąt miliwoltów.
Przyczyną potencjału spoczynkowego (napięcia spoczynkowego błony) jest nierówne rozdzielanie określonych jonów: przewaga ujemnych jonów w aksoplazmie i nadmiar jonów dodatnich (kationów) w środowisku zewnątrzkomórkowym. Wnętrze aksonu obfituje w jony potasu (K +), podczas gdy inne kationy występują, tu w bardzo niskich stężeniach. Zawartość jonów potasu jest w aksoplazrnie około 30 razy większa niż na zewnątrz neuronu. Jonami dla nich antagonistycznymi są duże aniony białkowe. W porównaniu z bardzo ruchliwymi, łatwo dyfundującymi na zewnątrz jonami potasu są one z powodu swych rozmiarów mało ruchliwe, a w przypadkach silnie związanych białek strukturalnych, zupełnie nieruchliwe. Przytrzymują one jednak wychodzące z komórki jony K +, przy zewnętrznej powierzchni błony. W środowisku zewnątrzkomórkowym występują natomiast w nadmiarze kationy sodowe (Na+) i aniony chlorkowe (CI-). Stężenie jonów sodowych jest ponad 10 razy, a jonów chlorkowych około 30 razy wyższe na zewnątrz aksonu. Jony Na+ i Cl-, okryte grubszym płaszczem wodnym, są większe o około 30% od jonów K + i tez wykazują dużą, ruchliwość
Z powodu nierównego stężenia jonów (przede wszystkim K + i Na +) wewnątrz i na zewnątrz neuronu jony te wykazują stałą tendencję do dyfuzji przez neurolemę, zgodnie z kierunkiem spadku ich stężeń. Dyfuzja ta może odbywać się poprzez pory występujące w neurolemie, będące w istocie wąskimi kanałami o ścianach naładowanych ujemnie. Wskutek tego odpychają one aniony, a ułatwiają przechodzenie przez nie określonym kationom. W każdym kanale rozróżnia się dwie części: wejście -pełniące rolę wybiórczego filtru dla jonów oraz furtkę -umieszczoną głębiej, zamykającą lub otwierającą przejście przez kanał. Istnieją oddzielne kanały dla jonów sodu i potasu. Średnica kanałów potasowych jest znacznie większa w stosunku do średnicy jonów K + niż średnica kanałów sodowych w stosunku do średnicy jonów Na +.
Błona aksonu w stanie spoczynku przepuszcza w obu kierunkach dyfundujące jony, choć w niejednakowym stopniu. Najłatwiej przenikają przez nią jony potasowe, następnie chlorkowe, a najtrudniej -sodowe. Kanały sodowe są bowiem wąskie a kanały potasowe znacznie szersze. Przepuszczalność błony dla jonów K+ jest 2 razy większa niż dla jonów Cl i około 50 razy większa niż dla jonów Na+. Prócz tego w warunkach utrzymywania się potencjału spoczynkowego o wiele więcej jest otwartych hipotetycznych furtek w kanałach potasowych niż sodowych. Z tego powodu w spoczynku pewna ilość K+ opuszcza bez przerwy neuron
I wspólnie z dodatnimi jonami sodu przyczynia się do powstania dodatniego potencjału elektrycznego na zewnątrz neurolemy Wnętrze aksonu natomiast wykazuje w spoczynku nadmiar jonów ujemnych (nie dyfundujących jon6w białkowych i wnikających do Środka jonów chlorkowych) Spoczynkowy potencjał błonowy swoiście hamuje przepuszczalność neurolemy dla jonów Na +, blokując wejście do kanałów sodowych, podobnie jak tetraetylammonium (TEA), selektywnie obniża przepuszczalność dla jonów K +. Wskazuje to na istnienie w błonie neuronu oddzielnych kanałów sodowych i potasowych. Kontrolę nad furtkami jonowymi mają sprawować wolne jony Ca2+. Nadmiar tych jonów w płynie
pozakomorkówym zmniejsza przepuszczalność błony neuronu dla jonów Na +, a mała ich ilość lub brak przepuszczalność tę zwiększa.
. Układ limbiczny.
W reakcjach emocjonalnych duże znaczenie maja 2 układy znajd. W mózgu: ukł.rąbkowy i siatkowaty.
Jedna z centralnych struktur podkorowych ukł. rąbkowego jest podwzgórze, otaczają go inne struktury podkorowe takie jak:
-pole przedwzrokowe
-przegroda przezroczysta
-pole przywęchowe
-nadwzgórze
-jądra przednie wzgórza- te części idą od góry
a od dołu
-hipokamp
-ciało migdałowate
Te części otacza kora rąbkowa (płat rąbkowy)
Płat rąbkowy ma połączenie z płatem czołowym, okolicą czuciowo-ruchowa, słuchowa i wzrokowa nowej kory.
W ukł.rąbkowym znajd.się ośrodki których drażnienie powoduje u zwierząt stan przyjemności lub przykrości, wywołuje strach lub wściekłość tego powodu nazwany jest MÓZGIEM EMOCJI.
Przy uszkodzeniu tego ukł. zwierzęta staja się nadmiernie agresywne lub nadmiernie łagodne, żarłoczność lub zanik apetytu, zboczenia płciowe itp.
Ukł. ten kontroluje również czynności autonomiczne i wewnątrzwydzielnicze stąd drażnienie różnych jego części ma wpływ na akcje serca, ciśn. Krwi, ruchu jelit.
W stanach emocjonalnych jest czynny tez ukł. siatkowaty od którego zależy poziom pobudzenia mózgu i całego org. A wiec poziom podniecenia. Zasadniczo podłożem motywacyjnego zachowania się zwierząt są występujące wpływy ośrodków podwzgórzowych, obejmujących ukł. rąbkowi i korę mózgową. Zakres rozprzestrzeniania się tych wpływów zależy od stopnia wyjściowego pobudzenia podwzgórzowych ośrodków motywacyjnych. Przy narastającym głodzie dochodzi do stopniowego włączania coraz to większej ilości elementów rąbkowych i korowych w obrębie motywacji pokarmowej. Przy dalszym wzroście natężenia pobudzenia wpływy te szerzą się na hipokamp i obejmują wszystkie struktury ukł. rąbkowego, powodując wys. U zwierzęcia reakcji orientacyjo-badawczej. Pobudzenie dalej dociera - z jednej strony do okolic kory mózgowej, z drugiej strony- do aktywujących aparatów ukł.siatkowatego śródmózgowia. W następstwie wytwarza się odpowiednia reakcja emocjonalna.
3.Podstawowe czynności mięśnia sercowego.
