
U
RSZULA
Z
IELENKIEWICZ
i
P
IOTR
C
EG£OWSKI
Zak³ad Biochemii Drobnoustrojów
Instytut Biochemii i Biofizyki PAN
Pawiñskiego 5a, 02-106 Warszawa
e-mail: ulazet@ibb.waw.pl
e-mail: piotr106@ibb.waw.pl
MECHANIZMY STABILNEGO DZIEDZICZENIA PLAZMIDÓW
LOSOWY I LEPSZY NI¯ LOSOWY ROZDZIA£ PLAZMIDÓW DO KOMÓREK POTOMNYCH
Podczas podzia³u komórek bakterii ist-
niej¹ce w nich plazmidy ulegaj¹ segregacji do
komórek potomnych. Naturalnie wystêpuj¹ce
plazmidy bakteryjne s¹ niezwykle trwale dzie-
dziczone przez komórki potomne i to w wa-
runkach braku presji selekcyjnej. Plazmidy
ma³e, o wielkoœci nie przekraczaj¹cej kilku ty-
siêcy par zasad, zawdziêczaj¹ to po prostu
znacznej liczbie kopii w jakiej wystêpuj¹ w ko-
mórce. Stwierdzenie to stanie siê jasne przy
za³o¿eniu, ¿e plazmidy mog¹ „rozchodziæ siê”
do komórek potomnych w sposób losowy.
Wtedy prawdopodobieñstwo, ¿e dany plazmid
nie znajdzie siê w komórce potomnej wynosi
0.5. Je¿eli w dziel¹cej siê komórce wystêpuje n
cz¹steczek plazmidowych (liczba kopii = n) to
prawdopodobieñstwo, ¿e ¿aden z nich nie trafi
do komórki potomnej wynosi (0.5)
n
. Poniewa¿
ka¿dy podzia³ komórkowy prowadzi do po-
wstania dwóch komórek, to prawdopodobie-
ñstwo, ¿e którakolwiek z nich nie bêdzie za-
wieraæ plazmidu (czyli tzw. czêstoœæ segregacji
plazmidu) wynosi: P = 2(0.5)
n
= 2
(1–n)
(
NORD-
STRÖM
i A
USTIN
1989). U¿ywaj¹c powy¿szego
równania porównajmy dwa plazmidy, z któ-
rych jeden wystêpuje w dwóch, a drugi w dwu-
dziestu kopiach w komórce. Komórki bezpla-
zmidowe powinny pojawiæ siê w przypadku
pierwszego z nich ju¿ po zajœciu dwóch po-
dzia³ów, a w przypadku drugiego z nich czê-
stoœæ segregacji wynosi mniej ni¿ 10
–6
, a wiêc
jest praktycznie niewykrywalna.
W naturze, równie¿ plazmidy wystêpuj¹ce
w niskiej liczbie kopii s¹ niezwykle trwale
utrzymywane w populacjach bakteryjnych,
niezale¿nie od braku presji selekcyjnej. Ozna-
cza to, ¿e musz¹ istnieæ specyficzne mechani-
zmy skutecznie przeciwdzia³aj¹ce utracie pla-
zmidów przez dziel¹ce siê komórki. Mechani-
zmy te mo¿na podzieliæ, w zale¿noœci od sposo-
bu dzia³ania, na trzy grupy: A, B i C (Ryc. 1).
Do grupy A nale¿¹ systemy miejscowo-spe-
cyficznej rekombinacji, dziêki którym, formy
oligomeryczne plazmidów powsta³e g³ównie
na drodze rekombinacji pomiêdzy plazmida-
mi, s¹ rozdzielane na pojedyncze cz¹steczki
plazmidowe, podlegaj¹ce nastêpnie losowej
dystrybucji do komórek potomnych.
Grupa B obejmuje systemy aktywnego roz-
dzia³u plazmidów (tzw. partycji) do dziel¹cych
siê komórek zapewniaj¹ce, ¿e w trakcie po-
dzia³u ka¿da komórka potomna otrzyma pla-
zmid.
W grupie C wystêpuj¹ tzw. mechanizmy ad-
dykcyjne, a wiêc uzale¿niaj¹ce komórki gospo-
darzy od wystêpuj¹cych w nich plazmidów. Po-
woduj¹ one œmieræ lub zahamowanie wzrostu
powsta³ych po podziale komórek bezplazmi-
dowych.
Nale¿y podkreœliæ, ¿e mechanizmy grupy A
wspomagaj¹ jedynie rozdzia³ losowy plazmi-
dów, podczas gdy te z grup B i C zapewniaj¹
dziedziczenie lepsze ni¿ losowe. Ponadto, o ile
mechanizmy A i B maj¹ bezpoœredni wp³yw na
Tom 51,
2002
Numer 3
(256)
Strony 297–304
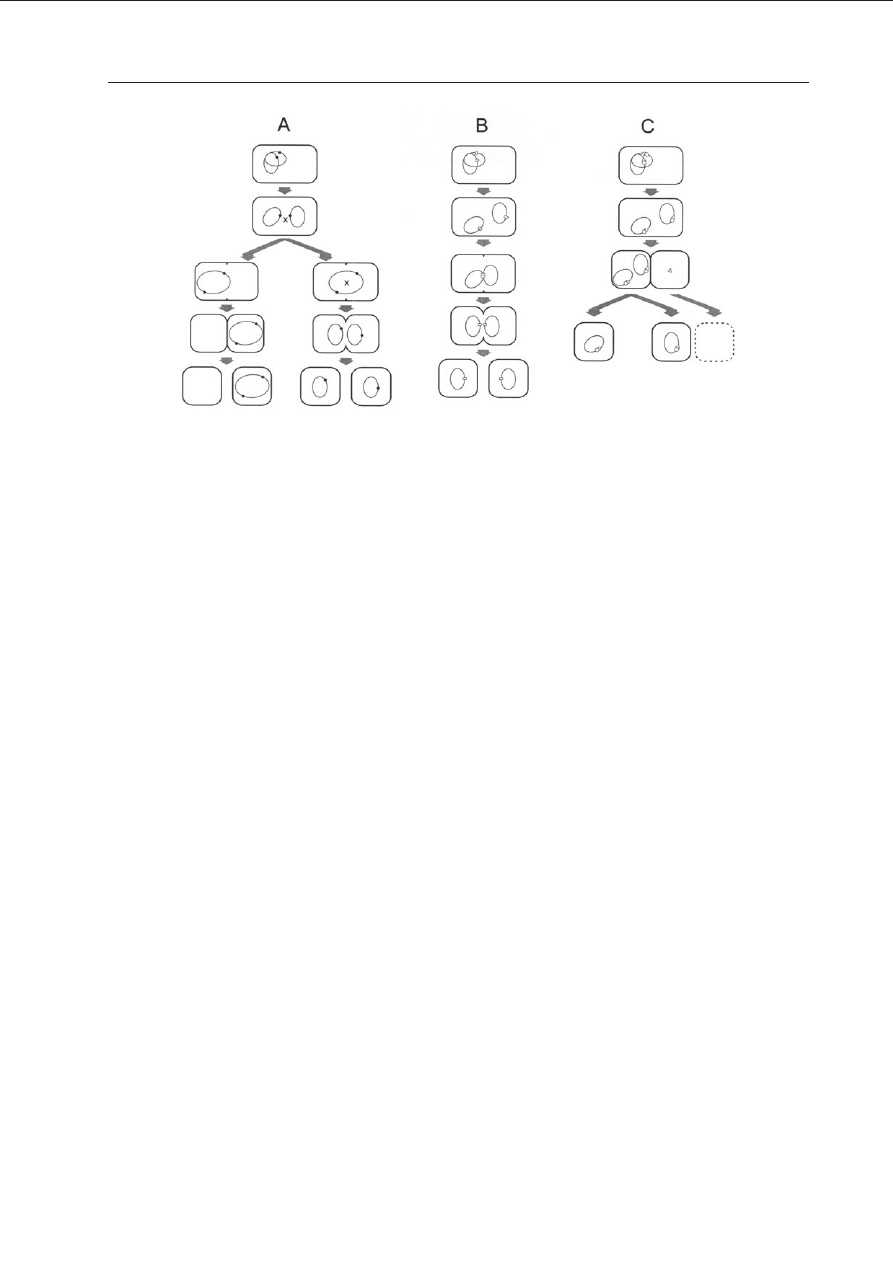
tzw. stabilnoœæ segregacyjn¹ plazmidu i dzia³aj¹
