Biologia komórki roślinnej
21.11.2012
Wykład 1
pierwsze były komórki prokaryotyczne
1 mld 400 mln lat temu powstały komórki eukaryotyczne
mykoplazmy - najprostsze
|
PROCARYOTA |
EUCARYOTA |
Wielkość komórki |
1-10 m |
10-1000m |
Metabolizm |
Anaeroby, aeroby |
Aeroby |
organella |
nukleoid, tylakoidy, mezosomy, rybosomy 70s |
Jądro, mitochondria, plastydy, ER, AG, rybosomy 80s |
cytoplazma |
Brak cytoszkieletu, brak ruchu, brak endo- i egzocytozy |
Cytoszkielet: mikrotubule, różne ruchy cytoplazmy, endo i egzocytoza |
DNA |
Koliste w cytoplazmie tylko egzony |
Bardzo długie, liniowe, introny i egzony, histony, otoczka jądrowa |
RNA i białka |
Transkrypcja i translacja w cytoplazmie |
Transkrypcja w jądrze, translacja w cytoplazmie |
Podział komórki |
Podział prosty komórki i nukleoidu |
Chromosomy, podziały: mitoza i mejoza, wrzeciono podziałowe: kariokinetyczne i cytokinetyczne |
Organizacja komórkowa |
jednokomórkowce |
Głównie wielokomórkowce, różne typy tkanek, 60 bilionów komórek |
Różnice zawartości DNA w komórkach :
PROCARYOTA
mała mykoplazma - 1,6 par zasad DNA x 10 do 6
pałeczka okrężnicy E.coli - 4 par zasad DNA x 10 do 6
duża cyjanobakteria - 16 par zasad DNA x 10 do 6
Eucaryota
mitochondrium drożdży piekarniczych- 2-3 par zasad DNA x 10 do 6
drożdże piekarnicze - 12-20 par zasad DNA x 10 do 6
pierwotniaki - 55-320 000 par zasad DNA x 10 do 6
komórki ssaków - 2 800 - 5 300 par zasad DNA x 10 do 6
Archebacteria- brak peptydoglikanu w ścianie komórkowej. Introny w DNA, różnice w rRNA, lipidach i enzymach. Baterie halofilne (barwnik bakteriorodopsyna), metanogenne, termoacidofilne (60-100stopni C przy pH 1,2)
Eubacteria- Peptydoglikan w ścianie komórkowej. Tylko egzony w DNA. Cyjanobacteria-sinice. Photochlorophyta. Bacteria klasy:Mycoplasmae, Bakterie właściwe, Myxobacterie (śluzowe), krętki, siarkobakterie, riketsje, baterie nitkowate.
Daty:
1665-Hooke użył prymitywnego mikroskopu do opisu małych latek w przekroju korka, którym dał nazwę komórki.
1674- Leeuwenhoek ogłosił swoje odkrycie pierwotniaków. 9 lat później zobaczył pierwsze bakterie.
1833 Brown opublikował swoje mikroskopowe obserwacje storczyków wyraźnie opisując jądro komórkowe.
1838- Schleiden i Schwann zaproponowali teorię komórkową, stwierdzając, że zawierająco jądro komórka jest uniwersalnym elementem budowy tkanek roślin i zwierząt.
1857- Kolliker opisał mitochondria w komórkach mięśni.
1879- Flemming bardzo wyraźnie opisał zachowanie się chromosomów podczas mitozy w komórkach zwierząt.
1881-Cajal i inni histologowie opisali metody barwienia, które ujawniły strukturę komórek nerwowych i organizację tkani nerwowej.
1898-Golgi po raz pierwszy opisał aparat Golgiego, zobaczył go dzięki wybarwieniu azotanem srebra.
1902- Boveri odkrył powiązania chromosomów z dziedzicznością, obserwując ich zachowania podczas rozmnażania płciowego.
1952- Palade, Porter i Sjostrand rozwinęli metody mikroskopii elektronowej, które pozwoliły po raz pierwszy zobaczyć wiele struktur wewnątrzkomórkowych. Wykazano, że mięsień zawiera układ fi lamentów białkowych co było pierwszym dowodem istnienia cytoszkieletu.
1957- Robertson opisał dwuwarstwową strukturę błony komórkowej.
1960- Kendrew stosując krystalografię rentgenowską uzyskał po raz pierwszy strukturę przestrzenną białka (mioglobina ze spermy kaszalota) o rozdzielczości 0,2nm. Perutz zaproponował strukturę hemoglobiny o mniejszej rozdzielczości.
1968- Petran i współpracownicy skonstruowali mikroskop konfokalny.
1974- Lazandes i Weber rozwinęli metody immunocytochemii stosując przeciwciała znakowane fluorescencyjnie do wykrywania cytoszkieletu.
1994- Chalfie i współpracownicy wprowadzili do badań mikroskopowych nowy znacznik - białko o zielonej fluorescencji (GFP).
Komórki eukariotyczne
Różnią się od prokariota wysoce skomplikowaną strukturą wewnętrzną.
Mają cytoszkielet, skomplikowany system organelli błonowych (ER, Ap.Golg) i organelli półautonomicznych (mitochondria, chloroplasty).
Mogą być samodzielnymi organizmami, tworzyć kolonie lub wielokomórkowe agregaty.
