Autekologia lasu
Autoekologia - nauka o przystosowaniu się organizmów żywych do ich środowiska
1. Siedlisko i środowisko leśne
siedlisko = ekotop - kompleks czynników abiotycznych
środowisko = biotop - czynniki abiotyczne ukształtowane przez biotyczne
ryc. 3 i 4 str 11.
2. Czynniki abiotyczne
Teoria czynników ograniczających - wszystkie elementy natury edaficznej jak i klimatycznej są dla rośliny bezwarunkowo potrzebne. Nie ma czynnika który można by zastąpić innym. W przyrodzie istnieje różnorodność i różne zależności, badania muszą być więc kompleksowe i należy badać współzależności (fitotron)
najbardziej czuły wskaźnik reagujący na układ warunków siedliskowych to roślina - wykorzystujemy ją w badaniach fenologicznych. Obserwujemy związek pomiędzy funkcjami fizjologicznymi roślin a warunkami klimatycznymi.
obserwujemy fenofazy: rozwój liści, kwitnienie, dojrzewanie, opadanie owoców i liści - mówią one o początku i końcu fenologicznych pór roku (przedwiośnie, pierwiośnie, wiosna, wczesne lato, lato, wczesna jesień, jesień i zima).
izofeny: linie łączące regiony o jednakowym terminie początku i końca fenofaz (b. przejrzysty wskaźnik)
zmiana siedliska przez człowieka: obróbka mechaniczna gleby, nawożenie, właściwy dobór roślin - ubożenie lasu: przerzedzanie drzewostanów wywóz drewna czyli zw. Mineralnych,
przykład - z każdym metrem sześciennym drewna sosnowego zabiera się z gleby następujące ilości składników (w gramach)
Ca P K
grubizna (stare drewno) 683 69 166
drobizna (młode drewno) 2150 629 793
a) stosunki klimatyczne w lesie
ilość promieniowania słonecznego a szerokość geograficzna
ruchy atmosfery a szerokość geograficzna (ryc. 5 i 6)
itd. Zeskanować
6. Właściwości biologiczne i wymagania ekologiczne najważniejszych gatunków drze leśnych
Szeregi ekologiczne wymagań drzew w odniesieniu do różnych czynników środowiskowych.
Podczas gdy gatunki lekkonasienne, zasiedlające wolne od roślin drzewiastych powierzchnie, wykazują duże wymagania w stosunku do światła, a małe w odniesieniu do wilgoci i obsiewają się na powierzchniach odkrytych, to gatunki ciężkonasienne, o dużych wymaganiach siedliskowych, odnawiają się zazwyczaj przy ograniczonym dostępie światła pod okapem drzewostanu macierzystego. Gatunki te są zwykle również wrażliwe na niekorzystne sploty warunków pogodowych, w szczególności na duże amplitudy termiczne i występowanie przymrozków.
Znajomość skal wymagań drzew leśnych w stosunku do poszczególnych czynników ekologicznych ma w hodowli lasu olbrzymie znaczenie praktyczne. Wymagania te według Puchalskiego i Prusinkiewicza (1968) przedstawiają się następująco:
1. Stosunek drzew do światła
Wyróżniamy tu trzy zasadnicze grupy ekologiczne (nie licząc podgrup):
Drzewa światłożądne (zaczynając od wymagających najwięcej światła: modrzew, brzoza brodawkowata, topole (bez osiki), sosna zwyczajna, olsza szara i robinia akacjowa.
Drzewa cienioznośne:
gorzej znoszące ocienienie - dąb szypułkowy, wiąz górski, limba, olsza czarna, jesion, osika, brzoza omszona,
lepiej znoszące ocienienie - grab, świerk, daglezja zielona, lipa drobnolistna, klon zwyczajny, klon jawor, dąb bezszypułkowy, dąb czerwony, sosna wejmutka.
Drzewa cieniolubne: buk, jodła i cis.
2. Stosunek drzew do ciepła
Wymagania drzew rozpatrywać tu musimy oddzielnie w klimatach niżowym i górskim, drzewa bowiem zachowują się odmiennie w tych makroklimatach.
W warunkach klimatu nizin, ze zmniejszającą się temperaturą powietrza od południowego zachodu ku północnemu-wschodowi:
Drzewa o dużych wymaganiach: lipa wielkolistna, jodła, buk, jawor, dąb bezszypułkowy.
Drzewa o średnich wymaganiach: grab, dąb szypułkowy, lipa drobnolistna, klon zwyczajny, jesion, olsza czarna, olsza szara, wiąz górski.
Drzewa o małych wymaganiach: świerk, sosna zwyczajna, modrzew europejski, osika, brzoza brodawkowata.
W warunkach klimatu podgórskiego i górskiego:
Drzewa o dużych wymaganiach: olsza czarna, grab, dąb szypułkowy, dąb bezszypułkowy, sosna.
Drzewa o średnich wymaganiach: olsza szara, jesion, lipa drobnolistna i wielkolistna, brzoza brodawkowata, wiąz górski, klon zwyczajny, jodła, buk, osika, jawor.
Drzewa o małych wymaganiach: modrzew europejski, limba, świerk, ja
3. Stosunek drzew do wody
Wyróżniamy tu także trzy podstawowe grupy ekologiczne:
Higrofity - drzewa o dużych wymaganiach wilgotnościowych: olsza czarna, brzoza omszona, jesion, wiąz górski, topole (bez osiki), dąb szypulkowy, świerk.
Mezofity - drzewa o średnich wymaganiach wilgotnościowych: jodła, klon zwyczajny, buk, grab, dąb bezszypulkowy, dąb czerwony, modrzew, sosna wejmutka, daglezja zielona.
Kserofity - drzewa o małych wymaganiach wilgotnościowych: robinia akacjowa, osika, brzoza brodawkowata, sosna zwyczajna, sosna czarna.
4. Stosunek drzew do gleby
Według wymagań w stosunku do zasobności gleby w składniki odżywcze można drzewa leśne podzielić również na trzy grupy ekologiczne:
Megatrofy (drzewa o dużych wymaganiach): jesion, jawor, wiąz górski, dąb szypulkowy, topole (bez osiki).
Mezotrofy (drzewa o średnich wymaganiach): dąb bezszypułkowy, buk, lipa drobnolistna, olsza czarna, grab, klon zwyczajny, jarząb pospolity, osika, topola biała i czarna, modrzew, jodła, daglezja, sosna wejmutka, świerk.
Oligotrofy (drzewa o małych wymaganiach): dąb czerwony, brzoza brodawkowata, olsza szara, robinia akacjowa, sosna zwyczajna i sosna czarna.
5. Stosunek drzew do wiatru
Pod względem stopnia odporności na wiatry i burze można drzewa leśne uszeregować (zaczynając od najodporniejszych) następująco:
Drzewa liściaste: dąb, klon, lipa, wiąz, jesion, olsza, buk, topola.
Drzewa iglaste: modrzew, sosna zwyczajna, sosna wejmutka, jodła, daglezja, świerk.
6. Odporność drzew na przymrozki
Według dotychczasowego stanu wiedzy drzewa pod względem stopnia ich wrażliwości na przymrozki można uszeregować następująco:
bardzo wrażliwe: buk, jodła i jesion;
mniej wrażliwe: świerk, dąb i daglezja;
rzadko cierpiące od późnych przymrozków: modrzew, klon, lipa;
odporne: grab, brzoza, osika, olsza, wiąz, jarząb i sosna.
Warto przypomnieć, że u dębu bezszypułkowego kwiaty i liście rozwijają się przeciętnie o 2 tygodnie później od dębu szypułkowego i stąd rzadziej cierpi on od przymrozków. Także w obrębie gatunku dębu szypułkowego wyróżnia się wcześnie pędzącą formę „precox" i późno pędzącą „tardiflora", która rozwija się po okresie wiosennych przymrozków. W tym zakresie istnieje wdzięczne pole do popisu dla selekcji drzew leśnych.
7. Odporność drzew na zanieczyszczenia powietrza
Aczkolwiek sprawa odporności drzew na trucizny emitowane przez przemysł w atmosferę jest rzeczą względną, gdyż przy pewnym dużym stężeniu wszystkie rośliny giną, to jednak przez porównanie można wyodrębnić gatunki bardziej lub mniej wrażliwe na zanieczyszczenia atmosfery. Dlatego stopniując wspomnianą wrażliwość, wyróżnia się pod tym względem cztery grupy ekologiczne:
Gatunki najodporniejsze: buk zwyczajny, klon polny, dąb czerwony, brzozy, topola berlińska, robinia akacjowa, lipa krymska, lipa srebrzysta, sosna czarna, cis pospolity.
Gatunki mniej wrażliwe: dęby szypułkowy i bezszypułkowy, klon jawor i klon zwyczajny, jesion wyniosły, olsze czarna i szara, topole czarna i niekłańska, daglezja zielona, modrzewie polski i europejski, jodła kalifornijska.
Gatunki wrażliwe: lipa drobnolistna, jesion amerykański, topole turkiestańska i Simona, kasztanowiec, wiązy, świerk kłujący.
Gatunki bardzo wrażliwe: jodła pospolita, świerk pospolity, sosna zwyczajna, sosna Banksa, lipa wielkolistna.
Przedstawione tu za Puchalskim skale wymagań ekologicznych drzew, ustalone na podstawie badań naukowych i obserwacji praktyków, nie są jednak niezmienne i ostateczne. Wiadomo bowiem, że amplituda ekologiczna niektórych gatunków jest bardzo szeroka i dlatego rosnąć one mogą zarówno na słabych, jak i zasobnych siedliskach. Poza tym kolejność wymagań, np. w odniesieniu do światła może nawet dość znacznie zmieniać się na różnej glebie (im lepsza gleba, tym większa wytrzymałość na ocienienie), bądź szerzej biorąc w zależności od optimum klimatyczno-edaficznego danego gatunku.
Niemniej znajomość przytoczonych tu skal jest dla hodowcy nieodzowna, gdyż bez niej niemożliwe jest zaplanowanie mieszanego składu gatunkowego uprawy, prowadzenie racjonalnych czyszczeń i trzebieży, wreszcie zupełnie niemożliwe staje się pokierowanie naturalnym odnowieniem lasu.
Generalnie rzecz biorąc, wszelkie czynności techniczne w lesie podbudowane być muszą gruntowną znajomością ekologii lasu - w szczególności znajomością właściwości biologicznych i wymagań ekologicznych drzew leśnych oraz zdolności wytwórczej siedliska.
Czynniki ekologiczne a naturalne odnowienie lasu.
Jeśli już mowa o naturalnym odnowieniu lasu, które jest w pełni zagadnieniem ekologicznym, stwierdzić trzeba, iż zależy ono w znacznej mierze od korzystnego układu warunków siedliskowo-drzewostanowych, a w lesie zagospodarowanym w równie dużej mierze od przyrodniczego przygotowania gospodarza lasu, który swą działalnością może wspomniane korzystne warunki stworzyć i odpowiednio je kształtować, zgodnie z potrzebami młodego pokolenia drzew.
Znaczenie poszczególnych czynników ekologicznych dla odnowienia naturalnego nie jest jednakowe w kolejnych etapach rozwoju, ale wszystkie one są ważne i decydują o powodzeniu odnowienia. Na pierwszy plan wysuwa się w okresie wschodów ciepło i wilgoć, później o utrzymaniu się nalotów przy życiu decyduje przede wszystkim światło.
Leśnik może regulować stosunki świetlne, cieplne i wilgotnościowe w lesie przez odpowiednie cięcia. Usuwając pewną liczbę drzew, zmniejsza zagęszczenie drzewostanu, przerywa zwarcie i w ten sposób zwiększa intensywność oddziaływania wspomnianych czynników w dnie lasu.
Szczególnie czynnik światła musi być umiejętnie dozowany, by pobudzić do przyrostu naloty, a nie doprowadzić do „zdziczenia" i zachwaszczenia gleby (Jaworski 1973). Idealne warunki świetlne w lesie panują wówczas, gdy gleba leśna pokrywa się skąpo roślinnością zielną. Dowodzi to korzystnego stanu biologicznego gleby, który w hodowli lasu nazywamy stanem lekkiego zazielenienia. Istotnie, gdy patrzeć z perspektywy na glebę, sprawia ona wrażenie jednolitego
W hodowli lasu wypracowano cale systemy i metody cięć sprzyjających odnowieniu naturalnemu, przy których każdy etap rozwojowy nalotu i podrostu wymaga innego natężenia światła. Przykładem takiego dozowania światła w dnie lasu jest klasyczna rębnia częściowa, w której wyróżniamy 4 rodzaje cięć: cięcia przygotowawcze, dozujące światło w celu pobudzenia drzew do kwitnienia i obradzania nasion oraz w dnie lasu celem doprowadzenia gleby leśnej do wspomnianego wyżej lekkiego zazielenienia (prawidłowo prowadzone trzebieże zastępują w zasadzie ten rodzaj cięć); cięcia obsiewne, zwiększające dopływ wilgoci i ciepła oraz światła w roku nasiennym, tak potrzebnych opadłym nasionom do skiełkowania i rozwinięcia pierwszych liści; cięcia odsłaniające, zmierzające do stopniowego usuwania drzewostanu macierzystego w miarę wzrostu i potrzeb nalotów i podrostów w stosunku do światła, i pomniejszania konkurencji korzeni starych drzew z młodym pokoleniem. Jednocześnie dokonuje się hartowanie podrostów i przystosowanie do samodzielnego życia na otwartej przestrzeni. Wreszcie ostatnim rodzajem cięć odsłaniających jest cięcie uprzątające, w którym usuwa się znad młodnika o średniej wysokości 2-3 m ostatnie przestoje drzewostanu macierzystego, a młode pokolenie, już zahartowane i wyprowadzone z niebezpiecznej przygruntowej warstwy powietrza (rosnące początkowo w łagodnym klimacie wnętrza drzewostanu), zaczyna samodzielne życie w pełnym oświetleniu.
Z podobnym, może jeszcze bardziej stonowanym dozowaniem światła spotkać się możemy przy odnowieniu krawężnym na północnym brzegu drzewostanu przy rębni brzegowo-smugowej Wagnera.
Jeśli idzie o temperaturę, to dla odnowienia naturalnego ma ona duże znaczenie w okresie kiełkowania. Niekorzystne oddziaływanie temperatury obserwujemy przy samosiewach bocznych na otwartej powierzchni, gdzie przymrozki spóźnione mogą wyrządzić duże spustoszenie wśród gatunków wrażliwych. Przy odnowieniu podokapowym tego rodzaju niebezpieczeństwo im nie grozi. Pośrednio wysoka temperatura może być niebezpieczna dla odnowienia naturalnego w przypadku wystąpienia długotrwałej suszy; płytko ukorzenione naloty mogą wtedy przepadać wskutek przegrzanej i przesuszonej wierzchniej warstwy gleby.
Znaczenia wilgoci dla odnowienia naturalnego nie trzeba specjalnie podkreślać -jest ono oczywiste i zasadniczo szansę powodzenia naturalnego odnowienia mają drzewostany w terenach o wystarczających i korzystnie rozłożonych w okresie wegetacyjnym opadach, a także pomyślnie układających się stosunkach wodnych gleby. Naloty powstające pod okapem drzewostanów macierzystych narażone są na konkurencję ze strony starych drzew. Ssące działanie korzeni drzew matecznych osusza glebę wokół drzew i dlatego często obserwuje się słaby przyrost bądź przepadanie nalotów i podrostów na talerzach wokół Pni starych drzew. Te charakterystyczne „talerze" ze słabo rozwijającym się nalotem wokół pni drzew obserwował już Fabricius (1929), który w sposób bardzo prosty udowodnił, że przyczyna tkwi w konkurencji korzeni, gdyż po przecięciu korzeni starych drzew w miejscach z zahamowanym przyrostem młodego pokolenia po krótkim czasie zabiedzone drzewka dogoniły wzrostem lepiej od początku wzrastające sąsiedztwo.
Jak podaje Kowalski (1981), proces odnowienia w lesie naturalnym przebiega w warunkach siedliskowych kształtowanych nrzp7 Hrzpwnstan
prawo minimum Liebiga
prawo głoszące, że wzrost i rozwój rośliny zależą od ilości tego składnika pokarmowego, którego w podłożu jest najmniej w stosunku do potrzeb. P.m.L. zostało sformułowane w 1840 przez chemika niemieckiego J. Liebiga. Jeśli w niezmienionych, optymalnych warunkach środowiska roślinie dostarczy się dużo azotu, fosforu, żelaza i jednocześnie zbyt mało potasu, to wzrost rośliny będzie ograniczony przez potas. W myśl p.m.L. rośliny uprawne należy nawozić tym pierwiastkiem, który w glebie znajduje się w niewystarczających ilościach. P.m.L. odnosi sig również do zwierząt. P.m.L. dotyczy tylko składników niezbędnych do wzrostu i rozwoju organizmów, do czynników środowiska zaś, których skutki działania szybko ulegają zmianie, odnosi się zasada -¦ tolerancji ekologicznej. P.m.L. znalazło praktyczne zastosowanie w nawożeniu roślin w rolnictwie, a także w leśnictwie i łowiectwie.
prawo Shelforda
prawo głoszące, że występowanie i rozwój organizmu w środowisku zależy od całego kompleksu czynników i zarówno nadmiar, jak i niedobór któregokolwiek z nich może działać ograniczająco. P.S. zostało sformułowane w 1913 przez ekologa amerykańskiego V.E. Shelforda. Czynniki abiotyczne środowiska określają tempo procesów biologicznych organizmu, możliwości przeżycia, rozrodu oraz decydują o jego rozmieszczeniu w środowisku. Oddziaływanie na organizm czynników środowiska zależy od ich natężenia. Każdy organizm ma określoną tolerancję względem czynników środowiska. Zob. tolerancja ekologiczna.
Wykład 2
1. Pojęcie biocenozy lasu i ekosystemu leśnego
Biocenoza - naturalny zespół organizmów żywych danego środowiska, powiązanych ze sobą przez różne czynniki ekologiczne i tworzących organiczną całość, która dzięki zdolności do rozmnażania i samoregulacji zagęszczenia utrzymuje się trwale w przyrodzie w stanie dynamicznej równowagi.
2. Ekoton (efekt stykowy biocenoz) - to strefa przejścia między dwiema biocenozami. Jest bogatsza w gatunki i ma bardziej skomplikowany układ warunków ekologicznych, niż każda z biocenoz sąsiednich.
