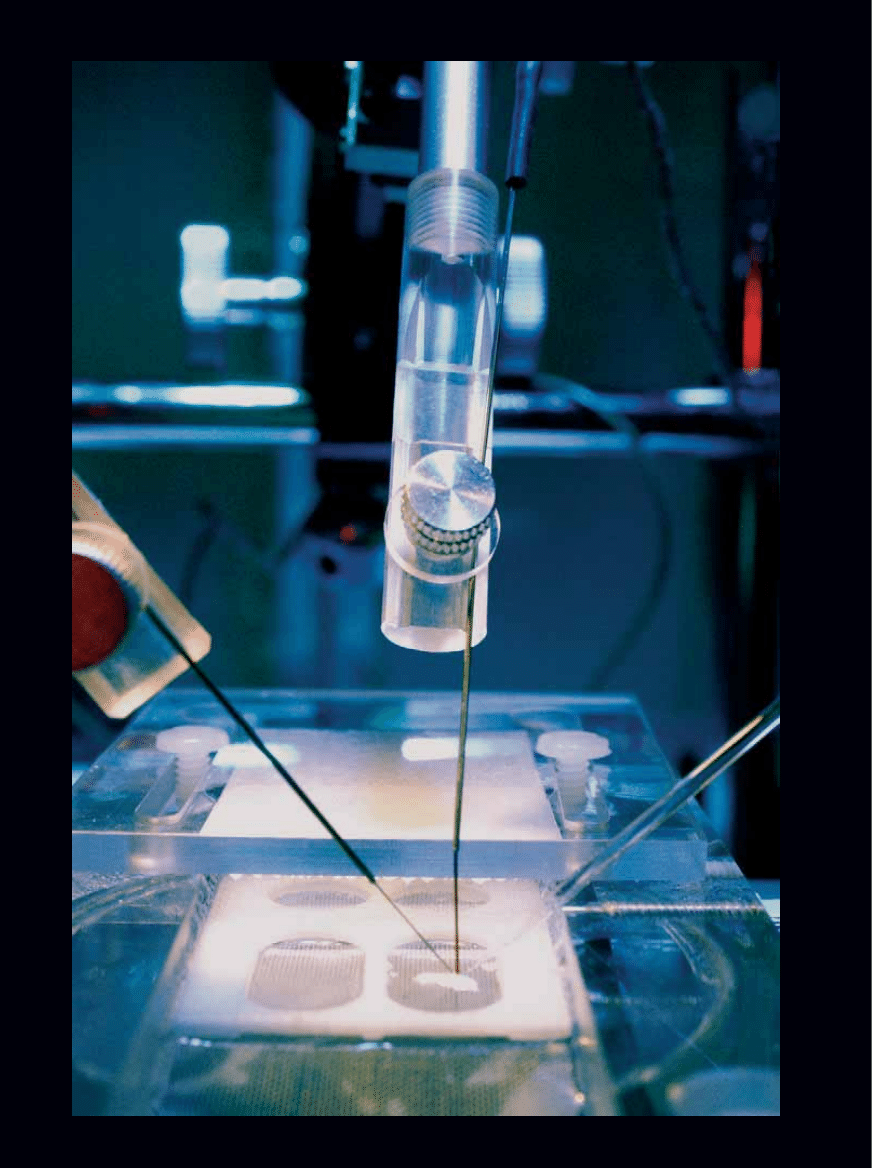
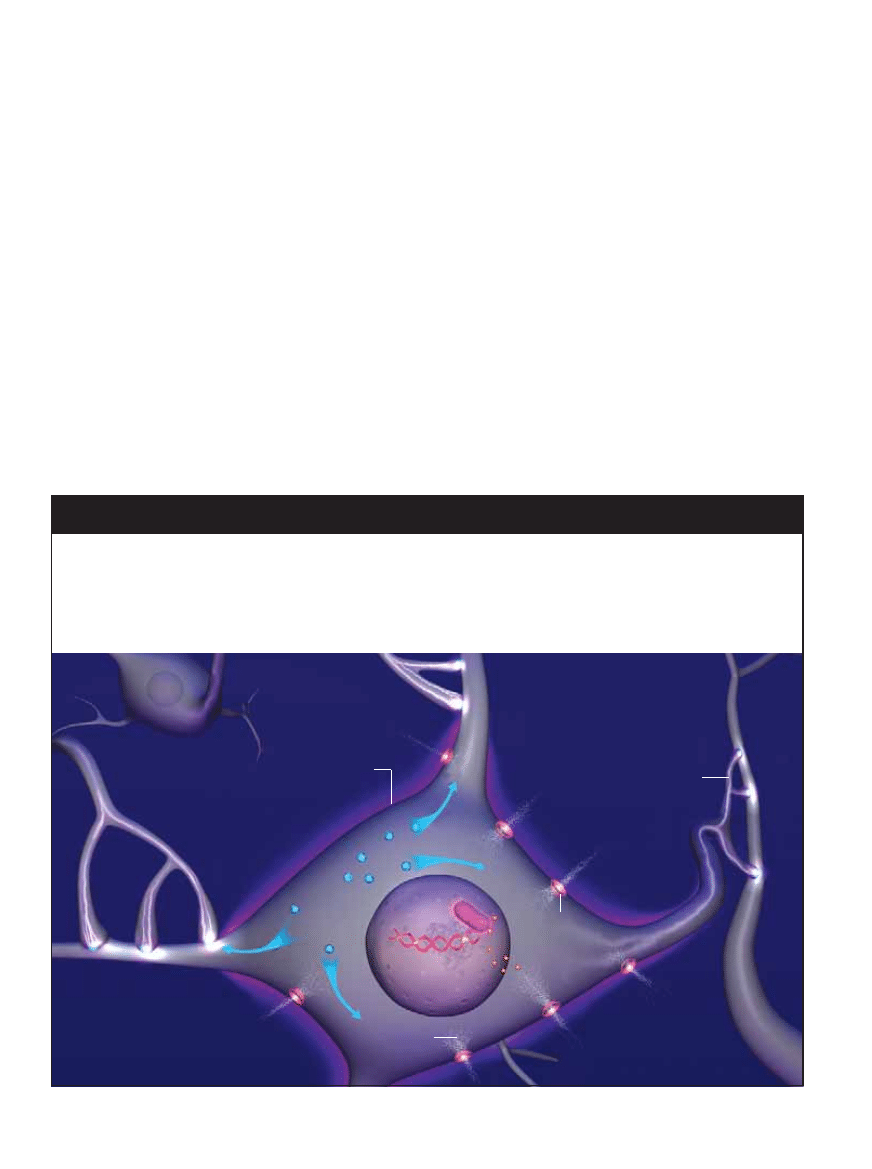
APARATURA elektrofizjologiczna
stymulujàc powstawanie pami´ci,
mo˝e pobudzaç i rejestrowaç sygna∏y
elektryczne w skrawkach hipokampa
wyizolowanego z mózgu szczura.