Komórka roślinna:
A)Protoplast
-cytoplazma (matrix cytoplazmatyczna, błony: plazmolemma i tonoplast, ER, rybosomy, struktury Golgiego, mikrociała/glioksysomy, peroksysomy, lizosomy, wakuole, mikrotubule i mikrofilamenty)
-jądro
-mitochondria
-plastydy (pro plastydy, leukoplasty, tioplasty, chloroplasty, chromoplasty)
Błony cytoplazmatyczne:
Zorganizowane, warstwowe, układy makromolekularne.
Nadają komórkom indywidualność przez oddzielenie od środowiska i pełnią funkcje niezbędne do życia komórek.
Wysoce selektywne bariery przepuszczalności.
Zawierają kanały, nośniki, pompy.
W komórkach eucariota tworzą oddzielne kompartmenty.
Wspólne cechy błon:
Grubość błon 6-10nm.
Zbudowane głównie z białek i lipidów w stosunku 1:4 do 4:1 oraz cukrowców.
Lipidy mają reszty hydrofobowe i hydrofilowe.
Mają specyficzne białka.
Są asymetryczne, spolaryzowane i półpłynne.
Są miejscem lokalizacji receptorów komórkowych.
Białka błonowe-białka związane ze strukturą błony biologicznej. Pełnią w błonach różne funkcje niezbędne do prawidłowego funkcjonowania komórki.
Występują jako:
-receptory-kontaktowanie komórki ze światem zewnętrznym, endocytoza
-enzymów-jak kompleksy białkowe syntetyzujące celulozę w komórkach roślinnych
-białka wiążące komórkę z innymi komórkami bądź elementami macierzy zewnątrzkomórkowej.
-białka uczestniczące w transporcie-kanały, przenośniki, pompy.
Transport przez błony
TRANSPORT |
||||
Przez błonę |
Pęcherzykowy (z fragmentami błon) |
|||
Bez nośników |
Z nośnikami |
Egzocytoza |
Endocytoza |
|
Dyfuzja prosta Dyfuzja złożona |
Dyfuzja ułatwiona |
Transport aktywny |
|
Pinocytoza Fagocytoza |
|
|
Pierwotny Wtórny Translokacja grupowa |
|
Endocytoza receptorowa |
Cytoplazma
Cytoplazma podstawowa(cytozol) oraz struktury błoniaste (organelle).
Cytozol- płynny, złożony koloid wodny, zawierający białka, lipidy, kwasy tłuszczowe, wolne aminokwasy oraz sole mineralne.
Ważny składnik cytoplazmy to sieć białkowych włókienek mikrofilamentów i mikrotubul, tworzących cytoszkielet.
W cytozolu są pozostałe plazmatyczne składniki komórek i zachodzi większość procesów metabolicznych.
Cytoplazma podstawowa- wypełniająca komórki substancja o pozornie jednolitej strukturze, mogąca przyjmować postać od cieczy do żelu. W niektórych typach cytoplazmy można zaobserwować rozróżnienie na dwa obszary:
-przylegającą do błony ektoplazmę
-wewnętrzną endoplazmę
Enzym markerowy cytoplazmy podstawowej- dehydrogenaza glukozo-6-fosforanowa (G6PD).
Cytoplazmę podstawową przenikają układy błonowe. OD zewnątrz ogranicza ją plazmalemma.
Cytoplazma podstawowa jest substancją koloidalną i w jej skład wchodzą:
-związki organiczne
-związki nieorganiczne pierwiastków: wapń, magnez, potas, cynk, miedź, mangan, fosfor, tlen, chlor, siarka, węgiel, bor, azot.
-faza rozpraszająca- woda
-organella- np. mitochondria, plastydy, peroksysomy, lizosomy, wakuole, cytoszkielet.
Kalmodulina- białko modulatorowe, występujące powszechnie u eucariota, działające w obecności jonów wapnia. Przyłącza 4 jony wapnia powodując zmianę swojej konformacji, w wyniku czego powstaje kompleks, który aktywuje i stymuluje działanie wielu enzymów i pomp błonowo-jonowych i innych białek. M.cz=16,7kDa, ma 148 aminokwasów.
Ubikwityna- małocząsteczkowe białko występujące tylko u eucariota. Pełni kluczową role w naznaczaniu (ubiwitynacji), które mają ulec nielizosomalnej proteolizie. 76 reszt aminokwasowych, 8,6kDa. Struktura ubikwityny jest wysoce konserwatywna- denaturacja nie następuje nawet po gotowaniu czy działaniu stężonym kwasem. Peptyd występujący w preteasomach.
Potranslacyjna modyfikacja białek w ubiwitynacji polega na przyłączaniu do reszt lizyny danego białka grupy karboksylowej C-końca reszty glicyny ubikwityny (gly-76).
Retikulum endoplazmatyczne
Wewnątrzkomórkowy i międzykomórkowy system kanałów odizolowanych od cytoplazmy podstawowej błonami biologicznymi. Tworzy nieregularną sieć cystern, kanalików i pęcherzyków. Szczególnie rozbudowane w komórkach, gdzie zachodzi intensywna synteza białek.