Mięsień sercowy przypomina swą budową mięsień szkieletowy. Jest poprzecznie prążkowany dzięki układowi nitek aktyny i miozyny w miofibrylach (zawiera również tropomiozynę i troponinę). Zwiera jednak większą liczbę mitochondriów co świadczy o dużej aktywności metabolicznej. Istotnym jednak elementem budowy mięśnia sercowego są wstawki łączące sąsiadujące komórki ze sobą. W ich obszarze występują złącza ścisłe (nexus) z koneksonami-kanały białkowe. Zapewniają niski opór elektryczny co umożliwia przewodzenie stanu czynnego pomiędzy komórkami.Dzięki temu komórki mięśnia są sprzężone elektrycznie, a mięsień sercowy stanowi czynnościową zespólnie (syncytium) i zachowuje się jak jedna wielka jednostka czynnościowa. Serce zbudowane jest z dwu oddzielnych zespólni: przedsionkowej i komorowej, rozdzielonych pierścieniem włóknistym. Pobudzenie pierwszej zespólni przewodzone jest do drugiej drogą wyspecjalizowanego układu przewodzącego, Z tego powodu mięsień sercowy zawsze odpowiada na bodziec skurczem maksymalnym zgodnie z prawem `' wszystko albo nic''.Bodźce dla mięśnia są wytwarzane i przewodzone przez odpowiednio wyspecjalizowane komórki samego mięśnia sercowego. Wytwarzane są przez komórki rozrusznikowe skupione w odpowiednich miejscach pomiędzy komórkami mechanicznymi.. Czas trwania potencjału czynnościowego w poszczególnych włóknach jest stosukowo długi (100-200 razy większy niż we włóknach mięśnia szkieletowego). Ponieważ jest on znacznie dłuższy od czasu szerzenia się impulsu skurczowego od pierwszego do ostatniego włókna, wszystkie włókna ulegają depolaryzacji w tym samym czasie. Główną rolę w skurczu i regulacji jego przebiegu odgrywają jony Ca2+. Wapń wiązany jest przez troponinę, co umożliwia interakcję miozyn i aktyny. W czasie pobudzenia wapń dyfunduje z przestrzeni zewnątrzkomórkowej do jej sarkoplazmy przez kanały wapniowe, napływa też z zewnątrz przez odwróconą wymianę Na/Ca, jak również jest uwalniany z siateczki sarkoplazmatycznej. Potencjał spoczynkowy włókien mięśnia sercowego wynosi od -85 do -90 mV. jest to potencjał ujemny.Pobudzenie prowadzi do powstania potencjału +30 mV.Ta depolaryzacja powoduje w efekcie utratę pobudliwości (okres skurczu mięśnia). Refrakcja bezwzględną (całkowity zanik pobudliwości) trwa około 200ms. Dalej następuje stopniowa repolaryzacja gdy mięsień jest już pobudliwy ale tylko na silne bodźce, a trwa to około 30ms—okres refrakcji względnej.Całkowity powrót do stanu wyjściowego, a więc pobudliwości spoczynkowej, następuje w momencie zakończonej repolaryzacji, to jest w okresie rozkurczu mięśnia sercowego.
Najważniejszą cechą mięśnia sercowego jest jego zdolność do samowytwarzania rytmicznie występujących stanów pobudzenia czynnościowego. Powstają dzięki temu rytmiczne skurcze przedsionków i komór. Serce zawdzięcza to w/w tkance bodźcotwórczej. Tkanka ta zbudowana jest z małej ilości miofibryli, dużym jądrze i słabo zaznaczonym poprzecznym prążkowaniu. Główne jej skupisko mieści się w przedsionku prawym tworząc węzeł zatokowy. Drugim skupiskiem jest węzeł przedsionkowokomorowy wraz z przedłużeniem w postaci pęczka przedsionkowokomorowego (Hisa). Wszystkie komórki tego układu mają zdolność do samowytwarzania stanu pobudzenia i mogą powodować wzbudzenie całego mięśnia sercowego. W warunkach fizjologicznych jednak tylko komórki węzła zatokowego inicjują swój rytm (depolaryzują się najszybciej).Stan pobudzenia szerzy się z prędkością około 1 m/s. Z tego powodu przedsionek prawy kurczy się ciut wcześniej niż przedsionek lewy. Pobudzenie z mięśni przedsionków dociera do węzła przedsionkowokomorowego pokonując tzw. strefę graniczną (elementy tkanki łącznej powiązane z licznie rozgałęzionymi włóknami mięśniowymi. Pokonanie tej strefy zwalnia szybkość depolaryzacji do 0,05 m/s, co powoduje wystąpienie skurczu komór serca po zakończeniu skurczu przedsionków. Można powiedzieć iż węzeł przedsionkowokomorowy jest podporządkowany węzłowi zatokowemu jednak gdy rytm pobudzeń z węzła zatokowego nie dociera, jest bardzo szybki lub zbyt wolny, węzeł przedsionkowokomorowy może wytwarzać impulsy pobudzające mięsień sercowy samodzielnie. Ponieważ między mięśniem sercowym przedsionków a komór istnieje pierścień włóknisty, pobudzenie z węzła przedsionkowokomorowego do ścian komór dociera jedynie przez pęczek przedsionkowokomorowy (za pośrednictwem komórek gruszkowatych-Purkiniego). Tutaj szybkość rozchodzenia się pobudzenia jest znaczna i wynosi od 1 do m/s.
Jeśli na serce w czasie skurczu działa dodatkowy bodziec to efekt działania bodźca jest uzależniony od okresu w którym on zadziałał. W okresie refrakcji bezwzględnej mięsień sercowy jest niepobudliwy. W końcowej fazie skurczu (refrakcja względna) oraz podczas rozkurczu wystąpi wówczas dodatkowy skurcz, czyli ekstrasystole. Pociąga to pewne konsekwencje a mianowicie iż jak każdy skurcz ma i on również swój okres refrakcji. Na ten okres może trafić fizjologiczne pobudzenie niedochodzące do skurczu. Powstaje wówczas tak zwana pauza wyrównawcza (kompensacyjna). Skurcze dodatkowe powstają przy nadpobudliwości układu nerwowego, zatruciach, niedotlenieniu serca itp. Mogą one być inicjowane w przedsionku, w węźle przedsionkowokomorowym i w samej komorze. Przy porażeniach prądem, schorzeniach naczyń wieńcowych oraz niektórych uszkodzeniach toksycznych może dojść do nierównomierności albo krańcowo do trzepotania lub migotania przedsionków (lawinowych skurczów całych przedsionków lub partii włókien mięśniowych. Najgroźniejsze dla życia jest trzepotanie lub migotanie komór, dochodzi wówczas z reguły do ustania krążenia.
Wlokno nerwowe pobudzenie i przewodzenie
Włókna nerwowe - wypustki kom. nerwowej (neuryty). Dł. ok. 1 metra. Jedne włókna mają osłonkę białkowo-lipidową (mielina) i są to włókna rdzenne które dzielimy na jednoosłonkowe i dwuosłonkowe. Inne jej nie posiadają i nazywają się wł. bezrdzennymi. Te z kolei dzielima na nagie (nie posiadające osłonki) lub tylko z jedną osłonką.