KA
Y CHERNUSH
W
thrillerze Memento g∏ówny bohater, Leonard,
pami´ta tylko to, co wydarzy∏o si´ do mo-
mentu urazu g∏owy, którego dozna∏ w noc
napaÊci na jego ˝on´. Osoby, które spotyka,
i wszystkie wydarzenia, które zasz∏y po tej
brzemiennej w skutki nocy, szybko ulatujà
z jego pami´ci. Leonard straci∏ zdolnoÊç
przetwarzania pami´ci krótkotrwa∏ej w d∏ugotrwa∏à. Pragnie
odnaleêç morderc´ ˝ony i pomÊciç jej Êmierç, jednak uwi´zio-
ny w teraêniejszoÊci musi znaleêç sposób na przechowanie
informacji kluczowych dla swojego Êledztwa. Wkrótce pokry-
wa tatua˝em ca∏e swoje cia∏o.
Ten poruszajàcy film zainspirowany zosta∏ historià cz∏owie-
ka znanego w literaturze medycznej jako HM, który w wieku
dziewi´ciu lat mia∏ wypadek rowerowy i w wyniku urazu
g∏owy zachorowa∏ na padaczk´. Aby z∏agodziç napady drgaw-
kowe, które nie ust´powa∏y po zastosowaniu innych metod,
lekarze zdecydowali si´ na usuni´cie cz´Êci mózgu zwanej hi-
pokampem oraz otaczajàcych jà struktur. Operacja rzeczywi-
Êcie przynios∏a ulg´, ale jednoczeÊnie upoÊledzi∏a niezbadany
mechanizm przetwarzania pami´ci krótkotrwa∏ej w d∏ugotrwa-
∏à. Informacje o ludziach, miejscach, wydarzeniach – czyli to,
co przechowujemy w mózgu jako pami´ç deklaratywnà – za-
nim zostanà zapisane w korze mózgowej, muszà przejÊç przez
hipokamp. Pami´ç o dawnych wydarzeniach, która zosta∏a
ju˝ zdeponowana w mózgu HM, pozosta∏a nietkni´ta, podczas
gdy wszystko to, co dzia∏o si´ w teraêniejszoÊci, szybko odp∏y-
wa∏o w niebyt. Pacjent co miesiàc odwiedza∏ swojego lekarza,
lecz za ka˝dym razem jego wizyta przebiega∏a tak, jakby si´
nigdy nie spotkali.
Problem przetwarzania wra˝eƒ odbieranych przez umys∏ w
trwa∏y Êlad pami´ciowy fascynuje neurobiologów ju˝ od
dawna. Nazwisko osoby, która w∏aÊnie zosta∏a nam przedsta-
wiona, przechowujemy w pami´ci krótkotrwa∏ej i po kilku
minutach mo˝emy mieç problem z jego przywo∏aniem, ale
takie informacje, jak nazwisko naszego najbli˝szego przyjacie-
la trafiajà do pami´ci d∏ugotrwa∏ej i zapewne nigdy ich nie tra-
cimy. Ale zanim lepiej uda∏o si´ poznaç mechanizm, dzi´ki
któremu mózg trwale rejestruje jedne wydarzenia, a inne od-
rzuca, naukowcy musieli rozwiàzaç pewien paradoks.
U podstaw zarówno pami´ci d∏ugotrwa∏ej, jak i krótko-
trwa∏ej le˝y specyfika po∏àczeƒ mi´dzy neuronami. W miejscu
styku neuronów, czyli synapsie, wypustka przekazujàca sy-
gna∏, zwana aksonem, napotyka dendryt – jeden z wielu roz-
ga∏´zionych wyrostków sàsiadujàcych neuronów [ilustracja
na stronie 69]. Podczas tworzenia pami´ci krótkotrwa∏ej po-
budzenie synapsy tymczasowo uwra˝liwia jà na nast´pnà sty-
mulacj´, wzmacniajàc jej dzia∏anie. Po pewnym czasie sytu-
acja wraca do stanu wyjÊciowego. Natomiast w przypadku
pami´ci d∏ugotrwa∏ej synapsy wzmacniane sà na sta∏e. Ju˝
od lat szeÊçdziesiàtych naukowcy wiedzà, ˝e do utrzymania
takich zmian w mózgu niezb´dna jest aktywnoÊç genów w
jàdrze komórkowym, czyli ich ekspresja prowadzàca do pro-
dukcji bia∏ek.
Badacze pami´ci zastanawiali si´, w jaki sposób ekspresja
genów ukrytych g∏´boko w jàdrze komórkowym mo˝e zawia-
dywaç odleg∏ymi synapsami. Skàd gen wie, kiedy trwale
wzmocniç synaps´, a kiedy pozwoliç nieistotnym wydarze-
niom na znikni´cie bez Êladu? I skàd bia∏ka kodowane przez
te geny wiedzà, którà z tysi´cy synaps nale˝y wzmocniç? Po-
MARZEC 2005 ÂWIAT NAUKI
67
Komórkowe
Êcie˝ki pami´ci
Niektóre wydarzenia zapami´tujemy
na zawsze, inne zaÊ nam ulatujà.
Przyczynà tego sà te same procesy,
które kszta∏tujà mózg podczas rozwoju
R. Douglas Fields

dobne problemy dotyczà p∏odowego etapu rozwoju mózgu,
kiedy zapada decyzja, które po∏àczenia synaptyczne zostanà
zachowane, a które nie. Podczas badaƒ nad tym zagadnie-
niem mój zespó∏ doszed∏ do intrygujàcego rozwiàzania jednej
z tajemnic pami´ci.
Pami´ç genetyczna
ROL
¢ GENÓW
w przekszta∏ceniu pami´ci krótkotrwa∏ej w d∏u-
gotrwa∏à odkryli ju˝ pionierzy biologii molekularnej. Do-
Êwiadczenia, podczas których zwierz´ta wykonywa∏y proste
zadania, pokaza∏y, ˝e do prawid∏owego uczenia si´ niezb´d-
na jest synteza nowych bia∏ek w mózgu i to w czasie kilku
lub kilkunastu minut po treningu, w przeciwnym razie pa-
mi´ç zanika [patrz: Bernard W. Agranoff „Memory and Pro-
tein Synthesis”; Scientific American, czerwiec 1967].
Aby powsta∏o bia∏ko, musi zajÊç transkrypcja odcinka DNA
znajdujàcego si´ wewnàtrz jàdra komórkowego, czyli przepi-
sanie kodu DNA na przenoÊnà postaç materia∏u genetyczne-
go zwanà informacyjnym RNA (mRNA). Informacyjny RNA
w´druje dalej z jàdra na zewnàtrz, do cytoplazmy, gdzie ma-
szyneria komórkowa dokonuje translacji, czyli zdekodowa-
nia instrukcji i biosyntezy nowego bia∏ka. Badacze odkryli,
˝e zarówno zablokowanie transkrypcji DNA na mRNA, jak i
translacji mRNA na bia∏ko uniemo˝liwia powstawanie d∏u-
gotrwa∏ych Êladów pami´ciowych, podczas gdy pami´ç krót-
kotrwa∏a pozostaje nienaruszona.