3. Nisze ekologiczne gatunku - niszą ekologiczna nazywamy sytuację lub umiejscowienie gatunku w biocenozie. N.E. gatunku zależy od tego gdzie on żyje i co robi. Siedlisko to „adres” organizmu, nisza to jego „zawód”. Jest to więc przestrzeń fizyczna zajmowana przez dany organizm, jak i funkcja tego organizmu w biocenozie oraz zależność od warunków bytowania. Są 3 etapy powstawania tego terminu:
nisza siedliskowa (mikrosiedlisko zajmowane przez organizm)
nisza troficzna (przemiany energetyczne organizmu)
nisza fizjologiczno-przystosowawcza (przestrzeń w obrębie której środowisko umożliwia utrzymanie się osobnika lub gatunku przy życiu.
4. Homeostaza ekosystemu, zasady regulacji biocenotycznej
Homeostaza - samoistna zdolność utrzymywania się i samoregulacji. Wg Oduma - tendencja układów biologicznych do opierania się zmianom i trwania w stanie równowagi.
Homeostazę określają 4 zasady regulacji biocenotycznej:
Zachowanie struktury. Dążność i zdolność do odtworzenia zniszczonej struktury.
Zachowanie obiegu materii i przepływu energii.
Zachowanie produktywności
Stabilizacja procesów.
5. Obieg materii i przepływ energii w biocenozie i ekosystemie
a) cykle biogeochemiczne i prawa termodynamiki
cykle biogeochemiczne - pierwiastki chemiczne krążą w biosferze od środowiska do rośliny i z powrotem tworząc obiegi zamknięte zwane cyklami biogeochemicznymi:
obieg typu gazowego realizujący się w atmosferze i hydrosferze [cykle azotu, tlenu, węgla (CO2)]. W skali Ziemi prawie doskonałe dzięki naturalnej regulacji na zasadzie sprzężenia zwrotnego.
obieg typu sedymentacyjnego dokonujący się na powierzchni skorupy ziemskiej [cykle fosforu, żelaza]. Są mniej doskonałe, krążenie ich łatwiej może ulec przerwaniu wskutek lokalnych zakłóceń.
Entropia - organizmy, ekosystemy i biosfera mają zdolność do tworzenia i utrzymywania stanu wewnętrznego uporządkowania (t.j. stanu niskiej entropii).
I prawo termodynamiki - energia może przemieniać się w inną postać, nigdy jednak nie powstaje ani nie ulega zniszczeniu. Energia słoneczna dochodzi do Ziemi i w większości zamienia się w energię cieplną. Część energii słonecznej absorbowana jest przez rośliny i magazynowana jako energia potencjalna. Zwierzęta pobierają chemiczną energię potencjalną w pokarmie, część jej jest związana w protoplazmie a większość ulega degradacji (przemianie w ciepło).
II prawo termodynamiki - każdy naturalny układ zamknięty, przez który odbywa się przepływ energii (np. Ziemia, las, jezioro) podlega przemianom dopóty, dopóki nie nastąpi trwałe dostosowanie się do otaczających warunków za pomocą mechanizmów samoregulujących. W układzie który osiągnął stan równowagi dynamicznej, przemiany energetyczne zachodzą w stały zawsze jednakowy sposób, w określonym tempie, zgodnie z zasadą stabilności.
Czy w naturze istnieje tendencja do wzrostu biomasy i zmniejszenia nakładów energii podtrzymującej jej istnienie czy też zwiększa się tylko przepływ energii?
b) poziomy troficzne w biocenozie i ekosystemie, łańcuchy pokarmowe i piramidy ekologiczne
Dla biocenozy najistotniejsza jest jej wewnętrzna organizacja, przejawiająca się w tzw. związkach biocenotycznych, które bywają bardzo różnorodne i skomplikowane. Jedne z najważniejszych to związki pokarmowe.
Stosunki pokarmowe mają tak duże znaczenie w dynamice i funkcjonowaniu żywej przyrody (biocenozy), że stworzone zostało w ekologii specjalne pojęcie struktury troficznej przejawiającej się w tzw. łańcuchach pokarmowych i piramidach ekologicznych. Wszelkie gatunki odnośnego zgrupowania tworzą szeregi łańcuchów pokarmowych, których całość składa się na tzw. cykl pokarmowy zgrupowania (rys. 66).
Podstawę cyklu pokarmowego tworzą zawsze organizmy autotroficzne, produkujące w procesie fotosyntezy substancję organiczną przyswajalną dla zwierząt roślinożerców (tzw. konsumentów I rzędu), które pożerane są z kolei przez małe drapieżniki (konsumentów II rzędu), te następnie przez większe drapieżniki mięsożerne i jeszcze większe, a na końcu łańcucha znajduje się jakiś wielki największy dla danego zgrupowania drapieżnik, który poza człowiekiem i chorobotwórczymi mikrobami nie ma już innych wrogów.
Łańcuchem pokarmowym nazywamy przepływ energii zawartej w pokarmie, począwszy od jej źródła, czyli roślin, poprzez szereg organizmów kolejno zjadających i zjadanych.
W przyrodzie mogą występować dwa rodzaje łańcuchów pokarmowych:
łańcuch spasania, zaczynający się na roślinach a kończący na wielkim drapieżniku (przykład na rysunku 66);
łańcuch detrytusowy, który wiedzie od martwej materii organicznej do drobnoustrojów (saprofagów) i zjadających je drapieżników.
Łańcuchy pokarmowe często się w przyrodzie krzyżują i wiążą wzajemnie. Taki układ powiązanych ze sobą łańcuchów w różnych ich ogniwach nazywamy w ekologii siecią troficzną lub siecią zależności pokarmowych. Mówiliśmy już, że w łańcuchach pokarmowych łatwo dopatrzyć się różnych poziomów troficznych.
poziom pokarmowy, poziom troficzny - [gr. trophe = pożywienie]- grupa osobników różnych gatunków spełniających podobną funkcję w procesie przepływu energii i obiegu materii w ekosystemie.
producenci - wszystkie organizmy samożywne wytwarzające materię organiczną w procesie fotosyntezy lub chemosyntezy.
konsumenci, czyli wszystkie zwierzęta. Zależnie od rodzaju pobieranego pokarmu wśród konsumentów można wyróżnić kilka p.p.:
konsumentów pierwszego rzędu (roślinożercy),
konsumentów drugiego rzędu (zwierzęta żywiące się roślinożercami)
k. dalszych rzędów (zwierzęta drapieżne, atakujące inne drapieżce lub pasożyty drapieżców).
reducenci (destruenci) organizmy żywiące się martwą substancją organiczną
Wyróżnienie p.p. umożliwia zilustrowanie w sposób graficzny struktury pokarmowej biocenozy (piramida ekologiczna).
piramida ekologiczna - graficzny sposób przedstawiania struktury pokarmowej biocenozy. P.e. ilustruje łączną wartość łańcuchów pokarmowych w całej biocenozie. Wszystkie organizmy samożywne (producenci) w danej biocenozie stanowią podstawę p.e. Kolejny poziom tworzą konsumenci pierwszego rzędu, do którego należą wszystkie organizmy roślinożerne; wszystkie drapieżniki tworzą poziom konsumentów drugiego i dalszych rzędów (rys.). Wyróżnia się piramidę biomasy, przedstawiającą ilość materii budującą każdy poziom pokarmowy, piramidę liczebności, obrazującą liczbę osobników każdego poziomu pokarmowego, piramidę energii, obrazującą przepływ energii przez poszczególne poziomy.
piramida biomasy - graficzny sposób przedstawiania ogólnej masy organizmów na każdym poziomie pokarmowym (troficznym) danej biocenozy, w danym czasie. P.b. określa masę producentów, konsumentów pierwszego i dalszych rzędów w biocenozie. Ilość biomasy można wyrazić w g lub kg suchej masy na m2 lub m3. P.b. może przyjmować różną postać. W biocenozie lądowej najczęściej biomasa producentów jest największa. Mniejszą biomasę stanowią konsumenci pierwszego rzędu, jeszcze mniejszą konsumenci drugiego rzędu (rys. a). Odwrócona p.b. jest charakterystyczna dla biocenoz wodnych, w których producentami są organizmy drobne, lecz szybko się rozmnażające (glony planktonowe), natomiast konsumentami (zwierzęta roślinożerne, drapieżne) są organizmy większe i o dłuższym czasie życia (rys. b).
piramida energii - graficzny sposób przedstawienia ilości energii zawartej w związkach organicznych przechodzącej z najniższego do najwyższego poziomu pokarmowego (troficznego) w danej biocenozie. Przekazywanie energii z poziomu na poziom odbywa się z pewnymi jej stratami. Część zasymilowanej energii na każdym poziomie pokarmowym ulega rozproszeniu, co powoduje zmniejszenie jej ilości dostępnej wyższym poziomom. Ilość przekazywanej energii z poziomu na poziom jest coraz mniejsza P.e. nigdy nie może być piramidą odwróconą, a jej kształt nie zależy ani od wielkości osobników, ani od tempa ich przemiany materii.
piramida liczebności - graficzny sposób przedstawiania liczby osobników należących do poszczególnych poziomów pokarmowych (troficznych) danej biocenozy (producentów i konsumentów). P.l. określa liczbę osobników poszczególnych poziomów w danym momencie na jednostkę powierzchni (rys. a). P.l. nie przedstawia tempa zmian liczebności osobników w czasie, dlatego może przyjmować postać piramidy odwróconej (rys. b), gdy niższy poziom pokarmowy ma mniejszą liczbę osobników niż wyższy, np. na jednym drzewie może żyć wiele owadów lub ptaków.
Ekosystem obejmuje zarówno organizmy żywe jak i ich abiotyczne otoczenie, czyli siedlisko. Oba te światy (ożywiony i nieożywiony) traktuje się tu jako równorzędnych partnerów. Jest to zamknięty układ ekologiczny, który z zewnątrz otrzymuje tylko energię promienistą Słońca (rys. 68).
Ekosystem oznacza każdą przestrzeń (stanowiącą pewną całość pod względem przyrodniczym), w której zachodzi stała wymiana materii pomiędzy jej żywą i nieożywioną częścią, jako wynik wzajemnego oddziaływania na siebie organizmów żywych i martwej materii
W ekosystemie do przytoczonych poziomów biocenotycznych (producentów, konsumentów i reducentów) dochodzi jeszcze jeden poziom w postaci czynników abiotycznych składających się na siedlisko.
6. Pierwotna i wtórna produktywność biocenozy
Poszczególne osobniki potrzebują stałego dopływu energii, by zrównoważyć jej wydatki ponoszone na metabolizm, wzrost i reprodukcję. Zużytkowują więc energię i przekształcają materię.
Istnieją dwa sposoby pobierania energii i materii przez organizmy:
Autotrofy pobierają energię słoneczną i związki nieorganiczne (rośliny zielone).
Heterotrofy pozyskują energie i materię żywiąc się materią organiczną.
Roślinożercy są heterotrofami, które odżywiają się roślinami, a drapieżniki są heterotrofami które zjadają inne zwierzęta. W skład biocenozy wchodzą i autotrofy i heterotrofy.
Energia i materia które docierają do biocenozy, są zużytkowywane przez osobniki i włączane w strukturę biologiczną, aby w końcu i tak zostały uwolnione do środowiska.
Ekosystem jest rodzajem transformatora który pobiera energię słońca, wiąże ją w procesie fotosyntezy i przenosi tę energię od roślin zielonych poprzez roślinożerców do drapieżników. Większość energii przepływa przez biocenozę tylko raz. Energia nie krąży, ale jest zamieniana na ciepło i tracona ostatecznie przez ekosystem. Tylko ciągły dopływ nowych porcji energii słonecznej podtrzymuje działanie biocenozy. Ryc.
Podstawą wszelkiego życia jest proces fotosyntezy
Fotosynteza jest procesem przekształcającym energię promieniowania słonecznego na energię chemiczną.
12H2O + 6CO2 + energia promieniowania słonecznego - chlorofil - C6H12O6 + 6O2 + 6H2O
enzymy
Rośliny równocześnie oddychają, zużywając energię do podtrzymania procesów życiowych. Oddychanie jest odwrotnością fotosyntezy:
C6H12O6 + O2 enzymy metaboliczne CO2 + H2O + energia zużywana na procesy życiowe
1. Produkcja pierwotna
produkcja pierwotna brutto = energia wiązana w procesie fotosyntezy w jednostce czasu
produkcja pierwotna netto = energia wiązana w procesie fotosyntezy minus energia tracona w procesie oddychania w jednostce czasu.
Produkcja pierwotna zmienia się w zależności od typu roślinności.
Największa jest w wilgotnym lesie tropikalnym i stopniowo zmniejsza się w stronę biegunów.
Produkcja oceanów jest bardzo mała, podobna jak arktycznej tundry. Zbiorowiska trawiaste są mniej produktywne niż lasy w tym samym rejonie.
Wydajność prod. pierw. brutto = energia wiązana w produkcji pierwotnej brutto/energia zawarta w docierającym promieniowaniu słonecznym.
Wydajność przekształcania energii przez roślinność jest nieduża:
Plankton = 0,5%, uprawy poniżej 1,5%, rośliny zielne 1-2%, lasy 2-3,5%.
Znaczna część tej energii jest tracona przy przekształcaniu energii słonecznej w energię chemiczną w procesie produkcji pierwotnej brutto (w trakcie oddychania samych roślin). Produkcja pierwotna netto jest jeszcze mniej wydajna. W lasach 50-75% p.p.b. jest tracona w czasie oddychania, czyli p.p.n. może stanowić 50-25% p.p.b.
W wielu biocenozach lądowych ok. 1% promieniowania słonecznego docierającego do roślin zostaje związana w energię chemiczną w procesie p.p.n.
A. Czynniki ograniczające produkcję pierwotną.
a) W ekosystemach wodnych:
światło - nawet w przejrzystych wodach na 1 metrze głębokości pochłaniana jest połowa światła. Zbyt duża intensywność światła także może hamować fotosyntezę. W tropikalnych wodach gdy promieniowanie jest nadmierne największa fotosynteza zachodzi kilka metrów pod powierzchnia wody.
W związku z tym powinien następować spadek produktywności z przesuwaniem się w stronę biegunów. Jednak nie jest tak.
Czynnikiem ograniczającym p.p są substancje pokarmowe (mała zawartość azotu i fosforu dla fitoplanktonu)
Wody Morza Sargassowego są czyste lecz ubogie i odznaczają się małą produktywnością. Jednak tu głównym czynnikiem ograniczającym wydaje się być żelazo, potem azot i fosfor.
Produkcja tam jest większa zimą (mieszanie się wód z różnych poziomów dzięki wiatrom, dolne poziomy wód zawierają większe ilości związków pokarmowych
Najbardziej produktywne są morza polarne, tam są najbardziej obfite połowy ryb.
b) W ekosystemach wód słodkich
światło (i temperatura)
fosfor
krzem
B. Czynniki czasem określające organizację biocenozy
Azot
Żelazo
Mangan
Molibden
C. Czynniki rzadko określające organizację biocenozy
węgiel
kobalt,
Siarka
mikroelementy.
c) w środowiskach lądowych
Opady ryc.
Temperatura ryc.
Lasy iglaste rosnące w tych samych warunkach klimatycznych są średnio bardziej produktywne.
Drzewa iglaste maja większą łączną powierzchnię liści, dłuższy okres produkcji
Produkcja pierwotna brutto w zbiorowiskach trawiastych jest ograniczana przez brak deszczu, temperatura ma mniejsze znaczenie, drugorzędne ograniczenia powodowane są przez typ gleby, jej pojemność wodną oraz dostępność substancji pokarmowych
Ryc. Przedstawiająca wpływ dodatku azotu i fosforu
Metody mierzenia produktywności:
l. Metoda zbioru plonu jest to metoda mająca praktyczne zastosowanie w leśnictwie, gdzie metodami dendrometrycznymi określa się wielkość produkcji drewna jako podstawowego produktu lasu. Ilość materiału roślinnego wyprodukowanego w jednostce czasu może być określana na podstawie różnic między ilością materiału w dwu momentach:
ΔB=B2-B1
B = zmiana biomasy w biocenozie pomiędzy czasem t1 a czasem t2
B1 = biomasa w czasie t1
B2 = biomasa w czasie t2
2. Metoda tlenowa polega na pomiarze uwalnianego w asymilacji tlenu, gdyż między ilością wytwarzanych związków pokarmowych a ilością uwolnionego tlenu istnieje ścisła zależność. Nie jest ona zbyt dokładna, gdyż wymiana tlenu z otoczeniem i zużywanie go przez rośliny oraz zwierzęta prowadzi do nieścisłości pomiarowych.
3. Metoda dwutlenkowa jest metodą fizjologiczną, polegającą na określaniu ilości wiązanego CO2 w procesie fotosyntezy liści. Ekologowie z powodzeniem zastosowali tę metodę w mierzeniu produktywności całych biocenoz.
4. Metoda szybkości zużywania substancji mineralnych zawartych w środowisku najczęściej stosowana w pomiarze produktywności zbiorników wodnych (oceanicznych), służąca do określania czystej produkcji całej biocenozy morskiej.
5. Metoda izotopowa z zastosowaniem radioaktywnego węgla C14 i fosforu P32. Polega ona na pomiarze szybkości obiegu znaczonych pierwiastków w środowisku. Ilość zasymilowanego pierwiastka promieniotwórczego ma być mniej więcej proporcjonalna do produktywności organicznej.
6. Metoda chlorofilowa nabrała dużego znaczenia w ekologii po odkryciu Gessnera (1949), który dowiódł, że na 1 m2 przypada w obrębie biocenozy jednakowa ilość chlorofilu i że ilość ta jest na ogół podobna w różnych biocenozach.
2. Produkcja wtórna
produkcja wtórna - materia organiczna (energia) gromadzona w ciele konsumentów i reducentów żyjących w danym ekosystemie. Konsumenci wykorzystują tylko pokarm uprzednio już wytworzony i przekształcają go we własne tkanki z dużymi stratami na oddychanie.
Produktywnością wtórną nazywamy szybkość magazynowania energii na troficznych poziomach konsumentów i reducentów (zwaną także "przyswajaniem"). Istnieje tylko jeden typ produktywności wtórnej, gdyż konsumenci zużytkowują uprzednio już wytworzony materiał pokarmowy, przekształcając go we własne tkanki ze znacznymi stratami energii na oddychanie.