Przewodzenie impulsu z kom. nerwowej na drugą lub z kom. nerwowej na efektor zapewniją styki zwane synapsami. Pierwsze to międzyneronalne a drugie neronowo-efektorowe. Błona plazmatyczna zakończenia neronu nazywa się błoną presynaptyczną. Wyróżniamy sanpsy chemiczne i elektryczne. W synapsie chem. między nueronami występuje wąska szczelina synaptyczna a przewodznie impulsu jest jednokierunkowe. W elek. błony przylegają do siebie a impuls jest dwukierunkowy.
Fazy wydzielania soku żoladkowego
głowowa - inaczej faza nerwowa, występuje wydzielanie soku żołądkowego pod wpływem impulsacji z nerwów błędnych. W fazie tej uczestniczą zarówno odruchy warunkowe jak i bezwarunkowe.
żołądkowa - pokarm pojawia się w żołądku, na skutek mechanicznego podrażnienia błony śluzowej żołądka zachodzi odruchowe i bezpośrednie oddziaływanie na komórki dokrewne G znajdujące się w błonie śluzowej żołądka. Wydzielają one do krwi gastrynę, która pobudza do wydzielania gruczoły błony śluzowej żołądka.
jelitowa - pod wpływem treści pokarmowej przechodzącej z żołądka do dwunastnicy na drodze nerwowej i humoralnej zachodzi zarówno pobudzanie jak i hamowanie czynności żołądka. Wytwarzane w dwunastnicy gastryna i cholecystokinina pobudzają wydzielanie soku żołądkowego natomiast sekretyna działa hamująco. Za pośrednictwem odruchu jelitowo-żołądkowego następuje hamowanie opróżniania żołądka i wydzielania soku żołądkowego
Swoista odpowiedz komorkowa
Chora komórka nowotworowa, produkcja cytokin, i polknięcie przez limfocyty Tcytotoksyczne
Budowa i czynności jajnikow
Jajnik (łac. ovarium) - parzysty narząd, występujący u kobiet i samic zwierząt. U dojrzałej kobiety ma kształt spłaszczonej elipsoidy o wielkości 5×3×1 cm i masie 6-8 g[1][2]. Jajniki leżą wewnątrz jamy otrzewnej[2] przy bocznych ścianach miednicy na tylnej powierzchni wiązadeł szerokich macicy, do których przywiązane są za pomocą krótkich krezek. Górne bieguny jajników objęte są przez jajowody. Jajniki służą podwójnemu celowi - wytwarzaniu komórek jajowych oraz wydzielaniu żeńskich hormonów płciowych, estrogenów, progesteronu, relaksyny i androgenów[1].
Od chwili osiągnięcia przez kobietę dojrzałości płciowej (pokwitanie) aż do wygaśnięcia jej funkcji rozrodczej (menopauza - przekwitanie), tj. przez okres około 35-40 lat - co ok. 28 dni dojrzewa w jajniku tzw. pęcherzyk Graafa, zawierający komórkę jajową. Dojrzały do pęknięcia pęcherzyk ma średnicę około 1 -1,5 cm, a komórka jajowa około 0,2 mm[3]. Dojrzewanie pęcherzyka zachodzi pod wpływem FSH, zaś pod wpływem LH dochodzi do zwiększenia ilości płynu w pęcherzyku i wreszcie jego pęknięcia[3]. Gdy pęcherzyk pęka, jajo dostaje się do jajowodu, a z pozostałej części pęcherzyka powstaje ciałko czerwone, natomiast z niego ciałko żółte, które wydziela progesteron konieczny dla umożliwienia wszczepienia zapłodnionego jaja w śluzówkę macicy [1].
4.Powstawanie i rola chłonki.
Powstawanie:
Płyn ten powstaje w obrębie krwionośnych naczyń włosowatych różnych tkanek, w szczególności w śródbłonkach nieciągłych. Przewaga filtracji nad resorpcją w tych śródbłonkach prowadzi do nadmiaru płynu międzykomórkowego, który wchłania się do obecnych tam ślepo zakończonych włosowatych naczyń chłonnych, zwanych terminalami. Jest włĄŚCIWEM wewnętrznym srd ustroju, powstaje z produktow metabolizmu Komorek i przesączu z naczyn krwionośnych. W warunkach fizjologicznych filtracja prawie zawsze przeważa nad resorpcją, co warunkuje ciągłe wytwarzanie chłonki, z okresowymi nasileniami towarzyszącymi nadmiernemu wzrostowi procesów filtracji. Powstająca chłonka jest kierowana z terminali do układu krążenia chłonki.
Jest wytwarzana w większości narządów, wyjątek stanowią: ośrodkowy układ nerwowy, szpik kostny, śledziona, łożysko, sznur pępowinowy oraz nabłonki chrząstek, rogówka i soczewka.
Rola:
Wspomaga wchłanianie i transport tłuszczów
Odbarcza układ żylny wrotny przewodu pokarmowego od transportu tych składników
Naczynia chłonne, którymi odpływa chłonka z miejsc jej wytwarzania stanowią swego rodzaju „bajpas” dla tworzącego się w nadmiarze płynu międzykomórkowego, co zapobiega obrzękom
Zaopatruje krew w leukocyty wytwarzane w węzłach i grudkach chłonnych
Bierze udział w odporności dzięki kierowaniu antygenów z chorych miejsc i narządów do odpowiednich węzłów chłonnych oraz doprowadza do ich kontaktu z miejscową pierwszą linią obrony. W konsekwencji powoduje to również skierowanie antygenu na tory odporności swoistej.
Pełni role pomostu pomiedzy krwia a komorkami
5.Przystosowanie do różnych temperatur otoczenia.
Temperatura wewnętrzna zwierząt homeotermicznych jest utrzymywana w granicach 35-42oC. Fakt istnienia gradientu temperatury między organizmami stałocieplnymi a ich otoczeniem determinował rozwój takich mechanizmów, które zapewniały sprawne oddawanie ciepła do środowiska w wysokich temperaturach oraz chroniły organizm przez zbyt dużymi stratami ciepła w niskich. Szczególnie trudne i niebezpieczne dla organizmów stałocieplnych są strefy o wysokich temperaturach, gdzie powietrze jest wysycone dużą ilością pary wodnej. Efektywność oddawania ciepła przez parowanie limitowana jest ilością wody w organizmie oraz zdolnością jej uzupełniania, jak również wilgotnością powietrza. W gorących i suchych środowiskach termicznych niska wilgotność powietrza ułatwia parowanie wody przez pocenie się i zianie, a tym samym oddawanie ciepła nawet wtedy, gdy temperatura powietrza jest zbliżona do temperatury skóry lub nawet ją przewyższa.
Skóra i pokrywa włosowa tworzą strefę kontaktu termicznego. Mają one podwójne znaczenie termoregulacyjne: zapewniają sprawne oddawanie ciepła oraz chronią organizm przed promieniowaniem słonecznym, będącym zewnętrznym źródłem ciepła. U zwierząt obserwuje się trzy typy pokrywy włosowej: 1) luźne i dosyć długie owłosienie, 2) ścisłe i zbite, 3)krótkie i gładkie.
