Ćwiczenia I - Mikroskopy - Ogólna budowa komórki roślinnej. Różnicowanie, Programowana śmierć komórki roślinnej (PCD).
Mikroskop fluorescencyjny to mikroskop świetlny używany w badaniach substancji organicznych i nieorganicznych, którego działanie oparte jest na zjawisku fluorescencji i fosforescencji, zamiast, lub razem ze zjawiskami odbicia i absorpcji. Fluoroscencja próbki może być pochodzenia naturalnego (np. fluoroscencja chlorofilu) lub być wynikiem dołączenia do elementów obserwowanej próbki fluoroforów, czyli substancji chemicznych, które fluoryzują po wzbudzeniu światłem o określonej długości.
Mikroskop polaryzacyjny - mikroskop ten posiada między okularem i źródłem światła dwa filtry polaryzacyjne. Jeden z nich jest nazywany polaryzatorem i znajduje się między źródłem światła i analizowaną próbką, a drugi jest nazywany analizatorem i znajduje się między próbką a tubusem. Polaryzator i analizator przepuszczają tylko tę część światła, która ma ściśle określoną polaryzację. W trakcie obserwacji są one skręcone względem siebie o 90° (są skrzyżowane), co powoduje, że jeśli próbka nie skręca płaszczyzny polaryzacji światła, to nie może ono przejść przez cały układ. Jeśli próbka jest aktywna optycznie, to wówczas przynajmniej część światła przechodzi przez analizator i dociera do oka obserwatora.
Mikroskop konfokalny (laserowy skaningowy) - nowoczesna odmiana mikroskopu fluorescencyjnego, w którym źródłem światła jest laser. Mikroskop ten umożliwia dokonywanie tzw. przekrojów optycznych preparatu, analizuje bowiem światło pochodzące z jednej jego płaszczyzny, eliminując światło docierające z warstw położonych wyżej lub niżej. Różnica między zwykłymi mikroskopami nawet fluorescencyjnymi polega na tym, że dzięki mikroskopowi konfokalnemu otrzymujemy obraz o lepszej rozdzielczości i kontraście. Dzięki tej metodzie możliwa jest analiza przekrojów optycznych w czasie ciągłym położonych na powierzchni lub w głębi preparatu.
Mikroskop kontrastu fazowego:
- badania przyżyciowe
- świetlny
- przesłona pierścieniowa i płytka fazowa
- przepuszcza światło na zasadzie interferencji
Charakterystyka ogólna komórki roślinnej:
W komórkach roślinnych miozyna występuje w niewielkich ilościach; stosunkowo łatwo można ją zidentyfikować w olbrzymich komórkach ramienic.
Komórki roślinne nie są samodzielne (bo rosną w układzie).
Ściany komórkowe sąsiednich komórek są połączone, tworzą system zwany apoplastem (system połączonych blaszkami ścian komórkowych, umożliwiający szybki transport wody, hormonów i transmiterów pomiędzy protoplastami komórek).
Ściana komórkowa stanowi środowisko zewnętrzne - podobne do ECM (przestrzeń / macierz zewnątrzkomórkowa). Szkielet ściany łączy się z mikrofilamentami wewnątrz protoplastu.
W ścianach komórkowych występują różnorakie enzymy (np. enzymy lignifikacji) oraz wypustki białkowe (receptory); procesy zachodzące w ścianie mogą mieć wpływ na metabolizm komórki.
Rośliny wyższe posiadają plastydy typu zielenicowego (zawierające chlorofil typu A, B oraz karotenoidy). Materiałem zapasowym jest skrobia magazynowana w amyloplastach.
Pomimo, że pierwotniaki uważane są za organizmy jednokomórkowe, u niektórych grup(Apicomplexa) występują pierwotne plastydy.
Centralne miejsce w komórce zajmuje ogromna wakuola wypełniona wodą; jej zadaniem między innymi jest nadawanie turgoru komórce - wakuola rozpiera komórkę i powoduje jej rozrost przestrzenny ( w przypadku zdegradowania ścian, komórka pęka).
Jądro komórkowe oddzielone jest od cytoplazmy podwójną błoną (nukleolemmą); system błon wewnętrznych (SER, RER, AG), mitochondria oraz peroksysomy; Wszystkie powyższe struktury są analogiczne do występujących w komórce zwierzęcej.
Immunolokalizacja
Specyficzność znakowania struktur, związków dzięki reakcji antygen-przeciwciało
Wyróżnia się:
- pośrednią (bardziej precyzyjna, najpierw przeciwciało pierwotne nie znakowane, następnie na nie przeciwciało wtórne znakowane i łączenie się z nim, następnie przyłącza się antygen)
- bezpośrednią (od razu przeciwciało wtórne, oznakowane)
Plazmoliza - proces tracenia wody w komórce w roztworze hipertonicznym. W wyniku tego następuje obkurczenie cytoplazmy od ścian komórki. Dotyczy ona wyłącznie komórek roślinnych.
* po dodaniu rodanku potasu (KCNS) następuje plazmoliza kątowa
* po dodaniu octanu amonu następuje plazmoliza kapturowa (wypukła)
Determinacja komórki - jest to określenie w co zostanie zróżnicowana komórka merystematyczna, na determinację ma wpływ:
- pochodzenie danej komórki (prokambium => elementy waskularne)
- roślin ważniejsza jest informacja pozycyjna czyli informacja o położeniu danej komórki
- nierównocenny podział komórki (zależność od podziału antyklinarnego i peryklinarnego - w inne struktury zostają przekształcone zależnie od typu podziału)
- o kierunki różnicowania decydują też komórki towarzyszące i ogólny plan budowy (na zewn. okrywające, wewn. np. przewodzące itp.)
- morfogeny - są to czynniki wpływające na różnicowanie, transportowane czynnie lub dyfunduje, komórki pod ich wpływem różnicują się zależnie od stężenia, takimi morfogenami możliwe że są hormony
PCD - jest to programowana śmierć komórki, nie mylić z apoptozą (to jest jeden z rodzajów tego procesu).
Występuje w komórkach które np. mają funkcjonować jako martwe (sklerenchyma, ksylem) - dochodzi do samotrawienia (lizy protoplastu).
Towarzyszy komórkom przechodzącym mejoze - 3 mejospory degenerują właśnie w sposób zaprogramowany, tylko 1 z nich zostaje, występuje też w rozwoju zarodkowym, jeśli jest coś niepotrzebne.
W kwiecie jednopłciowym (żeński + męski) jedna część z nich ulega programowanej śmierci.
GFP -
GUS -
Ćwiczenia II - Cytoszkielet. System błon wewnętrznych. Peroksysomy komórki roślinnej.
Cytoszkielet:
Do najważniejszych składników cytoplazmatycznych komórki eukariotycznej należy cytoszkielet (jest to zespół włókien białkowych o różnym składnie chemicznym, architekturze oraz funkcji).
Rozróżniamy trzy rodzaje włókien wchodzących w skład cytoszkieletu (podział ze względu na wielkość):
mikrotubule (MT) - średnica około 24 nm (budują np. wici i rzęski)
filamenty pośrednie (IF) - średnica 7 - 10 nm
mikrofilamenty (MF) - średnica 5 - 8 nm (budują np. mikrokosmki).
Mikrotubule zbudowane są z tubuliny - białka konserwatywnego genetycznie (wszelkie mutacje genów kodujących tubulinę są letalne). In vitro można dokonać polimeryzacji mikrotubul z podjednostek pochodzących z różnych źródeł. Mikrotubule mogą być stabilne (rzęski, wici) lub dynamiczne (np. wrzeciono podziałowe).
Filamenty pośrednie mogą być budowane z różnych rodzajów białek (o wspólnej domenie w części α-helikalnej oraz tym samym sposobie polimeryzacji). W wyniku rozpadu filamentów pośrednich, białka podjednostkowe są degradowane (nie są wtórnie wykorzystywane do syntezy filamentów, jak to jest w przypadku heterodimerów tubuliny lub G-aktyny). Aby wytworzyć włókno filamentu pośredniego, białka strukturalne muszą być syntetyzowane od nowa.
Mikrofilamenty zbudowane są polimerycznie z podjednostek G-aktyny (całe włókno nazywamy F-aktyną). Mikrofilamenty występują bardzo często w towarzystwie cząsteczek miozyny. Jako jedyne filamenty cytoszkieletu są elastyczne. Często łączą się w wiązki. Helikalność włókien aktynowych oznacza możliwość magazynowania energii (ciasny splot dwóch łańcuchów aktynowych - dużo energii; luźny splot - mało energii).
Sieć tensegralna („bierki połączone gumkami”) to sieć tworzona przez włókna cytoszkieletu, która tworzy trójwymiarową strukturę komórki (błona nie usztywnia komórki!!!). Obecność białek rozciągliwych (F-aktyna) i nierozciągliwych (mikrotubule, filamenty pośrednie) pozwala na wytworzenie tego typu sieci.