Główne procesy w ER wraz z enzymami
Rodzaj przemian metabolicznych |
Przykład enzymów |
-odcięcie odcinka sygnałowego -n-glikozylacja peptydów -modyfikacja łańcuchów oligosacharydowych peptydów |
-endopeptydaza sygnałowa -transferazy glukozylowe -glukozydaza I -mannozydaza I |
-synteza triglicerydów -synteza fosfolipidów -synteza sfingolipidów -synteza cholesterolu -modyfikacja kwasów tłuszczowych |
-syntetaza kwasów tłuszczowych -fosfotransferaza fosfatydylocholiny -metylotransferaza fosfatydyloetanoloaminy -acetylotransferaza sfingozyny -transacylazy
|
-hydroksylacja węglowodorów -utlenianie nienasyconych kwasów tłuszczowych -przemiany steroidów |
-oksydazy mikrosomowe |
-transport Ca2+ -transport glukozy |
-ATP-aza zależna od Ca2+ -glukozo-6-fosfataza |
-utlenianie -przyłączanie siarczanu -przyłączanie grup metylowych -acetylacja |
-oksydazy mirosomowe -transferaza siarczanowa -transferaza metylowa -acetylazy |
Aparat Golgiego
Organellum występujące niemal w każdej komórce eucariota, służące chemicznym modyfikacjom substancjo zużywanych przez komórkę lub wydzielanych poza nią.
Odbywa się tu:
-sortowanie i dojrzewanie białek i lipidów
-modyfikacje reszt cukrowych glikoprotein i glikolipidów
-synteza polisacharydów i mukopolisacharydów, glikozaminoglikanów, hemicelulozy, pektyny
Struktury błoniaste są dynamiczne, odbywa się przepływ substancji zawartych wewnątrz kanałów i pęcherzyków oraz błon.
W diktiosomie wyróżniamy dwa bieguny:
-cis (formowania)
-trans (dojrzewania)
Od cis do trans następuje wzrost stężenia lipidów. Po stronie cis: transferaza N-acetyloglukozaminy, galaktozylowa, fukozylowa, sialowa.
Sieć cis- przedział ratunkowy dla białe powstałych w ER, które przypadkowo złapały się w pęcherzyki płynące do ap. Golg. Są one wyłapane przez enzymy i skierowane z powrotem
Sieć trans-stacja rozdzielcza i sortująca. Produkty z wnętrza diktiosomu zostają rozsortowane zależnie od przeznaczenia i zapakowane do odpowiednich pęcherzyków:
-transportujących (dostarczają lipidy i białka do błony komórkowej)
-lizosomów i endosomy recyklujące
-egzosomy.
Przedział |
Typowe barwienie lub metoda histochemiczna |
Inne enzymy obecne w przedziale |
Powinowactwo do lektyn |
Najważniejsze procesy zachodzące w danym przedziale |
Bliższy cis |
Wyczernianie OsO4 Reakcja na G6Pazę (glukozo-6- fosfatazę) |
Oksydazy NADH : NADPH zależne Mannozydaza 1 Fosfotransferaza N-acetyloglukozaminylowa |
Konkawalina A ( ConA) |
-usuwanie zewnętrznych reszt mannozowych z oligosacharydów -przyłączanie kwasów tłuszczowych do seryny -przyłączanie acetyloglukozaminofosfor. Do enzymów lizosomalnych -przyłączanie N-acetlogalaktozaminy w procesie O- glikozylacji |
Środkowy |
Reakcja na NADPazę |
Transferaza N-acetyloglukozaminylowa Mannozydaza II |
Aglutynina z rącznika pospolitego |
-usuwanie dalszych reszt mannozowych z oligosacharydów -przyłączanie N-acetyloglukozaminy do oligosacharydó w procesie O-glikozylacji |
Dalszy (trans) |
Reakcja na TPP-azę Reakcja na NDP-azę |
Transferaza galaktozylowa Transferaza fruktozylowa Transferaza sjalylowa Glukozaminidaza-N-acetyloglukozaminy Siarko transferaza 5'nuleozydaza ATPaza |
Aglutynina z kiełków pszenicy |
-Przyłączanie koncowych reszt cukrowcowych w procesie O- i N-glikozylacji. -Usuwanie N-acetyloglukozaminy z enzymów liposomowych i ujawnienie ugrupowania mannozo-6fosforanowego. -Przyłączanie grup siarczanowych do białek, glikoprotein i proteoglikanów. -Kontrolowana proteoliza. -Segregacja białek błonowych i wydzielniczych |
Cytoszkielet
Sieć włóknistych struktur białkowych w komórce eukariotycznej, dzięki którym organella i substancje nie unoszą się swobodnie w cytoplazmie, ale zajmują określone miejsca. Cytoszkielet tworzą filamenty aktynowe (mikrofilamenty), mikrotubule (tu bulina) i fi lamenty pośrednie.
Filamenty aktynowe są strukturami statycznymi, zaś mikrotubule dynamicznymi, kurczącymi się i wydłużającymi, dzięki działalności białek motorycznych. Szkielet komórki jest elastyczny-szybka przebudowa zgodnie z potrzebami komórki.
Mikrofilamenty
Cienkie włókienka aktynowe. Pojedyncza cząsteczka aktyny jest białkiem kulistym. Każdy mirofilament składa się z wielu cząsteczek aktyny połączonych w dwa owijające się łańcuchy. Mirofilamenty mają średnicę 5-9nm, są giętkie i krótsze od mikrotubul.
Mikrotubule
Puste w środku rurki zbudowane z tu buliny. W komórce są 2 rodzaje tego białka - alfa i beta. Afla-tubulina łączy się z beta-tubuliną towrząc hetero dimery, z których powstają mikrotubule, a każda z nich ma 25nm średnicy. Komórka ciągle przebudowuje swoje mikrotubule dodając do nich nowe cząsteczki tu buliny (polimeryzacja) albo odrywając tu bulinę od mikrotubul (depolimeryzacja). Pojedyńcza mikrotubula żyje ok. 10 minut.