Fizjologiczną miarą przystosowania do niskiej temperatury otoczenia jest obniżenie dolnej temperatury krytycznej, która wyznacza strefę termicznej neutralności, czyli strefę komfortu cieplnego. Strefa termicznej neutralności obejmuje zakres temperatury otoczenia, w którym przemiana energii i zużycia tlenu oraz utrata ciepła przez parowanie pozostają na najniższym poziomie, a zmianom ulega jedynie przepływ krwi w powierzchownych warstwach ciała (skóra). Drugim czynnikiem określającym adaptację do niskiej temperatury jest wartość temperatury wyznaczającej maksymalny metabolizm, który utrzymuje zrównoważony bilans cieplny i stałą temperaturę wewnętrzną. Niektóre ssaki wodne jak np. wieloryby, gromadzą pod skórą duże ilości tłuszczu, który zapewnia bardzo dobrą izolację cieplną. Podobnie świnia domowa.
Innym typem ochrony przed stratami ciepła z organizmu jest wzrost izolacji cieplnej zewnętrznej warunkowany zmianami gęstości, ilości i rodzaju włosa lub upierzenia, mogących ujawnić się pod postacią porostu pokrywy puchowej.
Pokrywa włosowa niedźwiedzi polarnych całkowicie pochłania promieniowanie podczerwone emitowane przez zwierzęta, co chroni te zwierzęta przed stratami cieplnymi.
Zdolność podwyższania metabolizmu przez zaadaptowane do niskich temperatur zwierzęta jest jedną z podstawowych reakcji na działanie zimna. Procesowi temu towarzyszy wiele reakcji takich jak: 1) wzrost pobierania pokarmu, 2)podniesienie poziomu podstawowej przemiany energii, czyli metabolizmu podstawowego 3) wzrost wartości metabolizmu maksymalnego 4)przedłużenie czasu utrzymania metabolizmu maksymalnego, 5)zmniejszenie termo genezy drążeniowej, a powiększenie termo genezy bezdrżeniowej.
3.Dodatkowe gruczoły płciowe.
DODATKOWE GRUCZOŁY PŁCIOWE. Wytwarzają one wydzielinę, która razem z plemnikami tworzy nasienie, czyli spermę. Wydzielina ta zawiera szereg związków, które tworzą środowisko odpowiednie dla plemników, wpływając na ich żywotność i ruchliwość. Od wydzieliny zależy charakterys¬tyczna woń nasienia. W skład dodatkowych gruczołów płciowych wchodzą: gruczoły pęcherzykowe, gruczoł krokowy i gruczoły opuszko¬wo-cewkowe. Gruczoły pęcherzykowe leżą obok ujść nasieniowodów nad pęcherzem moczowym. Pozostałe dwa gruczoły leżą na przewodzie moczo¬wo-płciowym, do którego też uchodzi ich wydzielina.
5.Luteoliza.
Wytwarzany w ciałku żółtym progesteron przygotowuje drogi rodne do przyjęcia zarodka, jego implantacji i dalszego rozwoju ciąży. W przypadku braku ciąży utrzymanie ciałka żółtego jest nie potrzebne. W takiej sytuacji u Su w 13-14 dniu, u Bo 16-17 dniu od owulacji w kom. ciałka żółtego rozwija się proces zwyrodnienia tłuszczowego nazywany luteolizą. Ilość wytworzonego progesteronu gwałtownie maleje i maleje jego poziom we krwi. Proces ten jest wynikiem oddziaływania na ciałko żółte luteolitycznego hormonu prostaglandyny F2 ( PGF2 ), wytwarzanego w kom. błony śluzowej macicy. Komórki te uwalniają PGF2 do włosowatych naczyń krwionośnych i chłonnych błony śluzowej macicy, skąd- naczyniami żylnymi i chłonnymi przechodzącymi przez więzadło szerokie macicy- PGF2 podąża do krążenia ogólnego. Cząsteczki PGF2 nie mogą trafić do krwi tętniczej i są one rozkładane podczas przepływu krwi żylnej przez płuca. Krew tętnicza zaopatrująca jajniki nie ma czynnej PGF2. Aby PGF2 mogła dotrzeć do jajnika i spełnić swoje luteolityczne lokalne dla układu rozrodczego zadanie, przenika ona (na zasadzie mechanizmu przeciwprądowego) z krwi żylnej i z chłonki obszaru więzadła szerokiego macicy, spotyka się w krezce jajnikowej z drobnymi, pętlącymi tu odgałęzieniami tętnicy jajnikowej, przenika do ich wnętrza i z krwią tętniczą dociera do ciałka żółtego. Luteoliza w ciągu kilku dni powoduje całkowity zanik czynności wydzielniczej ciałka żółtego. Gdy wnikają do niego fibroblasty, cały gruczoł przekształca się w bezstrukturalne ciałko białawe(corpus albicans), pozostaje przez długi czas w jajniku. Gwałtowny spadek poziomu progesteronu we krwi po luteolizie jest sygnałem dla podwzgórza do zwiększenia częstotliwości i wielkości (amplitudy) pulsów wytwarzanej gonadoliberyny (GnRH). W rezultacie wolno rozwijające się dotychczas pęcherzyki jajnikowe są pobudzane przez FSH i LH do rozwoju i dojrzewania. W ciągu kilku dni wykształca się dojrzały pęcherzyk(i), osiągając stadium przedowulacyjne.
Gwałtowny spadek poziomu progesteronu we krwi powoduje również zmiany w ukrwieniu i czynności błony śluzowej macicy. Rozbudowana błona śluzowa, przystosowana do przyjęcia zarodka, staje się zbędna. U większości gatunków zwierząt następuje jej obumieranie i resorbcja. U naczelnych złuszczeniu błony śluzowej towarzysz pękanie drobnych naczyń krwionośnych. Ułatwia to wydalenie na zewnątrz, łącznie z krwią, martwej błony śluzowej. Jest to krwawienie menstruacyjne, które kończy jeden okres aktywności jajnika, a poprzedza rozpoczynający się nowy rozwój pęcherzyka jajnikowego.
1.Zmysły.
