9
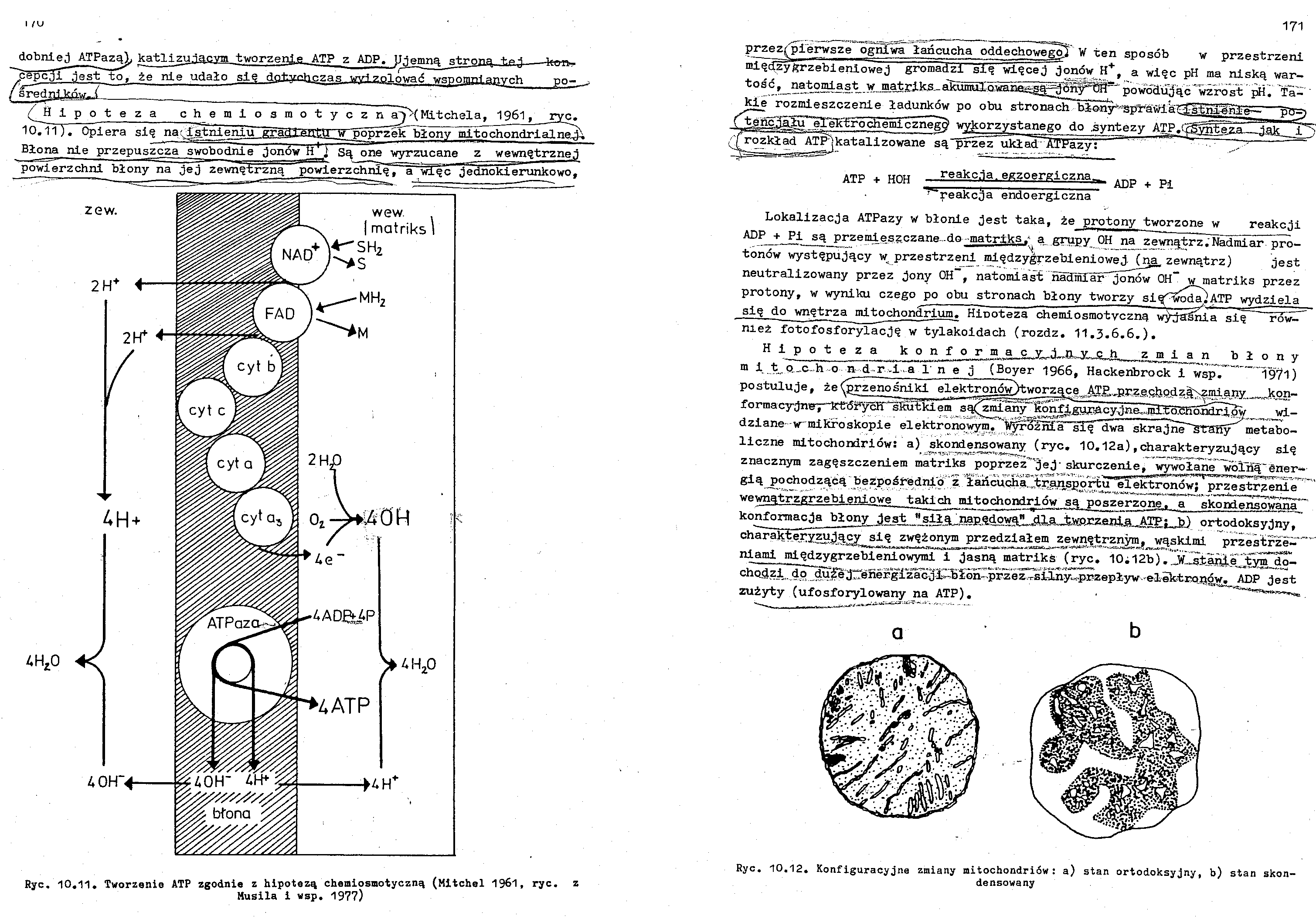
dobnlej ATPazą^ katlizu.iacvm tworzenie, ATP z ADP. Ujemna strona,,-taj--kon*
‘pepcjT^jesteto, że nie udało się dotychczas mzizalować wspQfflnl3£ych po-' sred ' /
Hipoteza chemloa m o t y c z njgK( Mitchela, 1961, _ryc.
