skanowanie0043
■
tatro komórkowe
|y Drugą strategią prowadzącą do amplifikacji genów rybosomowego RNA jest ■prielojkrotnienie rDNA w jądrze samej komórki jajowej. Tę strategię obserwuje się probu typach jajników, panoistycznym i meroistycznym. Amplifikacja rDNA w jąd-»ch tiocytów ma charakter pozachromosomowy. Kopie genów rybosomowych za-iparte są w licznych/ kolistych cząsteczkach rDNA, które pozostają poza strukturą ^feihosomów jąderkotwórczych (ryc. 4.2.17). Amplifikacja rDNA zachodzi podczas ^Hhezy w profazie podziału mejotycznego w stadium pachytenu (patrz rozdz. 8.2). Balistę cząsteczki zostają uwolnione z rejonu organizatora jąderka (NOR) w sposób MD końca niewyjaśniony. Replikacja uwolnionej kolistej cząsteczki rDNA zachodzi przez stałe dodawanie zasad DNA (fragmenty Okazaki) do otwartego końca 3' jej ni-^Wfechanizm replikacji kolistych cząsteczek rDNA przebiega zgodnie z zasadą „to-Bącego się koła" i jest powszechnie obserwowany zarówno wśród bezkręgowców, ^Riręgowców. Stopień zwielokrotnienia rDNA jest bardzo różny nawet u blisko ^Bcrewnionych gatunków. W oocytach pazurnicy (Kenopus laeoiś) poziom amplifi-BBranego rDNA wynosi 30-35 pg, co stanowi 1400-krotny przyrost rDNA w porowa mama z genomem haploidalnym tego gatunku. W przypadku pstrąga tęczowego Wtitmo gairdneri) przyrost rDNA w oocycie wynosi 20 pg, a w oocytach niektórych ^fe jesiotrowatych (Acipenseridae) jest 50-krotny. W jajach chrząszcza żółtobrzeżka tPytiscus marginaliś) ilość rDNA wytworzonego na drodze amplifikacji wynosi 67 pg, Komiast w oocytach niektórych mięczaków przyrost rDNA jest niewielki - 2-5-Nbrotny (małże, Bivalvia). Podobnie niski poziom zwielokrotnienia rDNA obserwuje pę wlbocytach ssaków. Stopień amplifikacji rDNA w oocytach pawiana i człowieka K tylko 4r-5-krotny.
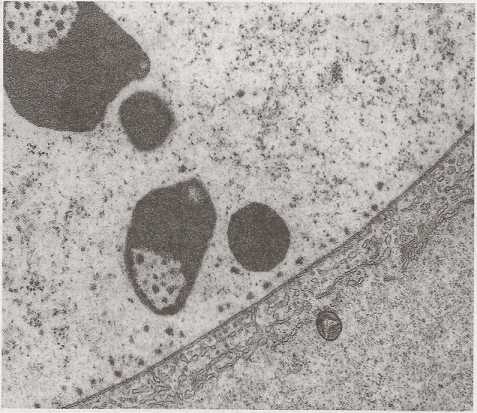
■jrc. 4.2.18. Fragment jądra oocytu z okresu prewitelogenezy wija Hanseniella nivea (Myriapoda) z czterema jąderkami wielokrotnymi. (Dzięki uprzejmości prof. Szczepana Bilińskiego)
Wyszukiwarka
Podobne podstrony:
skanowanie0045 172 Alkohol Ol A 0,6%o albo prowadzi do stężenia przekraczającego te wartości lub (2)
skanowanie0045 172 Alkohol Ol A 0,6%o albo prowadzi do stężenia przekraczającego te wartości lub (2)
65708 skanowanie0045 172 Alkohol Ol A 0,6%o albo prowadzi do stężenia przekraczającego te wartości l
skanowanie0006 15 □ Druga strategia zakłada, że systematyka jest nauką praktyczną
Jacek Rybicki, Beata Pawłowska M. Portera, że istnieją dwie fundamentalne strategie prowadzące do su
P4250127 214 Oczywiście druga definicja prowadzi do wyższej wartości ą, > 7,0, skoro Ht < Hm.
65708 skanowanie0045 172 Alkohol Ol A 0,6%o albo prowadzi do stężenia przekraczającego te wartości l
Logistyka jako strategia... 17 Do redukcji czasów realizacji możliwe jest zastosowanie procedury
iKAPITAŁ LUDZKI NARODOWA STRATEGIA SPÓJNOŚCI m Do wykonywania zadań zawodowych niezbędne jest
Wstyd i przemo0125 248 Wstyd i przemoc prowadzą do śmierci ofiary. Masochizm nie jest rozwiązaniem p
1624303p1158479924771?9213578 n W prowadzenie do prohlimalN ki ekonomiczne] Kkonomia jest nauką bada
więcej podobnych podstron