Węch i smak
Zmysły węchu i smaku umożliwiają ocenę pokarmów i płynów przechodzących przez jamę ustną. R e c e p t o r y w ę c h o w e znajdują się w błonie śluzowej grzbietowo-tylnej
części jamy nosowej, zwanej okolicą węchową (regio olfactoria), oraz w błonie śluzowej przegrody nosowej. Błona ta jest zgrubiała i ma żółtobrązowe
zabarwienie. Pokryta jest nabłonkiem węchowym. Jest on zbudowany z dwubiegunowych komórek węchowych i komórek podporowych. Komórki węchowe są komórkami nerwowymi, a ich włosowate pręciki są elementami pobudliwymi, funkcjonującymi podobnie jak receptory skory. Spełniają one rolę zewnętrznych chemioreceptorów pobudzanych przez cząsteczki chemiczne substancji zapachowych występujących we wdychanym powietrzu,
rozpuszczalnych w wodzie lub lipidach. Receptory węchowe drażnione są też przez cząsteczki zapachowe ulatniające się z pokarmów w czasie żucia, docierające poprzez nozdrza tylne okolicy węchowej. Cząsteczka chemiczna po rozpuszczeniu się w śluzie pokrywającym nabłonek węchowy drażni receptory, wywołując ich depolaryzację. Słabe pobudzenia węchowe inicjują węszenie u zwierząt, przy którym dochodzi do szybkiej wymiany powietrza w okolicy węchowej i silniejszego drażnienia
receptorów cząsteczkami substancji zapachowych. Pomiędzy komórkami węchowymi znajdują się też nagie zakończenia włókien nerwowych, podrażnienie których wywołuje odruch kichania. Włókna osiowe komórek węchowych opuszczają nabłonek i skupiają się w pęczki tworzące włókna węchowe. Włókna te poprzez otwory w kości sitowej docierają do opuszek węchowych, będących częścią płatów węchowych. Nabłonek węchowy zajmuje powierzchnię: u jamnika -75 cm2, u owczarka -150 cm2, u człowieka -5 cm2. Na tej powierzchni znajduje się komórek węchowych: u jamnika -125 mln, u owczarka -220 mln, u człowieka -około 60 mIn. U królika w nabłonku węchowym znajdować się ma około 100 mln receptorów. Zwierzęta z dobrze rozwiniętym węchem nazywane są węchowcami. Należą
do nich prawie wszystkie ssaki, a wśród nich i zwierzęta domowe..
Zwierzęta, kierując się węchem, znajdują pokarm, wodę, wykrywają z dużej odległości
niewidzialnych wrogów, odnajdują i dobierają osobników płci przeciwnej itp.
Z tego powodu receptory węchowe zaliczane są do tzw. receptorów dystansowych (telereceptorow). Komórki węchowe przesyłają powstające w nich informacje bezpośrednio do mózgu. Węchomózgowie stanowi bowiem filogenetycznie najstarszą część mózgowia. Drogi węchowe kończą się w polach węchowych kory mózgowej. W nich właśnie kształtują się odczucia węchowe umożliwiające rozpoznawanie rożnego rodzaju zapachów. Pobudzenia węchowe dochodzą też do podwzgórza i innych struktur układu rąbkowego (hipokampa i ciała migdałowatego). Węch w Życiu zwierząt odgrywa bardzo dużą rolę. Okolica węchowa jest u nich wielokrotnie większa niż u ludzi. Wykrycie kierunku prądów powietrza, które przenoszą ten czy inny zapach ułatwia śluzawica, płytka nosowa czy płytka ryjowa, zwilżane przez wydzieliny odpowiednich gruczołów. Utrata węchu jest dla psa, kania, krowy,
kozicy itp. Równie katastrofalna, jak utrata wzroku dla człowieka. Konie, kierując się węchem, dokładnie orientują się o rożnych zanieczyszczeniach wody. Normalne psy mogą wyczuć niektóre zapachy z odległości 1000 m. Pies może wykryć obecność jednej cząsteczki substancji zapachowej w jednym litrze powietrza. Psy tropiciele mają doskonale rozwinięta pamięć węchową. Substancje zapachowe mają w świecie zwierzęcym bardzo ważne znaczenie biologiczne. Wydzielanie swoistych substancji zapachowych przez jedne zwierzęta i odbieranie ich przez inne ma charakter pewnej przysyłanej z odległości wiadomości. Silne zapachy wydzielin gruczołów twarzowych, kopytowych i innych spełniają rolę środka informacyjnego wśród zwierząt i stanowią np. cechę rozpoznawczą wewnątrz stada. Informacja zapachowa spełnia ponadto bardzo ważną rolę w procesach rozrodu wielu gatunków zwierząt. Swoiste cząsteczki zapachowe (feromony) uwalniane przez wydzieliny rożnych gruczołów skórnych samic w czasie rui, przenoszone z prądem powietrza na dalekie odległości przyciągają i pobudzają samców. Zwierzęta domowe rozpoznają po zapachu swoje młode, a selekcja odpowiednich roślin przez pasące się krowy tez opiera się przede
wszystkim na kontroli węchowej, a następnie -na dotykowej i smakowej. Substancje zapachowe wywołują też pobudzenie układu węchowego u ptaków. Komórki receptorowe smaku, o kształcie wrzecionowatym, zaopatrzone w wypustki w postaci mikrokosmków, skupione Są w części centralnej specjalnych struktur -k u b k ó w s m a k o w y c h, otoczone komórkami podporowymi. Pobudzanie komórek receptorowych stanowi źródło wrażeń związanych z odczuwaniem czterech podstawowych smaków: słodkiego, kwaśnego, słonego i gorzkiego. Niektóre zwierzęta domowe mają prócz tego receptory smaku wody. Komórki smakowe, oplecione cienkimi włókienkami nerwów czuciowych, nie wykazu wybiorczej wrażliwości na poszczególne substancje smakowe, tylko wysyłają impulsację swoistą dla każdego rodzaju smaku. Rejestracja impulsacji z poszczególnych włókien nerwowych ośrodkowych wykazała swoistą częstotliwość dla rożnych bodźców smakowych. Złożone czucia smakowe tworzą się przez jednoczesne pobudzenie wielu receptorów smakowych. Podstawą odróżniania jakości smakowych ma być kod w potencjałach czynnościowych włókien nerwowych zaopatrujących kubki smakowe.
Komórki smakowe Są chemoreceptorami pobudzanym przez substancje rozpuszczalne w wodzie lub w ślinie, wnikające do tzw. jamki smakowej kubków. Kubki te znajdują się zasadniczo w ściankach brodawek grzybowatych, okolonych i liściastych języka, po ich bokach, na szczycie lub u podstawy. Brodawki nitkowate języka nie mają kubków smakowych. Oddzielne receptory smakowe występują w błonie śluzowej policzków,
podniebienia miękkiego, tylnej ściany gardła i nagłośni. Każdy kubek smakowy zawiera 12-20 komórek receptorowych, a w jednej brodawce może się mieścić około 200 kubków. Ogólna liczba kubków smakowych wynosi od u kury około 25, do u krowy -30000. Percepcja smakowa jest lepiej rozwinięta u zwierząt roślinożernych niż u mięsożernych, ponieważ pokarm tych ostatnich jest bardziej jednorodny. Bydło dobrze rozróżnia wszystkie cztery smaki podstawowe. Nie lubi smaku gorzkiego, bardziej toleruje słony, dużą wrażliwość wykazuje na smak kwaśny. Świnie lubią smaki słodki i gorzki. Wolą
raczej pokarm slodko-gorzki niż slodko-kwasny. Receptory smakowe psów odbierają smak cukru, lecz nie sacharyny. Koty bardzo słabo reagują na smak słodki, ale są bardzo wrażliwe
na smak chininy oraz chlorek potasu. U kotów, psów, świń, królików, gołębi i kur występują receptory smakowe pobudzane przez czystą wodę. Nie występują one u przeżuwaczy. Konie lubią tez smak słodki. Wszystkie zwierzęta najmniej lubią smak kwaśny. Przy brakach pewnych składników w środowisku wewnętrznym organizmu zwierzętom smakują pokarmy, które te składniki zawierają. Zwierzęta odczuwające braki pewnych ważnych składników, a pozbawione operacyjnie zdolności odróżniania smaków giną szybko, nie mogąc dobrać odpowiedniego pożywienia zawierającego te składniki.