Poniewa˝ jeden neuron mo˝e tworzyç dziesiàtki tysi´cy sy-
naps i raczej niemo˝liwe jest, aby dla ka˝dej z nich istnia∏
oddzielny, przyporzàdkowany gen, neurobiolodzy musieli wy-
jaÊniç, jak jàdro komórkowe kontroluje wzmocnienie poszcze-
gólnych synaps. Wysnuli hipotez´, ˝e odpowiednio silnie po-
budzona synapsa wytwarza jakàÊ czàsteczk´ sygna∏owà.
Podczas gdy tymczasowo wzmocniona synapsa utrzymuje
Êlad pami´ciowy, czàsteczka przemieszcza si´ do jàdra ko-
mórkowego neuronu i tam mo˝e aktywowaç geny niezb´d-
ne do produkcji bia∏ek utrwalajàcych wzmocnienie transmi-
sji synaptycznej. Nierozwiàzany pozosta∏ jednak problem, w
jaki sposób nowo wyprodukowane bia∏ko znajduje drog´ po-
wrotnà z cia∏a komórki do tej jednej spoÊród tysi´cy synaps,
która wys∏a∏a sygna∏.
W po∏owie lat dziewi´çdziesiàtych przebieg zdarzeƒ sta∏
si´ jaÊniejszy [ilustracja na stronie 70]. Badania wykaza∏y, ˝e
u gatunków zwierzàt tak filogenetycznie odleg∏ych od siebie
jak muchy i myszy kluczowà rol´ w przekszta∏caniu pami´ci
krótkotrwa∏ej w d∏ugotrwa∏à odgrywa ten sam czynnik
transkrypcyjny o nazwie CREB. Czynniki transkrypcyjne sà
bia∏kami wià˝àcymi si´ ze specyficznymi sekwencjami DNA,
a ich funkcjà jest w∏àczenie bàdê wy∏àczenie transkrypcji
okreÊlonych genów. Pobudzenie bia∏ka CREB w neuronie
prowadzi do ekspresji genów kodujàcych tajemnicze bia∏ka
wzmacniajàce transmisj´ synaptycznà. Efektem tego jest prze-
kszta∏cenie pami´ci krótkotrwa∏ej w d∏ugotrwa∏à.
W roku 1997 Uwe Frey z Leibniz-Institut für Neurobiolo-
gie w Magdeburgu oraz Richard G. M. Morris z University
of Edinburgh wykazali, ˝e „bia∏ka pami´ci”, jakiekolwiek
by one by∏y, wcale nie muszà szukaç drogi do tej jednej sy-
napsy. Docierajà po prostu do ka˝dego zakàtka komórki, ale
mogà oddzia∏ywaç wy∏àcznie na synapsy, które zosta∏y wczeÊ-
niej tymczasowo wzmocnione, i tam powodujà wzmocnienie
trwa∏e. Nadal jednak jedno istotne pytanie pozostawa∏o bez
odpowiedzi: czym jest czàsteczka sygna∏owa przekazujàca z
synapsy do jàdra komórki informacje o koniecznoÊci aktywa-
cji bia∏ka CREB w celu utrwalenia Êladu pami´ciowego?
W tym czasie ja i moi wspó∏pracownicy zajmowaliÊmy si´
podobnymi problemami, choç z nieco innej perspektywy.
W moim laboratorium w National Institute of Child Health and
Human Development dociekaliÊmy, jak powstajà po∏àczenia
neuronalne w mózgu rozwijajàcego si´ p∏odu. Tak wi´c gdy
badacze pami´ci zastanawiali si´, w jaki sposób nasze do-
Êwiadczenia ˝yciowe mog∏yby wp∏ywaç na geny, a te z kolei
modyfikowaç si∏´ po∏àczeƒ synaptycznych, my próbowaliÊmy
odpowiedzieç na pytanie, w jaki sposób geny determinujà mi-
liony po∏àczeƒ w kszta∏tujàcym si´ mózgu.
Wraz z innymi neurobiologami zajmujàcymi si´ rozwojem
mózgu sàdziliÊmy, ˝e doznania mogà odgrywaç rol´ w ulep-
szaniu planu po∏àczeƒ neuronalnych. W mózgu najpierw
powstaje zgrubna sieç neuronów, której kszta∏t okreÊlajà ge-
ny. W miar´ dojrzewania narzàdu po∏àczenia sà tworzone i te-
stowane; te bardziej efektywne zostajà zachowane, a mniej
efektywne wyeliminowane. Skàd jednak mózg wie, które po-
∏àczenia nale˝y zachowaç?
Budowanie mózgu
JU
˚ W ROKU
1949
psycholog Donald Hebb zaproponowa∏ pro-
stà regu∏´ wyjaÊniajàcà, w jaki sposób doznania wp∏ywajà
na sieç neuronów. Na podstawie s∏ynnych doÊwiadczeƒ Paw-
∏owa z psami wysnu∏ hipotez´, ˝e si∏a po∏àczenia mi´dzy
dwoma neuronami, które sà aktywne w tym samym momen-
cie, wzrasta. Przyk∏adowo neuron aktywowany dêwi´kiem
dzwonka oraz neuron aktywowany jednoczesnym podaniem
pokarmu powinny wzmocniç si∏´ po∏àczeƒ pomi´dzy sobà. W
ten sposób komórki tworzà obwód uczàcy si´, ˝e te dwa wy-
darzenia – dzwonek oraz podanie pokarmu – sà ze sobà po-
wiàzane.
Nie ka˝dy impuls docierajàcy do neuronu jest wystarczajà-
co silny, aby wywo∏aç jego reakcj´ w postaci przekazania sy-
gna∏u. Komórka nerwowa tak jak mikroprocesor nieustan-
nie odbiera i integruje tysiàce impulsów wejÊciowych
docierajàcych do niej z dendrytów, jednak ma tylko jedno
wyjÊcie – akson. Tak wi´c mo˝e odpowiedzieç tylko na zasa-
dzie wszystko albo nic: albo przekazaç informacj´ do nast´p-
nych neuronów za pomocà impulsu, albo nie.
Impulsy docierajàce do neuronów przez pobudzonà synap-
s´ powodujà lokalne zmniejszenie ró˝nicy potencja∏ów po
68
ÂWIAT NAUKI MARZEC 2005
n
Poszczególne neurony w jakiÊ sposób rozpoznajà, które
wspomnienia nale˝y zachowaç w postaci sta∏ych po∏àczeƒ
pomi´dzy jednà komórkà a drugà, a którym mo˝na pozwoliç
zaniknàç. Podobnie rozwijajàcy si´ mózg w pewien sposób
wybiera, które sieci komórek nale˝y pozostawiç,
a które wyeliminowaç.
n
W obydwu procesach niezb´dne jest przes∏anie z dalekich
kraƒców neuronów sygna∏ów elektrochemicznych, których
zadaniem jest w∏àczenie genów w jàdrze komórkowym, a tak˝e
przes∏anie przez w∏àczone geny odpowiedzi zwrotnej.
n
Jak w przypadku wielu decyzji ˝yciowych neurony decydujà si´
na utrwalenie po∏àczeƒ tylko wtedy, gdy sà pewne, ˝e jest to
rzeczywiÊcie wa˝ne.