na poziomie komórkowym, to mechanizm C —
poprzez eliminacjê komórek pozbawionych
plazmidów — dzia³a na poziomie populacyjnym.
Czasem u plazmidów, które zazwyczaj wystê-
puj¹ w 1–2 kopiach w przeliczeniu na chromo-
som gospodarza, funkcjonuj¹ wszystkie trzy ro-
dzaje mechanizmów wspomagaj¹cych dziedzi-
czenie, a niektóre z nich mog¹ byæ jeszcze zdu-
plikowane, co prowadzi, ca³kowicie na przekór
regu³om rachunku prawdopodobieñstwa, do
ich niezwyk³ej stabilnoœci segregacyjnej.
Poni¿ej nieco bardziej szczegó³owo omó-
wimy mechanizmy z grup A i C, natomiast sys-
temom partycji plazmidów poœwiêcony jest ar-
tyku³ M. £
OBOCKIEJ
i wspó³autorów w tym
zeszycie KOSMOSU. Tutaj nale¿y jedynie pod-
kreœliæ, ¿e systemy z grupy B s¹ niezwykle pre-
cyzyjne i skutecznie zapewniaj¹ wysok¹ stabil-
noœæ segregacyjn¹ plazmidów.
SYSTEMY MIEJSCOWO-SPECYFICZNEJ REKOMBINACJI
Identyczne kopie plazmidów czêsto ule-
gaj¹ miêdzy sob¹ rekombinacji i tworz¹ tym sa-
mym tzw. dimery lub formy oligomeryczne wy-
¿szego rzêdu (trimery, tetramery itd.). Tym sa-
mym zmniejsza siê liczba jednostek plazmido-
wych podlegaj¹cych segregacji do komórek
potomnych. Warto tu zaznaczyæ, ¿e system
kontroli liczby kopii plazmidu, który dzia³a po-
przez „odliczanie” miejsc startu replikacji (ori)
„nie dostrzega”, ¿e np. w dimerze dwa ori wy-
stêpuj¹ w jednej cz¹steczce DNA i nie urucha-
mia mechanizmów koryguj¹cych liczbê kopii
plazmidu. Natomiast, poniewa¿ ori wybierane
s¹ losowo do replikacji, prawdopodobieñstwo
wybrania w mieszaninie monomerów i dime-
rów plazmidowych danego ori do replikacji
jest dla dimeru 2-razy wy¿sze ni¿ dla monome-
ru. W konsekwencji, liczba powstaj¹cych dime-
rów bêdzie wzrastaæ. Wynikiem tej sytuacji,
okreœlanej mianem „katastrofy dimerowej”
(S
UMMERS
i wspó³aut. 1993) jest czêstsze ni¿
oczekiwane powstawanie komórek bezplazmi-
dowych.
Bardzo czêsto plazmidy koduj¹ specjalny
system miejscowo-specyficznej rekombinacji,
tzw. mrs (ang. multimer resolution system) sku-
tecznie przeciwdzia³aj¹cy utrzymywaniu siê
form multimerycznych plazmidów w komórce.
Proces przekszta³cania ich w formy monome-
ryczne odbywa siê na drodze wewn¹trzplazmi-
dowej rekombinacji. Przeprowadzaj¹ go miej-
scowo-specyficzne rekombinazy (tzw. resolwa-
zy) rozpoznaj¹ce okreœlone miejsca w plazmi-
dach, tzw. sekwencje res, które obejmuj¹ zwy-
298
U
RSZULA
Z
IELENKIEWICZ
i P
IOTR
C
EG£OWSKI
Ryc. 1. Mechanizmy stabilnego dziedziczenia plazmidów.
Dla uproszczenia wszystkie trzy mechanizmy przedstawiono dla plazmidów wystêpuj¹cych w jednej kopii w
przeliczeniu na chromosom gospodarza. (A) Miejscowo-specyficzna rekombinacja; ciemne kó³ka na plazmidach
oznaczaj¹ miejsca res; litera x pomiêdzy plazmidami lub wewn¹trz dimeru plazmidowego oznacza zachodzenie
rekombinacji. (B) Partycja; jasne prostok¹ty oznaczaj¹ tzw. miejsca centromerowe. (C) Mechanizm addykcyjny
czyli po-segregacyjne zabijanie komórek bezplazmidowych; kontury narysowane przerywan¹ lini¹ oznaczaj¹
œmieræ komórki na skutek utraty plazmidu.
kle kilkadziesi¹t par zasad. Systemy mrs s¹ za-
zwyczaj w ca³oœci kodowane przez plazmidy
(np. loxP-cre plazmidu P1, A
USTIN
i wspó³aut.