7. Dynamika biocenozy w ekosystemie leśnym
Fitocenoza leśna - jest najistotniejszą i najbardziej stabilną częścią biocenozy leśnej i tak jak cała biocenoza znajduje się w ciągłym rozwoju i przemianach.
Zmienia się ona zarówno w czasie jak i w przestrzeni, przy czym zmiany te mogą być różnokierunkowe i występować w różnym nasileniu, zależnie od kierunku i intensywności działających przyczyn.
Zmiany zachodzące w biocenozie dzieli się na trzy zasadnicze kategorie:
zmiany cykliczne, czyli wahadłowe (dobowe, sezonowe, roczne, okresowe),
zmiany regeneracyjne (zwane także sukcesją wtórną),
zmiany asymptotyczne (zmiany sukcesyjne i historyczne).
A - Zmiany cykliczne
Zmiany cykliczne powstają na skutek działania czynników okresowo przemijających i nie wykraczają poza normalny obręb szaty roślinnej.
Zaliczamy tu zmienność dobową, sezonową, roczną i okresową, dyktowaną różnym nasileniem w czasie czynników meteorologicznych.
Przez zmienność sezonową rozumiemy np. aspekty fenologiczne, występujące szczególnie wyraźnie w runie cienistych lasów liściastych, bukowych czy grabowych. Wyróżniamy tu dwa aspekty: wiosenny i letni.
Wiosenny złożony jest z roślin, które w krótkim czasie, tj. od wczesnej wiosny do chwili pokrycia się drzew listowiem, przechodzą pełny cykl życiowy (rozwojowy). Większość roślin należących do rodzin: Ranunculaceae, Primulaceae i Labiatae, a także Liliaceae (zawilce, przylaszczki, pierwiosnki, miodunki, kokorycze itp.).
Aspekt letni złożony jest z roślinności przystosowanej już do znikomej ilości światła, przefiltrowanej przez gęsty dach koron drzew (zachyłka, marzanna, gajowiec, dąbrówka, czerniec itp.).
Zmiany roczne powodowane są czynnikami meteorologicznymi. Zazwyczaj wywołuje je różna ilość lub rozkład opadów atmosferycznych, bądź różny przebieg temperatury.
Zmiany okresowe, również zależą od układu czynników pogodowych, meteorologicznych, działających jednak w okresach niestałych. Przykładem mogą tu być lasy zalewowe (olsy typowe lub łęgi), podlegające w okresach wilgotnych, np. wiosną lub jesienią, dłuższemu lub krótszemu zalewowi w zależności od opadów danego roku. Takie zespoły wykazują różną postaciowe, ekologicznie i florystycznie zmienność w okresach zalewu podczas lat suchych.
B - Zmiany regeneracyjne,
występujące pod wpływem przemijającego czynnika destruktywnego, zmierzają do powrotu zniszczonego lub zniekształconego zespołu (niekiedy poprzez szereg ogniw przejściowych) do typu zasadniczego. Obecnie zmiany te zalicza się do tzw. sukcesji wtórnej.
Przykładem mogą tu być stosunki zachodzące na wypalonej przez pożar powierzchni leśnej, zajmowanej przez las mieszany, np. świerkowo-jodłowo-bukowy. Na pożarzysku pojawiają się w krótkim czasie naloty brzozy i osiki, a więc gatunków stanowiących przejściowe ogniwo regeneracyjne do właściwego temu siedlisku zespołu, zwane leśnym ogniwem pionierskim. Pod okapem takiego „przedplonu naturalnego", brzozowego lub osikowego, stopniowo pojawiają się naloty buka, świerka i wreszcie jodły, które to gatunki z biegiem czasu całkowicie opanowują teren, tworzą zwarte piętro podokapowe, uniemożliwiając odnowienie pionierskim gatunkom światłożądnym. W ten sposób krótkowieczne, przejściowe ogniwo pionierskie zostaje wyparte, a las wraca do naturalnego składu sprzed wystąpienia pożaru.
C - Zmiany asymptotyczne,
czyli sukcesje ekologiczne, przebiegają niezmiennie w pewnym określonym kierunku, pod wpływem stale działającego czynnika. Prowadzą one do całkowitej zamiany jednej roślinności przez inną zupełnie różną od pierwotnej.
Typowym przykładem zmian sukcesyjnych jest nasuwanie się lasu na step. Wypieranie roślinności stepowej przez wyżej zorganizowaną asocjację leśną jest zjawiskiem naturalnym. Las jest zespołem potężniejszym niż roślinność zielna i wszędzie tam, gdzie typy te się stykają, nasuwanie się lasu na step jest oczywiste.
Te przemiany polegające na strefowym bądź lokalnym następstwie jednych biocenoz po drugich nazywamy właśnie sukcesjami.
Sukcesja ekologiczna to uporządkowany proces przemian, jakim podlega biocenoza.
Rozwój biocenozy przez oddziaływanie roślinności na środowisko, prowadzący do osiedlania się nowych gatunków, nazywa się sukcesją.
Motorem tych przemian są zmiany w siedlisku, wywołane przez czynniki fizykochemiczne i samą roślinność, które następnie pociągają za sobą dostosowywanie się biocenozy do nowych warunków otoczenia.
Podstawowymi terminami określającymi cały łańcuch przemian, jakim podlega biocenoza, są: „biocenoza pionierska", „stadia seralne" i „klimaks", czyli ogniwo kończące sukcesję ekologiczną. Cały zaś szereg biocenoz składający się na sukcesję zwiemy ogólnym mianem „sera”.
Istnieją dwa przeciwstawne, podstawowe pojęcia związane z sukcesją ekologiczną. Otóż sukcesję, która rozpoczyna się na martwym obszarze nie zajętym jeszcze nigdy przez żadną biocenozę (np. na nagiej skale lub plaży morskiej) nazywamy sukcesją pierwotną. Powierzchnia nagiego gruntu, pozbawiona swojej pierwotnej roślinności na skutek pożaru, powodzi czy zlodowacenia, nie pozostaje długo bez roślin i zwierząt. Szybko jest kolonizowana przez rozmaite gatunki, które następnie modyfikują jeden lub więcej czynników środowiskowych. Ta modyfikacja środowiska może z kolei umożliwić osiedlenie się następnych gatunków.
Jeśli zaś rozwój biocenozy zaczyna się od wyższego, a raczej wyżej zorganizowanego ogniwa przemian i ma charakter regeneracyjny, z którego biocenoza już istniejąca została w taki czy inny sposób usunięta (np. zaorane pole, wyrąbany lub spalony las itp), to taki proces nazywamy w ekologii sukcesją wtórną.
Sukcesja wtórna cechuje się zazwyczaj większą szybkością przemian w biocenozie, ponieważ na danym obszarze istnieje już gleba i kilka przynajmniej organizmów. Ponadto obszar, który uprzednio był już zajęty przez rośliny i inne organizmy zwierzęce sprzyja bardziej rozwojowi biocenozy aniżeli obszary całkowicie „jałowe". Ten właśnie typ sukcesji jest najczęściej obserwowany w przyrodzie.
Właściwością sukcesji ekologicznej jest również jej kierunkowość. W warunkach, w których proces ten jest już dobrze poznany, można w dowolnym czasie określić, z którym stadium seralnym aktualnie mamy do czynienia, a także można przewidzieć jego dalsze zmiany.
W sukcesji pierwotnej najpierw substrat zasiedlają ogniwa pionierskie. Jest to proces powolny, wymagający zwykle długiego okresu. Później pojawiają się dość szybko jedna po drugiej biocenozy seralne i wreszcie znów pod koniec przemian sukcesja zwalnia swe tempo tak, że klimaks stabilizuje się stopniowo w ciągu długiego okresu. Szybkość następowania kolejnych zmian jest różna w różnych etapach sukcesji, a wynika to ze stopniowej zmiany składników biocenoz początkowo krótkowiecznych na długowieczne i z faktu, iż wymianie w kolejnych stadiach podlega większość organizmów biocenoz seralnych.
Jeśli sukcesja przebiega od zbiorowisk prostych do coraz bardziej złożonych, trwałych i wysoko zorganizowanych pod względem ich wewnętrznej budowy, mówimy o sukcesji wstępującej, czyli „progresyjnej".
Bywa jednak, że na skutek degradacyjnej działalności człowieka na siedlisko (np. odwodnienie terenu przez przeprowadzoną nieumiejętnie regulację rzek czy w ogóle stosunków wodnych danego terenu) sukcesja może przebiegać w kierunku odwrotnym, biocenozy ubożeją i mamy do czynienia ze zjawiskiem przemian „regresyjnych". Typowym przykładem jest zakładanie monokultur na miejsce lasów mieszanych. Zakwaszający wpływ sosny czy świerka w litych drzewostanach na glebę prowadzi do jej degradacji i całkowitego przekształcenia lasów czy borów mieszanych na typowe bory iglaste.
Prawo sukcesji ekologicznej mówi, że naturalne biocenozy zmieniają się w pewien prawidłowy sposób, prowadząc do biocenoz stabilnych, znajdujących się w stanie dynamicznej równowagi.
Zespół klimaksowy nie jest jednak czymś bezwzględnie trwałym. Sukcesja ekologiczna doprowadza zawsze do powstania biocenozy ustabilizowanej i przystosowanej do istniejących warunków.
Istnieją jednakże dwie szkoły odmiennie interpretujące to zjawisko:
teoria monoklimaksowa, (sztafeta florystyczna, model ułatwiania) według której każdy region klimatyczny stwarza warunki dla powstania jednej jedynej biocenozy klimaksowej, do której na drodze naturalnej sukcesji zmierzają wszystkie istniejące na tym terenie biocenozy; Rozwój biocenozy odbywa się stopniowo i jest progresywny, od prostych biocenoz pionierskich do stadium ostatecznego, czyli klimaksu. Sukcesja taka przebiega wyłącznie dzięki zależnościom biotycznym — rośliny i zwierzęta stadiów pionierskich tak zmieniają środowisko, że staje się ono korzystne dla nowego zestawu gatunków, a cykl taki powtarza się do czasu osiągnięcia klimaksu. Sukcesja odwrócona (retrogresja; jest możliwa tylko w przypadku wystąpienia w środowisku zakłóceń, takich jak pożar, wypasanie czy erozja. Podstawowym założeniem jest to, że jeden gatunek zastępuje drugi, ponieważ w każdym stadium gatunki tak przekształcają środowisko, że dla nich samych staje się ono mniej odpowiednie, dla innych zaś bardziej odpowiednie. Wymiana gatunków jest więc procesem uporządkowanym i przewidywalnym oraz sprawia, że sukcesja jest procesem kierunkowym.
teoria poliklimaksowa, podług której zdążanie wszystkich biocenoz danego terenu do jednej „modelowej" biocenozy klimaksowej jest całkowicie nierealne, gdyż fizyczne warunki środowiska, przy ustalonym i jednakowym klimacie nigdy nie są wyrównane do tego stopnia, by mogły przekształcić się w krótkim czasie do jednego poziomu.
teoria kompromisowa, według której można przyjąć istnienie jednego teoretycznego klimaksu klimatycznego, w którego obrębie przejściowo w krótszych lub dłuższych okresach kształtują się zmienne, warunkowane zmiennością podłoża liczby klimaksów edaficznych.
Dysklimaks
Często organizmy, na których człowiekowi szczególnie zależy, nie występują w końcowych ogniwach sukcesji ekologicznej bądź biocenozie klimaksowej, lecz znaleźć je można w ogniwach pionierskich czy pośrednich stadiach seralnych. Wiele zwierząt łownych lub roślin drzewiastych rozwija się najlepiej w biocenozach przejściowych, krótkotrwałych.
Powstaje problem, jak zatrzymać sukcesję ekologiczną i utrzymać ogniwo seralne najbardziej pożądane przez człowieka, jednak niezmiernie trudno zrealizować to w naturze. Można by terytorialnie tak pokierować sukcesją biocenoz, by stale w kolejnych latach mieć wystarczająco duże obszary wchodzące w pożądane stadium seralne.
Biocenozę, która dla danego miejsca nie jest ani klimaksem klimatycznym ani edaficznym, a która utrzymuje się wysiłkiem człowieka, nazwać możemy dysklimaksem, czyli klimaksem sztucznym lub zniekształconym.
przykłady - nadmierne wypasanie bydła może doprowadzić do powstania biocenozy pustynnej w takich miejscach, gdzie klimat lokalny dopuszcza istnienie biocenozy stepowej jako zespołu klimaksowego. Biocenoza pustynna jest tu dysklimaksem wywołanym działalnością człowieka i utrzymywanym przez stały wypas, co świadczy o złej gospodarce ludzkiej, natomiast step jest tu rzeczywistym klimaksem klimatycznym. Tylko w głębi kontynentu, gdzie brak wilgoci (Azja, Australia) istnienie biocenozy pustynnej jest zjawiskiem naturalnym i klimaksowym.
w pewnych warunkach w prowincji Ontario trwałe lasy sosnowe utrzymują się na skutek powtarzających się pożarów leśnych. Bez tych pożarów sukcesja prowadziłaby do powstawania lasów liściastych. W tym przypadku dysklimaks ma większą wartość dla człowieka, gdyż surowiec z ogniwa regeneracyjnego jest o wiele cenniejszy od surowca z klimaksowego lasu liściastego.
Nie świadczy to jednak, że zawsze wartościowsze użytki pozyskiwać można z biocenoz seralnych. W niektórych rejonach naszego globu właśnie najcenniejsze drzewa z handlowego punktu widzenia uzyskujemy ze stadium klimaksowego i tam należy czynić wszystko, by przyspieszyć sukcesję i powrót biocenozy do stadium klimaksu. W innych rejonach ma się rzecz podobnie jak to przedstawiono przy dysklimaksach: cenniejsze użytki pozyskuje się z wcześniejszych stadiów seralnych. Stąd prawo sukcesji ekologicznej i koncepcja klimaksu klimatycznego i edaficznego mają tak duże znaczenie dla praktyki leśnictwa.
Wydaje się, że sukcesja jest właściwością samej biocenozy. Wynika ona z faktu, że oddziaływanie biocenoz na środowisko powoduje, iż obszar przez nie zajęty staje się mniej sprzyjający tej biocenozie, a bardziej korzystny dla innego zestawu organizmów; dzieje się to dopóty, dopóki nie zostanie osiągnięty stan równowagi, czyli klimaks. Biocenozy pionierskie wytwarzają znacznie więcej materii organicznej niż jej konsumują. Ten nadmiar materii organicznej może zmienić środowisko fizyczne do tego stopnia, że stanie się ono źródłem pokarmu, co odbije się na składzie gatunkowym biocenozy. Dopiero wtedy, gdy organizmy zdołają zmodyfikować środowisko tak dalece, jak to tylko jest możliwe, biocenoza może osiągnąć stan równowagi dynamicznej - klimaks.
Zagadnienia naturalnej sukcesji w bardzo plastyczny i interesujący sposób przedstawił Thomasius (1980).
Na pozbawionym roślinności (dziewiczym) siedlisku, cechującym się określoną zawartością składników pokarmowych, wilgotnością, temperaturą, promieniowaniem itp., a więc pewnymi warunkami wzrostu lasu, pod wpływem naturalnych czynników w obszarach humidowych pojawia się sukcesja, która prowadzi od:
1 - stadium inicjalnego (przedplonu),
2 - przez stadia przejściowe (lasu pośredniego),
3 - do stadium klimaksowego (lasu końcowego albo zrównoważonego); (rys. 74).
1. Stadium przedplonowe (pionierskie)
Sukcesja zaczyna się imigracją (wkroczeniem) i zasiedleniem (opanowaniem) siedliska przez gatunki o następujących właściwościach:
odporność na ekstremalne warunki klimatyczne (mróz, temperatura, susza, wiatr),
duża zdolność przystosowawcza do warunków glebowych (jałowa gleba, brak mikoryzy itp.),
ekstensywny system korzeniowy,
szybki wzrost w młodości, wczesna kulminacja przyrostu z wysokimi wartościami maksymalnymi i następnie z silnym spadkiem przyrostu (rys. 75),
wczesne dojrzewanie generatywne, częste i obfite obradzanie nasion, zdolność dobrego rozprzestrzeniania się nasion (częsta anemochoria),
krótka żywotność, niskie końcowe wartości przyrostu, relatywnie mała zdolność gromadzenia biomasy i energii w żywych organizmach.
Drzewostany zbudowane z takich gatunków pionierskich są złożone często z jednego lub niewielu gatunków przypadkowo znajdujących się w pobliżu. Ich zróżnicowanie wieku jest niewielkie, budowa pionowa prosta, przeważnie jednopiętrowa.
Produkcja pierwotna netto na jednostkę powierzchni w takich drzewostanach wzrasta gwałtownie do wysokich wartości po punkt kulminacyjny i w przeważającej mierze bywa akumulowana w biomasie lub energii (rys. 75).
Do rozpoczęcia takiej sukcesji pierwotnej potrzeba niewielkiej ilości martwej substancji organicznej (detrytusowej). Odpowiednio do tego, także respiracja przez destruenty na początku rozwoju lasu jest niższa. Ze wzrastającą akumulacją substancji organicznej rośnie masa organicznej materii odpadowej, co pociąga za sobą wzrost destruentów i ich respiracji (rys. 76).
2. Stadium lasu przejściowego
Zarówno między gatunkami, jak między osobnikami danego gatunku, występuje konkurencja o przestrzeń życiową, światło, wodę i składniki pokarmowe, która prowadzi do eliminacji słabiej przystosowanych osobników. Pod okapem w miarę ażurowego drzewostanu przedplonowego i w lukach po wcześniej obumarłych drzewach pionierskich zasiedlają się stopniowo klimaksowe gatunki drzewiaste o następujących właściwościach:
małe wymagania w stosunku do światła (szczególnie w młodości),
wrażliwość na ekstremalne warunki klimatyczne (mróz, spiekotę, suszę, wiatr), które często występują na otwartej przestrzeni,
duże wymagania w odniesieniu do gleby (większa zawartość próchnicy, dobra struktura, częste występowanie mikoryzy),
intensywny system korzeniowy,
wolny wzrost w młodości, późniejsza kulminacja przyrostu z niższymi wartościami maksymalnymi oraz utrzymywanie się przyrostu do późnej starości (rys. 75),
późniejsze dojście do dojrzałości generatywnej, mniej częste i nie tak obfite obradzanie nasion oraz mniejsza zdolność rozprzestrzeniania się nasion,
długowieczność, duże końcowe wartości wzrostu i duże zdolności gromadzenia przez organizmy żywe i biomasy, i energii.