Przeglàd /
Okablowanie
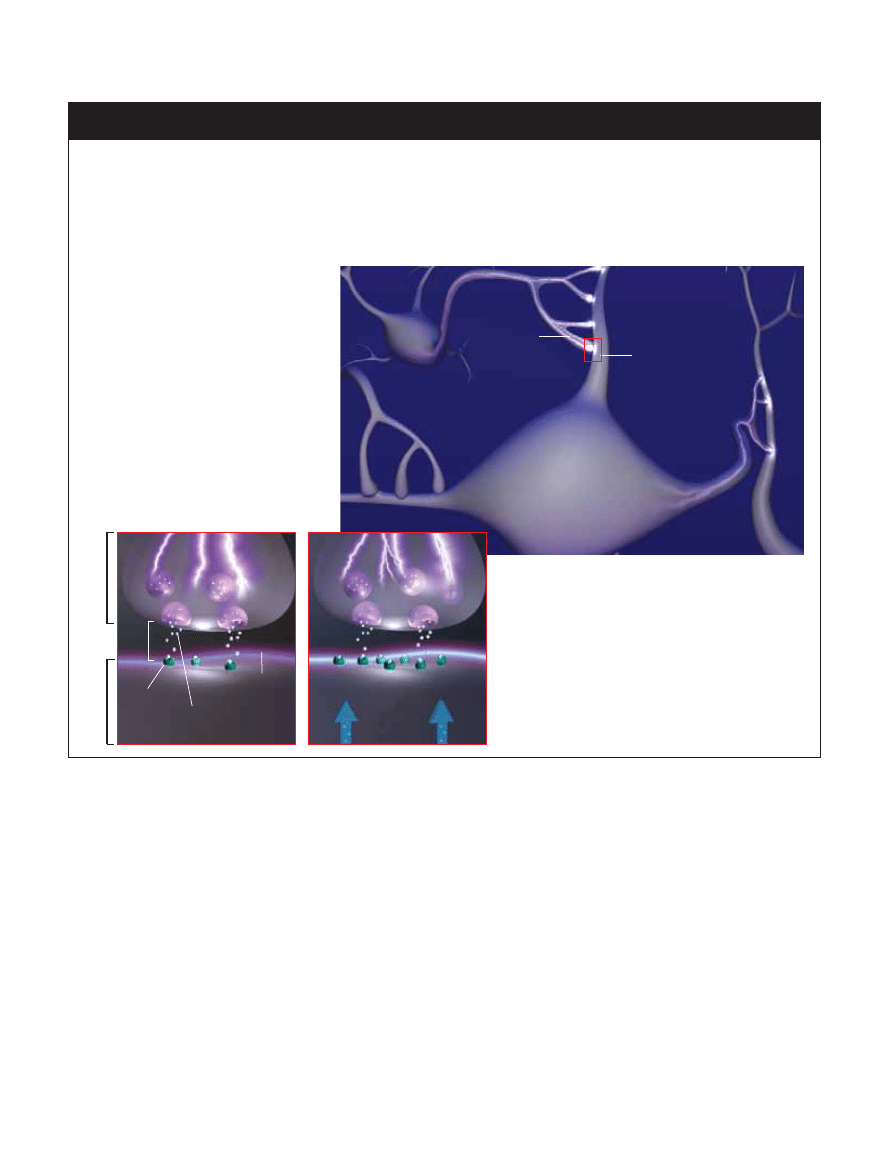
MARZEC 2005 ÂWIAT NAUKI
69
ALFRED T
. KAMAJIAN
JAK POWSTAJE PAMI¢å
Pami´ç powstaje, gdy komórki nerwowe tworzàce sieç wzajemnych po∏àczeƒ zwi´kszajà wra˝liwoÊç tych po∏àczeƒ zwanych
synapsami. W przypadku pami´ci krótkotrwa∏ej wzmocnienie transmisji synaptycznej utrzymuje si´ od kilkunastu minut do kil-
ku godzin. Synapsy bioràce udzia∏ w rozwoju pami´ci d∏ugotrwa∏ej wzmacniane sà na sta∏e. W powstawaniu pami´ci bierze
udzia∏ sam zapami´tywany sygna∏. Swojà podró˝ od neuronu do neuronu zaczyna on w chwili, gdy pierwsza komórka (komór-
ka presynaptyczna) wygeneruje impuls elektryczny zwany potencja∏em czynnoÊciowym (poni˝ej), który w´druje wzd∏u˝ wypust-
ki zwanej aksonem do zakoƒczenia synaptycznego.
PRZEKAèNICTWO SYNAPTYCZNE
Impuls docierajàcy do zakoƒczenia aksonu ko-
mórki presynaptycznej (wstawka poni˝ej) spra-
wia, ˝e ze specjalnych p´cherzyków do szczeliny
synaptycznej uwalniane sà substancje chemicz-
ne zwane neuroprzekaênikami. Neuroprzekaêni-
ki z kolei wià˝à si´ z receptorami znajdujàcymi si´
na cz´Êci postsynaptycznej, powodujàc lokalnà
depolaryzacj´ dendrytów drugiej komórki.
WZMACNIANIE SYNAPSY
Silne pobudzenie synapsy z wysokà cz´stoÊcià powoduje
tymczasowe uwra˝liwienie jej na nast´pny impuls, który
dzi´ki czemu mo˝e wyzwoliç wi´kszà depolaryzacj´ lokalnà.
To czasowe wzmocnienie wra˝liwoÊci synapsy jest podsta-
wà pami´ci krótkotrwa∏ej. Nie wiemy dok∏adnie, jak powsta-
je wzmocnienie d∏ugotrwa∏e, ale stwierdzono, ˝e w komór-
ce postsynaptycznej musi zajÊç biosynteza nowych bia∏ek
(z lewej). Bia∏ka te mogà wzmacniaç transmisj´ synaptycz-
nà przez zwi´kszenie iloÊci receptorów lub przebudow´ cz´-
Êci postsynaptycznej. Niewykluczone, ˝e mogà one te˝ wp∏y-
waç na funkcje cz´Êci presynaptycznej.
Neuron
K
omórka
postsynaptyczna
K
omórka
presynaptyczna
Neuron
Akson
Akson
Szczelina
synaptyczna
Dendryt
Dendryt
Receptor
Depolaryzacja
Neuroprzekaêniki
Bia∏ka
wzmacniajàce
transmisj´
synaptycznà
Synapsa
Potencja∏
czynnoÊciowy
obu stronach b∏ony komórkowej dendrytu, czyli depolaryza-
cj´. Kiedy synapsa pobudzana jest krótkimi seriami impul-
sów o wysokiej cz´stoÊci, wytwarza si´ stan chwilowego
wzmocnienia przekaênictwa charakterystyczny dla powstawa-
nia pami´ci krótkotrwa∏ej. Jednak krótka aktywnoÊç poje-
dynczej synapsy zwykle nie mo˝e zmusiç komórki do prze-
kazania dalej impulsu zwanego potencja∏em czynnoÊciowym
(AP – action potential). Dopiero wspólny wysi∏ek wielu rów-
noczeÊnie pobudzanych synaps zmienia potencja∏ b∏ony ko-
mórkowej w takim stopniu, ˝e powstaje potencja∏ czynno-
Êciowy i wiadomoÊç przekazywana jest do nast´pnego neuronu
w sieci.