1981;
parCBA plazmidu RK2, G
RINTER
i
wspó³aut. 1989). Znane s¹ jednak przypadki, ¿e
plazmid (np. plazmid ColE1) zawiera tylko miej-
sce res, a do rozdzia³u oligomerów wykorzystu-
je rekombinazê kodowan¹ przez genom gospo-
darza (S
UMMERS
i S
HERRATT
1984). Zdarza siê
te¿, ¿e kodowany przez plazmid system mrs wy-
maga chromosomalnego histono-podobnego
bia³ka komórkowego (np. bia³ko Hbsu B. subti-
lis dla rekombinazy beta plazmidu pSM19035,
A
LONSO
i wspó³aut. 1995).
Systemy miejscowo-specyficznej rekombi-
nacji s¹ czêsto jedynymi dodatkowymi mecha-
nizmami stabilizuj¹cymi plazmidy wystêpuj¹ce
w œredniej (15-30) liczbie kopii. Jednak¿e, w
po³¹czeniu z precyzyjn¹, pozbawion¹ fluktu-
acji kontrol¹ liczby kopii skutecznie zapobie-
gaj¹
one
utracie
plazmidów
z
populacji
(S
UMMERS
1998).
SYSTEMY ADDYKCYJNE
Chocia¿ powy¿ej opisane procesy zasadni-
czo zapewniaj¹ trwa³e dziedziczenie plazmi-
dów, to jednak z nisk¹ czêstoœci¹ pojawiaj¹ siê
komórki
bezplazmidowe.
Brak
plazmidu
zmniejsza obci¹¿enie metaboliczne komórki
co powoduje, ¿e bezplazmidowe bakterie
dziel¹c siê szybciej zdominowa³yby ca³¹ popu-
lacjê doprowadzaj¹c praktycznie do eliminacji
komórek plazmidowych. Bardzo szczególnym
sposobem
unikniêcia
takiej
sytuacji
jest
dzia³anie systemów addykcyjnych, zwanych
równie¿ systemami po-segregacyjnego zabija-
nia komórek bezplazmidowych, trucizny i od-
trutki, czy zaprogramowanej œmierci komórko-
wej. Systemy te doprowadzaj¹ do eliminacji ko-
mórki potomnej je¿eli nie odziedziczy³a ona
plazmidu. Ideê addykcji, czyli uzale¿nienia ko-
mórek od w³asnych plazmidów zaproponowa³
Koyama (K
OYAMA
i wspó³aut. 1975) zauwa-
¿aj¹c, ¿e je¿eli komórka trac¹ca plazmid ginie,
to w populacji nigdy nie znajdzie siê pozbawio-
nego plazmidów potomstwa.
Molekularn¹ podstaw¹ funkcjonowania sys-
temów addykcyjnych jest istnienie przynajm-
niej dwóch genów plazmidowych, warun-
kuj¹cych powstawanie trwa³ego czynnika tok-
sycznego oraz odtrutki — nietrwa³ego czynnika
przeciwdzia³aj¹cego powstawaniu lub dzia³a-
niu trucizny. Trucizna zawsze jest bia³kiem, a
odtrutk¹ mo¿e byæ albo bia³ko albo tzw. anty-
sensowny RNA. W komórce posiadaj¹cej pla-
zmid przebiega ci¹g³a, kontrolowana synteza
odtrutki zapewniaj¹ca jej nadmiar, dziêki cze-
mu komórka jest chroniona przed dzia³aniem
trucizny. Jednak¿e, z chwil¹ utraty plazmidu,
iloœæ odtrutki szybko siê zmniejsza na skutek de-
gradacji przez odpowiedni enzym hydrolitycz-
ny i dochodzi do syntezy lub uwolnienia z kom-
pleksu aktywnej trucizny. Pozbawiona plazmi-
du komórka wiêc ginie, a w populacji pozostan¹
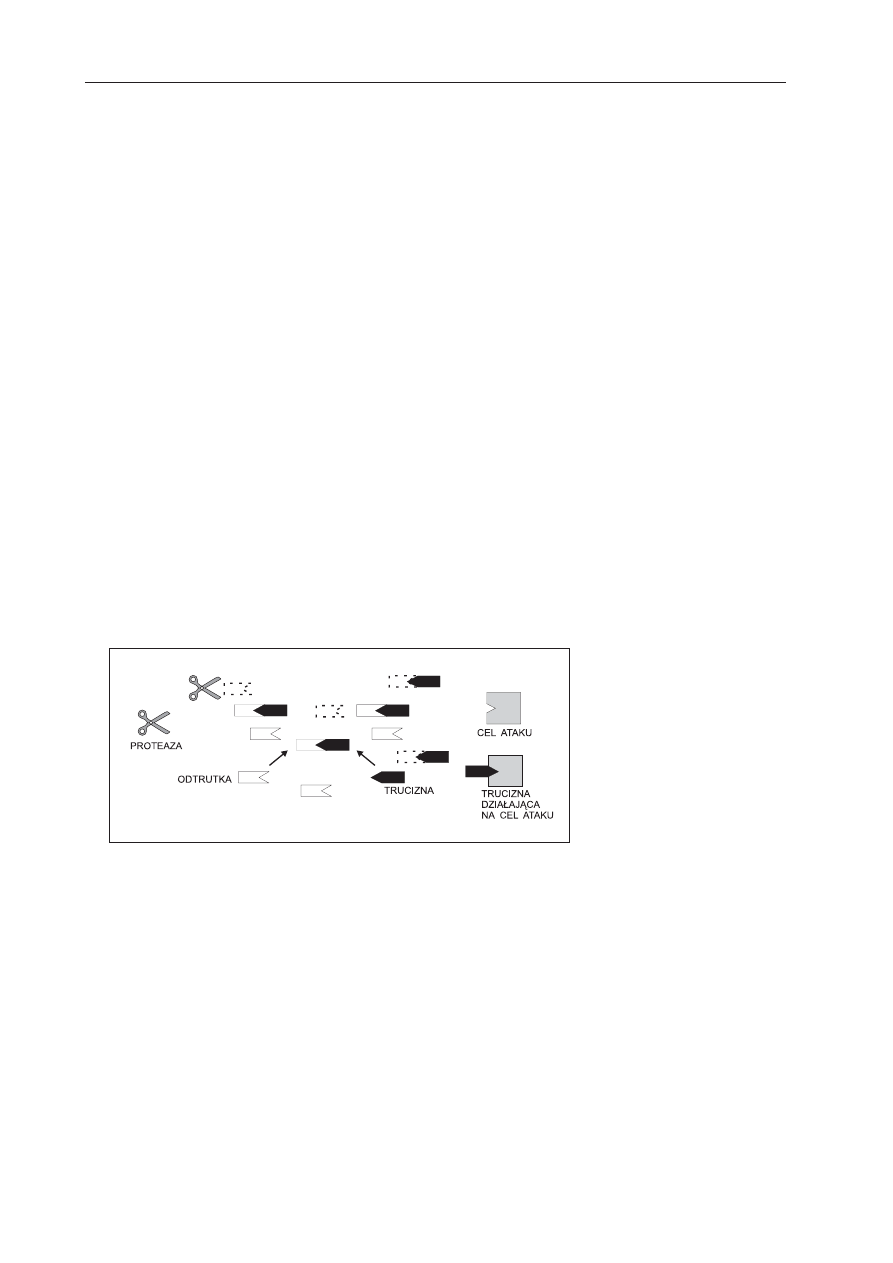
jedynie komórki zawieraj¹ce plazmid (Ryc. 2).