Drzewostany są tu najczęściej dwupiętrowe. Górne piętro jeszcze z drzewostanu przedplonowego zbudowane jest w przeważającej mierze z gatunków pionierskich. Jest ono początkowo względnie gęste i bogate w osobniki, przerzedza się jednak z biegiem czasu i w trakcie rozwoju traci w wielu partiach zwarcie. Dolne piętro, rekrutujące się z bardziej wymagających gatunków drzew lasu końcowego, jest mniej więcej równowiekowe, niskie i początkowo bez znaczenia. Z wiekiem rośnie na wysokość i w znaczeniu, aż do panującego drzewostanu włącznie.
Przyrost na jednostkę powierzchni spada w drzewostanie górnym po przekroczeniu punktu kulminacyjnego, ponieważ w dolnym piętrze przyrost jest jeszcze mały (rys. 76).
Z postępującym wiekiem w piętrze górnym drzewostanu występuje duża śmiertelność drzew, znaczny opad materii organicznej i wysoka respiracja przez destruenty. Prędkość akumulacji stopniowo się zmniejsza.
3. Stadium lasu końcowego (klimaks)
W tym stadium gatunki pionierskie przedplonu dawno już przepadły, a drzewostan składa się ostatecznie z gatunków klimaksowych, których wymagania ekologiczne odpowiadają warunkom klimatycznym i edaficznym siedliska (klimaks edaficzny i klimatyczny). W zależności od żyzności siedliska drzewostan składa się z większej lub mniejszej liczby gatunków, mniej lub bardziej różnowiekowych i pionowo zróżnicowanych. Wykształca się charakterystyczny klimat wnętrza lasu (fitoklimat leśny) i rozwija się urodzajna gleba leśna.
Produkcja materii i wiązanie energii na jednostkę powierzchni zbliżone są do wartości średniej przyrostu osobników różnego wieku i różnej światłożądności w różnym wysokościowym usytuowaniu w drzewostanie. Zrzut martwej materii i jej rozkład przebiegają mniej lub więcej równomiernie na określonym poziomie produkcji organicznej.
Stadium klimaksowe jest znamienne przez to, że produkcja materii lub wiązanie energii i gromadzenie materii lub wydzielanie energii w przybliżeniu wyrównują się, tzn. pn = e, tak że iloraz z produkcji pierwotnej netto i emisji równa się prawie l, co przedstawić można:
![]()
Tylko przy takim założeniu można mówić o zrównoważeniu ekosystemu. Odchylenie od stanu równowagi prowadzi przy Q < l do zużywania biomasy i przy Q > l do gromadzenia biomasy.
Iloraz Q =![]()
- jest równocześnie miernikiem dojrzałości naturalnego ekosystemu leśnego lub podziałką osiągniętego stadium sukcesji. W młodszych stadiach wartością wskaźnikową jest liczba l, a z wzrastającym stopniem dojrzałości ekosystemu zbliża się do l (klimaks; rys. 76d).
Podsumowując powyższe wywody Thomasius (1980) przytacza w miejsce ogólnych wniosków wypowiedzi Margalefa (1962) zawarte w pracy o naturalnym rozwoju ekosystemów leśnych:
sukcesja jest w zasadzie zawsze związana ze zwiększaniem się różnorodności i wzmacnianiem struktury biocenozy i ekosystemu,
sukcesja jest procesem przerywanym, wywoływanym przez imigrację gatunków drzewiastych o zróżnicowanych właściwościach,
sukcesja jest związana z akumulacją biomasy, gdzie krótkowieczne gatunki o niniejszych zdolnościach gromadzenia biomasy wypierane są stopniowo przez gatunki długowieczne o dużych zdolnościach przyrostowych; w parze z tym idzie zwiększanie stabilności ekosystemu leśnego,
sukcesje są przykładem nieodwracalnych procesów, które prowadzą do sytuacji z minimalnym zapotrzebowaniem energii — minimum przy entropii, a maksimum przy informacji.
8. Zmiany historyczne. Historia naturalna lasów europejskich po ostatnim zlodowacenia
Do zmian historycznych zaliczamy głównie zmiany szaty roślinnej zachodzące pod wpływem minionych epok geologicznych. Cała Środkowa Europa po Alpy, Sudety i Karpaty nosi piętno czterech zlodowaceń, które wydatnie zubożyły skład gatunkowy naszych lasów. Las środkowoeuropejski jest bowiem stosunkowo ubogi w gatunki drzewiaste, liczy ich zaledwie 40, podczas gdy lasy północnoamerykańskie złożone są z ponad 250 gatunków (mimo znacznie silniejszego i rozleglejszego zlodowacenia), a lasy azjatyckie mają ich aż około 500. Tłumaczy się to budową, a właściwie usytuowaniem wysokich gór, które w Europie przebiegają równoleżnikowo, podczas gdy w Ameryce Północnej i Azji układają się południkowe. Góry w Europie stanowią jakby barierę, na której co prawda zatrzymał się lodowiec w swej wędrówce na południe, jednak równocześnie uniemożliwiły one ucieczkę roślin do strefy cieplejszego klimatu, a później utrudniły im powrót na uprzednio zajmowane stanowiska. Takiej przeszkody drzewa nie napotkały na swej drodze ani na kontynencie amerykańskim ani na azjatyckim.
Do zmian roślinności leśnej powodowanych klimatem zmieniających się epok geologicznych w ostatnim ich etapie dołączył się jeszcze jeden czynnik, człowiek, który swą działalnością, głównie w ostatnim tysiącleciu, zmienił nie tylko stan powierzchniowy lasów, ale także ich budowę i skład gatunkowy. Ten czynnik zadziałał silniej na przestrzeni kilku stuleci, aniżeli klimat w okresie dziesiątków tysięcy lat.
W trwającym obecnie interglacjale, zapoczątkowanym przed z górą 20 000 lat, stopniowe ocieplanie się klimatu spowodowało, że las na powrót zdobył Europę, jednakże uboższy o wiele gatunków trzeciorzędowych z rodzajów: cypryśnik (Taxodium sp.), tulipanowiec (Liriodendron sp.), magnolia (Magnolia sp.), orzech (Juglans sp.) itp. Gatunki te nie występują już w naszych lasach, przestały istnieć, wymarły bądź nie mogły przekroczyć bariery wysokich gór.
Zaraz po ustąpieniu lodowca odkryta gleba w wyniku naturalnej sukcesji pokryła się karłowatą roślinnością występującą dziś jeszcze na dalekiej północy w pobliżu kręgu polarnego. Nastąpiła faza bezdrzewnej tundry z dębikiem ośmiopłatkowym (Dryas octopetala) jako rośliną przewodnią. W miarę ocieplania się klimatu w miejsce tundry wkroczyła roślinność drzewiasta, której rozwój podzielić można na cztery okresy:
I. Okres sosny i brzozy
Okres ten, zwany preborealnym, miał miejsce przed 8000 lat p.n.e. Klimat był wówczas suchy i zimny. W tundrze zasiedliły się leśne gatunki pionierskie: brzoza, sosna, osika w towarzystwie wierzb. Okres ten łączymy ze starszą epoką kamienną z koczującymi jeszcze myśliwymi i rybakami.
II. Okres leszczyny
Klimat w dalszym ciągu się ocieplał, jednak nadal brakowało wilgoci. W okresie tym, zwanym borealnym, poza leszczyną pojawiły się dalsze gatunki drzewiaste: dęby, lipy i wiązy, stanowiące niewielką jeszcze domieszkę do lasów sosnowo-brzozowych. Ta tzw. średnia epoka kamienna miała miejsce w czasie 8000—5500 lat przed naszą erą.
III. Okres mieszanych lasów dębowych
Klimat tego okresu cechował się optymalną od czasów lodowca temperaturą. Średnia temperatura roczna kształtowała się o 2 do 2,5°C wyżej aniżeli dzisiaj i lasy sięgały w górach od 200 do 400 m wyżej niż obecnie. Wyższa temperatura przy jednocześnie większej wilgotności sprzyjała rozwojowi lasu, stąd okres ten nazwano atlantyckim. Znaczną część niżową Europy Środkowej zajmowały w tym czasie mieszane lasy dębowe z domieszką innych gatunków liściastych: wiązu, lipy i graba. Sosna zepchnięta została na słabsze, piaszczyste gleby. Buk, był jeszcze gatunkiem stosunkowo rzadkim, a jodła występowała tylko na krańcach zachodnich: w środkowej Francji, w zachodniej Szwajcarii oraz w Pirenejach i w Czarnym Lesie. Jodła i buk stały się panującymi gatunkami w lasach środkowoeuropejskich. Świerk nasuwał się stopniowo na północno-wschodnie regiony Europy oraz zajął tereny górskie w Karpatach i Sudetach, w Alpach i w Górach Harzu. Była to tzw. młodsza epoka kamienna, trwająca od 5500—2000 lat p.n.e.
IV. Okres buka
Po atlantyckim optimum klimatycznym nastąpiły okresy subborealny i subatlantycki, w których klimat ulegał stopniowo pogorszeniu — szczególnie wyraźnie obniżyła się temperatura osiągając w końcu wartości dzisiejsze. Jodła, która wcześniej zasiedliła Wogezy i Czarny Las, obecnie rozszerzyła swój zasięg na Alpy i Karpaty, osiągając na północnym wschodzie dzisiejsze granice zasięgu. Około 1000 lat p.n.e. zaczęła się ekspansja buka, a jej kulminacja przypada około 500 lat p.n.e., tj. w ostatnim okresie prehistorycznego rozwoju lasów w Europie Środkowej. Świerk na północy Europy opanował Skandynawię i zajął północno-wschodnie tereny Polski. W górach gatunek ten utrzymał się w wyższych położeniach, gdyż położenie średnie (regiel dolny) opanował buk. Jest to tzw. epoka brązu i żelaza, rozpoczynająca się około 2000 lat p.n.e. i trwająca po dzień dzisiejszy.
Europa Środkowa znajduje się teraz w drugiej fazie interglacjału, w stale i stopniowo ochładzającym się klimacie, prowadzącym w kierunku następnego zlodowacenia. Ponadto silne wylesienia kontynentu doprowadziły do osuszenia i skontrastowania klimatu, czyli do jego kontynentalizacji nie sprzyjającej rozwojowi jodły.
Aczkolwiek wpływ człowieka na las zaznaczył się już w młodszej epoce kamiennej, to jednak destruktywne oddziaływanie czynnika ludzkiego, wpływające na kurczenie się areału leśnego, przejawiać się zaczyna wyraźnie dopiero od epoki brązu, kiedy to koczujące plemiona przeszły na osiadły tryb życia, połączony z uprawą roli oraz z hodowlą bydła i trzody.
Równocześnie z wylesieniem terenów pod uprawę rolniczą postępowały zmiany w budowie i składzie gatunkowym lasu. Na przykład Niemcy z leśnego ongiś kraju wskutek olbrzymich karczunków realizowanych w ostatnich wiekach zmieniły się nie do poznania, gdyż lesistość tego kraju z około 75% zmalała szybko do 25%, tj. mniej więcej do poziomu dzisiejszego. Podobnie wyglądała sytuacja w Polsce.
Inne było podejście człowieka do użytkowania lasu liściastego, a inne do iglastego. W pierwszym, po ścince drzew następowała naturalna jego regeneracja przez rozwijanie odrośli z pniaków. Odroślą te wraz z bujnym zielem i trawą stanowiły doskonałą paszę dla bydła. Taki pastwiskowy las liściasty, często krzaczasty i niskopienny, stanowił najpierwotniejszą formę lasu gospodarczego.
Z kolei rosnące zapotrzebowanie na budulec pokrywane było z wysokopiennego lasu iglastego, który jednak użytkowany był plądrowniczo, przez bezplanowe wycinanie pojedynczych drzew zależnie od potrzeby i wymiarów. Nie troszczono się zupełnie o jego odnowienie, które następowało i było możliwe w tym przypadku tylko na drodze samosiewu.
Człowiek karczując lasy, wypasając bydło w lesie i zmieniając sztucznie skład gatunkowy, w stosunkowo krótkim czasie przekształcił pierwotne lasy mieszane porastające Europę Środkową w drzewostany lite, w szczególności zaś lasy liściaste bądź mieszane z dużym udziałem gatunków liściastych na wielkopowierzchniowe monokultury sosnowe i świerkowe. Człowiek zmienił całkowicie obraz lasów europejskich, będących wynikiem wielowiekowej, ewolucyjnej sukcesji, realizującej się w naturze pod przemożnym wpływem zmian, jakim podlegał klimat w okresie tysięcy lat (rys. 81).
Wykład - Las jako zjawisko geograficzne
1. Ekologiczne formy i typy lasu wg ich rozmieszczenia poziomego
W różnych szerokościach geograficznych i na różnych kontynentach lasy różnią się od siebie tworząc liczne formy i typy.
Czynniki limitujące:
klimat (suma temperatur efektywnych) - zmieniają się od równika do biegunów - wpływa na równoleżnikowy układ lasów
opady i wilgotność - kierunek od brzegów do środka kontynentów im bliżej tym bardziej suche
stopień wykształcenia gleby i jej żyzność
1. Tropikalny las deszczowy - dżungla
strefa przyrównikowa +- 10° = Indie, Ameryka Środkowa, Australia, Afryka
opady min. 1600 mm śr. 2-3 tys. max. 16 tys. - Hawaje (równomierne w ciągu roku)
małe roczne wahania temperatur - max 6°C przy temp 22 - 28°C
gleby laterytowe czerwone i żółte, duża zawartość gliny, brak części organicznych - intensywny rozkład.
3-9 tys. gatunków pędowych, drzew ok. 2500, najwięcej w regionie malajsko-indonezyjski, najmniej w Afryce.
5 warstw roślinnych
1) najwyższe pojedyncze drzewa 40-60 m luźne
2) i 3) drzewiaste, zwarte korony: są liany i epifity
4) częściowo drzewiaste i wysokie zioła
5) zioła i młode drzewiaste
1% światła dochodzi do warstwy dna - półmrok
niezbyt gęsta roślinność dna
roślinność - brak pór roku - brak słojów rocznych
kwitnienie i owocowanie - periodyczność, lecz dżungla ciągle kwitnie i owocuje, wiecznie zielona
kora - gładka, jasna i cienka
ulistnienie - skórzaste, szarozielone
system korzeniowy płytki
produktywność ogólna produkcja suchej masy 500-600 t/ha
roczny przyrost masy zielonej 12-35 t/ha/rok
przyrost surowca drzewnego zbliżony dżungli i lasu bukowego europejskiego ok. 8 t/ha, występują straty na oddychanie
roczny przyrost grubizny mały (1m3/ha)
mahonie i palisandry.
1a. Tropikalny las deszczowy górski
wzrost wysokości do 3500 - spadek temperatury, opadów ok. 2000.
mniej gatunków, głównie paprocie drzewiaste
niższa wysokość lasu, mniej pięter lasu (4)
część zrzuca liście
2. Lasy tropikalne zielone w porze deszczowej - lasy monsunowe
monsuny - stałe wiatry wiejące na przemian w przeciwnych kierunkach
monsun lądowy - zimny - susza - 4-6 miesięcy
monsun morski - letni - deszcz
występują w pł-wsch. Azji, Afryka Równikowa, płn. Australia
a) lasy niższe niż dżungla, świetliste, mniej pięter, zrzucają liście
b) najniższe piętro gęste zioła i trawy - bambus, mniejsze liście, słoje
c) lasy wysokoprodukcyjne, hodowla, 6-10 m3/ha/rok, drzewo tikowe i sal
3. Tropikalne lasy suche
suche gleby i klimat - opady 700-1500 mm przy suszy 6-10 miesięcy
roślinność niższa max 20-25 m, gruba kora, drewno twarde lub zastąpione gąbczastą tkanką magazynującą wodę, korony spłaszczone, liście duże opadające lub małe skórzaste wiecznie zielone, liczba drzew na ha też mniejsza
występowanie: wsch i płd-zach Afryka, Indie centralne, Brazylia i Ameryka Środkowa i Australia
w Australii - Eucalyptus, w Afryce - akacje
znaczenie gospodarcze - drewno opałowe
formy:
a) las sawannowy
głównie trawy i rzadkie kępy drzew i krzewów kserotermicznych
Afryka - baobaby
Ameryka płd i Australia, Azja b. rzadko
znaczenie krajobrazowe
b) namorzynowy - zarośla mangrowe
płytkie brzegi mórz krajów tropikalnych
lasy lub zarośla rosnące w wodzie morskiej
zasięg - strefa przybrzeżna - przypływów morskich
halofity - słonorośla - zasolenie - liście są zrzucane, owłosione
występowanie: wsch i płd wsch Azja, płd Japonia, płw Synaj i płn Nowej Zelandii
drzewostany ok. 10-20 m wys.
korzenie powietrzne - szczudlaste
żyworodność - siewki rozwijają się z nasion na drzewach do kilku cm wbijają w szlam i zakorzeniają
użytkowanie w Azji - 30-letnia rębnia
c) tropikalne lasy iglaste
górskie tereny klimatu ciepłego, gleby ubogie
płd-wsch Azja i Ameryka Środkowa
głównie sosna: Pinus caribea, insularis, merkusii, oocarpa, Araukarie, Podocarpus
użytkowanie drewno i żywica, odnowienia naturalne i sztuczne
d) lasy bambusowe
60 rodzajów i ok. 700 gatunków
niskie gatunki 10-15 cm i wysokie do 30 m wzrost kilka m-cy
gleby dobre i przewietrzane
niziny, wymagają wys. temperatur i równomiernych opadów
użytkowanie na plantacjach na grunty wykarczowane i rolne
e) lasy galeriowe
wzdłuż rzek które zaopatrują je w wilgoć
roślinność podobna jak w dżungli
Afryka środkowa i Ameryka Płd.