Filamenty pośrednie
Włókienka o średnicy 10nm. Tworzą nieregularną, rozgałęzioną sieć w cytoplazmie. Jest och b.dużo w komórkach narażonych na mechaniczne oddziaływanie. Filamenty pośrednie są twarde i nadają sztywność komórkom.
Kinezyny
Białka motoryczne wykazujące powinowactwo do mikrotubul i mające zdolność do poruszania się w kierunku ich dodatniego bieguna. Wraz z dyneinami poruszającymi się do ujemnego bieguna biorą udział w wielu ważnych procesach zachodzących w komórce, m.in. podziale komórkowym, transporcie wewnątrzkomórkowym i poruszaniu się. Kinezyny są ATPazami poprzez hydrolizę ATP uwalniają energię niezbędną do ruchu. Długość 80nm, 360kDa.
Dyneiny
Białka motoryczne poruszające się do ujemnego bieguna mikrotubul. Dyneiny dzielimy na dwie klasy:
-dyneiny cytoplazmatyczne
-dyneiny rzęskowe
Są ATPazami.
Ściana komórkowa
zewnętrzny wytwór protoplastu
podlega bardzo dużym zmianom
skład : polisacharydy, lipidy, substancje aromatyczne, białka
lignina może być traktowana jako cześć sygnałowa, a nie tylko substancja utwierdzająca ścianę
związki fenolowe - cząsteczki sygnałowe
uczestniczy w funkcjonalnej specjalizacji komórki
w przypadku naczyń, podczas rozwoju komórki protoplast obumiera, a dojrzałą komórkę stanowi ściana komórkowa
jest dynamicznym komponentem matriks zew-kom, która zmienia się w czasie życia komórki
skład chemiczny : węglowodany : 11 cukrów i wiele połączeń 5x10 do 9
celuloza - główny składnik . 15-30%, związki aromatyczne
funkcje wynikają nie tylko ze specjalizacji strukturalnej ściany ale także obecności cząsteczek sygnałowych, które pełnią ważne funkcje w komunikacji kom / kom i jądro / kom.
Polisacharydy : celuloza, hemicelulozy, pektyny
Związki fenolowe : ligniny, kwasy fenolowe
Białka : obronne, hydrolazy, transportujące, strukturalne
28.11.2013
Wykład 2
Podział komórek- kondensacja chromatyny
Zmiany w otoczce jądrowej
Wrzeciono podziałowe tworzy się w miejscu jądra
Wrzeciono nie może mieć biegunów
U drożdży wrzeciono podziałowe wewnątrz jądra i zachowana jest otoczka jądrowa
MITOZA
Profaza
kondensacja chromosomów - kontrolowana przez odpowiednie geny oraz białko MPF inaczej p34. Aktywny MPF warunkuje także formowanie wrzeciona podziałowego i rozpad otoczki jądrowej, powoduje fosforylacje białek histonu H1 ( spiralizacja, upakowanie włókienek chromatynowych ) i H3
formowanie wrzeciona podziałowego Astralne ( centriola, zwierzęta ) lub nieastralne ( rośliny) Złożone z 1000 do wielu tyś mikrotubul
Chromosomy przyczepiają się do wrzeciona w kinetochorach i biegną do bieguna.
mikrotubule kinetochorowe
mikrotubule innerbiegunowe ( nie łączą się z chromosomami)
mikrotubule wolne
pula białek tubuliny
białka motoryczne ( kinezyna i dyneina)
→ Kinezyna popycha prążki wzdłuż układu, segregacja chromosomów i mitozie, reorganizacja cytoszkieletu, wydłużanie wrzeciona podziałowego poprzez tworzenie mostków.
→ Dyneina bierze udział w przemieszczaniu się chromosomów w kierunku biegunów wrzeciona podziałowego.
Klmodulina - regulacja puli jonów Ca2+
degradacja otoczki jądrowej i jąderka . Ustaje synteza rDNA, zmiany w białkach cytoszkieletu.
METAFAZA
przemieszczanie się wrzeciona podziałowego w miejsce zajmowane przez jądro
przyłączanie się chromosomów do wrzeciona podziałowego poprzez kinetochory
ruch chromosomów do płaszczyzny równikowej wrzeciona (kongresja)
Chromosomy przyłączają się do wrzeciona podziałowego w płaszczyźnie równikowej w taki sposób, kinetochory chromatyd siostrzanych się połączą z mikrotubulami biegnącymi do przeciwnych biegunów, co po rozpoczęciu anafazy zapewni rozdzielenie chromatyd do przeciwnych biegunów
ANAFAZA
Ruch chromosomów do przeciwnych biegunów zależy od wielu procesów i czynników ale nie zależy od własności wrzeciona podziałowego. Połączenia chromatyd są poza reginami centromerów.
Ruch chromosomów do biegunów jest uzależniony we znacznej mierze od mikrotubul wrzeciona podziałowego.
Trucizny mitotyczne
Kolichicyna zatrzymuje ruch.
Ruch anafazowy składa się z elementów:
redukcja odległości między kinetochorami i biegunami Anafaza A
wydłużanie odległości między biegunami
potem jest degradacja wrzeciona podziałowego.
Telofaza
Regulacja cyklu komórkowego
działanie cyklin i kinaz
wytworzenie aktywnej kinazy M - czynnik MPF
rozpad cyklin
INTERFAZA - produkcja cyklin G1
Bardzo ważne składniki odżywcze, czynniki wzrostowe.