SYSTEMY ADDYKCYJNE REGULOWANE PRZEZ
ANTYSENSOWNY RNA
Systemy addykcyjne regulowane przez an-
tysensowny RNA stanowi¹ spójn¹ grupê okre-
œlan¹ mianem rodziny hok-sok (ang. host kil-
ling, suppression of killing) opisuj¹cych geny
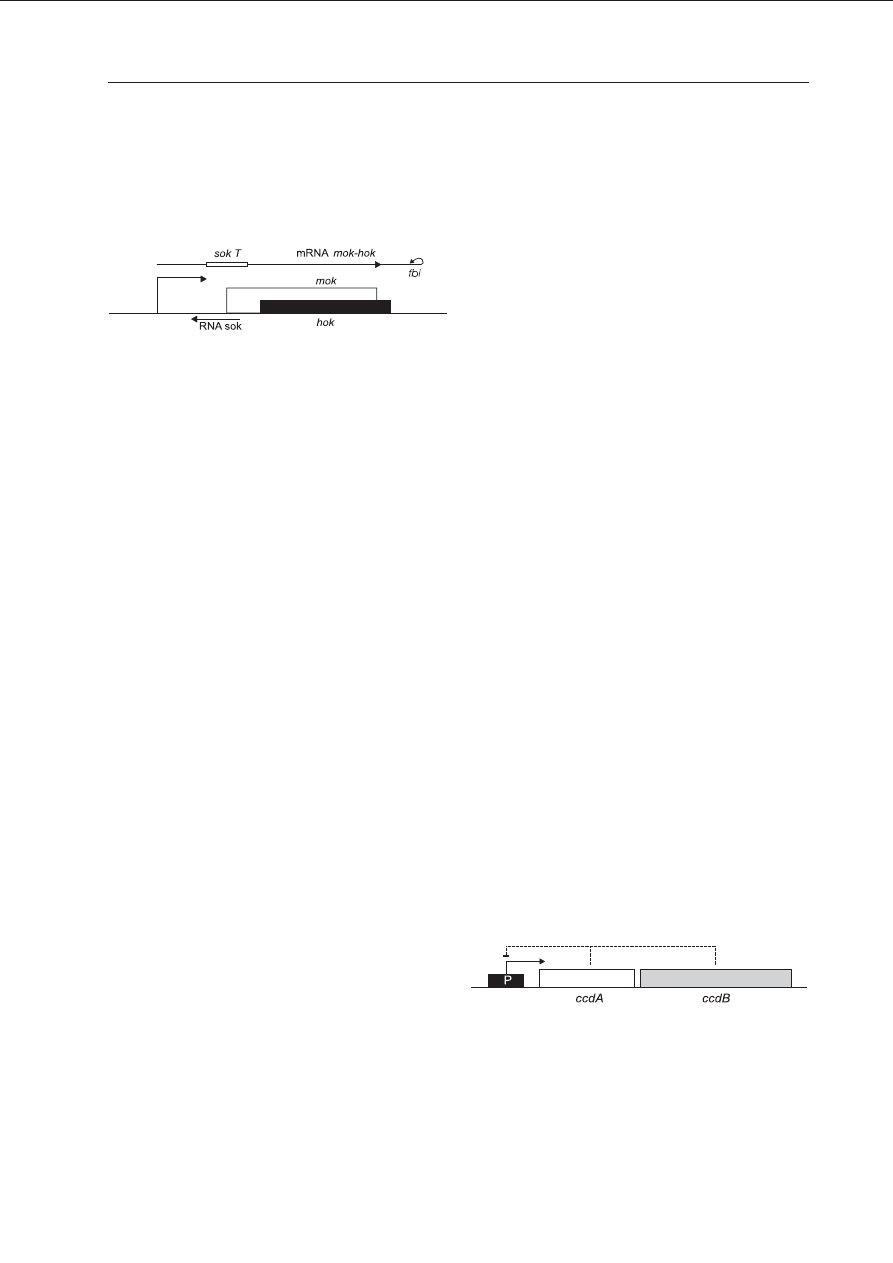
znalezione na plazmidzie R1 E. coli. Model or-
ganizacji genetycznej locus hok-sok przedsta-
wiono na Ryc. 3. Zawiera on trzy nak³adaj¹ce
siê geny: hok — koduj¹cy truciznê, mok - regu-
luj¹cy translacjê trwa³ego mRNA trucizny oraz
sok — koduj¹cy nietrwa³y, antysensowny RNA,
który jest komplementarny do obszaru 5’
mRNA mok-hok. mRNA trucizny zawdziêcza
swoj¹ wysok¹ trwa³oœæ wystêpuj¹cej na jego
Mechanizmy stabilnego dziedziczenia plazmidów
299
Ryc.
2.
Funkcjonowanie
plazmidowych
systemów
addykcyjnych u bakterii.
Na przedstawionym schema-
cie trucizna i odtrutka s¹
bia³kami. Kontur narysowany
przerywan¹ lini¹ oznacza de-
gradacjê odtrutki przez specy-
ficzn¹ proteazê.
koñcu 3’ strukturze fbi (ang. fold back inhibi-
tion). Ekspresja genu hok jest negatywnie regu-
lowana przez RNA Sok, który ³¹cz¹c siê z kom-
plementarnym odcinkiem mRNA genu mok
sprawia, ¿e utworzony kompleks, reprezen-
tuj¹cy dwuniciowy RNA, jest rozcinany przez
RNazê III i tym samym nie dochodzi do transla-
cji bia³ek Mok i Hok. W komórce bezplazmido-
wej istniej¹cy RNA Sok zostaje zdegradowany,
a niezwi¹zany mRNA hok ulega translacji. Po-
wstaj¹ce ma³e, zbudowane z 52 aminokwasów,
b³onowe bia³ko Hok powoduje œmieræ komó-
rek poprzez zahamowanie oddychania, obni¿e-
nie potencja³u b³onowego i wyp³yw zwi¹zków
niskocz¹steczkowych (T
HISTED
i wspó³aut.