Hebb doszed∏ do wniosku, ˝e synapsa, której aktywnoÊç
nie jest zsynchronizowana z innymi, b´dzie si´ negatywnie
odró˝niaç tak jak muzyk, który nie mo˝e zgraç si´ z orkie-
strà, i powinna zostaç usuni´ta. Natomiast synapsy, które
wspó∏grajà ze sobà, pobudzajàc komórk´ w sposób wystar-
czajàcy do wygenerowania potencja∏u czynnoÊciowego, b´-
dà wzmacniane. Tak wi´c mózg zmienia organizacj´ po∏à-
czeƒ, kierujàc si´ aktywnoÊcià rozwijajàcej si´ sieci neuro-
nów i doskonalàc jej wyjÊciowy kszta∏t.
Enzymy i inne bia∏ka, które w myÊl teorii Hebba wzmacnia-
jà lub os∏abiajà si∏´ po∏àczeƒ synaptycznych, muszà powstawaç
wskutek ekspresji konkretnych, aktywowanych genów. Nasz
zespó∏ podjà∏ si´ zadania zdefiniowania sygna∏ów w∏àczajà-
cych te geny.
Poniewa˝ informacja w uk∏adzie nerwowym kodowana jest
przez zmiany aktywnoÊci neuronów, za∏o˝y∏em, ˝e ekspresja
niektórych genów musi byç sterowana wzorem pobudzenia ko-
mórek nerwowych. ˚eby przetestowaç t´ hipotez´, wraz z Ko-
uichim Itohem odbywajàcym sta˝ podoktorski w moim labo-
ratorium izolowaliÊmy komórki nerwowe z p∏odów myszy,
hodowaliÊmy je na szalkach i stymulowaliÊmy pràdem tak, by
wzbudziç potencja∏ czynnoÊciowy. ZastosowaliÊmy ró˝ne wzo-
ry stymulacji i nast´pnie mierzyliÊmy poziomy mRNA genów, któ-
re pe∏nià wa˝ne funkcje w kszta∏towaniu sieci neuronów lub
adaptacji do Êrodowiska. Nasze przypuszczenia okaza∏y si´
s∏uszne. MogliÊmy w∏àczaç lub wy∏àczaç ekspresj´ konkretnych
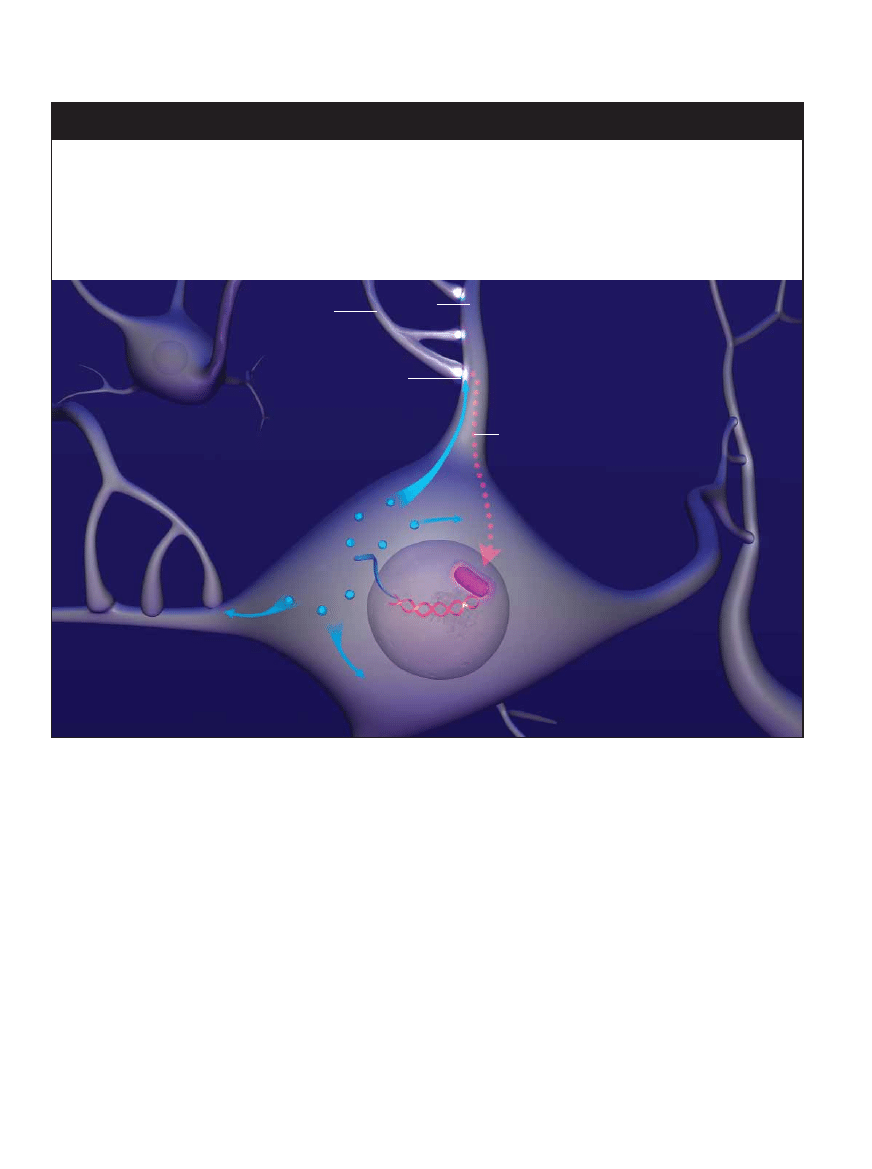
70
ÂWIAT NAUKI MARZEC 2005
ALFRED T
. KAMAJIAN
JAK GENY SCALAJÑ PAMI¢å
W latach szeÊçdziesiàtych odkryto, ˝e aktywacja genów prowadzàca do produkcji bia∏ek jest niezb´dna do wytworzenia pa-
mi´ci d∏ugotrwa∏ej. Od razu zrodzi∏o si´ pytanie, skàd gen mo˝e wiedzieç, kiedy ma uruchomiç produkcj´ bia∏ek majàcych zdol-
noÊç wzmocnienia synapsy (co le˝y u podstaw przekszta∏cenia pami´ci krótkotrwa∏ej w d∏ugotrwa∏à), a kiedy ma pozostaç
nieaktywny (pozwalajàc na zatarcie pami´ci krótkotrwa∏ej)? Czy istnieje jakaÊ dotàd niepoznana czàsteczka sygna∏owa, w´dru-
jàca od synapsy do jàdra komórkowego, która mówi komórce, ˝e ta ma uruchomiç produkcj´ bia∏ek niezb´dnych do wzmoc-
nienia transmisji synaptycznej? I jak bia∏ka majà rozpoznaç wÊród tysi´cy synaps t´ jednà, którà nale˝y wzmocniç? W po∏o-
wie lat dziewi´çdziesiàtych kilka pomys∏owych doÊwiadczeƒ rozjaÊni∏o te kwestie.
genów, ustawiajàc odpowiednià cz´stotliwoÊç na naszym sty-
mulatorze elektrofizjologicznym, tak jak dostrajamy radio do
okreÊlonej stacji nadawczej, aby s∏uchaç wybranego programu.