W szkielecie aktynowym mogą występować tzw. nadskręcenia. Jest to sytuacja analogiczna do nadskręceń występujących w kolistych cząsteczkach DNA u bakterii (zwijanie genoforu się w ósemki) - taka sytuacja następuje w wyniku np. wycięcia fragmentu cząsteczki DNA i wklejeniu go po obróceniu o 180°.
Funkcje cytoszkieletu w komórce roślinnej:
Generowania ruchu cytoplazmy (cykloza).
Reakcja grawitropijna (rejestracja kierunku działania grawitacji; biorą w tym udział amyloplasty statolitowe oraz mikrofilamenty).
Ruch organelli typu wici lub rzęsek.
Ruchy chromosomów wrzeciono kariokinetyczne).
Powstawanie polarności komórki (co jest ważne u komórek dzielących się, bo wyznacza kierunek podziału).
Kontrola kierunku podziałów komórkowych (zasada Errery - komórki zawsze dzielą się w miejscu; gdzie przyszła ściana komórkowa będzie miała najmniejszą powierzchnię).
Kierowanie procesem tworzenia ściany komórkowej.
Mikrofilamenty aktynowe
Białka oddziałujące z aktyną- ARP:
Inhibitory aktynowe- naturalne, produkowane przez grzyby, gąbki i rośliny w odpowiedzi na infekcje (jako broń chemiczna):
Latrunkulina-(Gąbki) blokujące podjednostki, uniemożliwia polimeryzację
Falloidyna-(grzyby) stabilizuje (usztywnia) MF, zapobiega depolimeryzacji-ważne w preparatyce, badaniach; zatrucia muchomorami sromotnikowymi;
Swinholid- fragmentuje MF
Cytochalazyna B-(grzyby) blokuje dodatni koniec MF
Reakcja grawitropijna.
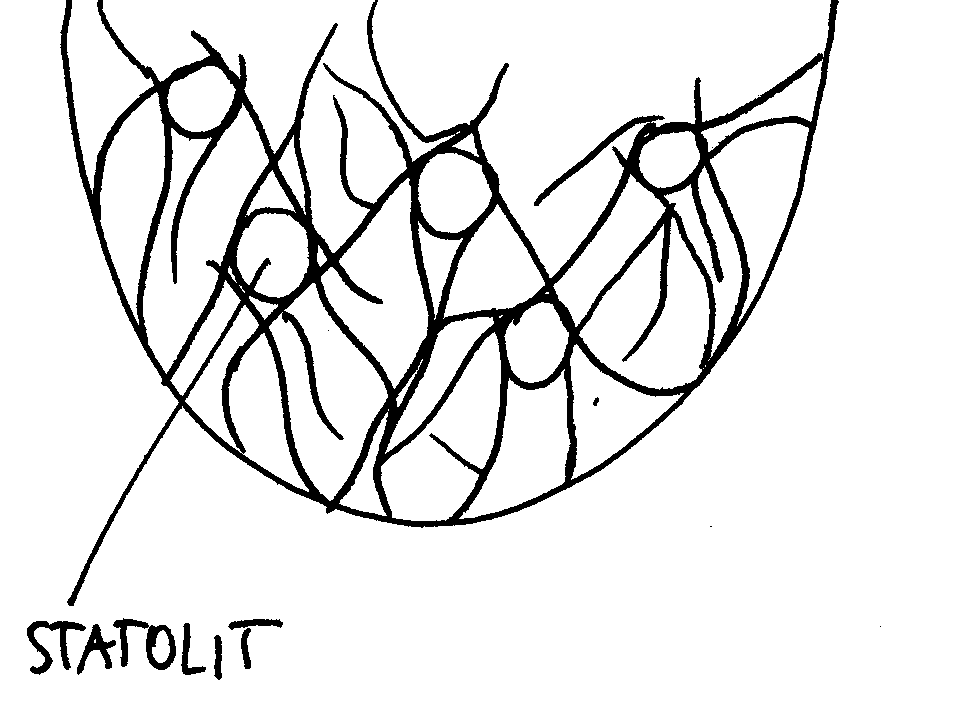
Za reakcję grawitropijną roślin odpowiada cytoszkielet; dzięki niej korzeń główny rośliny rośnie wgłąb ziemi. Odchylenie od pionu rejestrowane jest przez skrobię statolitową.
Już Darwin zaobserwował skrobię statolitową w komórkach czapeczki korzenia.
Za reakcję grawitropijną odpowiadają całe komórki (zwane statocytami), a nie skrobia (zmagazynowana w amyloplastach statolitowych).
Czapeczka korzeniowa pełni zatem rolę receptoru (statocyty ze statolitami); komórki czapeczki należą do jednego klonu (mają wspólne pochodzenie - ta sama komórka pierwotna).
Sygnał grawitropijny przechodzi od czapeczki do strefy wydłużeniowej korzenia; korzeń skręca, gdy komórki z jednej jego strony wydłużają się szybciej.
Chwytniki (ryzoidy) ramienic (np. Nitella sp. lub Chara sp.) również wykazują reakcję grawitropijną (więc można je nazwać statocytami). Ryzoid jest komórką o wzroście szczytowym (rośnie tylko wyróżniony fragment ściany). W ryzoidach występują tzw. ciężkie wakuole (wypełnione BaSO4), które spełniają funkcje statolitów.
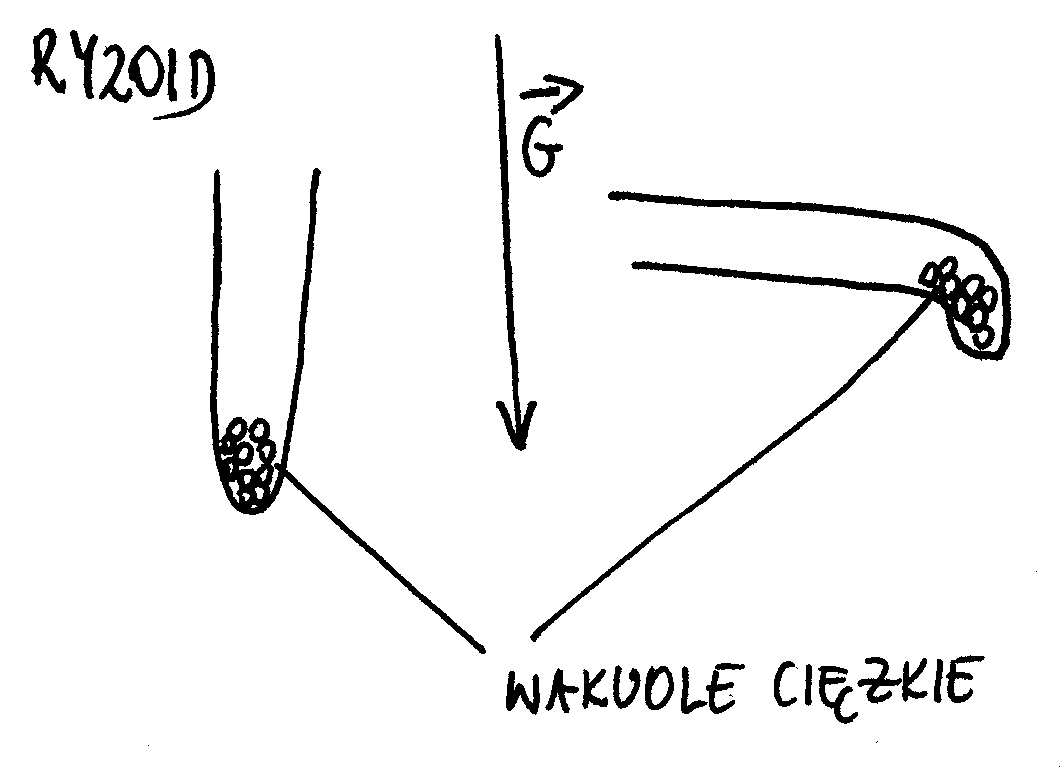
Rolą statolitów jest sedymentacja na powierzchni, która ma nie rosnąć (blokują one dopływ substancji transportowanych w pęcherzykach AG).
W wyniku zastosowania cytochalazyny B (trucizny mikrofilamentów) można upośledzić lub całkiem zahamować reakcję grawitropijną (doświadczenie to można wykonać np. na czapeczce korzeniowej Lepidium sp. taka sytuacja oznacza oczywiście, że w reakcji grawitropijnej bierze udział szkielet aktynowy (współpracujący ze statolitami).
Innymi przykładami reakcji grawitropijnej jest wzrost włośników lub łagiewki pyłkowej.