1995). Homologiczne geny hok-sok odnajdy-
wane s¹ na licznych plazmidach bakterii Gra-
m-ujemnych, a tak¿e w chromosomach wielu
bakterii, gdzie ich ewentualna funkcja stanowi
prawdziw¹ zagadkê.
Niedawno odkryto podobnie regulowany
poprzez antysensowny RNA, lecz niehomolo-
giczny do rodziny hok-sok, system stabilizacji
plazmidów u bakterii Gram-dodatnich: par pla-
zmidu pAD1 z Enterococcus faecalis (W
EAVER
i
wspó³aut. 1996). Region par koduje dwa prze-
ciwstawnie transkrybowane RNA: d³u¿szy RNA
I zawieraj¹cy gen toksyny fst (ang. faecalis pla-
smid-stabilizing peptide) oraz krótszy RNA II,
komplementarny do RNA I. Utworzenie kom-
pleksu obu RNA hamuje wytwarzanie trucizny
poprzez uniemo¿liwienie dostêpu rybosomów
do sekwencji inicjacji translacji fst. Równie¿ w
tym przypadku czas trwania czynnika zabez-
pieczaj¹cego RNA II jest znacznie krótszy ni¿
RNA I. Zarówno nadprodukcja RNA I, jak i pep-
tydu Fst powoduje œmieræ komórek, przed
któr¹ chroni dostarczenie nadmiaru RNA II
(G
REENFIELD
i wspó³aut. 2000).
BIA£KOWE SYSTEMY ADDYKCYJNE
W tym przypadku oba sk³adniki systemu
przeciwdzia³aj¹cego utracie plazmidów s¹
bia³kami. Organizacja genetyczna bia³kowych
kaset stabilizuj¹cych plazmidy jest zasadniczo
podobna — dwa (niekiedy trzy) geny stanowi¹
operon. Bardzo istotn¹ cech¹ tych operonów
jest autoregulacja transkrypcji: bia³ko odtrutki
samo lub w kompleksie z trucizn¹ dzia³a jako
represor. Wyj¹tkowo, w autoregulacji addyk-
cyjnego systemu
w-e-z plazmidu pSM19035 z
Streptococcus pyogenes nie bior¹ udzia³u ani
odtrutka ani trucizna, lecz oddzielne bia³ko re-
gulatorowe (D
E
L
A
H
OZ
i wspó³aut. 2000).
Bia³ka systemów addykcyjnych, poza jed-
nym wyj¹tkiem, s¹ raczej ma³e (70–130 amino-
kwasów), przy czym prawie zawsze bia³ka tru-
cizn s¹ nieco wiêksze od bia³ek odtrutek (Tabe-
la 1). Chocia¿ strukturalne i funkcjonalne po-
dobieñstwo licznych systemów addykcjnych
jest wyraŸne, nie ma znacz¹cych podobieñstw
sekwencji ich genów. Natomiast czêsto odnaj-
duje siê prawie identyczne kasety stabilizuj¹ce
wœród plazmidów pokrewnych, co zapewne
jest odbiciem horyzontalnego transferu genów
pomiêdzy ró¿nymi bakteriami i plazmidami
(G
ERDES
2000).
Jednym z najlepiej poznanych systemów
dzia³aj¹cych wed³ug przedstawionego powy-
¿ej schematu jest system addykcyjny ccd jedno-
kopiowego plazmidu F E. coli. Pocz¹tkowo zo-
sta³ opisany jako uzale¿niaj¹cy podzia³ komór-
kowy od wczeœniejszego zreplikowania pla-
zmidu (ang. coupled cell division, O
GURA
i
H
IRAGA
1983), a nastêpnie jako stabilizuj¹cy
utrzymywanie plazmidu poprzez eliminowa-
nie komórek bezplazmidowych (ang. control
of cell death, J
AFFE
i wspó³aut. 1985). W locus
ccd gen odtrutki ccdA poprzedza gen trucizny
ccdB. Ekspresja operonu jest negatywnie regu-
lowana poprzez kompleks bia³ek CcdA i CcdB
(Ryc. 4). Bia³ko toksyny CcdB doprowadza do
300
U
RSZULA
Z
IELENKIEWICZ
i P
IOTR
C
EG£OWSKI
Ryc. 3. Schemat systemu hok-sok kodowanego
przez plazmid R1.
Miejsce SokT w mRNA mok-hok oznacza region kom-
plementarny dla RNA Sok. W tym miejscu powstaje
dwuniciowy RNA, ktory jest degradowany przez RNazê
III.
Ryc. 4. Schemat systemu ccd kodowanego przez
plazmid F.
Zgiêta strza³ka wychodz¹ca z miejsca P oznacza pro-
motor transkrypcji. Jego aktywnoϾ jest reprymowana
przez kompleks utworzony przez produkty genów
ccdA ccdB.

œmierci komórek przez zablokowanie funkcjo-
nowania gyrazy — enzymu wprowadzaj¹cego
dodatkowe skrêty w helisie DNA. Funkcjono-
wanie gyrazy jest niezbêdne w wielu proce-
sach komórkowych, takich jak transkrypcja, re-
kombinacja czy rozdzia³ chromosomów do ko-
mórek potomnych. CcdB wi¹¿e siê trwale za-
równo do wolnej gyrazy, blokuj¹c jej aktyw-
noœæ, jak i do kompleksu gyrazy z naciêtym
DNA uniemo¿liwiaj¹c ponowne jego z³¹cze-
nie. Powsta³e w ten sposób pêkniêcia w DNA
wywo³uj¹ alarmow¹ reakcjê komórki, tzw. od-
powiedŸ SOS, polegaj¹c¹ m.in. na indukcji wie-
lu bia³ek naprawy DNA. Niemo¿noœæ usuniêcia
uszkodzeñ wywo³anych zablokowaniem ak-
tywnoœci gyrazy powoduje wstrzymanie po-
dzia³ów komórkowych, a nastêpnie œmieræ ko-
mórek.
Znane s¹ mutanty nietoksyczne bia³ka
CcdB oraz mutanty gyrazy niewra¿liwe na tê
truciznê. Dziêki temu oraz poznaniu krysta-
licznej struktury wiêkszej podjednostki gyra-
zy zaproponowano model tworzenia kom-
pleksu GyrA-CcdB, w którym C-koñcowy frag-
ment toksyny umieszczony jest w przestrzeni
centralnej dimeru gyrazy (L
ORIS
i wspó³aut.