CDK - aktywna kinaza fazy S
Kinaza p34
Synteza cyklin M
Cyklina Dcc 2 musi się połączyć z cyklinami ważnymi w procesie mitozy - tworzy się nieaktywny czynnik MPF - działanie kinaz MO15
MPF - 2 warstwy fosforanowe nieaktywny fosfataza gdy zadziała CD25 - aktywny MPF
MEJOZA
W tkance archesporialnej
1 profaza składa się z 5 stadiów : leptoten(długie cienkie nici), zygoten, pachyten(chromosomy oplatają się i uwidacznianie chromatyn), diploten(terminalizacja) i diapeneza
1 metafaza: ustawienie biwalentów
1 anafaza: rozchodzenie chromosomów
2 profaza jak w mitozie
(różnice między pierwszym podziałem mejotycznym a mitotycznym i w jakich tkankach występują)
05.12.2012
Wykład 3
Mitochondrium
Błona zewnętrzna-przepuszczalna dla cząsteczek do 5kDa, zawiera kanały utworzone przez porynę. Posiada tez enzymy syntezy lipidów i enzymy zmieniające substraty lipidowe w produkty metabolizowane w matrix.
Błona wewnętrzna-pofałdowana w grzebienie zwiększające powierzchnię całkowitą. Białka tej błony mają 3 funkcje: łańcuch transportów elektronów-utlenianie, synteza ATP, białka transportujące do i z matriks
Matriks- Zawiera wiele enzymów uczestniczących w utlenianiu pirogronianiu, kwasów tłuszczowych oraz cyklu kwasu cytrynowego. Zawiera kilka kopii mtDNA, rybosomy, tRNA.
Przestrzeń międzybłonowa- zawiera kilka enzymów wykorzystujących ATP wychodzący z matriks do fosforyzowania innych nukleotydów.
Mitochondria wykorzystują do przekształcania energii pirogronian i kwasy tłuszczowe, które wchodzą do mitochondriów i są rozkładane do acetylo-CoA i metabolizowane z CKT, który redukuje NAD+ do NADH.
Jest tez fosforylacja oksydacyjna:
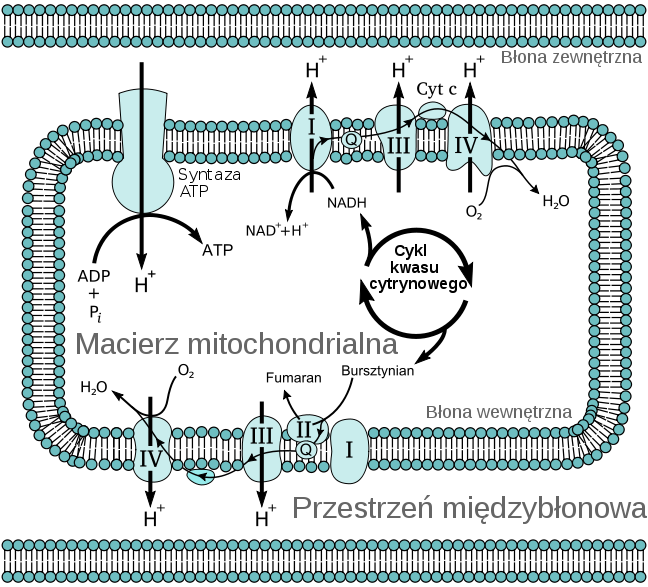
Z każdym przedziałem w mitochondrium są związane inne enzymy:
Błona zewnętrzna - oksydaza monoaminowa (marker), syntetazy acylo-CoA, oraz oksydoreduktazy.
Przestrzeń międzybłonowa - kinaza adenylowa (marker), kinaza nukleozydodifosforanowa.
Błona wewnętrzna - enzymy łańcucha oddechowego, syntetaza ATP, koenzym Q, cytochromy, ubichinon.
Matriks - enzymy cyklu kwasów trókarboksylowych, oksydacji kwasów tłuszczowych, syntezy DNA, RNA i białek.
Syntetaza ATP
Łańcuch przenośników elektronów
Charakterystyka mitochondriów
mogą występować w komórce bardzo często w dużej liczbie lub jest ich niewiele ( kom eukariotyczne ) - ich liczba jest zmienna
struktura może być różna, błona wewnętrzna może mieć bardzo regularny układ, lub błona wewnętrzna może mieć strukturę skondensowaną
rybosomy 70s
błona zewnętrzna o prostej budowie, błona wewnętrzna - złożona budowa
procesy : oddychanie komórkowe, biosynteza białek
liczba powstających cząsteczek ATP jest mała ( z jednej glukozy - 2 cząsteczki ATP)
produktem końcowym gklikolizy jest pirogronian - on trafia do mitochondrium ( ważna jest tutaj błona zewnętrzna i wewnętrzna oraz matriks)
błona zewnętrzna - bardziej przepuszczalna związana z transportem bardzo wielu cząsteczek
błona wewnętrzna - bardziej wybiórcza, na niej przenośniki elektronów, cytochromów, oksydaza cytochromu, syntaza ATP (z nią związany jest przepływ elektronów)
matriks - tu enzymy cyklu Krebsa, cząsteczka DNA, rybosomy 70s, jest tu proces replikacji DNA, transkrypcji i translacji. Tu również białka związane z skokiem termicznym i enzymy związane z utlenianiem kwasów tłuszczowych.