Włókna czuciowe od kubków smakowych biegną w nerwach twarzowym, językowo-gardlowym i błędnym. Uszkodzenia nerwu twarzowego mogą prowadzić do utraty zdolności dobierania smaków słodkiego, kwaśnego i słonego. Zaburzenia czynności nerwu językowo-gardlowego znoszą odczuwanie smaku gorzkiego. Korowa okolica odbiorcza impulsów smakowych znajduje się w sąsiedztwie obszaru czucia dla jamy ustnej,
zwłaszcza języka. Percepcja smakowa odgrywa bardzo ważną rolę w normalnej
pracy przewodu pokarmowego. Przyjemne dla zwierząt odczucia smakowe
pobudzają odruchowo wydzielanie soków trawiennych oraz wzbudzają apetyt.
Dzięki temu zwierzęta zjadają więcej karmy i dokładniej ją trawią.
Zmysł równowagi
Receptorami równowagi są specjalne p r o p r i o r e c e p t o r y znajdujące się wspólnie z narządem słuchu w uchu wewnętrznym, w przedsionku i przewodach półkolistych. Części te razem tworzą tzw. aparat przedsionkowy będący narządem równowagi. Rejestrują one ustawienie i przemieszczenie całego ciała wzglądem otoczenia, jak również ruchy głowy w stosunku do reszty ciała. Impulsy wysyłane przez receptory aparatu przedsionkowego wywołują powstanie błędnikowych odruchów tonicznych zapewniających lub przywracających normalną postawę, równowagę ciała i ustawienie gałek ocznych. W utrzymaniu równowagi uczestniczą tez odruchy rdzeniowe inicjowane przez pobudzenie receptorów mięśniowych oraz odruchy zachodzące przy udziale móżdżku.
Trzy przewody półkoliste: przedni, boczny (poziomy) i tylny, wypełnione
płynem zwanym śródchlonką (endolimfą), ustawione są w trzech płaszczyznach, prawie prostopadle w stosunku do siebie. W zależności od gatunku zwierzęcia przewód boczny leży, przy normalnej pozycji głowy, mniej lub więcej poziomo. Przewód przedni jest ustawiony prawie strzałkowo, a tylny - poprzecznie, przy czym oba te przewody łączą się ze sobą pod pewnym kątem w stosunku do wymienionych płaszczyzn. Specjalne ułożenie wzglądem siebie obu aparatów przedsionkowych powoduje, ze płaszczyzna tylnego przewodu po jednej stronie jest równoległa do płaszczyzny przewodu przedniego po drugiej stronie głowy. Podobnie w jednakowej płaszczyźnie lezą oba przewody boczne. Przewody leżące mniej
więcej w jednakowych płaszczyznach tworzą tzw. pary czynnościowe. Każdy przewód, rozszerzony u jednego końca w postaci tzw. banki, uchodzi do wspólnej komory nazywanej I a g i e w k ą, połączonej z następnym tworem błoniastym zwanym woreczkiem. Woreczek komunikuje się z narządem słuchu -ślimakiem. W bańkach przewodów półkolistych na występach zwanych g r z e b i e n i a m i są specjalne komórki receptorowe mające po około 50 rzęsek zlepionych substancją galaretowatą. Rzęski tkwiące w tej substancji tworzą wraz z nią w każdej bańce tzw. o s k l e p e k
Obroty głowy wokół osi poprzecznej, pionowej i przednio tylnej, wywołują przesuwanie się endolimfy wewnątrz określonych przewodów półkolistych. W początkowej fazie ruchu lub w przypadku jego przyspieszenia ruch płynnej endolimfy siłą bezwładności opóźnia się w stosunku do przemieszczenia całego przewodu i wywiera nacisk wsteczny na osklepek, przechylając go w stronę przeciwną. Zachodzące przy tym napinanie rzęsek wywołuje pobudzenie komórek receptorowych. Ponieważ przewody funkcjonują parami, w czasie każdego obrotu głowy ruchy osklepka po obu jej stronach odbywają się w kierunkach przeciwstawnych wzglądem odpowiednich przewodów. W ten sposób tworzą się wzorce impulsacji charakteryzujące każdą os obrotu Świadomość dotycząca położenia głowy istnieje także wówczas, gdy spoczywa ona nieruchomo i nie ma przemieszczeń endolimfy. Informacje w tych przypadkach pochodzą z rzęskowych komórek receptorowych znajdujących się w łagiewce i woreczku. Komórki te zgrupowane Są w nich w tzw. P l a m k a c h s t a t y c z n y c h. Rzęski tych komórek pokrywa również masa galaretowata, na której osadzone Są kryształki węglanu wapnia zwane otolitami Mechanizm pobudzenia polega tu na sprężystym odkształceniu rzęsek komórek receptorowych, powodowanym przez otolity, zgodnie z prawem ciążenia. Przy normalnej pozycji głowy jednakowe pobudzenia wychodzą z prawej oraz lewej plamki i identyczne sygnały docierają z obu stron do mózgu, natomiast przy skłonie głowy w lewo lub prawo pobudzenia te są zróżnicowane. Podobnie pochylenie czy odchylenie głowy zmienia kierunek nacisku otolitow na powierzchnie plamki. Plamki reagują prócz tego na ruchy endolimfy przy przyspieszeniach liniowych lub działaniu sil odśrodkowych. Komórki receptorowe baniek i plamek oplatają czuciowe włókna nerwowe przekazujące impulsy drogą nerwów przedsionkowych do jąder przedsionkowych leżących w rdzeniu przedłużonym. Z jąder tych impulsacja jest wysyłana do wszystkich części ośrodkowego układu nerwowego, które zawiadują postawą, równowagą i ruchami, w tym do: kory móżdżku, jąder ruchowych dla mięsni gałek ocznych oraz rdzenia kręgowego. Dzięki temu zmiana położenia głowy w stosunku do tułowia wywołuje natychmiastowe, odruchowe zmiany w napięciu mięśni kończyn i tułowia oraz odpowiednie
ustawienie gałek ocznych. Odruchowe rytmiczne ruchy wahadłowe gałek ocznych, wywołane podrażnieniem komórek nabłonka zmysłowego grzebieni kanałów półkolistych, nazywane są o c z o p I ą s e m.