Zmiana położenia statolitów zmienia stan naprężenia włókienka aktynowego połączonego z plazmalemmą. Stąd właśnie zniszczenie szkieletu aktynowego powoduje osłabienie reakcji grawitropijnej. Zmiana naprężeń błony może wpływać na przepuszczalność kanałów jonowych.
U mutanta Arabidopsis sp., który nie wytwarza skrobi w amyloplastach reakcja grawitropijna zachodzi pomimo tego, że amyloplasty są lekkie.Bramkowanie kanałów jonowych zależy od:
czynników elektrycznych
czynników chemicznych
naprężenia błony (czynników fizycznych)
Czapeczka korzeniowa
Komórki czapeczki korzeniowej ulegają apoptozie (nie jest to zniszczenie mechaniczne, jak pierwotnie sądzono).
Wszystkie komórki czapeczki posiadają skrobię statolitową.
Zaledwie 2-3 komórki w części centralnej czapeczki mają istotny wpływ na reakcję grawitropijną. Pozostałe komórki tylko wzmacniają sygnały.
Korzenie boczne są niewrażliwe na grawitację (rosną w bok od korzenia głównego). Czynnikiem modyfikującym to zjawisko jest światło. Gdy światła brakuje, korzenie rosną płasko; gdy jest światło - wzrost pod kątem GSA (jest to kąt odpowiedzi grawitropijnej specyficzny dla gatunku).
W czapeczkach korzeni bocznych amyloplasty statolitowe są skupione w pobliżu jądra komórkowego (a nie na dnie komórki, jak jest to w czapeczce korzenia gównego).
Rola mikrotubul i szkieletu tubulinowego:
Mikrotubule są najgrubszymi (do 25 nm) włóknami białkowymi; są puste w środku. Jedną miktotubulę tworzy 13 protofilamentów zbudowanych z heterodimerów tubuliny. Heterodimery odkładane są po helisie. Helisa posiada koniec A (+) i D (-).
Mikrotubule powstają jako struktury polarne (ponieważ polarne są heterodimery: α(-); β(+)).
Cytoszkielet tubulinowy można stabilizować przy pomocy taxolu (preparat uzyskiwany z cisu; głownie z kory). Taxol blokuje podziały komórkowe, co jest wykorzystywane w leczeniu nowotworów (np. raka piersi).
Kolchicyna blokuje polimeryzację mikrotubul; dzięki kolchicynie można uzyskać komórki poliploidalne (chromosomy nie rozchodzą się w trakcie podziału). Kolchicyna pochodzi od Colchicum autumnae (ziemowit jesienny; kwiat podobny do krokusa; śmiertelnie trujący; krowy go nie jedzą).
γ-tubulina to forma tubuliny występująca w MTOC, układająca się w struktury przypominające z wyglądu sprężynkę. γ-tubulina daje początek nowym mikrotubulom.
Protofilamenty mogą być składane w płaszczyznę, która następnie zwija się w rurkę (sposób częsty w układach wieloelementowych jak dublety w wiciach lub triplety w centriolach). Należy pamiętać, że jedynie zwierzęta i glony (poza krasnorostami) posiadają centriole).
Mikrotubula określonej długości jest zbudowana ze stale wymieniających się heterodimerów (ciągła polimeryzacja i depolimeryzacja). Mikrotubule mogą ulegać wydłużaniu lub skracaniu.
Polimeryzacja polega na przyłączaniu heterodimerów związanych z GTP (guanozynotrójfosforan). Na szczycie mikrotubuli znajduje się „czapeczka” GTP; jeśli w pobliżu nie ma nowych heterodimerów do polimeryzacji, mikrotubula traci GTP i zaczyna się rozpad struktury.
Z powierzchnią mikrotubul oddziałują różnego rodzaju białka z grupy MAP, min. białka motoryczne:
dyneina (przemieszcza się do końca „-”)
kinezyna (przemieszcza się do końca „+”)
Białka motoryczne mogą pełnić funkcje transportowe (np. transportują pęcherzyki AG); podobnie jak miozyna w połączeniu z mikrofilamentami. Układy transportowe działają zawsze poza jądrem komórkowym (aby DNA nie uległo uszkodzeniu; gdy komórka nie miała otoczki jądrowej; transport stanowił zagrożenie dla tożsamości DNA).
Stabilnym przykładem działalności budulcowej mikrotubul są wici i rzęski. Wić ma strukturę uniwersalną (9 X 2 + 2); szkielet tubulinowy jest spięta neksyną (białko elastyczne - rozciągliwość nawet ośmiokrotna) i otoczony plazmalemmą. Za ruch zginający rdzenia odpowiada dyneina rzęskowa.
Klejnotka (Euglena) porusza się za pomocą wici, w której mikrotubule składają się teleskopowo dzięki wędrówce haczyków dyneinowych po ich powierzchni (kierunek wędrówki zmienia się okresowo). Zatem dyneina jest bardzo ważnym białkiem biorącym udział w ruchu wici.
Gamety męskie (nawet tak wysoko uorganizowanych roślin jak sagowce i miłorzęby) są ruchliwe (posiadają wić).
Geny kodujące ważne dla organizmu białka są konserwatywne ewolucyjnie (niewrażliwe na mutacje; wszelkie mutacje są letalne), gwarantuje to stałość budowy białka (np. tubuliny).
Dublety mikrotubul występujące obwodowo w szkielecie wici zachodzą na siebie, co umożliwia im ślizganie się (jedna po drugiej) podobnie jak miozyna z aktyną w sarkomerach.
Wici poruszają się ruchem falistym, a rzęski wahadłowym.
System błon wewnętrznych
Błona elementarna zewnętrzna (plazmolemma)
System endomembranowy (GERL)
Reticulum endoplazmatyczne (ER)
Diktiosomy (G)
Pęcherzyki egzo- i endocytowe
Lizosomy (L)
Pęcherzyki egzocytowe biorące udział w procesie sekrecji (wydzielania) łączą się z plazmalemmą w momencie wylewania zawartości poza komórkę (powoduje to wzrost powierzchni błony).
Odwrotną sytuacją jest fagocytoza (i inne formy endocytozy), która powoduje zmniejszenie powierzchni błony poprzez utworzenie pęcherzyka endocytowego.
Biorąc pod uwagę oba powyższe zjawiska, można stwierdzić, że w pewnym sensie pazmalemma jest składnikiem systemu endomembranowego.
Kompartymentacja - powstanie pęcherzyków z dwuwarstwy lipidowej (oddzielenie części środowiska poprzez zamknięcie go wewnątrz pęcherzyka).
System błon wewnętrznych tworzy platformę dla reakcji zachodzących w komórce (tam zachodzi obróbka produkowanych substancji).
Każdy fragment błony posiada specyficzne markery białkowe (dzięki temu można odróżnić stronę cytoplazmatyczną i cysternową). Wnętrze pęcherzyka (lub strona od cysterny) jest częścią środowiska zewnętrznego - przy egzocytozie ta właśnie strona kontaktuje się ze środowiskiem zewnętrznym.
Fosfolipidy:
Ze względu na polarność cząsteczek fosfolipidów, hydrofilowe główki skierowane są do środowiska wodnego, a hydrofobowe ogonki skierowane są w stronę tłuszczową (kontakt z ciałem tłuszczowym lub innymi ogonkami fosfolipidów - tworzenie miceli i dwuwarstw).
Dwuwarstwy lipidowe spontanicznie zamykają się w formę pęcherzyka (liposom).
Ze względu na hydrofobowy charakter wnętrza błony fosfolipidowej, jest ona nieprzepuszczalną barierą dla polarnych cząsteczek; za transport tych cząsteczek odpowiadają białka tworzące:
kanały (mogą być otwarte lub zamknięte; gdy są otwarte, następuje transport bierny; płyną tędy substancje, które nie łączą się z białkiem kanałowym)
przenośniki / transportery (przenoszona cząsteczka łączy się z przenośnikiem - transport aktywny)
Poryny:
Transmembranowe części białek z grupy poryn mają budowę nie α-helikalną, ale β-kartkową (zwaną β-beczułką; beczułka wypełniona jest wodą).
Białka te tworzą duże (przepuszczające cząsteczki do 600 D), niespecyficzne pory w błonach zewnętrznych bakterii, mitochondriów i plastydów.
Akwaporyny:
Specjalnym rodzajem kanałów membranowych są akwaporyny (AQPs). Są to kanały wodne ułatwiające bardzo szybki wpływ lub wypływ wody np. w erytrocytach (lub w tonoplaście komórki roślinnej podczas zmian turgoru).
Białka akwaporyn składają się z 6 transbłonowych segmentów α-helikalnych.
Por ma średnicę 0,3 nm, co oznacza, że w świetle kanału mieści się dokładnie jedna cząsteczka wody (single file passage). Mimo to szybkość przepływu sięga kilku mld. cząsteczek na sekundę.