f) wtórne lasy tropikalne
pod wpływem człowieka
obiegowy wyrąb lasu - eksploatuje się najcenniejsze gatunki - wypalenie pozostałości - pojawienie się nowej biocenozy z gatunkami światłożądnymi, typowy dysklimaks, załamanie naturalnej sukcesji w wyniki czego uzyskuje się gatunki cenniejsze dla człowieka lecz niekorzystnie wpływa na glebę i roślinność
powszechnie stosowana w tropiku
g) lasy deszczowe ciepłej i umiarkowanej strefy klimatycznej
bogate i równomierne opady, ciepło, gleby dobre,
sklepienie luźne, świetliste, bujny wzrost roślinności zielnej, paproci palm, bambusów, niskich krzewów, ziół i traw
płd i wsch USA - dąb wirginijski i Magnolia grandiflora
Japonia, Korea i wsch Chiny też i sosna na glebach wilgotnych Taxodium, zielone dęby i magnolie
były też w Basenie Śródziemnomorskim
4. Twardolistne lasy ciepłej i umiarkowanej strefy klimatycznej
klimat: gorące i suche lata, krótkie, łagodne zimy - klimat łagodny
występowanie: Basen Morza Śródziemnomorskiego, płd Afryka, Kalifornia, Chile, Australia
roślinność; drzewa: 15-25 m wys. luźna zwartość, krzewy, bogate runo najniższa strefa, zioła byliny, podkrzewy, grube skórzaste liście
typowe: lasy oliwne i gaje cyprysowe
Basen Morza Śródziemnego - od Hiszpanii do Izraela w drzewostanach duży udział sosny: Pinus Halepensis, p. Bruttia, p. pinea
dęby (dolne piętro: Quercus ilex i Q. cocifera)
dąb korkowy Q. suber - Hiszpania, Portugalia
inne: rodzime i aklimatyzowane: kasztan jadalny, figi, palmy, drzewa poziomkowe, drzewiaste wrzosy, złotogłowy, oleandry,
wprowadzone: agawy, opuncje, pomarańcze, cytryny
zbocza gór: lasy usunięte, wypas, busz zarośli - makchia
pow. 800 m n.p.m. są lasy wysoko i niskopienne z kasztanem - są podobne do lasów środkowoeuropejskich liściasto - iglastych
śr. wys. - 40 m, zapas drewna 1250 m3/ha - Korsyka, śr. Włochy, Grecja
obecnie odbudowa lasów naturalnych w tej strefie
5. Lasy letnie - lasy liściaste umiarkowanej strefy klimatycznej zrzucajęce liście na zimę
występowanie: Europa Zach, Anglia, płd - wsch USA, Azja (Chiny i Japonia)
opady 700-1500 mm temp. - 5°C styczeń do + 20°C lipiec
wegetacja ok. 6- m-cy. susza fizjologiczna zimą - zrzucanie liści
las ten dzieli się na 2 strefy lasu cieplejsza Castanetum i chłodniejsza Fagetum
roślinność: gat lasotwórcze dęby i buk
iglaste sosny i świerki, bogaty podszyt i runo
aspekty sezonowe: wczesna wiosna (Anemone, Pulsatilla, Hepatica, Pulmonaria) i lato paprocie, widłaki, skrzypy i byliny
wys. drzew 25-30 m, 2-3 piętra, dobrze zwarte, najczęściej 1-piętrowe i jednogatunkowe
pierwotnie lasy bukowe w Bośni i dolne regle Karpat
próchnica nakładowa 10 - 20 t/ha
przyrost 10 - 15 m3/ha/rok
produkcyjność 5-8 m3/ha
6. Lasy mieszane liściasto - iglaste umiarkowanej strefy klimatycznej oraz lite lasy iglaste
występowanie: Europa płn i środk. Azja wsch. i częściowo Ameryka Płn
gorsze warunki cieplne i krótszy okres wegetacyjny
Ameryka płn - lasy sekwojowe z udziałem daglezji i thuji olbrzymiej, jodły olbrzymiej sekwoje mają wiek kilka tys. lat wys. do 110 m i średnica 10 m
Europa - lasy naturalne - przekształcone gospodarką człowieka w lasy iglaste
wprowadzone gatunki: sosna, jodła świerk, gat. szybkorosnące
rezerwaty i parki narodowe.
7. Lasy iglaste chłodnej strefy klimatycznej
występowanie: Europa płn, Azja i Ameryka Płn. 1/3 pow. leśnej świata to lasy iglaste.
przechodzi w tundrę najw. skupiska Azja i Ameryka Płn od Uralu do Oceanu Spokojnego ok. 5 tys km i tys km szerokości, aż za krąg polarny
gatunki drzew - sosna - siedliska ubogie: świerk, jodła syberyjska, modrzew syberyjski i limba, siedliska bogatsze, olsza i brzoza - podmokłe i jako towarzyszące
drzewa iglaste nie zrzucające liście, asymilacja wczesną wiosną i późną jesienią maksymalne wykorzystanie okresu wegetacyjnego
drzewa iglaste mniej narażone na śniegołomy
płd obszary tego typu lasu, duża produkcyjność, zapas grubizny
śr. 1000 m3/ha - max do 1500 m3/ha - stare drzewostany świerkowe najbardziej produkcyjne
inklinacja do jednogatunkowości
silne ocienienie gleby, zakwaszenie, runo ubogie
trudny rozkład materii organicznej
najbardziej wartościowe lasy.
Wykład - Las jako zjawisko geograficzne
2. Ekologiczne typy lasu wg rozmieszczenia pionowego
a) Zależność formacji roślinnej z wysokością n.p.m
Analogicznie jak w kierunku poziomym typy lasu zmieniały się przy posuwaniu od równika ku biegunom tak i w kierunku pionowym obserwuje się zmiany związane ze skracaniem okresu wegetacyjnego i pogarszaniem warunków glebowych. Za tym idzie zmienność roślinności leśnej w górach.
Humboldt zauważył że zmiany w roślinności wraz z wysokością odpowiadają tym które zachodzą wraz z szerokością.
Prawo Humboldta (1802 r.): ciepło zmniejszające się z wysokością n.p.m. i szerokością geograficzną wywołuje jednokierunkowe zmiany w szacie roślinnej uwidaczniające się w podobnym typie,
Nie jest zawsze prawdą, znaczenie ma także wielkość masywu górskiego, usytuowanie gór względem stron świata, prądy powietrzne i morskie, a głównie wpływ klimatu morskiego czy kontynentalnego. Granice typów roślinnych w górach wraz ze wzrastającą szerokością obniżają się aż na północy granica formacji leśnej schodzi do punktu zerowego n.p.m. i zbiega się z analogiczną dla stref poziomych.
Granica lasu na przykładzie buka:
Kilimandżaro, Himalaje 3500-4000 m.n.p.m.
Etna - Sycylia 37° szer. geogr. - wys. 1970 m.n.p.m.
Alpy Tyrolskie 46° szer. geogr. - wys. 1540 m.n.p.m.
Góry Harzu 52° szer. geogr. - wys. 650 m.n.p.m.
Norwegia 59° szer. geogr. - wys. 190 m.n.p.m.
b) Leśne piętra roślinności w górach w Polsce.
Zmiany w typach lasu jaki obserwujemy w miarę wznoszenia się nad poziomem morza.
1. piętro pogórza (Próg Sudecki Zgorzelec, Sobótka, Racibórz, próg Karpacki Zebrzydowice, Kraków, Rzeszów, Przemyśl -, wysokość 400-600 m.n.p.m.
Lasy mieszane z dużym udziałem gatunków typu liściastego dąb, grab, buk, sosna, jodła, charakterystyczny typ lasu to grąd
2. piętro regla dolnego - kraina buka do wysokości 1000-1250 m , typowy Fagetum, dominuje buk, domieszka jodła, świerk, jawor, wiąz, jesion, osika;
XVII/XIX w. duża przebudowa na rzecz świerka i jodły
obecnie buczyny stanowią rezerwaty ścisłe lub częściowe
problem w Sudetach - regiel dolny opanowany został przez świerczyny które uszkadzane są przez owady i grzyby - wymaga przebudowy.
3. piętro regla górnego - kraina świerka - Piceetum do wysokości 1250-1550 m dominuje świerk inne są domieszką. Z wysokością ustępuje jodła, modrzew, dorównuje limba i jarząb jako domieszka, limba w Tatrach, w Sudetach gatunek obcy.
4. piętro kosodrzewiny - sosna kosówka
powyżej górnej granicy lasu 1450-1800 m występuje w Sudetach, w Karpatach - brak w Bieszczadach - tu strefa zajęta przez olszę zieloną
znaczenie gospodarcze - zatrzymują lawiny śnieżne i podwyższają górną granicę lasu
zniszczona przez ludzi - łąki dla wypasu owiec
5. piętro hal wysokogórskich - piętro alpejskie
Sudety tylko Śnieżka 1603 m, Beskidy tylko Babia Góra - 1725, Tatry do wysokości 2300 m.
roślinność hal upodabnie je do tundry - dębik ośmiopłatkowy i wierzby karłowate i płożące.
6. piętro turni - granica wiecznych śniegów
>2300 m, tylko Tatry, nagie strome skały
mchy i porosty, skalnice, szarotka
3. Granice lasu
Las jest potężną i dynamiczną formacją roślinną która wykazuje w stosunku do czynników środowiskowych duże wymagania i pod wpływem zbyt wysokiej lub niskiej temperatury, niedoboru lub nadmiaru wilgoci, jałowej gleby itp. może zostać wyparty lub zastąpiony przez inne formacje roślinne.
cechy lasu krańcowego:
zmniejszenie się liczby gatunków budujących las, gatunki graniczne na płn: sosna zwyczajna, świerk pospolity, świerk syberyjski, modrzew syberyjski, brzoza brodawkowata; w górach europejskich gatunki graniczne: sosna karłowata, sosna limba, świerk pospolity, modrzew europejski, jarząb pospolity, po nich gatunki krzewiaste salix, alnus, vaccinium.
wzrost drzew na wysokość jak i grubość wyraźnie maleje,
zagęszczenie drzewostanu zmienia się (głównie maleje), przyczyną jest głównie susza (rzeczywista i fizjologiczna) i skład gleby.
a. Granica lasu powodowana niską temperaturą powietrza (granica chłodu)
granica lasu pokrywa się na półkuli północnej z izotermą lipca = 10°C i na terenie równinnym do 22°szer.
wraz z obniżaniem się temperatury towarzyszą:
maleje produkcja materii organicznej
pogorszenie warunków egzystencji
mniejsza odporność na szkody
mniejsze obradzanie
osłabienie siły regeneracyjnej
ma miejsce gwałtowny przeskok od zwartych wysokich drzewostanów do słabych zarośli leśnych „strefy walki” - nie wyjaśnione
w Beskidzie Żywieckim świerk osiąga pierśnicę 1,3 m na wysokości
600 m.n.p.m. - 10 lat
1300 m.n.p.m. - 20 lat
1300 -1400 m.n.p.m. - 30 lat
b. Granica powodowana fizjologiczną suszą mrozową
istotą zjawiska jest mrozowa susza (fizjologiczna) wynikająca z wysokiej już ciepłoty igliwia i zmarzniętej jeszcze gleby. Stan gleby nie pozwala na uzupełnienie braków wilgoci w pędach nad glebą = efekt obumierania pędów
szkody od suszy mrozowej zależą od:
zabezpieczenia drzew przed nadmierną transpiracją
możliwości gromadzenia wody w tkankach
stopnia podatności pędów wzrostowych na obsychanie pod wpływem wiatrów.
c. Granica lasu powodowana suszą rzeczywistą
wywołana - nikłymi opadami, płytką glebą, płytkie systemy korzeniowe są silniej narażone na wahania wilgoci w glebie.
d. Granica lasu powodowana nadmiarem wilgoci.
przyczyna: stagnujące wody bagienne pozbawione tlenu. Czynnik ograniczający to brak tlenu.
częściowo znoszą to wierzby i olchy.
4. Las krańcowy na przykładzie lasu górskiego
Trudno znaleźć wyraźną granicę lasu, na pewnym pasie ma miejsce stopniowe zmniejszanie przeciętnej wysokości oraz oddalanie się poszczególnych egzemplarzy czyli rozluźnianie zwarcia i karlenie.
Stosunki zwarcia i wysokości drzewostanu wyróżnia się idąc ku górze:
pas zwartego lasu
pas luźnego drzewostanu (świerki osiągają ok. 8 m. wys.)
pas świerków pojedynczych wys. 8 m
pas grup karłów tj. świerków niższych od 8m, nie pokrzywionych
pas krzywulców tj. małych pokrzywionych drzew.
Czynniki które decydują o położeniu wysokościowym górnej granicy lasu.
Położenie geograficzne (szerokość geograficzna) - wraz z oddalaniem od równika górna granica lasu obniża się, w okolicach polarnych schodzi do poziomu morza.
Wielkość masywu górskiego (zmiany klimatyczne). W dużych kompleksach górskich granica lasu podnosi się wyżej aniżeli w małych izolowanych kopcach górskich.
Ekspozycja (wystawa stoków). Stoki południowe są cieplejsze, więc granica lasu na ogół jest wyżej (wyjątek Tatry Wysokie - zmienia to kierunek wiatru)
Nachylenie stoków. Strome urwiste nie sprzyjają rozwojowi lasów. Spychają go niżej. Współdziała tu wiatr i różnicuje sytuację w zależności od stopnia nachylenia stoków odwietrznych.
Wiatr. W górach nabiera szczególnego znaczenia. Wywiera ujemny wpływ na zasięg lasu na stokach odwietrznych. Na zboczach dowietrznych podnoszą się i las ku górze, na zboczach odwietrznych (wschodnich) będą obniżać granicę lasu.
Konkurencja między zbiorowiskami biotycznymi na styku piętra leśnego regla górnego i piętra kosodrzewiny istnieje strefa walki. Kosodrzewina ma mniejsze wymagania glebowe i wilgotnościowe jest lepiej przystosowana do surowego klimatu może zejść poniżej klimatycznej granicy lasu i zahamować mu dojście do wysokości.
Człowiek. (wyręby, karczunki, wypas bydła, utrzymywanie zwierzyny) - obniżenie i zdeformowanie granicy -zagrożenie lawinowe
Wykład - Las jako obiekt zainteresowań człowieka
2. Ekologia populacji drzewiastych
l. Pojęcie drzewostanu
Najwyższą i najważniejszą warstwą w zespole leśnym, jest warstwa drzew, która z ekologicznego punktu widzenia warunkuje istnienie pozostałych warstw.
„Drzewostanem nazywamy ogólnie zespół (skupienie, zbiór) drzew tworzących główny składnik leśnej szaty roślinnej, które rosnąc w bliskim sąsiedztwie, wzajemnie oddziałują na siebie waz z innymi komponentami lasu kształtują w specyficzny sposób warunki środowiska leśnego, a przy tym same w swym indywidualnym i zespołowym życiu pozostają pod wielostronnym wpływem tego środowiska".
To specyficzne, wzajemne uwarunkowanie powstaje dzięki przemożnemu wpływowi wysokich i długowiecznych drzew na kształtowanie się warunków klimatycznych i glebowych w lesie.
Podstawowe cechy mikroklimatu wnętrza drzewostanu.
Sklepienie koron drzew, tworzących zazwyczaj mniej lub więcej gęsty pułap, oddziela warstwę powietrza ponad lasem od warstwy znajdującej się poniżej sklepienia koron, w obrębie pozbawionych gałęzi pni drzew.
RYC Kontaktowanie się wolnej atmosfery z powietrzem wnętrza lasu dokonuje się nie tylko poprzez ażurowe sklepienie koron, lecz także z boku, od strony otwartej powierzchni graniczącej z lasem. Las przeciwstawia się również bocznym wpływom atmosfery, wykształcając na obrzeżu tzw. okrajek leśny, zbudowany z zewnętrznego szeregu drzew o gęstych i nisko osadzonych koronach, sięgających często do samej ziemi. W ten sposób strefa zieleni, utworzona przez okrajek i sklepienie koron drzew przedstawia się jako „zamknięta czasza" albo odwrócone naczynie, zamykające w swym ażurowym wnętrzu część wszechobecnej atmosfery.
Z powodu wspomnianego silnego osłabienia kontaktu z wolną atmosferą powietrze znajdujące się wewnątrz lasu staje się swoistym tworem fizykalnym, o odmiennych parametrach nasilenia poszczególnych czynników klimatycznych.
Powietrze wnętrza lasu charakteryzuje się:
nieco większą wilgotnością względną,
większą zawartością CO2,
niższą i bardziej wyrównaną temperaturą,
małą i do tego silnie skontrastowaną ilością światła (mozaiką świetlną)
oraz przede wszystkim zaciszną atmosferą, co stanowi jedną z najważniejszych cech klimatu lasu.
Ponieważ główną przyczyną tych zmian jest roślinność leśna, można ten specyficzny układ warunków klimatycznych, panujących wewnątrz lasu, nazwać fitoklimatem leśnym.
Pojęcie okrajka leśnego oraz ściany i brzegu drzewostanu.
Drzewa obrzeżne tworzące okrajek leśny, wykształcają charakterystyczny pokrój (habitus), wskutek graniczenia z powierzchnią pozbawioną roślinności leśnej RYC. O ile budowa części nadziemnej drzewa kształtowana jest tu głównie jednostronnym lepszym oświetleniem, o tyle jego część podziemna jest formowana brakiem konkurencji od strony pola i bardzo dużą konkurencją o wodę i pokarmy od strony wnętrza lasu. To nierównomierne oddziaływanie światła nad glebą i konkurencji korzeniowej w glebie staje się przyczyną asymetrii w pokroju tego drzewa. Od strony pola wykształca ono nisko osadzoną koronę oraz silnie rozwija system korzeniowy. Od strony lasu natomiast, wskutek bocznego ocienienia, korona utrzymuje się tylko na wierzchołku drzewa - również system korzeniowy jest wykształcony wyraźnie słabiej; drzewo upodabnia się od strony otwartej przestrzeni do drzew wyrosłych w odosobnieniu, od wnętrza lasu przybiera natomiast pokrój typowy dla drzew wyrosłych w silnym zagęszczeniu przy ograniczonym dostępie światła.
Niezależnie od asymetrii w budowie korony, drzewo okrajkowe, podlegając fototropizmowi oraz nierównomiernemu obciążeniu koroną, wychyla się wyraźnie w kierunku pola, skąd otrzymuje nieograniczoną ilość pełnego światła.