loTu)-. Opiera się narfatnienlu gra
__ _ "poprzeŁ^błony mltochondrlalnej^
Błona nie przepuszcza swobodnie jonów H ] Są one wyrzucane z wewnętrznej powierzchni błonyna jej zewnętrzną powierz chnl^T^ więc "jednokierunkowo,
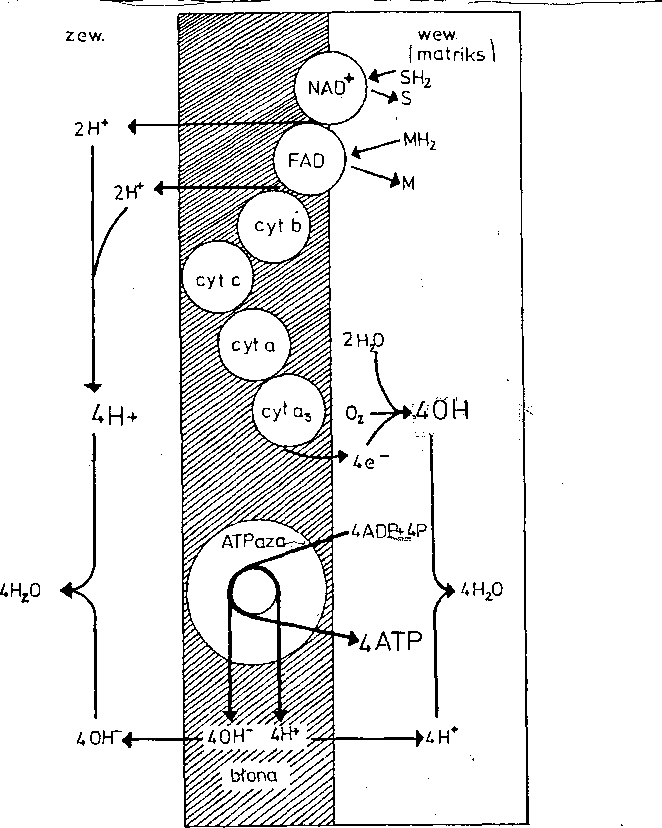
Ryc 10.11. Tworzenie ATP zgodnie z hipotezą chemiosmotyczną (Mitchel 1961, ryc. z
Husila i wsp. 1977)
pr2e2(^pierws2e ognlwaT łańcucha oddechowegal W ten sposób w przestrzeni inięcizygrzebieniowej gromadzi się więcej-jonów H*, a więc pH ma niską wartość ^naJio^as^.w..mąt^kjs._aKimiSbSfflne^^g^i^wŁ' po\rod^ąc'^3TOst pH. Ta- ' kie rozmieszczenie ładunków po obu stronach bł^ny^^pTdł/iallU^— tjh^i^r^^fFo^h^mlczn^g^ wykorzystanego do .syntezy ATP.ci^^tejZZIi^c
rozkład^Fjkatalizowane są'^rzez^Ikład~ATPazy:
ADP + Pi
ATP + HOH reakcja.egzoergiczna.
T reakcja endoergiczna
reakcji
Lokalizacja ATPazy w błonie jest taka, że protony tworzone w
ADP + Pi są przemieszczane-do -matriks .* a grupy OH na zewnątrz. Nadmiar protonów występujący ws przestrzeni roiędzy^rzebieniowej- (na zewnątrz) jest neutralizowany przez jony OH", natomiast nadMar^jonów OH" w matriks przez protony, w wyniku czego po obu stronach błony tworzy się^^rodajATP wydziela się do wnętrza mitochon^rium^ Hipoteza chemiosmotyczną wihtaŚnia sle~^ rów-~~ nież fotofosforylację w tylakoidach (rozdz. 11.3-6.6.).
Hipoteza konform a c y j„.n.v c h z m i_a n błony m i .t, o.-c-h o a-.d-r.-i-.-a l'n e j (Boyer 1966, Hackeribrock i wsp. " ~19?1)
postuluje, że (przenośniki elektronów)tworzące _Affi^prze,QhPdz^xzmiany_____kon-
formacy,jne7^k1fófiych^ŚlnJtki em są<^Sany“£onfi^picyjne,,Jmitb.chondri^ _ widzi anew^ mikro skopi e el ektronowym. ^ftfr ozS a^Tę^dwa skra jne^Tany^ metaboliczne mitochondriów: a) skondensowany (ryc, 10,12a),charakteryzujący się znacznym zagęszczeniem matriks poprzez^jej- skurczenie, wywołane l/6Xną;~dner-gią jjochodząćą bezpośrednio 'z, łanću^ą tę elektronów; przestrzenie
wewnątrzgrzebieniowe takich mitochondriów są poszerzone, a skoMensowana"~ konformacja błony jest ttslłą;,napgdpwĄ!L^Xa^PJgZghi^^^r.b) ortodoksyjny, charakteryzujący się zwężonym przedziałem zewiętrzr^^ wąskimi przestrze-'" niami międzygrzebieniowymi i jasną matriks (ryc, 10;12b).
chodzJ^^do^di^j^fiei^za^i^loh-pp^ez^iiriy^pa^epływ^el-^tEoj^Ws ADP jest zużyty (ufosforylowany na ATP),

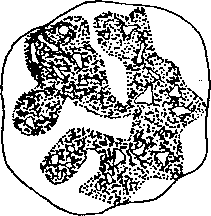
Ryc. 10.12, Konfiguracyjne zmiany mitochondriów: a) stan ortodoksyjny, b) stan skondensowany
Wyszukiwarka
Podobne podstrony:
4t©»- dobnle j ATPazą^ katlizu.iacym tworzenia &TP % ADP» Pjemną stroną tftj—-^C^cnl^.lsst Że ni
4t©»- dobnle j ATPazą^ katlizu.iacym tworzenia &TP % ADP» Pjemną stroną tftj—-^C^cnl^.lsst Że ni
DSC09391 Ryc- 12*8. Rola cyklu ATP/ADP w przenoszeniu fosforanu bogatoenergetycznego. Należy zwrócić
60311 PICT5906 tekstu określonych wyrazów, znaków, cyfr. liter itp. Ujemną stroną tych zadań jest to
P1080957 szybkość tworzenia ATP w różnych przemianach Maksymalna szybkość tworzenia
78075 P1080838 ATP-aza miozynowa ATP c=== S=»- ADP + PI + Q (energfaj PROGI RESYfrr V ATP W MIĘŚNIAC
0000077 2 //O <p—o~CE> H-C-OH H2C-0-® +E-SH HC-OH —* HjC-O-® ATP ADp //O C-S-E H-Ć-OH
r CO2 Ryc. 9.2. Schemat uzyskiwania i zużytkowania energii przez komórkę przy udziale cyklu ATP-ADP
DSC00693 glukozo-ó-/1 glukozo-1-Z’+ ATP ADP-glukoza f+ akceptor *-1
więcej podobnych podstron