GERL:
System endomembranowy ma związek z plazmalemmą oraz nukleolemmą, tonoplastem, błoną mitochondrialną itd.
Kształt ER nadają mu aktynowe pęczki (platformowanie ER przez aktynę). ER można obserwowac stosując barwnik fluorescencyjny DiOC6.
Ruch anterogradowy (do przodu) - pęcherzyki od ER łączą się ze stroną cis AG. W cysternach odbywa się dojrzewanie białek; od strony trans odrywają się pęcherzyki, które wędrują do plazmalemmy, wakuoli lub ruchem retrogradowym (do tyłu) do ER.
kierunku ruchu pęcherzyka decyduje etykieta białkowa (np. klatryna).
Ciała tłuszczowe w komórkach roślin oleistych powstają z SER jako pęcherzyki wypełnione trójglicerydami, otoczone jednowarstwą fosfolipidową naszpikowaną stabilizującymi „pinezkami” oleozynowymi.
Ciała białkowe tworzące się z RER w komórkach tkanki odżywczej (endosperm), tworzą się poprzez magazynowanie białek syntetyzowanych na rybosomach RER.
Każda błona systemu GERL jest asymetryczna (asymetria cis-trans).
Zasada transportu pęcherzykowego gwarantuje zachowanie orientacji błon.
Ciała białkowe to magazyny białka otoczone dwuwarstwą lipidową; ciała olejowe otoczone są jednowarstwą.
Ruch od ER do AG nazywamy anterogradowym, a odwrotny - retrogradowym.
Polisomy (wiele rybosomów połączonych razem) to główna postać, w jakiej występuja rybosomy na RER.
Pęcherzyki oderwane od ER zawierające białko wędrują dalej do AG, gdzie białko dojrzewa (dołączane są reszty cukrowe, lipidowe lub inne; zmienia się też struktura przestrzenna białka).
Ekstensyna transportowana jest pęcherzykowo do ściany komórkowej.
Istnieje możliwość zmiany liczby diktiosomów (mogą się rozrywać).
Przy użyciu izotopów radioaktywnych można śledzić obieg substancji wewnątrz systemu GERL.
Przekazywanie zawartości pomiędzy cysternami następuje na drodze pęcherzykowej. Możliwe jednak, że transport może odbywać się na zasadzie dynamiki całego układu; nowe pęcherzyki tworzą nową cysternę i całość przesuwa się; na stronie trans cysterna ulega rozpadowi. Taka sytuacja oznaczałby, że zawartość pęcherzyków w zasadzie nie zmienia lokalizacji (nie przechodzi pomiędzy cysternami) - można to porównać do mikrotubuli, która budowana jest z coraz to nowych heterodimerów.
Białka receptorowe umieszczone na plazmalemmie muszą być transportowane w specyficzny sposób (bo muszą być odpowiednio zorientowane na powierzchni błony komórkowej.
Kierunek wędrówki pęcherzyków zależy od etykiet białkowych na powierzchni pęcherzyków. Białka z grupy SNARE rozpoznają miejsce, do którego ma trafić pęcherzyk.
V-SNARE (vesicule SNARE) - białko na pęcherzyku; rozpoznaje błonę docelową
T-SNARE (target SNARE) - białko rozpoznające białka V-SNARE
Połączenie V-SNARE i T-SNARE powoduje zlanie się obu błon w miejscu połączenia.
Lizosomy
Markerem lizosomów jest kwaśna fosfataza (jeden z enzymów wewnątrz liposomu).
Lizosomy są szczególnymi pęcherzykami odpączkowującymi od strony trans AG.
Zawierają hydrolazy; ich funkcją jest degradacja makrocząsteczek wchłoniętych do komórki.
Nowo syntetyzowane enzymy są znakowane specjalną resztą cukrową (tak, aby mogły być związane do błony przy pomocy specyficznego receptora), we wnętrzu AG enzym łączony jest z przyszłą błoną lizosomu. Dopóki jest on związany z błoną, nie ma właściwości hydrolitycznych. Gdy lizosom odrywa się od AG, następuje aktywacja pomp protonowych (zakwaszanie środowiska wewnątrz lizosomu), białko enzymatyczne odłącza się od receptora błonowego i następuje podział lizosomu na dwie części (jedna zawiera enzym, a druga receptor, który wędruje do AG - taki komórkowy recycling :P)
Białka z grupy COP
COP1 - odpowiada za transport retrogradowy (od AG do ER)
COP2 - odpowiada za transport anterogradowy
Endocytoza receptorowa:
Pęcherzyki sekrecyjne docierające do błony komórkowej wbudowują swoją błonę do plazmalemmy (powiększenie powierzchni). Aby błona nie pękła, równolegle biegną procesy odwrotne (endocytoza). Oba te procesy zapewniają ciągły ruch systemu GERL.
Proces endocytozy prowadzony jest tylko na niektórych obszarach błony.
Połączenie się ligandu z receptorem powoduje rozpoczęcie wpuklania błony; bierze w tym udział białko markerowe - klatryna.
Triskeliony klatryny budują kosz klatrynowy wokół pęcherzyka.
Przewężenie szyjki dołka opłaszczonego powodujeinne białko - dynamina.
Kosz klatrynowy zapobiega pęknięciu pęcherzyka w cytoplazmie.
Z pęcherzykiem endosomalnym łączy się lizosom dając endosom wtórny, który traci klatkę klatrynową..
Klatka klatrynowa jest podobna do struktury węgla C60 (fullerenu) - piłka złożona z 12 pentagonów zamykających heksagony w kulę.
Klatryna jest białkiem zbudowanym z 6 łańcuchów (struktura 4-rzędowa); ramiona triskelionów są elastyczne (mogą tworzyć 5 lub 6-kątne elementy).
Mikrosomy
Otoczone są pojedynczą błoną elementarną.
Często zawierają inkluzje krystalicznego białka.
Wyróżniamy:
Peroksysomy: liściowe, glioksysomy, brodawek korzeniowych
Sferosomy - rezerwy energetyczne komórek (silnie załamują światło)
Hydrogenosomy - prowadzą oksydację pirogronianu
Glikosomy - katalizują beztlenowy rozkłąd glukozy
Peroksysomy
Peroksysomy są bardzo podobne do lizosomów (bark różnic strukturalnych).
Białka peroksysomów mają skłonność do krystalizacji.
Peroksysomy mogą mieć różny charakter. Peroksysomy liściowe utylizują tlen powstały w procesie fotosyntezy; w komórkach brodawek korzeniowych obrabiają związki azotowe.
U zwierząt peroksysomy występują w dużych ilościach w wątrobie i nerkach; u glonów i roślin - w komórkach prowadzących fotosyntezę.
Markerem peroksysomów jest rozkładająca nadtlenek wodoru katalaza (stanowiąca do 15% składy białkowego peroksysomu). Występuje ona wraz z licznymi oksydazami generującymi nadtlenek wodoru; w ten sposób komórka jest odizolowana od toksycznego tlenu i wody utlenionej.
Peroksysomy wspomagają pracę mitochondriów.
Triton WR-1339 - detergent akumulujący się w lizosomach i zwiększający ich ciężar.
Peroksysomy różnią się od lizosomów wrażliwością na inny detergent - digitoninę; aby wyzwolić katalazę z peroksysomu trzeba użyć około 10 razy więcej digitoniny niż jest potrzebne do wyzwolenia kwaśniej fosfatazy z lizosomu. Gdyby obie substancje były w takich samych pęcherzykach, zostałyby uwolnione przy tym samym stężeniu detergentu.
Jeśli peroksysomy zawierają rdzeń krystaliczny, łatwo je odróżnić na zdjęciach TEM; jeśli nie mają rdzenia, stosuje się test DAB (reakcję z diaminobenzydyną).
W reakcji DAB, po utlenieniu diaminobenzydyny przez katalazę, powstaje polimer łączący się z czterotlenkiem osmu (standardowa substancja kontrastująca)
W komórkach roślin rdzeń krystaliczny peroksysomu tworzy katalaza (marker peroksysomów).
W komórkach zwierząt rdzeń peroksysomu tworzy oksydaza moczanowa (nieregularny kryształ).
Urikaza - oksydaza moczanowa przeprowadza w peroksysomach oksydację produktów przemiany kwasów nukleinowych (puryny) i niektórych białek.
Wszystkie peroksysomy mają zdolność rozkładania H2O2 za pomocą katalazy.
H2O2 -> 0,5 O2 + H2O
Funkcje peroksysomów:
Metabolizm nadtlenku wodoru.
Detoksykacja komórki (utlenianie różnych substancji).
Oksydacja kwasów tłuszczowych.
Metabolizm związków azotu.
Katabolizm substancji niezwykłych np. D-aminokwasów albo ksenobiotyków (alkany), np. teflonu.