Szyfr czasowy
KIEDY ZAOBSERWOWALIÂMY
, ˝e ekspresja genów w komórkach
nerwowych mo˝e zale˝eç od wzorca aktywnoÊci neuronu,
postanowiliÊmy zbadaç powa˝niejszy problem. W jaki spo-
sób wzór elektrycznych depolaryzacji powierzchni b∏ony ko-
mórkowej wp∏ywa na geny po∏o˝one g∏´boko w jàdrze neuro-
nu? Aby odpowiedzieç na to pytanie, musieliÊmy przyjrzeç
si´ uwa˝nie cytoplazmie, przez którà wiedzie szlak sygna∏u z
powierzchni komórki do jàdra.
Okaza∏o si´, ˝e nie znajduje si´ tam pojedyncza Êcie˝ka,
lecz plàtanina krzy˝ujàcych si´ ze sobà szlaków biochemicz-
nych, które rozprowadzajà informacj´ z b∏ony komórki po
ca∏ym jej wn´trzu. W jakiÊ sposób sygna∏ elektryczny o ró˝-
nych cz´stoÊciach potrafi∏ w tym cytoplazmatycznym gàsz-
czu odnaleêç prawid∏owà drog´ z b∏ony do jàdra neuronu.
ChcieliÊmy zrozumieç, jak to si´ dzieje.
G∏ównà Êcie˝kà, po której informacja o stanie elektrycznym
b∏ony dociera do systemu szlaków biochemicznych w cy-
toplazmie, sà b∏onowe kana∏y wapniowe zale˝ne od napi´-
cia. St´˝enie jonów wapnia wewnàtrz komórki utrzymuje
si´ na bardzo niskim poziomie – jest ich tam 20 000 razy
mniej ni˝ na zewnàtrz. Kiedy depolaryzacja b∏ony osiàga
krytycznà wartoÊç, wyzwolony zostaje potencja∏ czynnoÊcio-
wy. Jest to jednoczeÊnie sygna∏ do otwarcia na krótkà chwi-
l´ kana∏ów wapniowych, które do komórki wpuszczajà wiel-
ki strumieƒ jonów wapnia. W ten sposób impuls elektryczny
za ka˝dym razem przekszta∏cany jest w wiadomoÊç zrozu-
mia∏à dla maszynerii biochemicznej znajdujàcej si´ we wn´-
trzu neuronu.
Dalsze wydarzenia przypominajà przekazywanie pa∏eczki
w biegu sztafetowym. Nap∏ywajàcy do cytoplazmy wapƒ akty-
wuje nast´pny element szlaku przekaênictwa sygna∏owego –
enzymy zwane kinazami bia∏kowymi. Dobudowujà one gru-
p´ fosforanowà do innych enzymów (cz´sto tak˝e kinaz), w
ten sposób w∏àczajàc je lub wy∏àczajàc. Te zaÊ oddzia∏ujà na
kolejne bia∏ka. Koƒcowym etapem tego wielostopniowego pro-
1
Silna bàdê wielokrotnie
powtarzana stymulacja
tymczasowo wzmacnia synaps´
i w ten sposób informuje jàdro,
˝e nale˝y stworzyç trwa∏y
Êlad pami´ciowy
2
Aby nastàpi∏o
trwa∏e wzmocnienie
synapsy, musi dojÊç
do pobudzenia aktywnoÊci
bia∏ka CREB
5
Bia∏ka mogà zadzia∏aç tylko
na synapsy b´dàce w stanie
tymczasowego wzmocnienia
wywo∏anego wyjÊciowà
stymulacjà
3
Wewnàtrz jàdra
komórkowego CREB
aktywuje wybrane geny,
powodujàc ich transkrypcj´
na mRNA, który nast´pnie
w´druje do cytoplazmy
4
Maszyneria komórkowa
rozpoczyna translacj´ mRNA,
produkujàc bia∏ko wzmacniajàce
transmisj´ synaptycznà, które
swobodnie rozprasza si´
po ca∏ej komórce
Akson
Dendryt
Synapsa
CREB
mRNA
Gen
Jàdro
komórkowe
Bia∏ka
wzmacniajàce
transmisj´
synaptycznà
Hipotetyczne czàsteczki przekazujàce
sygna∏ od synapsy do jàdra komórkowego
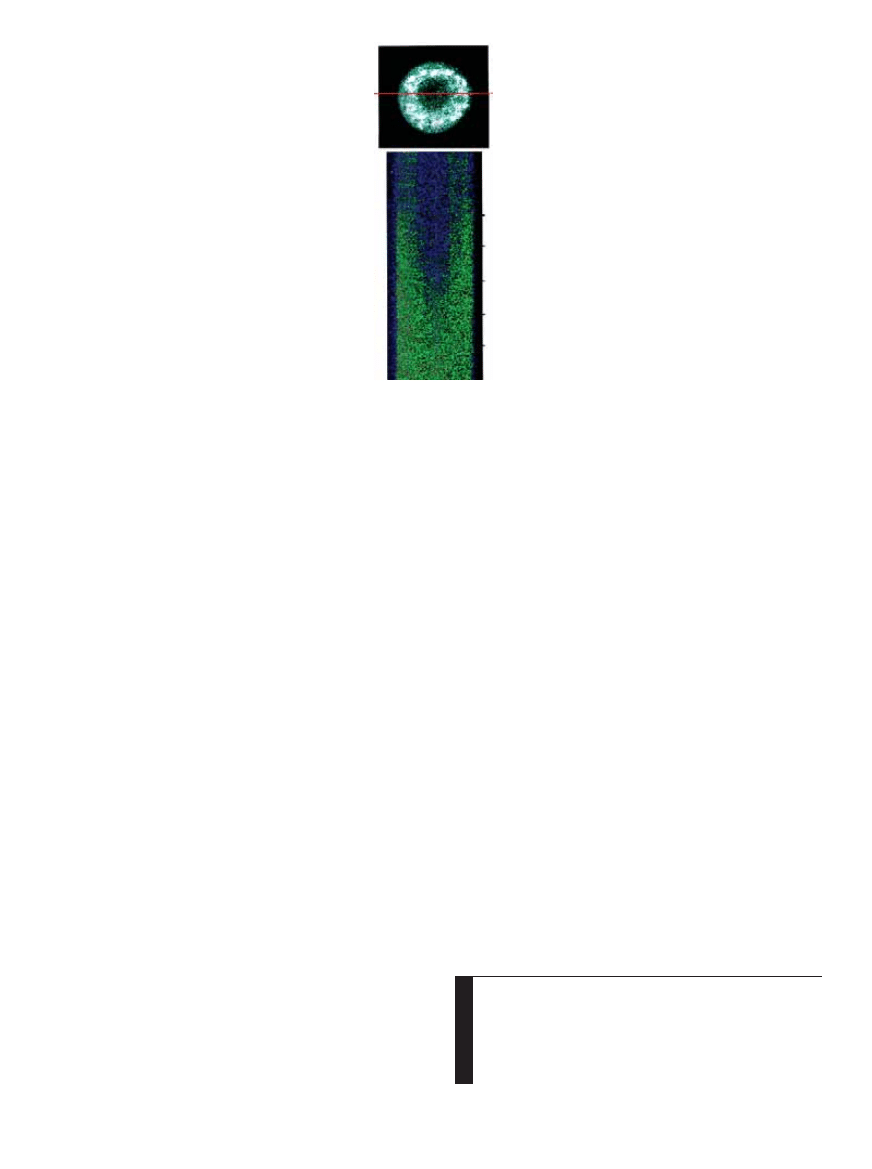
cesu jest aktywacja czynników transkrypcyjnych.