Wyładowania z narządu równowagi docierają też do kory mózgowej. Przy uszkodzeniach lub zniszczeniach aparatu przedsionkowego niemożliwe jest utrzymanie równowagi ciała oraz wykonanie skoordynowanych ruchów. Po jednostronnym zniszczeniu narządów równowagi zwierzęta przybierają asymetryczną postawę wskutek przechylenia głowy i tułowia na stronę operowaną. Prowadzi to do rożnego rozdziału napięcia w mięśniach kończyn prawej i lewej polowy ciała: po stronie uszkodzenia wzrasta napięcie mięśni prostowników, po stronie zdrowej -mięśni zginaczy. Z tego powodu ulegają zaburzeniu ruchy zwierzęcia. Żaby podczas pływania lub wykonywania skoku obracają się wokół długiej osi ciała. U innych gatunków zwierząt obserwuje się krążenie w koło, tendencję do upadku lub przetaczania się wokół długiej osi ciała, wszystkie skierowane na stronę uszkodzenia. Obustronne uszkodzenie aparatu przedsionkowego nie wywołuje tak dużych zmian w ruchach lub postawie, pogarsza natomiast wyraźnie orientację zwierzęcia w przestrzeni.
Wzrok
U zwierząt duża część informacji o zmianach w otaczającym środowisku dociera do mózgu za
pośrednictwem receptorów wzrokowych. Receptory te, zwane f o t o r e c e p t o r a m i, znajdują się w siatkówce. Fotoreceptory Są pobudzane przez kwanty (fotony) energii promienistej. Fotony to składniki fal świetlnych, elektromagnetycznych, z których tylko
energia fal o długości 350--740 nm może pobudzać receptory wzrokowe. Fale te wchodzą w skład tzw. widzialnej części widma. Zasadniczym zjawiskiem w powstawaniu odczuć wzrokowych jest pochłanianie kwantów światła przez specjalne barwniki fotoreceptorow (barwniki wzrokowe).
Fotoreceptorami siatkówki Są czopki i pręciki.
C z o p k i to Fotoreceptory dzienne, natomiast p r ę c i k i to Fotoreceptory działające w mroku. Oko ludzkie zawiera 20 razy więcej pręcików niż czopków. Zwierzęta nocne (nietoperz, jeż i kret) mają w siatkówce tylko pręciki. Głównie pręciki mają tez sowy, króliki i koty. Ptaki dzienne mają w siatkówce głównie czopki. Największe zagoszczenie czopków występuje w centralnej części siatkówki zwanej p I a m k ą o c z n ą Jest to miejsce najostrzejszego widzenia. Na samym obwodzie siatkówki znajdują się tylko pręciki. Na pozostałym obszarze siatkówki występują zarówno czopki, jak i pręciki. Czopki zapewniają precyzyjne widzenie przedmiotowe i widzenie barw. Pręciki odbierają minimalne bodźce świetlne, jakie daje otaczający świat w czasie zmroku i nocy. I Wykazują one duży stopień wrażliwości, ale mimo to widzenie przedmiotowe za ich pomocą jest mało dokładne i nie zapewnia widzenia barw. W pręcikach występuje barwnik wzrokowy zwany c z e r w i e n i ą w z r o k o w ą lub r o d o p s y n ą. Rodopsyna jest złożonym białkiem zbudowanym z grupy białkowej -opsyny pręcikowej (skotopsyny) oraz barwnika zwanego retinalem. Retinal powstaje w drodze przemian chemicznych z witaminy A, Rodopsyna pod wpływem światła rozkłada się na swe części składowe, natomiast w ciemności ulega resyntezie. Podczas rozpadu rodopsyny dochodzi do pobudzenia pręcików. Z barwników wzrokowych występujących w czopkach znana jest j o d o p s y n a. Prawdopodobnie podczas jej rozpadu na opsynę czopkową i retinal powstaje ciało chemiczne pobudzające zakończenia nerwowe komórek zawierających czopki. Czopki bowiem, podobnie jak pręciki Są tylko wypustkami komórek nerwowych warstwy zewnętrznej siatkowi. W ten sposób jodopsyna odgrywałaby taką samą rolę w procesie widzenia dziennego, jak rodopsyna odgrywa w widzeniu o zmroku. Nie wiadomo jednak, czy jodopsyna występuje w siatkówce ssaków. Przy przystosowywaniu się fotoreceptorów siatkówki do warunków odbierania promieni świetlnych o małym natężeniu, czyli przy tzw. a d a p t a c j i do c i e m n o s c i, dochodzi do resyntezy barwnika wzrokowego we wszystkich fotoreceptorach, tak że minimalne natężenie promieni jest odbierane przez maksymalną liczbę fotoreceptorów. Przy adaptacji do si1nego światła dochodzi najpierw do nadmiernego pobudzenia receptorów wskutek szybkiego rozkładu barwnika wzrokowego. Widzenie w warunkach intensywnego oświetlenia staje się możliwe dopiero wtedy, gdy nadmiar tego barwnika zostanie unieczynniony w przeważającej części fotoreceptorow.
Do syntezy rodopsyny i jodopsyny konieczna jest obecność witaminy A, tak ze jej
niedobory powodują niedowidzenie nocne -kurzą ślepotę. Trzy typy czopków umożliwiają widzenie barw dzięki znajdującym się w nich barwnikom, wychwytującym wybiórczo kwanty czerwieni, zieleni i błękitu. Barwniki te to tzw. e r y t r o 1 a b, c h I o r o 1 a b i c y a n o 1 ab
Inne barwy widma widzialnego mogą powstawać przez odpowiednie mieszanie
wymienionych trzech kolorów. Reakcje powodujące rozpad barwników wzrokowych pobudzają fotoreceptory, wywołując w nich depolaryzację i w ten sposób energia świetlna przetwarzana jest w energię elektryczną impulsów nerwowych. Wynika z tego, ze energia świetlna nie pobudza bezpośrednio receptorów wzrokowych, lecz pobudzanie ich odbywa się za chemicznym pośrednictwem barwników wzrokowych. Jeden czopek nadaje informacje do 1000 lub więcej komórek okolicy wzrokowej kory mózgowej, natomiast impulsy nerwowe od 10--100 pręcików odprowadzane są z siatkówki tylko przez jedno włókno osiowe. W tarczy nerwu wzrokowego w siatkówce brak fotoreceptorow i stąd w jej miejscu występuje tzw. plamka s1epa. Światło dociera do fotoreceptorow, przenikając do wnętrza przez całą grubość siatkówki, natomiast impulsy powstające w nich przebiegają w kierunku odwrotnym, przez łańcuch złożony z trzech neuronów. Zdolność do rozróżniania barw występuje nie u wszystkich gatunków zwierząt Ptaki domowe mają wzrok bardzo dobry i widzę na dużą odległość, natomiast źle widzą przy słabym oświetleniu Nie wszystkie ptaki są w stanie odróżnić kolor czerwony od zielonego Prócz światłoczułej siatkówki można w oku rozróżnić „układ optyczny" składający się z rogówki, soczewki, ciała szklistego oraz cieczy wodnistej. Mają one rożne współczynniki załamywania światła. Promienie świetlne wpadające do oka są tak załamywane przez układ optyczny, by obraz na siatkówce powstał w plamce żółtej. Na zasadzie osobniczego doświadczenia jest on interpretowany zgodnie z rzeczywistą pozycją. Przedmioty znajdujące się w niewielkiej odległości od oczu są ogniskowane na siatkówce dzięki zwiększaniu się głównie przedniej krzywizny soczewki. Na wskutek takiej zmiany jej kształtu zwiększa się siła załamywania (refrakcji) układu optycznego oka. Przy patrzeniu w dal przednia krzywizna soczewki staje się mniej wypukła. Przy ogniskowaniu na przedmiot bliski miesień rzęskowy kurczy się odruchowo i rozluźnia więzadło zawieszające, a elastyczna soczewka zwiększa krzywiznę przednią. W czasie patrzenia na przedmioty odlegle soczewka się spłaszcza. W ten sposób odbywa się
nastawianie, czyli a k o m o d a c j a, układu optycznego oka do patrzenia na przedmioty znajdujące się w rożnej odległości od oka. Akomodacji na przedmioty bliskie towarzyszy zbieżność oczu (konwergencja), a także odruchowe zwężenie źrenicy. Sprawia to, ze obraz oglądanego przedmiotu pada na plamki żółte obu siatkówek.