U grzybów w peroksysomach znajdują się enzymy pozwalające na rozkład krótkołańcuchowych węglowodorów. Może się to przyczyniać do oczyszczania wycieków ropy.
Glioksysomy
W komórkach roślinnych specjalne peroksysomy zwane glioksysomami przeprowadzają konwersję lipidów w węglowodany podczas kiełkowania nasion roślin oleistych (soja, orzeszki ziemne, słonecznik), gdzie materiałem zapasowym w komórkach liścieni lub endospermu są kropelki tłuszczu (trójglicerydy). Dzieje się to przy udziale typowo roślinnych enzymów.
Glioksysomy są nietrwałe; czynne w czasie kiełkowania siewki (1-2 tygodnie), następnie przekształcają się w peroksysomy liściowe. Spektrum enzymów zmienia się w ontogenezie.
Peroksysomy brodawek korzeniowych
W peroksysomach brodawek korzeniowych znajdują się aminotransferazy; dzięki czemu może tam zachodzić degradacja lub tworzenie aminokwasów.
Biogeneza peroksysomów
Powstają poprzez podział (są uważane za najstarsze endosymbionty komórki eukariotycznej; oksydaza D-aminokwasowa jest być może dowodem tego, że peroksysomy są najstarszymi endosymbiontami).
Białka peroksysomowe są dostarczane posttranslacyjnie, ich transport do wewnątrz odbywa się z użyciem energii z ATP.
Białka peroksysomowe są syntetyzowane na rybosomach cytoplazmatycznych; muszą być zaopatrzone w sekwencję sygnałową SKL zbudowaną z trzech aminokwasów.
S - small uncharged (mały, nienaładowany) - seryna, prolina, alanina
K - kation charged (+) - lizyna, arginina
L - lipidlike (hydrofobowy) - metionina, leucyna
Pozbawienie sygnału SKL powoduje pozostanie białka w cytozolu, natomiast dołączenie sygnału SKL do obcego białka pozwala wprowadzić je do peroksysomu (uniwersalizm SKL polega na tym, że można tą drogą wprowadzać bardzo różne białka; podany tutaj przykład z lucyferazą, świetlikami i transgenicznym tytoniem
Adrenoleukodystrofia - defektywne białko integralne błony peroksysomu nie pozwala na transport do wnętrza kwasów tłuszczowych, które nagromadzając się w płynach ustrojowych niszczą osłonki mielinowe nerwów.
Ćwiczenia III - Podział komórkowy. Ściana komórkowa.
Cykl komórkowy - od podziału do podziału, wyróżnia się 4 fazy:
#Faza G1
- komórka jest mała, następuje jej wzrost
- w tej fazie najłatwiej jest zahamować, spowolnić czy przyśpieszyć cykl
- tu występuje translacja i transkrypcja
CDK + cykliny G1 = kompleks umożliwiający przejście do fazy S
#Faza S
- tutaj następuje replikacja materiału genetycznego (semikonserwatywnie) przez aparat replikacyjny (ligazy, helikazy), zaczyna się w konkretnym miejscu,
- tworzone są białka histonowe, kohezyjny które łączą ze sobą 2 siostrzane chromatydy
- kończy się gdy mat. genetyczny zostaje zreplikowany
Kinaza + cyklina A = kompleks zatrzymujący replikację i umożliwiający tym samym wejście w następną fazę
#Faza G2
- synteza białek wrzeciona podziałowego (mikrotubul i filamentów aktynowych)
- pod koniec fazy uaktywnia się kinaza fazy M i powoduję fosforylację i defosforylację białek -rozpoczęcie mitozy
Cykliny mitotyczne + kinaza = przejście do fazy M
#MITOZA:
- Kariokineza:
1. Profaza:
- kondensacja chromatyny - powstają chromosomy mitotyczne poprzez fosforylację histonu H1, H2A, H3 (przez kinazę fazy M)
- po obu stronach centromeru tworzy się kinetochor który ma strukturę blaszkową (miejsce wiązania mikrotubul wrzeciona)
- depolimeryzacja mikrotubul kortykalnych - z nich powstaje wrzeciono kariokinetyczne:
* MT biegunowe
* MT kinetochorowe (łączą się z kinetochorem - liczba MT łączących się z kinetochorem jest różna)
2. Prometafaza:
- otoczka jądrowa ulega fragmentacji - zapoczątkowuje to aktywacja kinazy fazy M (fosforyzuje i zmienia konformację białek blaszki)
- zanikanie jąderka przez udział nukleolin.
3. Metafaza:
- kończy się kondensacja chromatyny
- chromosomy ustawiają się w płaszczyźnie równikowej, stała pozycja
- składniki cytoplazmy rozdzielają się po równo do biegunów komórki
4. Anafaza:
- nagłe rozdzielenie chromatyd siostrzanych w miejscu centromerów
- jest kilka teorii ruchu tych chromatyd:
* ruch ślizgowy - polega na ślizganiu się mikrotubul biegunowych względem siebie (podobny ruch MT jaki występuje w wiciach)
* mechanizm kieratowy - polega na wydłużaniu mikrotubul, z przodu następuje polimeryzacja a na tylnej części depolimeryzacja przy czym jest ona wolniejsza.
* mechanizm zamka błyskawicznego - występuje tutaj boczne oddziaływanie między MT biegunowymi a MT kinetochorowymi.
* teoria kołnierza ślizgowego - kinetochor ślizga się po MT-ach
5. Telofaza:
- po doprowadzeniu chromatyd następuje dekondensacja chromatyny
- synteza rRNA jest wstępem do odtwarzania jąderka
- reorganizacja cytoszkieletu komórki
- wrzeciono zanika , rekonstrukcja jądra
- Cytokineza:
Z wrzeciona kariokinetycznego powstaje tutaj wrzeciono cytokinetyczne (ono tylko inicjuje podział)
Powstaje pierścień kurczliwy (opaska preprofazowa)
- składa się z miozyny i aktyny
- zaciska się podobnym mechanizmem jak u nas w sarkomerze
- filamenty się nie tworzą od nowa tylko reorganizują
- obkurczając się tworzy się bruzda podziałowa
ER wysyła pęcherzyki do centrum (centryfugalnie) - sekrecja różnych subst. (pektyn)
Tworzy się przegroda pierwotna
- w niej są pektyny (półpłynne)
- rozrasta się na boki
- po styknięciu się z oryginalną ścianą, masa ta gęstnieje i tworzy się blaszka środkowa
Tworzona jest następnie ściana pierwotna (elastyczna)
Zanika fragmoplast
Mikrotubule odbudowywują cytoszkielet
Jest hamowana przez cytochalazynę B (powstaje komórka dwujądrowa, nie wstrzymuje kariokinezy)
Ruch chromosomów:
Mikrotubule odpowiedzialne są również za ruch chromosomów - mikrotubule dynamiczne polimeryzujące w MTOC.
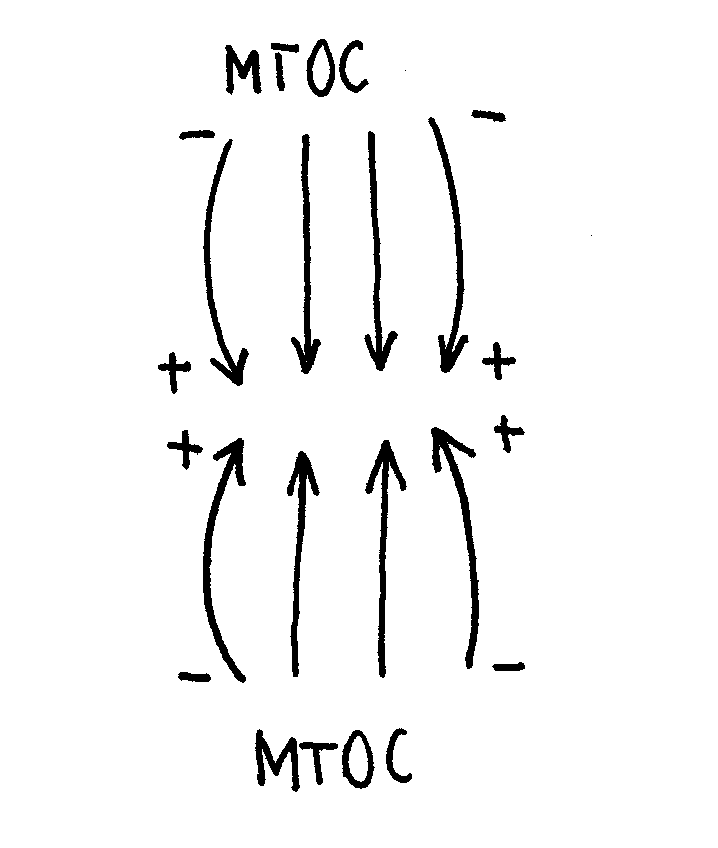
Przewężenie pierwotne chromosomu (centromer) otoczony jest białkami kinetochorowymi, które mają zdolność łączenia się z mikrotubulami. Białka kinetochorowe można wykrywać (tak jak inne białka) metodą immunocytochemiczną.