Na przyk∏ad czynnoÊç bia∏ka CREB jest pobu-
dzana przez fosforylacj´ kinazami zale˝nymi od
wapnia i wygaszana przez enzymy, które usuwa-
jà grup´ fosforanowà. Jednak w komórce znajdu-
jà si´ setki innych czynników transkrypcyjnych
oraz kinaz bia∏kowych. ChcieliÊmy si´ dowie-
dzieç, w jaki sposób wzór aktywnoÊci elektrycz-
nej neuronu, regulujàc nap∏yw wapnia, pobudza
konkretne kinazy, które z kolei docierajà do w∏a-
Êciwych czynników transkrypcyjnych i za ich po-
Êrednictwem kontrolujà odpowiednie geny.
˚eby przeÊledziç, jak ró˝ne wzory aktywnoÊci
elektrycznej neuronów przek∏adane sà na che-
miczny sygna∏ zmian st´˝enia wewnàtrzkomór-
kowego wapnia, wype∏niliÊmy neurony barwni-
kami, które pod wp∏ywem wapnia Êwiecà na
zielono. Mo˝na by∏o przypuszczaç, ˝e poszcze-
gólne geny odpowiadaç b´dà na ró˝ne st´˝enie wapnia we-
wnàtrz neuronów. ZaobserwowaliÊmy jednak coÊ o wiele bar-
dziej interesujàcego. Amplituda wzrostu st´˝enia wapnia w
neuronie mia∏a dla regulacji aktywnoÊci genów mniejsze zna-
czenie ni˝ przebieg zmian st´˝eƒ w czasie, b´dàcy odbiciem ak-
tywnoÊci elektrycznej komórki.
Feleke Eshete podczas sta˝u podoktorskiego w moim la-
boratorium identyfikowa∏ Êcie˝ki sygna∏owe wiodàce od na-
p∏ywu wapnia do komórki przez aktywacj´ enzymów a˝ do
pobudzenia czynników transkrypcyjnych. Stopniowo zaczy-
naliÊmy uÊwiadamiaç sobie, jak ró˝ne wzorce przekazywa-
nych przez neuron impulsów mogà pobudzaç poszczególne
drogi przesy∏ania sygna∏u. Najwa˝niejszym czynnikiem oka-
za∏ si´ czas.
OdkryliÊmy, ˝e nie da si´ przedstawiç szlaku wiodàcego z
b∏ony komórki do DNA jako prostego ∏aƒcucha reakcji bioche-
micznych. Na ka˝dym kroku, poczynajàc od nap∏ywu wap-
nia do neuronu, droga przekazu informacji rozga∏´zia si´ w
sieç silnie powiàzanych ze sobà Êcie˝ek. Ka˝da z nich ma in-
ne ograniczenia szybkoÊci reakcji determinujàce zdolnoÊç
odpowiedzi na sygna∏y o ró˝nych wzorcach. Te w∏aÊciwoÊci
decydujà, jakà drogà sygna∏ o okreÊlonej cz´stoÊci przekaza-
ny zostanie do jàdra.
Niektóre Êcie˝ki sygna∏owe szybko odpowiadajà na sygna∏
i natychmiast wracajà do stanu wyjÊciowego. Dzi´ki temu
nadajà si´ do przekazywania sygna∏u o wysokiej cz´stoÊci,
ale nie mogà podtrzymaç sygnalizacji podczas d∏u˝szych
okresów braku aktywnoÊci elektrycznej, rozdzielajàcych serie
potencja∏ów czynnoÊciowych. Z kolei inne, powolne Êcie˝ki
nie nadà˝ajà za szybkimi seriami impulsów, ale te˝ nie potra-
fià natychmiast wygaszaç aktywnoÊci, co gwarantuje pod-
trzymanie sygna∏u w okresach przerw we wzmo˝onej czynno-
Êci elektrycznej neuronu. Geny kontrolowane przez te Êcie˝ki
b´dà zatem pobudzane w sytuacjach, gdy sygna∏ dociera do
komórki wielokrotnie, lecz niezbyt cz´sto – w∏aÊnie tak jak
podczas zapami´tywania powtarzanej informacji.
Inaczej mówiàc, zaobserwowaliÊmy, ˝e sygna∏y o ró˝nych
wzorcach czasowych aktywujà przystosowane do dane-
go wzorca szlaki przekaênictwa, co umo˝liwia zró˝nicowane
w∏àczanie czynników transkrypcyjnych i genów. Na przyk∏ad
nasze badania pokaza∏y, ˝e aktywacja bia∏ka CREB nast´pu-
je bardzo szybko po stymulacji neuronu i utrzymuje si´ doÊç
d∏ugo po zaprzestaniu pobudzania komórki.
CREB mo˝e wi´c dzia∏aç podczas trwajàcej na-
wet ponad 30 min przerwy pomi´dzy powtarza-
nymi seriami stymulacji. Przerwy tej d∏ugoÊci sà
charakterystyczne dla nauki lub treningu.
To nas zaciekawi∏o. Skoro CREB gra tak wielkà
rol´ w procesie zapami´tywania, to czy Êcie˝ka
przekazywania sygna∏u, którà badaliÊmy, aby zro-
zumieç procesy rozwoju mózgu, nie ma równie˝ zwiàzku z
pami´cià? PostanowiliÊmy to sprawdziç.
Pami´ç w probówce
STRUKTUR
¢ MÓZGU
, którà usuni´to pacjentowi HM – hipokamp
– mo˝na wyizolowaç z mózgu myszy, pociàç na plasterki i
utrzymywaç przy ˝yciu w odpowiednim roztworze, a za po-
mocà mikroelektrod pod∏àczonych do wzmacniaczy elektro-
nicznych rejestrowaç aktywnoÊç elektrycznà pojedynczych
synaps. Podajàc seri´ wielu impulsów elektrycznych w krót-
kim czasie (tzw. stymulacja t´˝cowa lub wysokocz´stotliwo-
Êciowa), które silnie aktywujà synaps´, mo˝emy jà wzmoc-
niç. Wtedy zastosowanie pojedynczego impulsu elektrycznego
wywo∏a oko∏o dwukrotnie wi´kszy potencja∏ elektryczny ni˝
przed stymulacjà z wysokà cz´stoÊcià.