Łańcuch oddechowy - kończy oddychanie
Genom mitochondrialny drożdży ma więcej intronów niż człowieka
Proces biosyntezy białek w mitochondriach
Struktura genomu jest kolista
W matriks może odbywać się replikacja i transkrypcja
Część powstałego mRNA wchodzi w skład podjednostki rybosomalnej, a białka pochodzą z cytoplazmy
Część białek ( np. rybosomów, polimeraza RNA) pochodzą z cytoplazmy
Mitochondria mają częściową autonomie
tRNA wytwarza na podstawie genomu mtDNA
powstałe białka będą stanowiły składnik łańcucha oddechowego - będą wbudowane w wewnętrzną błonę mitochondrium (ale jest to białko złożone, musi się połączyć z innym białkiem pochodzącym z cytoplazmy- tzn to białko powstałe z mtDNA)
proces związany i importem białek
CHLOROPLASTY
błona wewnętrzna, zewnętrzna i stroma
tylakoidy stromy i tylakoidy gran
zwykle w ich pobliżu położone są peroksysomy i mitochondria
peroksysomy nie mają własnych rybosomów i DNA, otoczone pojedynczą błoną
pomiędzy tymi organellami ( + jądro) może nastąpić wymiana materiału genetycznego
Budowa cząsteczki chlorofilu :
pierścień porfiryczny
atom Mg
grupy funkcyjne ( różnią cząsteczki chlorofilu :A, B, C, D, E, F, G )
proces fotosyntezy jest związany z błoną tylakoidów gran w których są anteny fotosystemu I i II odpowiedzialne za pobieranie kwantów energii świetlnej - tworzą się związki wysokoenergetyczne
transport elektronów i protonów w błonie tylakoidu
INNE PLASTYDY
proplastydy
leukoplasty
amyloplasty
lipidoplasty ( z kropelkami tłuszczu )
etioplasy = ciało prolamylarne
proplastydy (w obecności światła tworzą się z nich chloroplasty a jeżeli brak światła to etioplasty)
chloroplasy mogą się zmieniać w etioplasty i odwrotnie
chromoplasty - prosta budowa wewnętrzna, karoteny i ksantofile często w formie kryształów
12.12.2012
Wykład 4
Peroksysomy
enzymy przyczyniające się do syntezy i rozpadu nadtlenku wodoru ( katalazy i peroksydazy )
wytworzenie bursztynianu, który potem w mitochondriach bierze udział w cyklu Krebsa
wymiana związków związanych z fotooddychaniem
odbywa się tu pierwotne oddychanie
otoczone pojedynczą błoną
brak rybosomów i własnego DNA
ściśle związane metaboliczne z mitochondriami i chloroplastami
podobne pochodzenie do mitochondriów i plastydów
import białek
jeśli powstanie Ag powierzchniowe - ich ekspresja
lokalizacja jonów wapnia, precypitaty Ca przy plasmodesmach, inkluzje w chloroplastach, pojawienie struktur białkowo- tubularnych
JADRO KOMÓRKOWE
diploidalna forma lub poliploidalna ( do kilku tysięcy n)
przyjmuje różne kształty
zawiera jąderko, chromatynę, sok jądrowy,
otoczka jądrowa jest porowata, są to kompleksy odpowiednio rozmieszczone w otoczce
8 obwodowych białek +1 - wydostawanie się mRNA rybosomów
jądro - struktura nitkowata ( chromocentryczna )
białka - laminy w zwierzęcych a w roślinnych MFP-1 i MAF1(też typu lamin)
jąderko: części globularne i fibrylarne, wakuole jąderkowe
pory jądrowe- na zewnątrz b.fibrylarne a do wewnątrz koszyczek → transport białek oraz kw.nukleinowych i wirusy np.hiv, oncovirusy, influenza
macierz jądrowa: struktura włóknisto-ziarnista, głównym składnikiem białka oraz RNA - 60-80% to hnRNA (heterogenny), udział w organizowaniu struktury DNA
obszar międzychromatynowy: włókna okołochromatynowe, ziarnistości okołochromatynowe
HISTONY; BIAŁKA CHROMATYNY
DNA - długa, liniowa cząsteczka 2m DNA, 2nm średnicy
Upakowanie DNA - nukleosom, solenoid, pętle, minipasma, chromosomy
Białka zasadowe
Różnią się liczbą aminokwasów oraz stosunkiem Agr/Lys
Heterochromatyna-późna faza a euchromatyna-wczesna
Transkrypcja a histony rdzeniowe
połączenie DNA z histonami ogranicza dostęp białkowych regulatorów transkrypcji, do ich promotorów ( są czynniki zdolne do zmiany struktury nukleosomów i ułatwiają transkrypcję - białko SWJ / SWF ) To białko jest częścią polimerazy II i nadaje zdolność do reorganizacji nekleosomów
inne kompleksy - nucleosome remodelling factor ( NURF)
Rola histonu H1 ( in vitro represor )
wpływ na kondensacje chromatyny w zależności od stężenia NaCl
H1 łączy się z DNA lawinowo
H1 ma warianty sekwencyjne
Ulega modyfikacjom potranslacyjnym
Wczesne i późne H1
Specyficzność tkankowa ( brak H1 u drożdży )
6-9 wariantów sekwencyjnych w komórce
ulega różnym modyfikacjom wynikiem jest zmienna długość łącznikowego DNA
FUNKCJA CHROMATYNY
DNA w jądrze człowieka 2 m długości, jądro średnica - 3,4 do 20 m, struktura nukleosomowa bardzo ważna w zwiększaniu zachowania DNA w jądrze.