Złapanie chromosomu (połączenie się MT z kinetochorem) przed ustawieniem w płaszczyźnie równikowej nie powoduje przerwania ani zmiany w podziale; sytuacja taka prowadzi jedynie do dodatkowej polimeryzacji mikrotubuli i właściwego ustawienia chromosomu.
Dzięki połączeniu chromosomu z mikrotubulami (różną ilością) z przeciwnych biegunów, w kinetochorze tworzy się silne naprężenie. Anafaza trwa bardzo krótko (chromatydy bardzo szybko się rozchodzą).
Mechanizm ściągania chromosomów do biegunów komórki:
Hipoteza pęczków mikrotubul (odciąganie odbywa się na zasadzie składania teleskopowego, analogicznie do sytuacji w obwodowych dubletach MT w wiciach).
Eksperyment polegający na laserowym przecięciu połowy wrzeciona w momencie największego naprężenia udowadnia, że mikrotubula (ta nie potraktowana laserem) działa jak gumka i ściąga cały chromosom do jednego z biegunów). Można zatem dojść do wniosku, że istnieje elastyczne białko, na którym umieszczone są mikrotubule wrzeciona.
p34 - kinaza występująca przy podziale/cyklu komórki
p34 + cyklina mitotyczna (G2) = MPF
MPF (M-phase promoting factor) - kompleks wprowadzający do fazy M
p34 + cyklina mitotyczna (G1) = SPF
MPF i SPF odpowiadają za fosforylację odmiennych substratów
Wrzeciono cytokinetyczne:
Mikrotubule wrzeciona cytokinetycznego zdają się być niezbędne do przemieszczania pęcherzyków AG (wypełnionych pektynami) do płaszczyzny podziału, gdzie tworzy się pektynowa przegroda pierwotna.
Najnowsze badania obalają jednak ten pożałowania godny mit , bo zniszczenie mikrotubul wrzeciona cytokinetycznego nie powoduje zahamowania tworzenia się przegrody.
Wrzeciono cytokinetyczne posiada inną odporność i właściwości niż wrzeciono kariokinetyczne (odpowiadają za to białka z grupy MAP).
Cytokineza może zachodzić na trzech drogach:
Gdy wrzeciono kariokinetyczne nie rozpadnie się, tylko zmieni nieco kształt (tzw. fragmoplast); przegroda pierwotna rośnie centyfugalnie (odśrodkowo).
Gdy wrzeciono kariokinetyczne nie rozpadnie się, ale nie zmieni nawet swego kształtu; przegroda pierwotna rośnie centrypetalnie (dośrodkowo).
Gdy wrzeciono kariokinetyczne rozpadnie się, a mikrotubule ustawią się równolegle do płaszczyzny podziału (tzw. fikoplast); jądra komórkowe zbliżają się do siebie, ale nie wiem, w którą stronę następuje wzrost przegrody pierwotnej.
Ułożony jest on tuż pod plazmalemmą; mikrotubule są często połączone z błoną za pomocą transbłonowych łączników.
Cytoszkielet kortykalny jest najlepiej widoczny w czasie interfazy; układ mikrotubul jest często wysoce uporządkowany (są ułożone równolegle do siebie).
Układ mikrotubul kortykalnych wskazuje oś polarności komórki (jest do niej prostopadły).
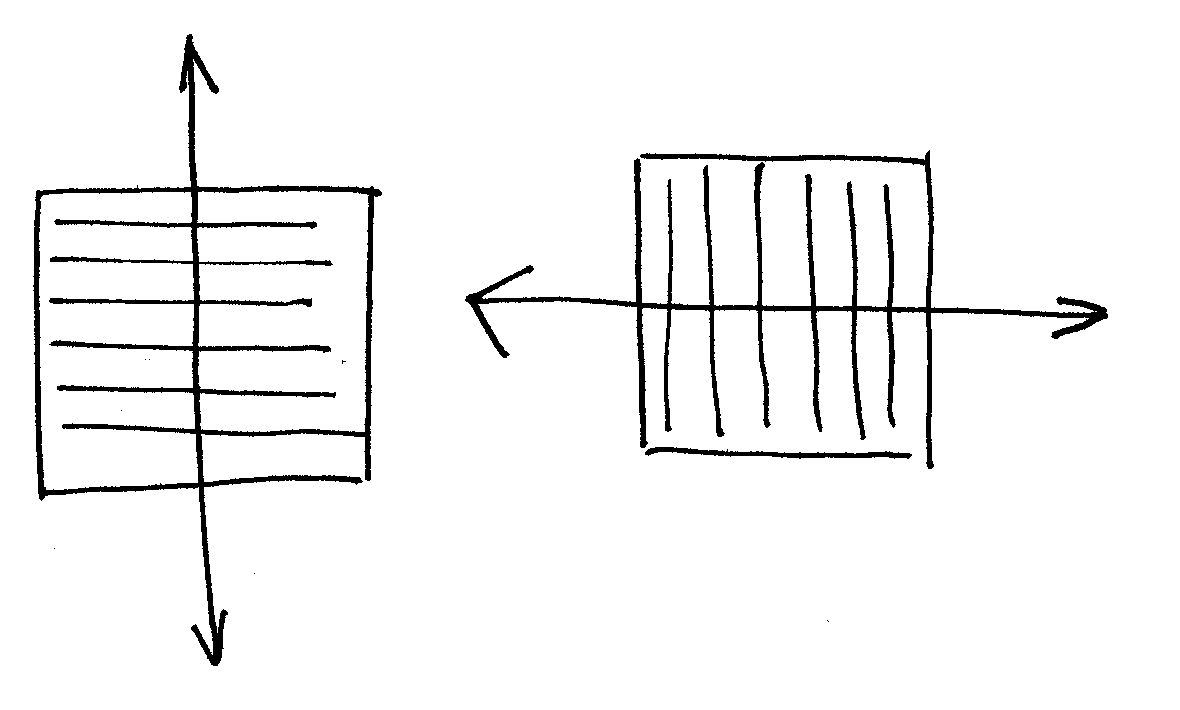
Roślina może kontrolować polarność komórek regulując stężenie hormonów roślinnych (drobnocząsteczkowych); można to udowodnić przedstawiając choćby doświadczenie wykonane na hypokotylu owsa.układ mikrotubul poziomy (więc oś polarności pionowa) - gdy traktujemy komórki gibereliną GA3 (promocja wydłużania).
układ mikrotubul pionowy (przebudowa o 90°) - gdy potraktujemy komórki etylenem
Na merystemach apikalnych, w komórkach rosnących izometrycznie (równomiernie we wszystkie strony), układ mikrotubul kortykalnych jest sieciowy (nieuporządkowany); również sieciowy jest układ mikrofibrylli celulozowych ściany.
Intrususcepcja - wzrost poprzez rozciąganie turgorowe (np. wzrost ścian pierwotnych w komórkach rosnących izometrycznie.
Na bocznej powierzchni merystemów zachodzi już wzrost spolaryzowany.
Mikrofibrylle celulozowe tworzą się równolegle do układu mikrotubul kortykalnych; dzięki takiemu ułożeniu turgor może rozciągać komórkę tylko w jedną stronę.
W komórkach podlegających ukierunkowanemu wzrostowi (w organach osiowych) układ mikrotubul kortykalnych jest wysoce uporządkowany (są równoległe do siebie).
W preprofazie cytoszkielet ulega depolimeryzacji; w fazie tej mikrotubule kortykalne ulegają „collapsowi”, tworzy się opaska preprofazowa.
W szkielecie kortykalnym interfazowym mikrotubule są oddalone o około 25 nm. Pod wpływem „collapsu” mikrotubule z biegunów „zsypują” się na środek.
Opaska preprofazowa (PPB - preprophase band) nie występuje np. w komórkach kambium, mimo że są to komórki z definicji dzielące się (jednak to, że nikt tego nie zobaczył, wcale nie znaczy, że tego nie ma).
W komórkach kamforowca układ mikrotubul kortykalnych w interfazie jest helikalny; nie znaleziono opasek preprofazowych w komórkach kamforowca (istnieje hipoteza, że opaska może być tworzona przez pojedyncze mikrotubule, dlatego nie widać jej pod mikroskopem.
Położenie opaski wyznacza płaszczyznę podziału i położenie przegrody pierwotnej (jest odzwierciedlaniem polarności komórki).
Opaska całkowicie depolimeryzuje z chwilą wejścia komórki w profazę (dzięki temu zwiększa się pula heterodimerów potrzebnych do syntezy wrzeciona).