1999).
Odtrutka CcdA nie tyko zapobiega tworze-
niu kompleksu CcdB z gyraz¹ wi¹¿¹c siê trwale
z trucizn¹, ale uwalnia równie¿ CcdB z ju¿ ist-
niej¹cych kompleksów. Odtrutka CcdA jest de-
gradowana przez proteazê Lon, a jej okres
trwania jest dwukrotnie krótszy ni¿ okres trwa-
nia trucizny CcdB.
Warto zaznaczyæ, ¿e system ccd nie jest je-
dynym przyczyniaj¹cym siê do trwa³ego utrzy-
mywania plazmidu F. Poza nim, plazmid F po-
siada kilka miejscowo-specyficznych rekombi-
naz, system partycji sop oraz dwa, homologicz-
ne do rodziny hok-sok, systemy regulowane an-
tysensownym RNA (N
ORDSTRÖM
i A
USTIN
1989). Niewielki, w porównaniu np. z sop, sto-
pieñ stabilizacji poprzez ccd, prawdopodobnie
wynika z tego, ¿e wywo³uje w pierwszej kolej-
noœci odpowiedŸ SOS, a nie natychmiastow¹
œmieræ komórek bezplazmidowych. Postuluje
siê wiêc, ¿e jego rol¹ mo¿e byæ, po czêœci, za-
pewnienie komórkom zmiennoœci genetycz-
nej (C
OUTURIER
i wspó³aut. 1998).
Pomimo intensywnych badañ prowadzo-
nych przez ostatnie lata nad ró¿nymi systema-
mi trucizna-antidotum tylko dla nielicznych z
nich uda³o siê ustaliæ molekularny mechanizm
dzia³ania trucizny. Obok opisanego powy¿ej
mechanizmu dzia³ania bia³ka CcdB plazmidu F
poznano tak¿e cel ataku trucizny systemu
pemI/pemK (inaczej okreœlanego jako kis/kid)
plazmidu R1 (R
UIZ
-E
CHEVARRIA
i wspó³aut.
1995). PemK (Kid) blokuje aktywnoœæ DnaB —
enzymu helikazy DNA, który rozplata dwuni-
ciowy DNA podczas inicjacji replikacji chro-
mosomu i niektórych plazmidów E. coli. Ostat-
nio ustalono, ¿e trucizna ParE plazmidu RK2,
podobnie jak CcdB, jest inhibitorem gyrazy
(J
IANG
i wspó³aut. 2002). Molekularne mecha-
nizmy dzia³ania trucizn innych systemów nie
zosta³y jeszcze ustalone. Nieco lepiej przedsta-
wia siê wiedza dotycz¹ca mechanizmów zwiê-
Mechanizmy stabilnego dziedziczenia plazmidów
301
Tabela 1. Wybrane przyklady bialkowych systemów stabilizujacych utrzymywanie plazmidów; kom-
pilacja wg G
ERDESA
2000 oraz
Z
IELENKIEWICZ
i
C
EG£OWSKIEGO
2001.
System
Plazmid
Trucizna
Odtrutka
Proteaza degra-
duj¹ca odtrutkê
Cel
ccd
F
CcdB (101)
CcdA (72)
Lon
gyraza
pem/parD
R100/R1
PemK/Kid (110)
PemI/Kis (84)
Lon
helikaza (DnaB)
phd-doc
P1
Doc (126)
Phd (76)
ClpXP
translacja?
parDE
RK2/RP4
ParE (103)
ParD (83)
nieznana
gyraza
pas
pTF-FC2
PasB (90)
PasA (74)
Lon
nieznana
relBE/stbDE
P307/R485
RelE/StbE (95)
RelB/StbD (83)
Lon
translacja?
hig
Rts1
HigB (92)
HigA (104)
nieznana
nieznana
w-e-z
pSM19035
Zeta (287)
Epsilon (90)
nieznana
nieznana
W nawiasach podano wielkoœci bia³ek w liczbach aminokwasów.

kszonej wra¿liwoœci odtrutek w porównaniu
do toksyn plazmidów E. coli — s¹ one najczê-
œciej degradowane przez proteazê Lon, a od-
trutka Phd systemu phd/doc jest substratem
dla proteazy ClpXP (Tabela1).
GENY TRUCIZNA-ODTRUTKA UMIEJSCOWIONE W
CHROMOSOMACH
Wiele plazmidowych genów addykcyjnych
ma swoje homologi w chromosomach bakte-
ryjnych. Systematyczne przeszukiwania kom-
puterowych baz danych pokazuj¹, ¿e s¹ one
obecne w ka¿dej grupie taksonomicznej Proka-
ryota tak¿e u Archaea. Niekiedy liczne, po-
krewne geny systemów trucizna-odtrutka po-
wtórzone s¹ w tym samym chromosomie
(G
ERDES
, 2000). Rola jak¹ geny te mog¹ pe³niæ
w chromosomach bakterii jest ca³kowicie nie-
jasna. Ich funkcjonowanie jako systemów
trwa³ego dziedziczenia chromosomów nie
mia³oby ¿adnego sensu, bo komórki pozbawio-
ne chromosomów tak czy inaczej czeka nie-
chybna œmieræ. Jednak¿e, chromosomalne ho-
mologi genów addykcyjnych, przeniesione na
plazmidy wydatnie zwiêkszaj¹ ich stabilnoœæ
segregacyjn¹ (A
IZENMAN
i wspó³aut. 1996,
G
OTFREDSEN
i G
ERDES
1998).
BAKTERIOCYNY ORAZ INNE CZYNNIKI PLAZMIDOWE DZIA£AJ¥CE Z ZEWN¥TRZ
Nazw¹ bakteriocyny oznacza siê produko-
wane przez bakterie zwi¹zki hamuj¹ce wzrost
s¹siaduj¹cych komórek zazwyczaj blisko spo-
krewnionych gatunków. Czêsto zdolnoœæ pro-
dukowania truj¹cej bakteriocyny niesiona jest
przez plazmidy, zawsze razem z odpowiednim
genem(i) koduj¹cym(i) odpornoœæ na dzia³a-
nie tego czynnika na komórkê producenta.