Enzymy uczestniczące w replikacji DNA
helikaza - rozwinięcie heliksu DNA, odseparowanie nici rodzicielskiego DNA
prymaza DNA - synteza starterowych nici DNA
polimerazy DNA - synteza nowych nici DNA, alfa - replikacja nici opóźnionej DNA, beta jądrowego DNA
rybonukleaza
endonukleazy
ligaza
topoizomeraza
JĄDERKO
cześć granularna( 2 podjednostki rybosomalne), włóknista
„wakuole”
centrum fibrylarne z DNA jąderkowym
jąderko ma swój cykl jąderkowy
w interfazie 1-4 jąderka
rozproszenie jąderka ( dezintegracja )
potem asocjacja jąderka
na jedną z par chromosomów organizator jąderka NOR (w przewężeniu wtórnym )
połączenie tego organizatora jąderka pod otoczka jądrowa -
białka LAMINY - umocowywanie, kondensacja składników jąderka
duża podjednostka: 3 rodzaje RNA
mniejsza podjednostka: 1 rodzaj RNA
jest warunkiem koniecznym prawidłowego rozwoju np. w embriogenezie, homeostazie dojrzałych organów, prawidłowe funkcjonowanie ksylemu, aerenchymy itd.
apoptosis like - podobna do zwierzęcej ale są pewne różnice
procesy autolizy i autofagii towarzyszące śmierci kom. mają duży udział w uśmiercaniu komórek roślinnych
oligonukleosomalna fragmentacja DNA
udział Ca2+
zmiany poziomu fosforylacji białek
degradacja kom. spełniających tymczasowe funkcje
eliminacja tapetum
degradacja kom. bielma
degradacja protoplastu jako końcowy przejaw różnicowania kom. - elementy naczyniowe
powstawanie aerenchymy
kinazy białkowe zależne od cyklin - Cdk - indukują etapy poprzez fosforylacje reszt tyrozynowych i treoninowych wybranych białek, stanowią podjednostkę katalityczną MPF (Maturation promoting factor)
cykliny - białka regulatorowe, które przez przyłączenie do Cdk umożliwiają kinazom dokonanie fosforylacji innych białek
System naprawy uszkodzonego DNA - telomeraza - u zwierząt w kom. płciowych a u roślin we wszystkich
Nieprzypadkowa segregacja chromatyd w wierzchołkach paprotników
Pola morfogenetyczne w szczytowych częściach organów roślin nasiennych (zwolnienia podziałów komórkowych)
Dryft inicjałów w wierzchołkach korzeni czy pędów, chimery (organizmy zbudowane z komórek różniących się genetycznie)
Informacja cytoplazmatyczna - genom w mitochondriach i plastydach; obecność RNA i określonych białek w cytoplazmie oraz czas ich trwania i rozkładu w komórkach
Receptory sygnałów - kompetencja kom.
Sygnały bliskiego zasięgu
Sygnały dalekiego zasięgu
przez nicienie do włośników
400 gatunków roślin podlega infekcji
pałeczki +ssRNA 4-10 genów
składa się z dłuższej i krótszej cząsteczki, krótsza zawiera informacje o białkach kapsydu oraz białka związane z transportem
karłowatość
nekrozy
zmiany na organach
żółknięcie
enzymy w jąderku - polimeraza DNA I - numatryna ( dojrzewaniem rybonukleoproteidów )
nukleoina ( udział w transkrypcji rRNA)
fibrylaryna
rybocharyna
Rybosomy
19.02.2013
Wykład 5
Programowana śmierć komórki podczas rozwoju roślin
Dwie formy uśmiercania: fizjologiczna i patologiczna
Fizjologiczna
zaprogramowana w genomie podczas rozwoju i morfogenezy organizmu
→ w celu kontroli liczby komórek
→ usunięcie kom. uszkodzonych
→ usunięcie kom. potencjalnie szkodliwych
PCD - program śmierci komórki - towarzyszy rozwojowi sporofitu i gametofitu, morfogenezie tkanek embrionalnych, homeostazie organów, starzeniu tkanek roślinnych lub w odpowiedzi na infekcję mikroorganizmami.
np.: rozwój gametofitu żeńskiego, degradacja wieszadełka, kom. bielma, eliminacja pręcików w kom. żeńskiej, tapetum, kom. czapeczki, różnicowanie naczyniowych elementów ksylemu
Patologiczna
nekroza - powstaje wskutek mechanicznych urazów lub stresów abiotycznych: temp., związki toksyczne itp.
Aktywatory i inhibitory śmierci kom. można wyróżnić u roślin i u zwierząt.
Cechy apoptozy u zwierząt zaobserwowane u roślin:
Zależna od kaspaz - proteinaz cysteinowych
Rośliny mają dużą liczbę genów inhibitorów proteaz cysteinowych umieszczonych specyficznie w organach roślin.
System kaspaz i ich inhibitorów jest regulowany przez wiele czynników np. stres oksydacyjny.
Geny programu śmierci komórki u zwierząt: dad-1, lsd-1, acdl-1 a u roślin bardzo podobne.
Mitochondria - kluczowe organella w kontroli śmierci kom. (cytochrom c, AIF, Apaf-1, pory w zew. błonie, białka bcl, bcl-1)
Udział wakuol w których są enzymy trawiące białko cytoplazmatyczne.