Aparat szparkowy u cebuli powstaje prostopadle do opaski preprofazowej.
Położenie opaski preprofazowej jest „zapamiętywane”; nawet po wirowaniu przegroda pierwotna tworzy się w miejscu wyznaczanym wcześniej przez opaskę.
Do tworzenia wrzeciona cytokinetycznego używane są heterodimery, które wcześniej tworzyły wrzeciono kariokinetyczne.
Ułożenie mikrotubul kortykalnych jest związane z ułożeniem mikrofibrylli celulozowych (ściana powtarza układ mikrotubul). Zależność ta zaobserwowana została w dojrzewającym protoksylemie.
Na powierzchni błony komórkowej istnieją enzymy (syntazy celulozowe - w postaci 6-promiennych rozet), które syntetyzują łańcuch celulozowy z glukozy dostarczanej przez pęcherzyki AG.
Syntazy celulozowe nie zawsze tworzą 6-promienną rozetę; czasem występuje linearny system syntaz (np. u krasnorostów); taki układ syntaz daje tasiemkowaty układ mikrofibrylli celulozowych.
Na powierzchni komórek niektórych zielenic występują układy heksagonalne zawierające wiele 6-promiennych rozet syntaz celulozowych. Może to sugerować, że kompleks taki syntetyzuje mikrofibrylle w różnych kierunkach (bo poszczególne warstwy ściany są ułożone w różnych kierunkach).
U roślin wyższych celuloza powstaje na zewnątrz komórki. U niektórych glonów jednak celuloza powstaje w cysternach AG; tworzone są tam łuseczki celulozowe, które transportowane są na zewnątrz komórki.
Blaszka środkowa
Ściana pierwotna z pierwotnymi polami jamkowymi, cienka, rozciągliwa, powstaje w czasie cytokinezy, rośnie wraz z komórką. Safranina + błękit cyjaninowy barwi celulozę na niebieskawy kolor. Po ukończeniu formowania powstaje:
Ściana wtórna (warstwy S1, S2 i S3) z jamkami prostymi lub lejkowatymi, ulega drewnieniu, odkładana jest tutaj lignina która barwi się na kolor czerwony poprzez dodanie safraniny
System połączonych ścian komórkowych nazywamy apoplastem.
Utrzymywanie komórki w stanie turgoru (bez ściany komórka by pękła).
Określenie właściwości mechanicznych tkanek (często wraz z wakuolą).
Regulacja kierunków wzrostu komórki.
Zabezpieczenie przed czynnikami środowiska, oraz aktywne zabezpieczenie przed patogenami.
Udział w przenoszeniu / odbiorze sygnałów - narzuca drobno cząsteczkowość sygnałów roślinnych; hormony (jak auksyny) wędrują w ścianie jak neurotransmitery pomiędzy neuronami.
Kontakt apoplastu z symplastem (komórki transferowe) - tam, gdzie protoplast dotyka ściany.
składniki szkieletowe celuloza
macierz ściany hemicelulozy, pektyny, glikoproteiny
substancje inkrustujące lignina, krzemionka, węglan wapnia, tiaminy, żywice
substancje adkrustujące suberyna, kutyna, sporoplenina, śluzy, gumy, kaloza (buduje glukan, wiązanie beta)
Składniki inkrustujące wnikają pomiędzy elementy strukturalne ściany; lignina umożliwia utworzenie tkanki waskularnej.
Substancje adkrustujące tworzą warstwę na powierzchni; sporoplenina nie ulega biodegradacji.
Hemicelulozy przyklejone są do mikrofibrylli celulozowych. Z hemicelulozami związane są neutralne pektyny, a z nimi pektyny kwaśne.
Glikoproteiny to białka elastyczne przeplecione przez mikrofilamenty.
F-aktyna (mikrofilamenty)
neksyna (spina dublety w szkielecie wici)
ekstensyny (przepleciona przez mikrofibrylle; pełni funkcję strukturalną; zawiera dużo proliny i hydroksyproliny - jak w kolagenie).
Zawartość białek w ścianie komórkowej zależy od stanu dojrzałości komórki, typu ściany, jej stymulacji (zranienia, atak patogenów itp.)
Rodzaje białek w ścianie (wartość procentowa określa zawartość procentową cukru w glikoproteinie):
HRGP (55%; bogate w hydroksyprolinę; występują we floemie, kambium; inaczej nazywane ekstensynami).
PRP (0-20%; bogate w prolinę; występują w ksylemie, włóknach i korze pierwotnej).
GRP (0%; bogate w glicynę, występują w ksylemie).
AGP (90%; bogate w galaktozę i arabinozę; istotne w adhezji i rozpoznawaniu komórek np. prowadzenie łagiewki pyłkowej przez tkanki słupka).
Udział w budowaniu ściany (ze składników dostarczanych przez komórkę).
Ekspansyny - odpowiadają za wzrost powierzchni ściany oraz rozpad ściany w trakcie maceracji i zrzucania organów, pozwalają na rozluźnienie struktury ściany komórkowej (ich działalność przejawia się np. powstawaniem bocznych struktur na powierzchni merystemu.
Ekstensyny to białka elastyczne spinające mikrofibrylle celulozowe w ścianie komórkowej, nie można ich mylić z ekspansynami.
W ścianie znajdują się enzymy szlaku syntezy ligniny, a także degradujące ścianę.
Komórki posiadające ściany komórkowe posiadają receptory (w ścianie rzecz jasna), które rozpoznają komórki sąsiednie.
Plastydy powstają wskutek różnicowania się form prekursorowych - proplastydów (w tkankach merystematycznych)
Namnażanie przez podział
Różnicowanie się proplastydów następuje tkankowo, przez udział światła - prowadzi do powstania form dojrzałych:
Ostatnią fazą jest faza starzenia
Mogą się odróżnicowywać w inne typy plastydów (np. chromoplast w chloroplast)
Każdy plastyd ma: podwójną błonę, zachowują własne DNA, posiadają macierz - stromę.
Genom plastydowy jest pojedynczą cząsteczką DNA, znajdują się po kilkanaście w pojedynczym obszarze nukleoidopodobnym
Białko PEND - białko które odpowiada za replikację, segregację i transkrypcję DNA
W otoczkach błonowych znajdują się unikalny skład lipidowy: znaczna ilość sulfolipidów i galaktolipidów przy niskim poziomie fosfolipidów.
Podział następuje poprzez mechanizm rozszczepienia
kształt sferyczny
niewielka ilość rybosomów
jeden lub kilka obszarów nukleoidopodobnych (ON)
pojedyncze ziarna skrobi i plastoglobule (struktury osmofilne zawierające estrowe pochodne karotenoidów, plastochinony i kwasy tłuszczowe)
słabo rozwinięty system błon, rurkowe struktury przypominające MT
Ciała kształtu soczewkowatego, zbudowane z otoczki (2 błonowa), stromy oraz silnie rozbudowanego systemu spłaszczonych obłonionych pęcherzyków nazywanych tylakoidami - część może łączyć się w stosy zwane granami
Pochodzą z proplastydów
Etioplasty są charakterystyczne poprzez posiadanie ciał prolamellarnych, które stanowią silnie rozbudowaną przestrzennie parakrystaliczną sieć błonowych rurek (cztero lub cześcio ramienna jednostka rurkowa (rurku :D nie wkurwiaj mnie, dobrze?) łącząc się ze sobą ramionami, jednostki te tworzą sieć przestrzenną, błony tubulin zawierają oksydoreduktazy NADPH - enzym ten katalizuje zależną od światła redukcję protochlorofilidu do chlorofilidu - prekursora chlorofilu.
Etioplasty mogą przekształcić się w chloroplasty poprzez ekspozycję na światło (protochlorofilid przekształca się w chlorofili a ten w chlorofil) następnie ulega zmianom struktura wewnętrzna tego plastydu tworząc dojrzały chloroplast, taki sam jak ten nieprzerwany epizodem ciemności.
Chloroplasty to jedyne plastydy zawierające chlorofil i prowadzące fotosyntezę, której istotą jest wykorzystanie energii świetlnej do redukcji pobieranego z atmosfery CO2 do poziomu cukrów. Reakcja przebiega ta w 2 fazach:
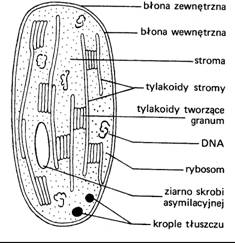
Tylakoid - składa się z błony tylakoidowej oraz światła (wnętrza tylakoidu), stos tylakoidów tworzy grana. Wyróżnia się dwa typy:W nich znajdują się barwniki - najpowszechniejszy chlorofil (a, b, c, d) oraz inne barwniki (karoteny, fikobiliny i ksantofile).