Bia³ka bakteriocyn wydzielane s¹ na zewn¹trz
produkuj¹cych je komórek i adsorbowane na
powierzchni komórek wra¿liwych, których
wzrost ulega zahamowaniu. Produkuj¹ce bak-
teriocyny komórki s¹ na nie niewra¿liwe, po-
niewa¿ jednoczeœnie aktywne s¹ geny warun-
kuj¹ce opornoœæ. Rezultatem jest wzglêdna
nieobecnoœæ komórek bezplazmidowych, nie-
chronionych przed obecn¹ w pod³o¿u tru-
cizn¹. Podobieñstwo dzia³ania bakteriocyn i
systemów addykcji jest tylko powierzchowne.
Plazmidowe systemy addykcyjne nie dopusz-
czaj¹ do powstania bezplazmidowego potom-
stwa, natomiast produkowanie toksyn zew-
nêtrznych broni g³ównie komórki posiadaj¹ce
plazmid przed konkurencj¹ nowych, bezpla-
zmidowych komórek.
SYSTEMY RESTRYKCJI-MODYFIKACJI JAKO CZYNNIKI STABILNOŒCI PLAZMIDÓW
Systemy restrykcji-modyfikacji DNA (R-M)
wystêpuj¹ powszechnie u Prokaryota zarówno
w chromosomach, jak i na plazmidach. Para ge-
nów R-M koduje dwa enzymy: endonukleazê
(R) rozpoznaj¹c¹ specyficzn¹ krótk¹ sekwen-
cjê nukleotydow¹ w dwuniciowym DNA i
zdoln¹ do jego przeciêcia w obrêbie lub po-
bli¿u tej sekwencji oraz metylazê (M), która po-
przez metylacjê tej sekwencji czyni DNA nie-
wra¿liwym na dzia³anie endonukleazy restryk-
cyjnej. Przyjmuje siê, ¿e obecnoœæ systemów
R-M zabezpiecza komórkê przed inwazj¹ obce-
go DNA (np. wirusów czy plazmidów). Jednak
hipoteza „obrony komórkowej” nie wyjaœnia
ró¿norodnoœci ani specyficznoœci enzymów
restrykcyjnych, zw³aszcza tych rozpoznaj¹cych
sekwencje 8-nukleotydowe, które niezwykle
rzadko lub w ogóle nie wystêpuj¹ w ma³ych ge-
nomach plazmidów i wirusów, przed którymi
maj¹ chroniæ komórki.
N
AITO
i wspó³aut. (1995) wykazali, ¿e obec-
noœæ genów systemów R-M, EcoRI lub PaeR7 w
plazmidzie wydatnie podnosi jego stabilnoϾ
segregacyjn¹. Proponowany mechanizm stabi-
lizacyjny równie¿ funkcjonuje na zasadzie
po-segregacyjnego zabijania komórek bezpla-
zmidowych, jednak nale¿y podkreœliæ tu wyra-
Ÿn¹ ró¿nicê w porównaniu do dzia³ania kla-
sycznych systemów addykcyjnych. W przypad-
ku systemów R-M nie jest potrzebna ró¿nica w
stabilnoœci metylazy DNA (odpowiednik od-
trutki) i endonukleazy restrykcyjnej (odpo-
wiednik trucizny). Obydwa enzymy mog¹
mieæ tak¹ sam¹ trwa³oœæ w komórce, jednak
postêpuj¹ca ich inaktywacja lub rozcieñczanie
w dziel¹cych siê komórkach bezplazmidowych
osi¹gaj¹ takie stadium, ¿e niektóre sekwencje
w chromosomie nie zostan¹ ju¿ zmetylowane.
Do przeciêcia chromosomu w niezmetylowa-
nych sekwencjach i wywo³ania œmierci komór-
302
U
RSZULA
Z
IELENKIEWICZ
i P
IOTR
C
EG£OWSKI

ki wystarczy tu nawet œladowa aktywnoœæ en-
donukleazy restrykcyjnej.
Widziane w ten sposób systemy R-M, po-
dobnie jak systemy addykcyjne, rozwa¿ane s¹
jako samolubne elementy, które utrwali³y siê w
ewolucji jako modu³y zaprogramowanej œmier-
ci komórek, a które zosta³y wykorzystane przez
plazmidy do zapewniania ich trwa³ego bytowa-
nia w komórkach bakterii (Y
ARMOLINSKY
1995, K
OBAYASHI
1998).
MECHANISMS OF PLASMID STABLE INHERITANCE
S u m m a r y
The stable inheritance of bacterial plasmids is
achieved by a number of different mechanisms.
Among them are: resolution of plasmid multimers into
monomers, selective killing of plasmid-free segregants
and active plasmid partitioning into dividing cells. The
first two mechanisms are discussed in this article. The
multimer resolution systems (mrs) consist of a site
specific recombinase (resolvase) and the defined nu-
cleotide sequence res located on the plasmid. By spe-
cific recombination between repeated res sequences
the recombinase resolves plasmid oligomers to mono-
mers. This maximizes the number of plasmid units
prior to cell division and considerably contributes to
stable maintenance of plasmid in bacterial cells. The
post-segregational killing systems involve a stable poi-
son and an unstable antidote. The antidotes nautralize
their cognate poisons or prevent their synthesis. The
different decay rates of the poisons and the antidotes
underlie the molecular mechanisms of poison activa-
tion in plasmid-free cells. By killing and eliminating
plasmid-free cells from the population of plasmid-
bearing ones the poison-antidote couples act there-
fore as plasmid addiction systems. While the mrs maxi-
mize the random plasmid distribution into the divid-
ing cells addiction systems assure better-than-random
plasmid distribution.
LITERATURA
A
IZENMAN
E., E
NGELBERG
-K
ULKA
H., G
LASER
G
.,
1996. An
Escherichia coli chromosomal “addiction modu-
le” regulated by guanosine 3’,5’-bispyrophospha-
te: a model for programmed bacterial cell death.
Proc. Natl. Acad. Sci. USA 93, 6059–6063.
A
LONSO
J. C., W
EISE
F., R
OJO
F.
,
1995. The Bacillus subti-
lis histone-like protein Hbsu is required for DNA
resolution and DNA inversion mediated by the
beta recombinase of plasmid pSM19035. J. Biol.
Chem. 270, 2938–2945.
A
USTIN
S., Z
IESE
M., S
TERNBERG
N
.
, 1981. A novel role for
site-specific recombination in maintenance of
bacterial replicons. Cell 25, 729–736.
C
OUTURIER
M., B
AHASSI
E., V
AN
M
ELDEREN
L., 1998. Bacte-
rial death by DNA gyrase poisoning. Trends
Microbiol. 6, 269–275.
D
E
L
A
H
OZ
A. B., A
YORA
S., S
ITKIEWICZ
I., F
ERNANDEZ
S.,
P
ANKIEWICZ
R., A
LONSO
J. C., C
EG£OWSKI
P
.,
2000.