T ę c z ó w k a spełnia rolę przesłony, regulując ilość światła wpadającego do oka przez z r e n i cę. Pod wpływem światła źrenice się zwężają (miosis), w ciemności -rozszerzają (mydriasis). Wielkość źrenicy zależy od napięcia dwu mięśni gładkich: mięśnia zwieracza źrenicy, zajmującego wolny brzeg tęczówki, i mięśnia rozszerzającego źrenicę, mającego włókna ułożone promieniście. Miesień zwieracz źrenicy jest unerwiony przez włókna przywspółczulne, miesień rozszerzający -przez włókna współczulne. Jeśli załamywanie promieni świetlnych w oku jest zbyt silne lub gałka oczna jest zbytnio wydłużona, promienie równolegle wpadające do oka skupiają się nie na siatkówce, lecz przed nią. Jest to przyczyną wady refrakcji oczu zwanej k r o t k o w z r o c z n o s c i ą. Jeśli zdolność do załamywania promieni w oku jest zbyt słaba lub gałka oczna jest zbyt skrócona, promienie równolegle skupiają się poza siatkówką. Mamy wtedy do czynienia z nieprawidłowością refrakcji oka zwaną d a I e k o w z r o c z n o s c i ą. Innym rodzajem nieprawidłowości refrakcji jest n i e z b o r n o s c 0 k a (astygmatyzm). Polega ona na tym, ze skutkiem nienormalnej krzywizny rogówki albo rzadziej zmian w innych częściach układu optycznego oka, załamujących promienie świetlne, promienie te skupiają się w dwóch lub więcej ogniskach. Obrazy oglądanych , przedmiotów tworzące się wtedy na siatkówce nie są ostre.
Zakres otaczającej przestrzeni, której obraz pada na siatkówką nieruchomego oka, nazywany jest p o l e m w i d z e n i a. Każde oko ma swoje pole widzenia, ale na skutek odpowiedniego ustawienia i ruchów gałek ocznych, jednooczne pola widzenia nakładają się w partiach przyśrodkowych mniej lub bardziej na siebie, dając rożnie rozlegle obuoczne pole widzenia. Przedmioty występujące w tym polu wykrywane są jednocześnie obu oczami. Widzenie obuoczne pozwala na dość dokładną ocenę odległości i wielkości oglądanych przedmiotów, daje dokładniejszą ocenę głębi i proporcji. Widzenie jednooczne nie jest tak dokładne, ale pozwala oku, nawet bez poruszania głową, wykryć np. nagle pojawienie się nowego obiektu w jego polu widzenia lub wystąpienia jakiegoś nieistniejącego dotąd ruchu.
U zwierząt domowych w zależności od przedniego lub bocznego umiejscowienia oczu na głowie rożną wielkość mają jedno- i obuoczne pola widzenia. Kot i niektóre psy mają oczy umiejscowione bardziej z przodu, stąd mają duże obuoczne i małe jednooczne pola widzenia, królik natomiast z oczami umieszczonymi po bokach głowy ma wąskie dwuoczne i rozlegle jednooczne (panoramiczne) pola widzenia. Szerzej rozstawione, przesunięte na boki głowy, wypukle oczy u zwierząt roślinożernych stwarzają im duże panoramiczne pola widzenia pozwalające na śledzenie wszystkiego co się wokół nich dzieje, przy nieznacznych tylko ruchach głowy. U królika po uniesieniu głowy powstaje nawet z tyłu wąskie, obuoczne pole widzenia. Większość zwierząt o rozległych obuocznych polach widzenia orientuje się w środowisku głównie za pomocą wzroku, zwierzęta zaś o bocznie umiejscowionych oczach -głównie za pomocą węchu
SŁUCH
Wyszukiwarka
Podobne podstrony:
Fizyka - plan programu2008 09, biologia, Biologia I rok, od adama, studia, semestr I, Fizyka
oceny, biologia, Biologia I rok, od adama, studia, Antropologia
Biologia gr VIII, biologia, Biologia I rok, od adama, studia, semestr I, Fizyka
kolokwium 1. zagadnienia, biologia, Biologia I rok, od adama, studia, Antropologia
Sprawozdanie ćwiczenia 1, biologia, Biologia I rok, od adama, studia, semestr I, Chemia, Laborki
Sprawozdanie ćw. 3, biologia, Biologia I rok, od adama, studia, semestr I, Chemia, Laborki
Sprawozdanie. Cw.2.destylacja, biologia, Biologia I rok, od adama, studia, semestr I, Chemia, Labork
Biologia Komórki. zaganienia wykładowe, biologia, Biologia I rok, od adama, studia, biologia komórki
zagadnienia na II koło z immuno, biologia, Biologia I rok, od adama, studia, immunologia
bphz sciaga cz 2, biologia, Biologia I rok, od adama, studia
pyt bio 2006, biologia, Biologia I rok, od adama, studia, semestr I, Fizyka
SERCE, biologia, Biologia I rok, od adama, studia, Antropologia
WSKAŹNIKI JAKOŚCI GLEB, biologia, Biologia I rok, od adama, studia, semestr I, Ekologia, ćwiczenia
oceny2a, biologia, Biologia I rok, od adama, studia, Antropologia
Fizyka-egzamin przyład, biologia, Biologia I rok, od adama, studia, semestr I, Fizyka
Biologia Komorki. zaganienia wykladowe, biologia, Biologia I rok, od adama, studia, biologia komórki
ZAGADNIENIA do testu BIOLOGIA 2008, biologia, Biologia I rok, od adama, studia, semestr I, Chemia
Biocenoza, biologia, Biologia I rok, od adama, studia, semestr I, Ekologia, ćwiczenia, Podst Ekol II
Dyfuzja ruchliwosc, biologia, Biologia I rok, od adama, studia, semestr I, Fizyka
więcej podobnych podstron