Duża i asymetrycznie zbudowana korona drzewa okrajkowego przyczynia się również do wykształcenia grubszego, aczkolwiek bardziej zbieżystego pnia w porównaniu z drzewami rosnącymi wewnątrz lasu. Silne, jednostronne ugałęzienie drzewa okrajkowego powoduje nierównomierny obieg wody i asymilatów, co przejawia się na przekroju poprzecznym RYC, który jest eliptyczny z mimośrodowo usytuowanym rdzeniem.
Przesunięcie rdzenia w kierunku lasu wywołane jest lepszym odżywieniem i bujniejszym przyrostem drewna od strony dużej powierzchni asymilacyjnej drzewa, co uzewnętrznia się nakładaniem szerokich słojów rocznych od strony pola i wąskich słojów od strony lasu.
Tak pokrój, jak i budowa wewnętrzna drzew okrajkowych sprawiają, że okrajek leśny spełnia ważną funkcję osłony przeciwwiatrowej lasu. Składają się na to następujące elementy:
1) drzewa są wychylone w kierunku otwartej przestrzeni, a więc przeciw wiatrowi (może się jednak zdarzyć, że od strony działania stałych wiatrów mogą wykazywać wychylenie przeciwne, jak inne drzewa rosnące wewnątrz drzewostanu - w tym przypadku działanie wiatru równoważy bądź przewyższa fototropizm);
2) duże, jednostronne i nisko osadzone korony drzew okrajkowych oraz związana z tym zbieżystość pnia sprawiają, że ich środek ciężkości jest nisko umiejscowiony, co utrudnia wywalenie ich przez wiatr;
3) dzięki wspomnianej większej zbieżystości pni drzewa okrajkowe są znacznie elastyczniejsze - łatwiej się gną aniżeli łamią na wietrze;
4) eliptyczny przekrój pni tych drzew usztywnia je w kierunku głównego działania wiatru;
5) siłom łamiącym wiatru przeciwdziałają pewne właściwości fizyczne drewna drzew okrajkowych, w szczególności duża wytrzymałość na rozrywanie (rozciąganie) partii drewna szerokosłoistego (wykształconej od strony pola) i znaczna wytrzymałość na zgniatanie drewna wąskosłoistego (odkładanego przez drzewa od strony wnętrza drzewostanu);
6) silne zakotwiczenie tych drzew przez długie korzenie rozwinięte w kierunku pola, uniemożliwiające wyważenie i obalenie ich przez wiatr w stronę drzewostanu.
W ten sposób okrajek leśny nie tylko chroni wnętrze lasu od wpływów powietrza atmosferycznego spoza lasu, lecz także zabezpiecza mało odporne drzewa wnętrza lasu przed wywalającym lub łamiącym działaniem wiatru (przy wierzchołkowych koronach i wąskich systemach korzeniowych drzew wyrosłych w zagęszczeniu moment siły wywalającej wiatru jest dla lasu bardzo niekorzystny).
Okrajki z tego samego gatunku co planowany drzewostan lity powstają analogicznie jak uprawa, w której od pierwszych zabiegów dąży się do tego, by osiągnąć większe odstępy drzew obrzeżnych. Jeśli przy zakładaniu uprawy od razu posadzi się drzewa obrzeżne w większych odstępach, trzeba się liczyć z większym zachwaszczeniem. Nawet większe wypady wśród drzew obrzeżnych, szczególnie gdy zastosowano więźbę trójkątną, nie mają większego znaczenia, gdyż drzewa rosnące za okrajkiem funkcjonalnie podporządkowują się okrajkowi. Również przy zabiegach pielęgnacyjnych w fazie młodnika, gdy wykształcony został jeden rząd drzew okrajkowych, należy zwrócić baczną uwagę na drzewa za nim rosnące, by przystosowały się do roli wspomagającej, aby powstający okrajek rozwinął się w gęsty, zamknięty, tylko od pola ugałęziony „zielony mur" (RYC).
Jeśli na obrzeżu drzewostanu iglastego ma być posadzony rząd drzew liściastych, co może mieć miejsce równocześnie z uprawą lub nieco później, to należy się liczyć, niezależnie od wyższych kosztów samych sadzonek, również z większymi nakładami pracy i kosztów podczas pielęgnacji. Tworząc okrajki z innego gatunku niż drzewostanu, z punktu widzenia lepszej ochrony drzewostanu i większej różnorodności krajobrazowej, nie należy zapominać jednak o krajobrazowo-estetycznej roli również tych gatunków, z których zbudowany jest sam drzewostan (RYC).
O odrębnościach fitoklimatycznych decyduje także wielkość powierzchni zajmowanej przez las. Małe skupienie drzew w dużo mniejszym stopniu modyfikuje układ czynników klimatycznych aniżeli skupienie duże, zajmujące większą powierzchnię.
Z okrajkiem, a ściślej rzecz biorąc z granicą lasu z polem lub innym środowiskiem nieleśnym, wiążą się dalsze dwa pojęcia: ściana i brzeg drzewostanu (RYC).
Ściana drzewostanu (RYC) jest to boczna powierzchnia styku lasu z jego sąsiedztwem. W zależności od charakteru sąsiedztwa wyróżniamy:
l) ścianę odkrytą, gdy drzewostan bezpośrednio graniczy z polem (powierzchnią otwartą) lub
2) ścianę zakrytą, jeśli w bezpośrednim sąsiedztwie rozpatrywanego drzewostanu rośnie inny drzewostan, odróżniający się wyraźnie taką czy inną charakterystyczną cechą, np. wiekiem lub gatunkiem drzew.
Odkrytą ścianę drzewostanu tworzy zazwyczaj okrajek leśny, aczkolwiek w lasach zagospodarowanych w miejscach graniczenia starego drzewostanu z powierzchnią zrębową lub uprawową spotkać się można z nieuodpornioną ścianą odkrytą.
Zakryta ściana drzewostanu z reguły nie ma wykształconego okrajka (RYC). Ściana drzewostanu (lasu) w sposób istotny zmienia warunki ekologiczne przyległej powierzchni. Rzuca na nią cień, co w głównej mierze zależy od jej ekspozycji, zatrzymuje wiatr, zmienia także stosunki termiczne oraz wilgotność powietrza i gleby, zarówno przez bezpośrednie oddziaływanie roślinności leśnej, jak i pośrednio poprzez wymianę między powietrzem atmosferycznym w lesie a powietrzem zalegającym ponad koronami drzew. Stąd duże znaczenie ekologiczne ściany lasu, również jako niszy ekologicznej dla zwierząt (gryzonie).
Całkowicie ochronny wpływ ściany lasu na przyległą powierzchnię przejawia się na stosunkowo wąskim pasie, szerokości równej połowie wysokości drzewostanu. W praktyce hodowlanej wykorzystuje się ten wpływ dla wprowadzenia na przyległą uprawę gatunków wrażliwych (np. buka, jodły, dębu itp.), a przy odnowieniu naturalnym, głównie dla samosiewnego odnowienia świerka, zasiedlającego chętnie otwartą powierzchnię przy północnej ścianie lasu.
Brzeg drzewostanu, obejmujący po obu stronach skrajnego rzędu drzew obrzeżną partię lasu szerokości 5-10 m, może być otwarty lub zamknięty (RYC).
Otwarty brzeg drzewostanu występuje często w lasach gospodarczych, głównie tam, gdzie gospodaruje się zrębami zupełnymi, najczęściej w jednopiętrowych i jednogatunkowych lasach niżowych. Cechuje się on brakiem wykształconego okrajka i wysoko osadzonym sklepieniem koron drzew, które rosnąc gęsto na zajmowanej powierzchni uformowały wysokie i pozbawione gałęzi pnie. Pozbawiona podszytów i podrostów przestrzeń między glebą leśną a pułapem koron nie stanowi żadnej zapory dla wnikania do wnętrza lasu wiatru, światła oraz innych wpływów z sąsiadującej z lasem otwartej powierzchni. Otwarty brzeg drzewostanu nie zabezpiecza lasu przed szkodami ze strony wiatrów i nie sprzyja powstawaniu w jego wnętrzu specyficznego klimatu (RYC)
Zamknięty brzeg drzewostanu (RYC) z wykształconym okrajkiem oraz wypełnioną koronami drzew i podszytów partią brzeżną lasu, sięgającą od ściany w głąb drzewostanu co najmniej na odległość równą połowie wysokości starych drzew (10 m i więcej), zabezpiecza w znacznym stopniu drzewostan przed szkodami powodowanymi przez wiatr i przyczynia się do powstawania w jego wnętrzu zacisznej atmosfery.
Gospodarz lasu powinien dążyć do tego, by las na granicy z polem miał zamknięty brzeg drzewostanu. Osiąga się go w drodze celowej pielęgnacji obrzeżnej partii drzewostanu już od najmłodszych lat, sadząc na brzegu lasu gatunki o znacznej odporności na działanie wiatrów, wykształcające głębokie systemy korzeniowe i gęste korony. W dalszym ciągu formuje się brzeg drzewostanu za pomocą odpowiednich cięć. W partii brzegowej przeciwdziała się zwarciu i zamieraniu dolnych gałęzi w drodze systematycznego rozluźniania sklepienia koron. W ten sposób drzewa zachowują nisko osadzoną koronę przez długi okres życia i wypełniają biomasą całą przestrzeń brzegu drzewostanu (RYC).
Z wiekiem, nawet przy silnym rozluźnianiu zwarcia, dochodzi jednak do obumierania dolnych gałęzi i to tym szybszego, im większa jest światłożądność rosnących na brzegu gatunków. W takim przypadku koniecznością staje się wprowadzanie podszycia w celu zamknięcia otwierającego się od dołu brzegu drzewostanu.
Przy klasyfikacji brzegów lasu należy uwzględniać:
1) zajmowaną powierzchnię (jednorzędowe okrajki i wielorzędowe brzegi smugowe),
2) stan ulistnienia (wiecznie zielone i zielone tylko latem),
3) stosunek gatunków brzegowych do gatunku tworzącego drzewostan (brzegi z tego samego gatunku co drzewostan i z gatunków innych, jak: brzoza, osika, dąb lub buk są odmiennie ukształtowane, spełniają różne funkcje i wymagają innego podejścia pielęgnacyjnego).
Szczególne znaczenie mają zamknięte brzegi drzewostanów na ekspozycjach silnie narażonych na działanie wiatrów burzowych, gdzie spełniają przede wszystkim rolę opaski przeciwwietrznej (RYC). W terenach o szczególnym zagrożeniu przez wiatr często nie wystarcza sam zamknięty brzeg z dobrze wykształconym okrajkiem. W celu zapobieżenia niebezpiecznym uderzeniom wiatru o ścianę lasu wychodzi mu się tutaj naprzeciw, sadząc przed ścianą drzewostanu pas roślinności krzewiastej, lokalizując wysokie krzewy w partii przy samej ścianie i coraz niższe krzewy w miarę oddalania się od ściany lasu. Powstaje w ten sposób utworzona z roślinności równia pochyła, po której wiatr, miast uderzać w wysoką i pionową ścianę drzewostanu, podnosi się płynnie ku górze (opływając tylko las i posuwając się nad jego sklepieniem) nie wyrządzając żadnych szkód.
Z powyższego wynika, że warstwa drzew ma wielkie znaczenie socjologiczne i ekologiczne dla lasu. Mając ponadto na względzie wielorakość i różnorodność spotykanych w naturze drzewostanów, które należy wyodrębnić i usystematyzować w toku prac urządzeniowych w celu prawidłowego ich zagospodarowania, można w konkluzji sprecyzować następującą definicję. Drzewostanami nazywamy części lasu różniące się od swego otoczenia sposobem powstania, wiekiem, składem gatunkowym, zwarciem, zagęszczeniem, jakością techniczną drzew, siedliskiem, obecnością nalotów lub podrostów itp., a które dzięki zajmowaniu większych powierzchni pozwalają na odrębne sposoby ich zagospodarowania.
Tak rozumiany drzewostan (w ujęciu taksacyjnym pododdział, wydzielany w urządzaniu lasu na tle siedliskowego typu lasu) stanowi również jednostkę hodowlaną, natomiast z punktu widzenia produkcji jest ogniwem w większej jednostce, którą przedstawia obręb produkcyjny lub gospodarstwo. Aby mówić o drzewostanach, a także o czynnościach hodowlanych wynikających z różnych sposobów ich zagospodarowania, trzeba poznać przede wszystkim charakterystyczne cechy drzewostanów, według których się je rozróżnia i klasyfikuje. Są to: l) pochodzenie, 2) wiek, 3) wysokość (grubość), 4) wielkość, 5) skład gatunkowy, 6) budowa, 7) zagęszczenie, 8) zwarcie, 9) jakość i 10) struktura.
II. Cechy drzewostanu
l. Pochodzenie drzewostanu
Drzewostan pochodzenia naturalnego i sztucznego. Pochodzenie drzewostanu jest bardzo istotną cechą, lecz nie zawsze łatwą do określenia w terenie. Drzewostany mogą powstawać przez odnowienie naturalne (samosiew), przeważnie jednak pochodzą z bardzo rozpowszechnionego w ostatnim stuleciu odnowienia sztucznego (przez siew lub sadzenie).
Sztucznie odnowione drzewostany dość łatwo rozpoznać po regularnej więźbie drzew i przez długie lata zaznaczających się rzędach, a ponadto po równomiernym wzroście na całej powierzchni (rys. 172),
Drzewostany pochodzące z samosiewu wykazują zawsze bardzo dużą różnorodność, w szczególności silne zróżnicowanie zagęszczenia, wieku oraz wysokości i grubości drzew (rys. 173).
Rozpoznanie pochodzenia drzewostanu jest łatwe prawie wyłącznie w jego okresie młodocianym. W starszych, wysokich i z natury bardzo już przerzedzonych drzewostanach dopiero wnikliwe badanie poparte analizą drzew próbnych lub metodą nawiertów drzew świdrem przyrostowym (umożliwiającą określenie wieku poszczególnych drzew) rozstrzyga często o rzeczywistym pochodzeniu drzewostanu. Jednakże nawet stwierdzona równowiekowość nie przesądza nieraz o sztucznym pochodzeniu drzewostanu,
Nawet silnie zróżnicowane w młodości drzewostany gospodarcze, powstałe przez samosiew, mogą pod wpływem cięć pielęgnacyjnych (trzebieży dolnych) upodobnić się w starszym wieku do drzewostanów pochodzenia sztucznego (wykazywać niewielkie zróżnicowanie wysokości i grubości).
Drzewostan pochodzenia wegetatywnego i generatywnego. Niezależnie od pochodzenia z różnych sposobów odnowienia, drzewostan może być pochodzenia wegetatywnego (z odrośli), tworząc tzw. las niskopienny, lub pochodzenia generatywnego (z nasion), w wyniku czego mamy do czynienia z lasem wysokopiennym. Obie wymienione wyżej formy lasu mogą występować na tej samej powierzchni, tworząc tzw. las połączony. W tym przypadku określenie pochodzenia drzewostanu nie nastręcza na ogół większych trudności.
Drzewostany pochodzenia wegetatywnego cechuje w młodości bardzo szybki wzrost. Odrośle nie muszą bowiem wysilać się w pierwszych latach życia na budowę korzeni, lecz korzystają w tym czasie z olbrzymiego - w stosunku do własnych wymiarów - systemu korzeniowego ściętego, starego drzewa matecznego. Dzięki bardzo obfitemu zaopatrzeniu w wodę i składniki pokarmowe odrośle bujnie przyrastają, wielokrotnie przewyższając wzrostem ziarnówki, np. odrośle osiki przyrastają nieraz 30 razy szybciej od równowiekowych osobników wyrosłych z nasienia.
Oprócz różnic we wzroście istnieją jeszcze pewne cechy morfologiczne, po których poznajemy odrośle np. kształt i wielkość liści. Zjawisko to szczególnie wyraźnie występuje u osiki, ale stwierdzamy je także u lipy, dębu i wiązu. W praktyce leśnej najczęściej mamy do czynienia z odroślami z pniaka, rzadziej z korzenia. Liczba pędów odroślowych na jednym pniaku jest nieraz znaczna; np. na pniaku ściętego dębu stwierdzano nieraz po 100 i więcej odrośli. Liczba ta z wiekiem ulega stałej redukcji i dochodzi zazwyczaj do kilku grubych pni, a przy prowadzonej pielęgnacji gospodarz lasu często pozostawia tylko jeden pęd z pniaka. Wówczas o pochodzeniu drzewostanu mówią nam przyrosty grubości (tzw. słoje roczne), bardzo szerokie w partii wewnętrznej przyrdzeniowej i szybko zwężające się w późniejszych latach, gdy drzewo przestało już korzystać z systemu korzeniowego pniaka matecznego i wykształciło własny, znacznie słabszy system pobierania. Najczęściej jednak w lesie odroślowym pozostawia się z każdego pniaka po 2 lub 3 pędy, wyrastające w postać „bliźniaka" lub „gniazda" (rys. 174).
Największą zdolność odroślową przejawiają drzewa w okresie energicznego wzrostu na wysokość, na żyznych siedliskach oraz w pełnym świetle (jałowe siedlisko i cień ją hamują, a mają tę zdolność tylko niektóre z naszych gatunków drzew leśnych, jak olsze, dęby, wiązy, osika i grab.
Drzewostany pochodzenia wegetatywnego żyją krócej, wykazują mniejszą zdrowotność i produkują małowartościowy surowiec drzewny (głównie opał), skutkiem czego hodowla takich drzewostanów ma uzasadnienie głównie w okolicach o znacznym zapotrzebowaniu na drewno opałowe. Przyczyną krótkiej żywotności drzewostanów odroślowych jest to, że połączenie odrośli ze starym pniakiem, po wykształceniu przez nie własnego systemu korzeniowego, staje się bramą dla inwazji patogenów, głównie grzybów pasożytniczych, atakujących rdzeniową partię szyi korzeniowej drzewa. Oprócz tego dołącza się tutaj jeszcze tzw. zmęczenie korzeni i stadialne starzenie się komórek, gdyż wiek osobników kolejnych pokoleń przy wegetatywnym mnożeniu sumuje się; prowadzi to często do degeneracji.
W praktyce leśnej najczęściej mnoży się wegetatywnie przez odrośle z pniaków olszę czarną, grab i dąb. Poza lasem, w wysokopiennych uprawach plantacyjnych rozmnaża się wegetatywnie przez sadzenie zrzezów (sztobrów) gatunki o największej sile odroślowej: topole i wierzby (rys. 175). Z gatunków iglastych występujących w naszych lasach zdolność pędzenia odrośli mają tylko cis, sosna smołowa i tuja (wszystkie zresztą bez większego znaczenia gospodarczego).