W komórkach zwierzęcych fagocyty usuwają umierające protoplasty - heterofagia
U roślin autofagia - enzymy wakuoli, rzadziej autoliza
Rodzaje RNA
mRNA - przekazywanie informacji - kolejność aminokwasów w biosyntezie białek
rRNA - struktura rybosomu, aktywność transferazy peptydowej, translokacja
tRNA - cząsteczki przenoszące aminokwasy w biosyntezie białek i w innych szlakach metabolicznych
Niskocząsteczkowe RNA:
U1, U2, U4/U6, U5 snRNA - wycinanie intronów
U3 snRNA - dojrzewanie rRNA
U7 snRNA - obróbka końcowa 3'mRNA histonowego
M1 RNA - komponent kataboliczny RNazy P biorącej udział w obróbce tRNA
Regulacja cyklu komórkowego
System sterujący cyklem ma specyficzne punkty kontrolne wielkości komórki, które zatrzymują cykl, czekają aż komórka osiągnie wielkość krytyczną.
1. w fazie G1 przed replikacją DNA
2. w fazie G2 przed wejściem w mitozę
Białka cyklu komórkowego
09.01.2013
Wykład 6
Wzrost i różnicowanie
Rośliny wyróżnia nieograniczony wzrost. Zapewniają to merystemy - obszary kom. O cechach embrionalnych zdolne do ciągłych podziałów.
Merystemy apikalne i boczne.
Różnicowanie komórek roślinnych jest procesem w którym pochodzące z merystemów, podobne komórki z tą samą informacją genetyczną, w wyniku niejednakowej ekspresji genów zaczynają syntetyzować inne białka, wykształcają odmienne cechy, inne pod względem struktury jak i funkcji.
Proces różnicowania komórek
W organizmach wielokomórkowych podlega regulacji ponadkomórkowej, ważny jest w tym system sygnalizacji.
Czynniki transkrypcyjne lub RNA - zdolność regulowania ekspresji genów podrzędnych.
Działanie tych czynników i hormonów jest dla komórki źródłem informacji pozycyjnej (los komórki zależy do jej pozycji w organizmie roślinnym). Ekspresja genów regulatorowych zależy od sygnałów docierających do roślin z jej środowiska zewnętrznego.
Badania nad mechanizmami specyfikacji losów kom. oraz mechanizmami prowadzącymi do powstania określonego fenotypu prowadzone w warunkach modelowych - zawiesina komórek miękiszowych, komórki glonów.
Determinanty losów komórki: wewnętrzne i zewnętrzne
Wewnętrzne - informacja genetyczna i jej ochrona - genety - w najstarszych drzewach najmłodsze pędy nadal wytwarzają pełnowartościowe nasiona.
Zewnętrzne
-plazmodesmy
-domeny izolowane symplazmowo
-system apoplastowy
-fala auksynowa
-sygnały elektryczne
-substancje transportowane w ksylemie i floemie
-sygnały ze środowiska zewnętrznego o charakterze chemicznym, temp., grawitacja, dł. fali świetlnej
Powstawanie informacji pozycyjnej zróżnicowanej czasowo-przestrzennie, mapy morfogenetyczne i zegary biologiczne.
16.01.2013
Wykład 7
TRV - wirus nekrotycznej kędzieżawki tytoniu
Choroba wywołana przez wirusa ma znaczenie ekonomiczne
TRV
Porażenie przez wektor który znajduje się w glebie
lub
inakulacja mechaniczna
lub
inserty+infektory+zdrowa część
Infekcja:
Zmiany w komórkach:
W epidermie korzenia - cząstki TRV krótsze i dłuższe oraz zmiany w ER
W miękiszu - b.dużo cząstek wirusa w cytoplazmie oraz w apoplaście
W młodych naczyniach transportowane cząstki wirusa, nekrotyczne kom. miękiszowe, dużo nieopłaszczonych cząstek w naczyniach.
Ściana komórkowa rozluźniona - opłaszczone cząstki
Ściany w bulwach dużo grubsze niż w liściach.
Często cząstki krótkie oddzielają się od długich.
Mitochondria - tworzą się pęcherzykowate struktury, najczęściej opłaszczone krótkie cząstki.
Jądro kom. - nieopłaszczone cząstki wewnątrz
Przy infekcji roślin tym wirusem organella na pewno są miejscem obecności cząstek wirusa.
10
Wyszukiwarka
Podobne podstrony:
Wykład 5, Biologia UWr, II rok, Biologia Komórki Roślinnej
Fizjologia roślin wykłady, Biologia, fizjologia roślin
Biologia komórki roślinnej, Biologia komórki roślinnej #1, Biologia komórki roślinnej prof
Biologia komórki roślinnej, Biologia komórki roślinnej #1, Biologia komórki roślinnej prof
Biologia komórki roślinnej, Biologia komórki roślinnej #8, Każda błona systemu GERL jest asymetryczn
Biologia komórki roślinnej, Biologia komórki roślinnej #3, Reakcja grawitropijna
cwiczenia - notatki, II ROK, III SEMESTR, Biologia komórki roślinnej
wyklad z biologii komorki!, Ogłoszenia, plany, ważne, Ogłoszenie
Pytania na egzamin, pytania - biologia komórki roślinnej 2007, 1
BIOLOGIA, komorki rosline i zwierzece
Fizjologia roślin wykłady, Biologia, fizjologia roślin
Wykład 3 1 Biologiczna Ochrona Roślin
BIOLOGIA KOMÓRKI ROŚLINNEJ
wykład 6 fizj roślin, biologia, fizjologia roślin
wyklad 4 fizj roślin, biologia, fizjologia roślin
Wykład piąty biologia komórki
więcej podobnych podstron