Stroma - zawiera liczne struktury zawieszone w substancji podstawowej:
Otoczka - charakterystyczna jest u chloroplastów ze względu na przebieg wielu skomplikowanych i ważnych procesów. Znajduję się tu CFC (centra formujące chlorofil - są to kompleksy wieloenzymatyczne otoczki)
Plastyd nie zawierający chlorofilu, kształt sferyczny, otoczka 2 błonowa, wnętrze wypełnia stroma (w niej ON - obszary nukleoidopodobne, rybosomy i plastoblobule a także najbardziej istotne ziarna skrobi)
Pojedynczy amyloplast może zawierać pojedyncze duże ziarno lub kilka mniejszych.
Ziarna powstają poprzez odkładanie (apozycję) skrobii wokół niednego lub kilku tzw. ośrodków znajdujących się w stromie.
Ziarna można podzielić na: proste (jeden ośrodek) i złożone (kilka takich ośrodków) rzdko występują półzłożone (ośrodki których warstwy odkładające są wspólne)
Apozycja skrobi niejednakowo intensywna prowadzi do powstania ziaren o uwarstwieniu ekscentrycznym (np. u ziemniaka), podczas gdy równomierne odkładanie warstw nazywa się koncentrycznym ułożeniem (np. miękisz liścieni roślin motylkowych)
W pełni rozwinięte amyloplasty spotyka się tylko w tkankach spichrzowych u roślin lub czapeczki gdzie pełnią wtedy funkcje statolitu, percepcja bodźca grawitacyjnego.
Plastydy nie zawierające chlorofilu, rozpowszechnione w tkankach roślinnych, zazwyczaj znajdują się w sąsiedztwie jądra komórkowego, otoczka 2 błonowa, wnętrze wypełnia stroma z niewielką ilością plastoglobuli i ON, nie zawierają rybosomów.
Występują przedewszystkim w komórkach kory pierwotnej łodygi, korzenia, epidermie i jej wytworach (włośniki), gruczołach wydzielających olejki eteryczne, i bielmie.
Słabo poznane funkcje, oprócz tego że udowodniono ich rolę w wytwarzaniu olejków eterycznych w młodych owocach cytrusowych.
Zawierają znaczą ilość karotenoidów, mimo tego nieaktywny fotosyntetycznie - brak chlorofilu. Kształtu sferycznego, elipsoidalnego bądź ameboidalnego, powstają na wskutek przekształcenia najczęściej chloroplastów lub amyloplastów
W pełni rozwinięte chromoplasty znajdują się przedewszystkim w płatkach korony oraz w owocach, przy współudziale antocyjan znajdujących się w wakuoli nadają różnorodne żywe barwy.
Typowo otoczka 2 błonowa, wnętrze wypełnia stroma oraz struktury magazynujące karotenoidy, jest tu ON, plastoglobule, bardzo mała ilość rybosomów oraz pokłady fitoferrytyny.
Wyróżnia się kilka typów tych plastydów:
Białkami swoistymi dla chromoplastów jest fibryllina (wyizolowana z papryki), białko CHRA, CHRB, CHRC
Są ostatnim ogniwem ciągu biogenetycznych powiązań między plastydami, mogą niekiedy przekształcać się w aktywnie fotosyntezujące chloroplasty.
Cytoszkielet tubulinowy kortykalny
Polarność komórki
Mikrotubule kortykalne, a układ mikrofibrylli w ścianie:
Ułożenie cytoszkieletu kortykalnego a mikrofibrylle celulozowe
Układ syntaz celulozowych:
Ściana komórkowa
Funkcje ściany komórkowej:
Sklerenchyma posiada grubą ścianę komórkową, która zapewnia sztywność nawet w warunkach stresu wodnego (np. u kserofitów).
Klasyfikacja składników strukturalnych ściany komórkowej:
Białka elastyczne występujące w komórkach roślinnych:
Białka strukturalne ściany:
Białka enzymatyczne:
Droga transportu ekspansyny jest zawsze drogą najkrótszą; transkrypt wędruje do najbliższego diktiosomu, gdzie dojrzewa. Nie wiadomo skąd komórka wie, że dana droga jest najkrótsza i że daną ścianę należy modyfikować ekspansyną.
w środowisku kwaśnym oddzielają hemicelulozy od mikrofibrylli (rozkładają wiązanie wodorowe; ekspansyny nie są białkami enzymatycznymi, ponieważ wiązanie wodorowe nie jest typowym wiązaniem chemicznym).
* Transglukozydaza ksyloglukanu - enzym rozcinający wiązania glikozydowe między hemicelulozą (ksyloglukanem), zmienia układ struktury i wiązań.
* CKI - białka inhibitujące kolejne podziały komórkowe.
* NOR - (nucleosome organizer region) - w tym regionie zakłada się jądro po podziale
* UDP-glukoza - ufosforylowana forma, prekursor do budowy celulozy. MT kortykalne określają gdzie się utworzą włókna (Kolchicyna hamuje ten proces)
Ćwiczenia IV - Plastydy.
- amyloplasty
- leukoplasty
- chloroplasty (tylko one chlorofil mają - fotosynteza)
- chromoplasty
1. Proplastydy
2. Chloroplasty
proplastyd -> plastyd ameboidalny -> plastyd przedgranowy -> chloroplast (jeśli ma dostęp (sferyczny) (ameboidalny) (następuje wpuklanie do światła)
błony tworząc pęcherzyki)
\
\/
etioplast
(jeśli brak dostępu do światła)
- Faza świetlna - zachodzi niemal w całości w błonach tylakoidów i wymaga stałego dopływu światła (produktami są ATP i NADPH)
- Faza ciemna - zachodzi całkowicie w stromie i nie wymaga dopływu światła, faza ciemna wykorzystuje produkty z fazy świetlnej (jasnej) do produkcji cukrów z użyciem CO2
- tylakoidy gran (tworzą właśnie te skupiska)
- tylakoidy stromy (przenikają swobodnie stromę)
- obszary nukleoidopodobne, rybosomy, ziarna skrobi, plastoglobule, depozyty fitoferrytyny (białko wiążące nadmiar jonów żelaza)
3. Amyloplasty
4. Leukoplasty
5. Chromoplasty
- globularne (powstaje w wyniku transformacji chloroplast -> chromoplast, posiada więcej plastoglobuli niż chloroplast, wysoki stosunek lipidy/białko)
- tubularne (tutaj funkcje gromadzenia karotenoidów mają tabule, niski stosunek lipidy/białko, długie i cienkie struktury, cylindryczne, współistnieją z plastoglobulami)
- krystaliczne (tutaj karotenoidy skumulowane są w formie kryształów, otoczone błoną podobną do błony tylakoidów)
- błonowe (struktury gromadzące barwniki mają postać błoniastą, tworzą retikulum chromoplastowe, powstają przez inwaginację wewnętrznej błony plastydu)
Wyszukiwarka
Podobne podstrony:
ĆWICZENIE 14, Studia - Chemia kosmetyczna UŁ, II rok, III semestr, CHEMIA NIEORGANICZNA laboratorium
ĆWICZENIE 11, Studia - Chemia kosmetyczna UŁ, II rok, III semestr, CHEMIA NIEORGANICZNA laboratorium
notatki pirożenko22 10 09, Psychologia, Uczelnia, II rok, III semestr, Psychologia rozwoju człowieka
PIII - teoria, Studia, SiMR, II ROK, III semestr, Elektrotechnika i Elektronika II, Elektra, Elektro
elektra P4, Studia, SiMR, II ROK, III semestr, Elektrotechnika i Elektronika II, Elektra, Elektronik
Ekonomika ochrony srodowiska wyklad 18.04.05, administracja, II ROK, III Semestr, rok II, sem IV, Ek
elektra M4, Studia, SiMR, II ROK, III semestr, Elektrotechnika i Elektronika II, Elektra, Elektronik
Prawo finansowe W I, administracja, II ROK, III Semestr, rok II, sem IV, prawo
Egz mech 2(1), Studia, SiMR, II ROK, III semestr, Mechanika Ogólna II, Mechanika 2, Mechanika
jasiek pytania, Studia, SiMR, II ROK, III semestr, Elektrotechnika i Elektronika II, Elektra, Elektr
BIOCHEMIA- pojęcia, Studia, II rok, II rok, III semestr, Biochemia
inż, administracja, II ROK, III Semestr, podstawy budownictwa + inżynieria komunikacyjna
M2, Studia, SiMR, II ROK, III semestr, Elektrotechnika i Elektronika II, Elektra, Elektronika i Elek
zagadnienia - wyklad 5, II ROK, III SEMESTR, Fizjologia zwierząt
ATT00008, administracja, II ROK, III Semestr, Socjo wsi
Socjologia wsi 4, administracja, II ROK, III Semestr, Socjo wsi
Genetykda 2, Studia, II rok, II rok, III semestr, Genetyka
więcej podobnych podstron