Drzewostany pochodzenia generatywnego są zdrowsze od odroślowych i charakteryzują się długowiecznością. W młodości przyrastają znacznie wolniej od odroślowych. Drzewa na przekroju wykazują równomierny układ wąskich słojów rocznych, a pod względem morfologii wykształcają wysokie, proste i gonne pnie (strzały) i wysoko osadzone wierzchołkowe korony, mają prawidłowy pokrój, osiągają znaczne rozmiary i produkują cenne drewno (rys. 176 i 177).
Warunkiem rozmnażania generatywnego jest osiągnięcie przez drzewa wieku fizjologicznej dojrzałości, w którym zaczynają kwitnąć i obradzać nasiona.
Wiek ten jest różny dla poszczególnych gatunków drzew, jednak u wszystkich zaobserwować można, iż najwcześniej zakwitają osobniki rosnące w odosobnieniu i korzystające z pełnego światła. Także wcześniej niż wewnątrz drzewostanu zakwitają drzewa okrajkowe, korzystające jednostronnie z większego dostępu światła. Najpóźniej zaczynają obradzać nasiona drzewa rosnące w głębi zwartego lasu. Stąd wniosek, że światło jest czynnikiem przyspieszającym wiek dojrzewania fizjologicznego drzew.
Lata obfitego obradzania nasion w drzewostanach zwą się w hodowli lasu latami nasiennymi. W zależności od urodzaju wyróżniamy lata nasienne: słabe, średnie i dobre. Między latami nasiennymi występują lata głuche, w których tylko mała liczba drzew skąpo obradza, natomiast okres dzielący lata nasienne nazywamy okresem nasiennym. Częstość i obfitość obradzania nasion zależy od wielu czynników, głównie jednak od właściwości genetycznych i odległości występowania danego gatunku od jego optimum klimatycznego.
W cieplejszym klimacie drzewa obradzają częściej. Jako przykład posłużyć tu może sosna zwyczajna, która w naszych szerokościach geograficznych obsiewa się obficie co 2 lub 3 lata. W środkowej Finlandii jej okres nasienny wydłuża się już znacznie do 10, a nawet 20 lat, a jeszcze dalej na północ na obfitszą produkcję nasion liczyć można zaledwie raz na 100 lat. Podobnie dzieje się ze świerkiem, który u nas obradza co 5 lub 6 lat, a w Norwegii już tylko co 9 lat.
2. Wiek drzewostanu
A. Drzewostany równo i różnowiekowe
równowiekowe - lasy gospodarcze - tam gdzie stosuje się zręby zupełne i odnowienia sztuczne z siewu i sadzenia
różnowiekowe - lasy naturalne i pierwotne oraz gospodarcze zagospodarowane rębnią przerębową lub zrębowo-przerębową.
ściśle równowiekowe drzewostany są praktycznie niemożliwe: różny jest czas kiełkowania nasion, wypadanie, dosadzanie, samosiew; brzoza, osika, sosna.
w hodowli lasu za równowiekowe uważa się: gdy w okresie młodszym różnice wieku poszczególnych drzew dochodzą do 10 lat, a w starszych do 20 lat.
klasy wieku Ia, Ib, IIa, IIb, itd.
Oznaczanie wieku drzewostanów
na podstawie ksiąg gospodarczych, w których jest historia drzewostanu (rok założenia i sposób powstania)
na ściętych drzewach licząc słoje + 2-5 lat na siewkę
drzewo stojące - nawiert świdrem przyrostowym w szyi korzeniowej i policzenie słojów rocznych na wałeczku drewna
młodsze drzewostany iglaste: policzenie okółków gałęzi + dodanie 2-5 lat na wyrośnięcie 1-go okółka.
B. Fazy rozwojowe drzewostanu
Podstawą wyróżnienia fazy są kryteria biologiczne i gospodarcze
1. Uprawa lub nalot (uprawa - sadzenie, nalot - samosiew)
trwa od chwili skiełkowania lub podsadzenia do chwili zetknięcia się koron drzew czyli wystąpienia zwarcia. Zależy np. od
siedliska - sosna - na żyznym siedlisku osiągnie zwarcie w 7 lat, na ubogim siedlisku do kilkunastu lat,
więźby - czyli gęstości sadzenia. Przy więźbie prostokątnej (nierównych odstępach sadzonek między rzędami i w rzędach) zwarcie w rzędach nastąpi wcześniej niż między rzędami.
tempo wzrostu - jako cecha gatunkowa, światłożądne zwykle rosną na początku szybciej niż cienioznośne. Tempo wzrostu zmienia się też z czasem np. świerk do 10 lat rośnie wolniej niż buk, lecz później dogania go i przegania pod tempem wzrostu.
ta faza decyduje o udatności drzewostanu, ważna jest znajomość ekologiczna drzew i dobra diagnoza siedlisk.
jest to okres indywidualnego wzrostu drzew, brak powiązań socjologicznych, drzewa nie wchodzą między sobą w stosunki konkurencyjne i nie oddziałują na klimat.
2. Młodnik lub podrost (do ok. 20-25 lat)
Młodnik pochodzi z odnowienia sztucznego na powierzchni otwartej a podrost z odnowienia naturalnego pod okapem drzewostanu.
to pierwsza faza gdzie uwidacznia się konkurencja i drzewa oddziałują na siebie, w wyniku zwarcia i ocienienia gleby we wnętrzu młodnika panuje odrębny fitoklimat,
trwa od zwarcia drzew koron do chwili obumierania dolnych gałęzi, czyli tzw. oczyszczania się, w tej fazie korony szybko rosną do góry,
cechą tej fazy jest szybki wzrost - ma tu miejsce kulminacja rocznych przyrostów,
wytwarzają się warstwy drzew - w zależności od składu gatunkowego drzewostan różnicuje się na 2-3 warstwy.
3. Tyczkowina (do ok. 30-35 r. i pierśnicy do ok. 10 cm)
wysokie tempo wzrostu na wysokość, „stagnujący przyrost na grubość”, niekorzystny stosunek grubości do wysokości.
pnie smukłe, sosna ma koronę z 6 okółków, dolne obumierają wskutek intensywnego oczyszczania się drzew
narażony na wiatry i śniegołomy
4. Drągowina (od ok. 30 lat do 50 lat) pierśnica 11-20 cm
koniec intensywnego wzrostu na wysokość i rozpoczęcie przyrostu na grubość
słabną procesy oczyszczania się i wydzielania drzew,
osiąganie wieku dojrzałości fizjologicznej.
5. Drzewostan dojrzewający (od ok. 50 lat do ok. 80 lat i pierśnicy 21-35 cm)
wysokość drzew jest już ustalona i przyrosty są minimalne
utracenie zdolności rozrostu koron i ich regeneracji
przerwanie pułapu koron to powstawanie luk w sklepieniu leśnym
drzewa regularnie kwitną i obradzają
wzmożony przyrost grubości
jeśli są cięcia pielęgnujące to występuje zjawisko przyrostu z prześwietlenia, czyli wzmożony przyrost średnicy drzew,
6. Drzewostan dojrzały (80-100 lat, pierśnica 36-50 cm)
osiągnięcie kulminacji przyrostu grubości
nadaje się do użytkowania rębnego
drzewostan wszedł w okres odnowienia naturalnego
7. Starodrzew (pow. 100 lat, pierśnica pow. 50 cm)
traci zdolność przyrostu, oznaki starzenia się
obumierają pojedyncze drzewa, zwarcie stopniowo się rozluźnia.
Są to oczywiście cechy uśrednione, istnieje zróżnicowanie międzygatunkowe: są gatunki długowieczne, np. dąb oraz krótkowieczne np. brzozy - są więc różnice w długości faz.
C. Pielęgnacja drzewostanu w fazach rozwojowych
1. Faza uprawy i nalotu
prace pielęgnacyjne: obróbka powierzchniowa gleby, zwalczanie mechaniczne i chemiczne chwastów, przerzedzanie za gęstych siewów, nawożenie organiczne lub mineralne
czyszczenia wczesne (cięcia pielęgnacyjne): regulacja składu gatunkowego, usuwa się gatunki niepożądane pionierskie np. brzozy, osikę, a także odrośla i przedrosty (starsze osobniki tego samego gatunku które zasiedliły teren wcześniej i górują rozmiarami nad sadzonkami) i przerosty (osobniki tego samego gatunku jednak charakteryzujących się nadmiernie bujnym wzrostem).
2. Młodniki i podrosty
czyszczenia późne, celem ich jest:
regulowanie składu gatunkowego
wyrównanie tempa wzrostu, usuwanie przerostów (mała wartość drewna) i rozpieraczy
normowanie zwarcia
poprawa jakości i zdrowotności poprzez usuwanie osobników niepożądanych
3. Tyczkowina i drągowina
do tej pory czyszczenie to była głównie „selekcja negatywna"
od tej klasy wieku prowadzi się trzebierze tj. cięcia pielęgnacyjne w nawrocie 3-5 letnim jako trzebierze wczesne
• określa się ją jako selekcję pozytywną (wybiera się do dalszej hodowli osobniki dorodne, najlepsze), zadania:
ochrona najwartościowszych drzew
poprawa jakości surowca drzewnego
poprawa stanu sanitarnego i biologicznego
zwiększenie przyrostu miąższości
zachowanie siły produkcyjnej gleby
dostarczenie cienkich użytków międzyrębnych
4. Drzewostan dojrzewający
trzebierze późne powtarzane co 5-8 lat, cięcia w tym okresie trwale przerywają dach koron nie mogą być stosowane więc tak często. Te cięcia pielęgnacyjne mają na celu:
skrócenie okresu produkcji przez osiąganie przyrostu z prześwietlenia, czyli grubości drzew
wzmożenie sił wytwórczych siedliska poprzez zintensyfikowanie procesów glebowych (zwiększenie ilości światła, ciepła i wilgoci docierającej do gleby).
przygotowanie drzewostanu do odnowienia: skrócenie cyklu produkcyjnego (naturalne odnowienie pod okapem drzewostanu macierzystego)
w drzewostanie o specjalnym znaczeniu - ochronnym spotęgowanie ochronnej roli lasu
zwykle ta faza kończy się wyrębem lasu
jeśli życie drzewostanu ma być przedłużone (np. produkcja bardzo cennych sortymentów , mają być to lasy ochronne czy rezerwatowe to pielęgnacja się nie kończy. Prowadzi się dalsze cięcia pielęgnujące zapas oraz cięcia sanitarne (wycinanie posuszu i drzew chorych, usuwanie śniego i wiatrołomów)
3) Wysokość drzewostanu
Jest to wypadkowa wysokości wszystkich drzew tworzących drzewostan.
I. Przyczyny różnej wysokości drzewostanów:
Wiek: jego konsekwencją jest wzrost i rozwój drzewostanu, ok. 50 - 60 lat ustala się wysokość drzewostanu
Bonitacja siedliska. Żyzność siedliska stwarza zróżnicowane warunki wzrostu na wysokość
• na jałowych piaskach drzewostan osiągnie 1/2 wysokości tej co na glebach próchnicznych, tak więc średnia wysokość drzew jest także miernikiem żyzności siedliska.
3. Gatunek drzewa
występują różnice indywidualne
różne jest tempo wzrostu: jodła powoli przyrasta w młodości, później nadrabia ten okres,
Zmienność osobnicza (różne są cechy genetyczne i właściwości biologiczne gatunku i osobników)
Cięcia pielęgnacyjne. W zależności od metody cieć średnia wysokość może być podwyższana lub obniżana:
silna trzebież górna - obniżenie śr. wysokości drzewostanu
silna trzebież dolna - podwyższenie śr. wysokości drzewostanu (usuwanie drzew przygłuszonych i niwelowanie pionowego zróżnicowania).
Związek wysokości z grubością drzewostanu (krzywa wysokości)
Stosunek wysokości drzewa do grubości jego pnia na wysokości piersi pozwala na określenie wysokości drzew w drzewostanie na podstawie pomiaru niewielkiego procentu drzew o różnych pierśnicach a następnie naniesienie pomierzonych wysokości jako odnośnych punktów w polu współrzędnych, odpowiadających na osi odciętych ich pierśnicom. Linia wyrównująca układ punktów w polu nosi miano krzywej wysokości drzewostanu.
Ryc.
Grubość drzew koreluje z wysokością lecz skala grubości jest bardziej zróżnicowana, gdyż zdolność przeżycia drzewa jest mniej zależna od grubości niż od wysokości.
Związek pomiędzy grubością drzewa a korona jest silniejszy. Na podstawie grubości drzewa można oszacować jego klasę biologiczna i wielkość korony.
4. Wielkość i kształt powierzchni drzewostanu
Wyróżniamy drzewostany duże i małe.
Wg Bauera duży drzewostan o powierzchni co najmniej 1 ha, zajętej przez skupienie drzew różniące się wyraźnie od swego otoczenia wiekiem, gatunkiem lub kombinacją gatunków, budową pionową bądź odrębnością siedliska (bonitacji). Na pojęcie drzewostanu składają się:
do 5 arów - mała grupa
5-10 arów - duża grupa
10-50 arów - kępa
0,5 - 1,0 ha - drzewostan mały
> 1 ha - drzewostan
Drzewostan duży jest ekologicznie samodzielny tzn. wpływy boczne (wnikanie światła bocznego, przenikanie wiatru czyli przenoszenie obcych nasion, wywiewanie ściółki) nie sięgają wnętrza drzewostanu. Zwykle wpływy te nie sięgają dalej niż wysokość drzewostanu.
zalety dużych drzewostanów
lepszy fitoklimat
uproszczenie prac gospodarczych i administracyjnych
wady
większe narażenie na pożary, wiatry burzowe, szkodniki.
5. Skład gatunkowy drzewostanów
Jest to najważniejsza cecha drzewostanu, od niej zależą inne a także uzależnione są czynności gospodarcze w lesie.
drzewostany jednogatunkowe - lite
wielogatunkowe - mieszane
a także
liściaste
iglaste
jest to podział mało precyzyjny gdyż i tu mogą występować i lite i mieszane. Drzewostan lity jest gdy inne gatunki stanowią do 10% liczby drzew, powierzchni lub masy
Na skład gatunkowy drzewostanów rzutują warunki środowiskowe - przy bardzo trudnych i skrajnych drzewostany mogą być lite.
A. Stopień zmieszania (w drzewostanach wielogatunkowych)
zmieszanie określamy liczbowo od 0 do 10 (pełny skład to 10).
równomierne np. 5 i 5
nierównomierne np. 7 i 3 czyli 1 gatunek przeważa
B. Formy zmieszania gatunków drzew
1. Rozmieszczenie przestrzenne
jednostkowe (pojedynczo lub po kilka w małych skupieniach)
rzędowe (rzędy gatunków rosną na przemian)
smugowe i pasowe (po kilka lub kilkanaście rzędów tego samego gatunku)
- grupowe gdy gatunki domieszkowe tworzą lite skupienia o wielkości jak w podziale Bauera.
- kępowe duże skupienia do 50 arów
2. Czas trwania domieszki
trwałe - zazwyczaj produkujące cenny użytek
przejściowe stosowane w pewnym celu np. dla ochrony lub pielęgnacji cennych gatunków głównych
przedplon: wstępna agregacja roślin pod której okapem czyli w korzystnych warunkach klimatycznych wprowadza się właściwy dla siedliska drzewostan cieniolubny i wrażliwy na przymrozki (jodła, buk). Przedplonem są gat. pionierskie światłożądne - brzoza, sosna, osika - zagłuszają chwasty i stwarzają korzystny mikroklimat.
podgon: jest to domieszka gatunków szybko rosnących w młodości w celu pobudzenia gatunków głównych do energiczniejszego wzrostu na wysokość. Są to zwykle gat. szybko rosnące (sosna, brzoza, robinia) pobudzają do wzrostu takie drzewa jak dąb, buk czy jodła (do 10 lat),
domieszki są cenne z ekologicznego punktu widzenia zwiększają heterogenność drzewostanu.
Dddddddddddddd uzupełnić
6. Budowa pionowa lasu
Budowa pionowa lasu to wykształcanie się w drzewostanie pięter drzewiastych - przyczyną jest wiek i gatunek drzew.
Zróżnicowanie wieku zależy od stosowanej rębni i odnowienia:
rębnia zupełna i sztuczne odnowienie - jednolity wiek
rębnia stopniowa lub przerębowa - zróżnicowany wiek
Gatunek: w drzewostanach mieszanych - różne gatunki mają różne właściwości ekologiczne tempo wzrostu i rozwoju - różne piętra. Gatunki światłożądne dystansują gatunki cienioznośne i budują górne piętro.
Budowa pionowa decyduje o stosowanych czynnościach gospodarczych (sposobie pielęgnacji)
W lasach zagospodarowanych wyróżniamy: drzewostany jedno-, dwu-, i wielopiętrowe.
a. Jednopiętrowe. Klasyfikacja drzew w drzewostanie.
Korony drzew obok siebie, prawie w jednej słabo zróżnicowanej warstwie, przestrzeń między koronami jest prawie nie wykorzystana, pomiędzy koronami występuje silny ucisk boczny,
W drzewostanie równowiekowym z czasem następuje rozwarstwienie pułapu koron, w efekcie powstaje zmienność ekologiczna poszczególnych osobników.
Występuje proces wydzielania się drzew dlatego utworzono klasy biologiczne.
Seebach - 1843: wyróżnił drzewa dominujące, opanowane, wydzielone, zagłuszone - podział ten miał ułatwić prace pielęgnacyjne - trzebieże.
Międzynarodowy Związek Leśnych Instytutów Badawczych - 1956 r. Klasyfikacja:
stanowisko socjalne: panujące (100), opanowane (200), przygłuszone (300).
wartość hodowlana: dorodne (10), pożyteczne (20), szkodliwe (30),
morfologia strzały i korony: dobra (1), średnia (2), zła (3),
Liczby odpowiadające poszczególnym cechom się sumuje i np. 231 oznacza drzewo współpanujące o nieprawidłowej strzale i dobrze rozwiniętej koronie.
b. Drzewostany dwupiętrowe
Korony drzew tworzą dwa odrębne piętra:
piętro górne - gat. światłożądne - tworzą okap lasu
piętro niższe - gat. cieniste - piętro podokapowe
dwupiętrowe są najczęściej skutkiem działalności gospodarczej człowieka.
okres międzywojenny - b. dużo klęsk: zaczęto w drzewostanach sosnowych robić podsiewy z liściastych (dąb, buk, grab) - najwięcej na terenach Wielkopolski.
c. Drzewostany wielopiętrowe
3 i więcej pięter drzewiastych b. rzadkie - regle dolne.
powstają w efekcie rębni jednostkowo-przerębowej
są to zazwyczaj drzewostany różnowiekowe, zbudowane głównie z gatunków cienistych.
d. Budowa lasów pierwotnych
W większości są różnowiekowe, różnogatunkowe i wielopiętrowe.