Plasmid copy-number control and better-than-
random segregation genes of pSM19035 share a
common regulator. Proc. Natl. Acad. Sci. USA 97,
728–733.
G
ERDES
K., 2000. Toxin-antitoxin modules may regula-
te synthesis of macromolecules during nutritio-
nal stress. J. Bacteriol. 182, 561–572.
G
OTFREDSEN
M., G
ERDES
K. 1998. The Escherichia coli
relBE genes belong to a new toxin-antitoxin gene
family. Mol. Microbiol. 29, 1065–1076.
G
REENFIELD
T. J., E
HLI
E., K
IRSHENMANN
T., F
RANCH
T.,
G
ERDES
K., W
EAVER
K. E., 2000. The antisense RNA
of the par locus of pAD1 regulates the expression
of a 33-amino-acid toxic peptide by an unusual
mechanism. Mol. Microbiol. 37, 652–660.
G
RINTER
N. J., B
REWSTER
G., B
ARTH
P. T., 1989. Two me-
chanisms necessary for the stable inheritance of
plasmid RP4. Plasmid 22, 203–214.
J
AFFE
A., O
GURA
T., H
IRAGA
S., 1985. Effects of the ccd
function of the F plasmid on bacterial growth. J.
Bacteriol. 163, 841–849.
J
IANG
Y., P
OGLIANO
J., H
ELINKI
D.R., K
ONIECZNY
I., 2002.
ParE toxin encoded by the broad-host-range pla-
zmid RK2 is an inhibitor of Escherichia coli gyra-
se. Mol. Microbiol. 44, 971–979.
K
OBAYASHI
I., 1998. Selfishness and death: raison d’-
etre of restriction, recombination and mitochon-
dria. Trends Genet. 14, 368–374.
K
OYAMA
A. H., W
ADA
C., N
AGATA
T., Y
URA
T., 1975. In-
direct selection for plasmid mutants: isolation of
ColVBtrp mutants defective in self-maintenance
in Escherichia coli. J. Bacteriol. 122, 73–79.
L
ORIS
R., D
AO
-T
HI
M. H., B
AHASSI
E. M., V
AN
M
ELDEREN
L.,
P
OORTMANS
F., L
IDDINGTON
R., C
OUTURIER
M., W
YNS
L., 1999. Crystal structure of CcdB, a topoisomera-
se poison from E. coli. J. Mol. Biol. 285, 167–1677.
N
AITO
T., K
USANO
K., K
OBAYASHI
I., 1995. Selfish be-
havior of restriction-modification systems. Scien-
ce 267, 897–899.
N
ORDSTROM
K., A
USTIN
S. J., 1989. Mechanisms that con-
tribute to the stable segregation of plasmids.
Annu. Rev. Genet. 23, 37–69.
O
GURA
T., H
IRAGA
S., 1983. Mini-F plasmid genes that
couple host cell division to plasmid proliferation.
Proc. Natl. Acad. Sci. USA 80, 4784–4788.
R
UIZ
-E
CHEVARRIA
M. J., G
IMENEZ
-G
ALLEGO
G., S
ABARIEGOS
-
J
ARENO
R., D
IAZ
-O
REJAS
R.
,
1995. Kid, a small prote-
in of the parD stability system of plasmid R1, is an
inhibitor of DNA replication acting at the initia-
tion of DNA synthesis. J. Mol. Biol. 247, 568–577.
S
UMMERS
D. K
.,
1998. Timing, self-control and a sense of
direction are the secrets of multicopy plasmid sta-
bility. Mol. Microbiol. 29, 1137–1145.
Mechanizmy stabilnego dziedziczenia plazmidów
303

S
UMMERS
D. K., B
ENTON
W. H., W
ITHERS
H. L., 1993. Mul-
ticopy plasmid instability: the dimer catastrophe
hypothesis. Mol. Microbiol. 8, 1031–1038.
S
UMMERS
, D. K., S
HERRATT
D. J., 1984. Multimerization
of high copy number plasmids causes instability:
CoIE1 encodes a determinant essential for pla-
smid monomerization and stability. Cell 36,
1097–1103.
T
HISTED
T., S
ORENSEN
N. S., G
ERDES
K., 1995. Mechanism
of Post-segregational Killing: Secondary Structure
Analysis of the Entire Hok mRNA from Plasmid R1
Suggests a Fold-back Structure that Prevents
Translation and Antisense RNA Binding. J. Mol.
Biol. 247, 1960–1968.
W
EAVER
K. E., J
ENSEN
K. D., C
OLWELL
A., S
RIRAM
S. I.,
1996. Functional analysis of the Enterococcus fae-
calis plasmid pAD1-encoded stability determi-
nant par. Mol. Microbiol. 20, 53–63.
Y
ARMOLINSKY
M. B., 1995. Programmed cell death in
bacterial populations. Science 267, 836–837.
Z
IELENKIEWICZ
U., C
EG£OWSKI
P., 2001. Mechanisms of
plasmid stable maintenance with special focus on
plasmid addiction systems. Acta Biochim. Polon.
48, 1003–1023.
304
U
RSZULA
Z
IELENKIEWICZ
i P
IOTR
C
EG£OWSKI
Wyszukiwarka
Podobne podstrony:
Mechanizmy chorób dziedzicznych 1
Mechanizmy chorˇb dziedzicznych
Mechanizmy chorób dziedzicznych 2
Podstawy mechanizmów zmienności i dziedziczenia
Mechanizmy dziedziczenia
mechanizmy dziedziczenia cukrzycy
Mechanizmy dziedziczenia odp
,gospodarka osadowa, Mechanizm procesu stabilizacji tlenowej
Wytrzymałość materiałów, Wytrzymałość materiałów - dziedzina wiedzy inżynierskiej, cześć mechaniki t
Mechanizm dziedziczenia, BIOLOGIA MOJE PRYWATNE
Wytrzymałość materiałów, Wytrzymałość materiałów - dziedzina wiedzy inżynierskiej, cześć inżynierii
Badanie stabilizatora parametrycznego, Akademia Morska -materiały mechaniczne, szkoła, Mega Szkoła,
STABIL, Akademia Morska -materiały mechaniczne, szkoła, Mega Szkoła, szkola1, III, AUTOMATY, STABILN
Stabilność układu, Studia PWR Tranposrt Mechaniczny Politechnika Wrocławska, Semestr VII, Automatyka
RYSOWANIE POŁĄCZEŃ CZĘŚCI MASZYNOWYCH pliki, Wytrzymałość materiałów, Wytrzymałość materiałów - dzie
więcej podobnych podstron