W warunkach skrajnych występują jako jednogatunkowe, jednopiętrowe - np. tajga, lite sośniny na wydmach, las krańcowy w Europie - sosny, lite buczyny w Rumunii i Bośni.
Las pierwotny wykazuje przestrzenne zróżnicowanie w etapach rozwoju, występują mozaikowato 3 człony wiekowe: odnowienie naturalne, optymalny wzrost i zamieranie starych drzew. Te człony występują grupowo lub kępowo.
7. Zagęszczenie drzewostanu.
a. Pojęcie zagęszczenia w ekologii
Liczba osobników drzew na jednostkę powierzchni (1ha)
przy odnawianiu naturalnym - miliardy nasion/ha - z czego mały % w glebie kiełkuje, ale i tak daje setki tys. siewek/ha.
przy odnowieniach sztucznych sadzi się kilkanaście tys. sadzonek/ha, z tego w drzewostanie dojrzałym 100 letnim pozostaje kilkaset drzew/ha.
jest to skutkiem konkurencji o światło, wodę, pokarm - pozostają najlepsze, najbardziej przystosowane.
zagęszczenie gat. światłożądnych jest niższe niż cienioznośnych.
b. Problem więźby przy sztucznym odnowieniu
Z zagęszczeniem związany jest problem więźby (rozmieszczenia sadzonek)
Przy sztucznym odnowieniu mamy do czynienia z mniejszą liczba drzewek, od więźby początkowej zależy czas od posadzenia do osiągnięcia zwarcia - powinien on być najkrótszy.
Więźba początkowa zależy od:
sposobu przygotowania gleby pod uprawę (pełne umożliwia dowolną więźbę)
jakości siedliska, (żyźniejsze - gęstsza, gdyż inaczej więcej gałęzi - więcej sęków) i tak:
Bs - słaba żyzność siedliska - 10 tys. sadzonek w więźbie 1,4 x 0,7
na żyznych siedliskach sosnowych - 17 tys. sadzonek - 1,4 x 0,4m
wieku (wymiarów) sadzonek,
biologii gatunku (gęstej wymagają np. sosna dąb, buk; rzadkiej - modrzew, świerk)
początkowego tempa wzrostu - pionierskie rosną szybciej, więc sadzone powinny być rzadziej.
c. Zagęszczenie a konkurencja i kooperacja u drzew.
W drzewostanach gospodarczych rozmieszczenie najpierw jest regularne jednak później przekształca się w skupinowe.
Najostrzejszą konkurencję obserwuje się przy równomiernym rozmieszczeniu drzew w litych drzewostanach sztucznego pochodzenia. Różnicowanie wysokości prowadzi do zagłuszania niższych i słabszych osobników.
Jeszcze wyraźniej konkurencja występuje wśród dwóch gatunków, które dążą do rozdzielenia terytorialnego. W naturze konkurencja prowadzi do sukcesji ekologicznej. Nieregularne rozmieszczenie drzew i powstawanie skupinowej struktury przestrzennej jest interakcją dodatnią - sprzyja współpracy międzyosobniczej - drzewa opierają się skuteczniej niekorzystnym układom bio- i abiotycznym środowiska.
d. Grupowe życie drzew
Na skutek wydzielania się drzew słabszych pozostałe osobniki tworzą biologiczne skupienia tzw. biogrupy
Biogrupy maja jakby wspólną koronę - odcinającą od zewnętrznych wpływów atmosferycznych, zacieniają glebę - brak chwastów, zrastają się korzeniami - tworzą jakby jednolity organizm.
Optymalnym zjawiskiem jest gdy drzewostan składa się z 300-400 biogrup/ha a każda liczy 5-6 drzew.
Biogrupowość występuje powszechnie w lasach pierwotnych np. łączenie się świerków w górnej granicy lasy w tzw. roty czyli gęste skupiny złożone z kilku- kilkunastu drzew o jakby jednej wspólnej koronie.
Zwarcie drzewostanu
a. Pojęcie i sposoby określania zwarcia drzewostanu
Wpływa na ilość i jakość produkcji drewna. Drzewa powinny rosnąć względnie blisko siebie, stykając się brzegami niezbyt dużych, ale gęstych i symetrycznych koron.
zwarcie definiowane jest jako:
wzajemne położenie koron drzew
stopień wypełnienia przestrzeni nad glebą koronami drzew
stopień ocienienia gleby przez rosnący na niej drzewostan
stosunek łącznej powierzchni rzutów koron drzew rosnących na jednostce powierzchni do tej powierzchni.
Zwarcie zmienia stosunki świetlne w drzewostanie - ma wpływ na pokrój drzewa
Drzewa rosnące bez zwarcia - niekorzystny stosunek grubizny do drobnicy - miąższość drobnicy przekracza miąższość grubizny.
Drzewa w zwarciu - miąższość grubizny stanowi do 90%.
Określanie zwarcia
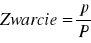
rzutowanie koron drzew na glebę
p - łączna powierzchnia rzutów koron, P - powierzchnia 1 ha.
zdjęcia lotnicze, planimetrowanie rzutów koron
szacunki zwarcia na oko przy długoletnim doświadczeniu
Skala szacunkowa zwarcia
silne (1,2-1,1) - korony zachodzą na siebie
pełne (1,0-0,9) - korony brzegami się stykają
umiarkowane albo rozluźnione (0,8-0,7) - między koronami przerwy mniejsze niż 0,5 m,
przerywane 90,6-0,5) - przerwy są takie że mogłyby się pomieścić korony drzew,
luźne (0,4) odległości między koronami takie że mogłoby się zmieścić więcej niż jedno drzewo
bez zwarcia (0,3 i poniżej) - drzewa stoją pojedynczo bez wpływu na siebie.
b. Przyczyny różnego zwarcia
jakość siedliska - na bogatszych korony są większe,
gatunek - gęsto ulistnione gatunki cieniste lepiej wypełniają przestrzeń niż światłożądne
wiek - silnie zwarte są zwykle młode drzewostany (młodnik i tyczkowina). Z wiekiem przerzedzają się.
zabiegi gospodarcze - z czasem prowadzą do rozluźnienia drzewostanu.
c. Wpływ zwarcia na drzewostan i glebę.
Wpływ zwarcia na drzewostan
Ze wzrostem zwarcia:
długość i wielkość koron się zmniejsza
grubość drzew maleje
strzały stają się pełniejsze i lepiej oczyszczone z gałęzi,
wąskie słoje roczne
mniejsza produkcja
drzewa osiągają większą miąższość pnia
opóźnia dojrzewanie i owocowanie.
Wpływ zwarcia na glebę
jest żyzna gleba o jednakowej miąższości
zapobiega erozji na stokach
drzewostan wykazuje właściwości retencyjne w stosunku do opadów atmosferycznych
nadaje glebie strukturę gruzełkowatą
hamuje parowanie wody
wpływa na runo
d. Zwarcie koron drzew a ażurowość sklepienia leśnego oraz stosunek zwarcia do zagęszczenia drzewostanu.
Ażurowość - luki miedzy koronami oraz w ich obrębie.
Jest to inaczej przepuszczalność świetlna pułapu koron drzew
Między zwarciem a zagęszczeniem drzewostanu istnieje związek polegający na tym, że bardziej zagęszczone drzewostany wykazują zwykle silniejsze zwarcie koron drzew.
Jakość drzewostanu
a. Pojęcie i przyczyny różnej jakości drzewostanu.
Dla hodowcy cecha jakości wiąże się z drewnem bezsęcznym, regularnie wąskosłoistym o prostej strzale. Drzewa w lesie gospodarczym powinny wykazywać pokrój monopodialny, mieć koronę wysoko osadzoną, długi odcinek oczyszczonego pnia a także cechować się równomiernym przyrostem na wysokość i grubość.
Przyczyny dobrej lub złej jakości.
Właściwości genetyczne osobników tworzących drzewostan - pewne cechy morfologiczne są dziedziczne.
Pochodzenie (proweniencja) drzew - pewne rasy klimatyczne jednego gatunku pochodzące z odmiennych warunków przyrodniczych wykazują negatywne skłonności, inne rasy mają dobre cechy i prawidłowy wzrost
Zgodność wymagań ekologicznych ekotypu z siedliskiem - wprowadzanie gatunku o małych wymaganiach na żyzne siedliska skutkuje pniami zbieżystymi i z dużą liczbą sęków. Wprowadzenie gatunku o dużych wymaganiach na siedliska ubogie powoduje karłowaty wzrost, krzaczasty pokrój i surowiec złej jakości.
Wzrost w odpowiednim zagęszczeniu - zastosowanie odpowiedniej więźby początkowej upraw zgodnej z wymaganiami biologicznymi gatunku prowadzi do przyśpieszenia oczyszczania się drzew i poprawy jakości technicznej drzewostanu.
Pielęgnacja - systematyczna pielęgnacja, normująca tempo wzrostu drzew i zagęszczenie drzewostanu prowadzi do poprawy jakości i wartości drzewostanu
10. Struktura drzewostanu
a. Prawo Queteleta oraz pojęcie struktury drzewostanu
Lambert Quetelet - Belg, statystyk stwierdził, że najwięcej spotyka się osobników reprezentujących średnie wymiary, a w kierunku wyższych i niższych wartości cechy liczba osobników maleje, osobników o skrajnych cechach jest bardzo mało. Po wykreśleniu otrzymujemy krzywą prawdopodobieństwa Gaussa czyli rozkład normalny.
b. Struktura litego drzewostanu równowiekowego
Struktura grubości drzew
Krzywa frekwencji drzew w klasach grubości jest krzywą binominalną, jednoszczytową. Różni się od krzywej Gaussa asymetrią, gdyż lewe ramię jest krótsze. Rozkład drzew jest dodatnio skośny
Struktura grubości drzewostanów równowiekowych wyróżnia się tym, że jest mniej drzew grubszych od grubości średniej a więcej cieńszych. Można wyznaczyć średnią arytmetyczną odliczając w uporządkowanym szeregu 40% drzew począwszy od najgrubszych lub 60% drzew od najcieńszych.
Z wiekiem drzewostanu - spłaszczenie wierzchołka i przesunięcie na wykresie w prawo
Struktura wysokości drzew
rozkład asymetryczny do struktury grubości
struktura grubości - charakter awansowy
struktura wysokości - charakter spadkowy - wydzielanie drzew
b. Duża asymetria w młodnikach w starszych mniejsze
Struktura biologiczna drzewostanu
klasy biologiczne drzewostanu wg Krafta uwzględniają wysokość i pierśnicę drzew
z wiekiem drzewostanu zanikają najniższe klasy - wydzielają się.
Struktura drzewostanu różnowiekowego
odnosi się do lasów naturalnych i sztucznych przez odnowienia naturalne i po odnowieniu zrębowo-przerebowym i przerębowych
struktura wg Paczoskiego - las naturalny
wg Kowalskiego - Puszcza Białowieska
krzywa z 3 wierzchołkami - zróżnicowana budowa piętrowa
w drzewostanach gospodarczych krzywa regularna bez wierzchołków
opracowano modele - maleje postęp geometryczny i logarytmiczny
Struktura drzewostanu wielogatunkowego
ujmuje się każdy gatunek oddzielnie
buk najwieksza dynamika wzrostu - dędzie dominować
jodła obecnie panująca nie odnawia się i jest zagrożona
świerk - gatunek ustępujący
w sukcesji - jodła może się odnawiać - cienilubna - prod biol - świerk zostanie wyparty
Zabiegi pielęgnacyjne
III. Dynamika rozwoju
Składa się z 4 faz
- proces naturalnego przemieszczania się drzew w warstwach drzewostanu - pionowo
- proces naturalnego wydzielania się drzew - poziome różnicowanie populacji
- proces naturalnego oczyszczania się pni z gałęzi - zmiany pokroju
proces naturalnego starzenia się
Proces przemieszczania się drzew w warstwach drzewostanu
wzrost drzew z wiekiem - to przemieszczenie się
wzrost nie jest równomierny, wiek 6 lat - 30-40 cm, 12 lat 2-3 krotnie wyższe przyrosty
wiek 6 lat drewnienie opóźnione, 12 lat różnice pogłębiają się
więźba |
Wiek 7 lat |
Drzewa porastają |
awansują |
spadają |
posusz |
Wiek 12 lat |
1 x 0,3 |
100% |
45,2% |
7,5 |
27,3 |
20,0 |
80 % |
1,2 x 0,5 |
100% |
34,5% |
13,2 |
36,2 |
16,0 |
84% |
z wiekiem przemieszczanie się maleje - zwiększa się wydzielanie
najbardziej intensywne w uprawie i młodniku
Proces naturalnego wydzielania się drzew
jest to efekt przemieszczania się pionowego z klas wyższych do niższych
proces ten rozpoczyna się po zwarciu
faza uprawy - indywidualny rozrost, brak konkurencji i obumierania o wypadają
faza młodnika - silna konkurencja i ciągła redukcja osobników - wydzielania się
faza uprawy - stosunek grubości do wysokości 1:50 1 cm średnicy - 50 cm wysokości
faza młodnika - stosunek grubości do wysokości 1:70 - efekt pogoni za światłem
ryc.
Liczba wypadów w uprawie odwrotnie proporcja do wielkości
Największe wydzielanie w młodniku - kulminacja wzrostu
Mniejsze wydzielanie - tyczkowina - 20-30 lat
Wydzielanie w drągowinie małe
Liczba drzew na 1 ha sosny
1 r - 12-30 tys
10 - 9,2-18 tys
30 - 2,3-4,1 tys.
60 - 730-177 szt.
80 - 480 - 880 szt.
tyczkowina - stosunek grubości do wysokości 1:90 : 1: 120
w tych fazach młodnik, tyczkowina i drągowina istotne prace pielęgnacyjne.
Proces oczyszczania się pni drzew z gałęzi
bardzo ważne dla jakości surowca
zaczyna się od fazy młodnika i trwa do drągowiny
w fazie młodnika i tyczkowiny ciągle 6-7 żywych okółków - dolne zamierają
oczyszczanie się zależy od:
światłożądność - im bardziej światłożądny tym krótsza żywotność gałęzi
więźba początkowa gęsta - szybciej
właściwości genetyczne gatunku, odmiany rasy klimatyczne
zasobności siedliska - żyźniejsze - gorzej się oczyszczają - może być nawet krzywy wzrost drzewa (sosna na glinie)
prace pielęgnacyjne - czyszczena trzebieże
oczyszczanie się gałęzi wynika z bilansu procesu fotosyntezy (A) i zapotrzebowania (B)
A>B gałęzie pozostają
A=B wzrost maleje
A<B obumierają
Efekt zacienienia i dostępności światła
Starzenie się drzewostanu
najbardziej uwidacznia się w lasach naturalnych
system korzeniowy najczęściej zamiera
mniejsza zdrowotność i podatność na choroby
obumieranie rozpoczyna się od korzeni.
Leśnictwo Wykład 1 - Las jako system ekologiczny 6
Zagadnienia autoekologii lasu
Leśnictwo Wykład 2 - Las jako system ekologiczny 25
Zagadnienia synekologii lasu
Leśnictwo Wykład 3 - Las jako zjawisko geograficzne 34
Leśnictwo Wykład 3 - Las jako zjawisko geograficzne 35
Leśnictwo Wykład 3 - Ekologiczne typy lasu wg rozmieszczenia poziomego 56
Leśnictwo Wykład 5 - ogiczne typy lasu wg rozmieszczenia pionowego 60
Leśnictwo Wykład 5 - ogiczne typy lasu wg rozmieszczenia pionowego 35
Leśnictwo Wykład 5 - Autoekologia 61
![]()
Wyszukiwarka
Podobne podstrony:
ekonomiczne uwarunkowania uzytkowania lasu, Ekonomika lenictwa - Tuchola i Poznan2008 (97) pelna2, E
Lasy Polski, leśnictwo wykłady
Podstawy leśnictwa wykład 1
Podstawy leśnictwa wykład 2
wykład VII, LEŚNICTWO SGGW, MATERIAŁY LEŚNICTWO SGGW, Transport, TRANSPORT
INŻYNIERIA LEŚNA Wykład I 29, AR Poznań - Leśnictwo, inżynieria leśna, Inżynieria
Wyklady z ekonomiki, Leśnictwo, Ekonomika w leśnictwie
OPISY do Projektu!!!!, LEŚNICTWO SGGW, MATERIAŁY LEŚNICTWO SGGW, Urządzanie, Wykłady, PROJEKTY sem 8
VI- Małopolska, LEŚNICTWO SGGW, MATERIAŁY LEŚNICTWO SGGW, Urządzanie, Wykłady, PROJEKTY sem 8 Rok Ak
TRANSPORT Wykład XI7, AR Poznań - Leśnictwo, Transport leśny, Transport - egz, Transport leśny
GTD, LEŚNICTWO SGGW, MATERIAŁY LEŚNICTWO SGGW, Urządzanie, Wykłady, PROJEKTY sem 8 Rok Akademicki 20
Opisy do wieków dojrzałości, LEŚNICTWO SGGW, MATERIAŁY LEŚNICTWO SGGW, Urządzanie, Wykłady, PROJEKTY
wykład III, LEŚNICTWO SGGW, MATERIAŁY LEŚNICTWO SGGW, Transport, TRANSPORT
Pytania z wykładu 10 - grupa 5, geodezja, rok III, Podstawy Rolnictwa i Leśnictwa
wykład nr 5, geodezja, rok III, Podstawy Rolnictwa i Leśnictwa
Wykłady z 29.05.09 z gleboznastwa, Leśnictwo, Gleboznawstwo leśne
wyklad7, STUDIA SGGW Wydział Leśny leśnictwo, Semestr 1 i 2, Zoologia, Zoologia egzamin niezbędnik
więcej podobnych podstron