Taki wzrost efektywnoÊci synapsy, nazywany d∏ugotrwa-
∏ym wzmocnieniem synaptycznym (LTP – long term poten-
tiation), pomimo swojej nazwy, mo˝e trwaç doÊç krótko. Je-
˝eli po stymulacji t´˝cowej podawaç b´dziemy pojedyncze
testowe impulsy w sta∏ych odst´pach, zaobserwujemy, ˝e de-
polaryzacyjna odpowiedê synapsy powoli si´ zmniejsza, po
kilku godzinach powracajàc do poziomu wyjÊciowego. Tym-
czasowe wzmocnienie synapsy, znane jako wczesne LTP, jest
komórkowym modelem pami´ci krótkotrwa∏ej.
Co ciekawe, jeÊli t´ samà stymulacj´ t´˝cowà powtórzymy
kilka razy (w naszym doÊwiadczeniu trzykrotnie), to wzmac-
niamy synaps´ na sta∏e, otrzymujàc póêne LTP. Poszczególne
stymulacje nie mogà jednak nast´powaç bezpoÊrednio jedna
po drugiej – pomi´dzy ka˝dà serià impulsów musi byç odpo-
wiednia przerwa (w naszym przypadku 10 min). Kiedy do so-
li fizjologicznej, w której trzymamy skrawki hipokampa, do-
damy zwiàzki chemiczne blokujàce powstawanie mRNA lub
bia∏ek, to nawet po powtarzanej kilkakrotnie stymulacji t´˝-
cowej wielkoÊç wywo∏ywanych potencja∏ów wróci do stanu
MARZEC 2005 ÂWIAT NAUKI
71
WED¸
UG FELEKE ESHETE I R
. DOUGL
ASA FIELDSA;
JOURNAL OF NEUROSCIENCE
, TOM 21, NR 17, 1 IX 2001. ©
SOCIETY FOR NEUROSCIENCE
R. DOUGLAS FIELDS jest kierownikiem Sekcji Rozwoju i Plastycz-
noÊci Uk∏adu Nerwowego w National Institute for Child Health and
Human Development i wyk∏adowcà Neurosciences and Cognitive
Science Program w University of Maryland. W swoim poprzednim
artykule dla Scientific American [„Niedoceniane komórki mózgu”;
Âwiat Nauki, maj 2004] opisa∏ rol´ gleju w procesach myÊlenia i
uczenia si´.
O
AUTORZE
JONY WAPNIA widoczne na przekroju neuronu dzi´ki
barwnikowi wra˝liwemu na ich obecnoÊç (z lewej ciem-
ne jàdro komórkowe). Autor u˝ywa∏ mikroskopu konfo-
kalnego do wizualizacji rzutów dokomórkowego nap∏ywu
wapnia, wywo∏anych potencja∏ami czynnoÊciowymi. Obra-
zy przekrojów poprzecznych uzyskiwane co 2 ms zosta-
∏y z∏o˝one (poni˝ej z lewej), ukazujàc zmiany st´˝enia we-
wnàtrzkomórkowego wapnia w czasie (zielony i czerwony).
0 ms
250
500
750
1000
wyjÊciowego po dwóch, trzech godzinach. Tak jak w ca∏ym
organizmie komórkowy model pami´ci krótkotrwa∏ej nie za-
le˝y od jàdra, podczas gdy d∏ugotrwa∏ej zale˝y.
Frey i Morris u˝yli tej techniki, aby pokazaç, ˝e bia∏ka od-
powiedzialne za póêne LTP mogà dzia∏aç na ka˝dà tymczaso-
wo wzmocnionà synaps´. Najpierw stymulowali jednà synap-
s´ pojedynczà serià impulsów, aby uzyskaç wczesne LTP, które
normalnie trwa kilka godzin. Potem stymulowali drugà sy-
naps´ tej samej komórki w taki sposób, ˝eby powsta∏o na niej
póêne LTP, stosujàc trzy serie impulsów oddzielone od siebie
dziesi´ciominutowymi przerwami. W efekcie otrzymali póê-
ne LTP na obu synapsach. Silna stymulacja by∏a dla jàdra sy-
gna∏em do produkcji bia∏ek pami´ciowych, które odnalaz∏y
drog´ do ka˝dej synapsy gotowej do ich u˝ycia.
Zacz´liÊmy si´ zastanawiaç, czy rzeczywiÊcie do powsta-
nia pami´ci d∏ugotrwa∏ej jest potrzebna czàsteczka sygna∏o-
wa wys∏ana z synapsy do jàdra. OpieraliÊmy si´ na w∏asnych
badaniach, z których wynika∏o, ˝e ró˝ne przebiegi impulsów
mogà aktywowaç poszczególne geny, uwzgl´dnialiÊmy tak˝e
teori´ Hebba, wed∏ug której aktywnoÊç neuronu jest niezb´d-
na do okreÊlenia, jakie po∏àczenia zostanà wzmocnione. Mo-
˝e zatem silna aktywacja jednej synapsy albo synchroniczna
aktywacja wielu synaps wzbudza potencja∏ czynnoÊciowy w´-
drujàcy po aksonie, co wywo∏uje nap∏yw wapnia przez
kana∏y zale˝ne od napi´cia, po∏o˝one w ciele komórki, w po-
bli˝u jàdra? I to w∏aÊnie ten sygna∏ dociera opisanymi wcze-
Êniej Êcie˝kami do bia∏ka CREB?
Razem ze sta˝ystkà Serenà Dudek postanowiliÊmy doÊwiad-
czalnie sprawdziç naszà hipotez´, umieszczajàc skrawki mózgu
w roztworze substancji blokujàcej funkcjonowanie synapsy.
Aby wygenerowaç potencja∏ czynnoÊciowy, bezpoÊrednio sty-
mulowaliÊmy elektrycznie cia∏o komórki lub akson. W ten spo-
sób bez udzia∏u synaps otrzymywaliÊmy potencja∏ czynnoÊcio-
wy w neuronie. JeÊli do wytworzenia póênego LTP (naszego
komórkowego modelu pami´ci d∏ugotrwa∏ej) niezb´dna by∏a-
by czàsteczka sygna∏owa w´drujàca od stymulowanej synapsy
do jàdra, wy∏àczenie synapsy powinno zablokowaç powstanie
trwa∏ego wzmocnienia transmisji. JeÊli natomiast to potencja-
∏y czynnoÊciowe wywo∏ujà sygna∏y docierajàce do jàdra, za-
blokowanie synapsy nie powinno mieç wp∏ywu na aktywnoÊç
genów w jàdrze. Gdy zbadaliÊmy, czy w skrawkach mózgu za-
sz∏a aktywacja czynnika transkrypcyjnego CREB, stwierdzili-
Êmy, ˝e w niewielkim obszarze, w którym stymulowaliÊmy po-
tencja∏y czynnoÊciowe przy ca∏kowicie wy∏àczonych synapsach,
czàsteczki bia∏ka CREB mia∏y do∏àczone grupy fosforanowe, co
znaczy, ˝e zosta∏y prze∏àczone w stan aktywny.
72
ÂWIAT NAUKI MARZEC 2005
ALFRED T
. KAMAJIAN
SKÑD JÑDRO WIE, KIEDY WZMOCNIå SYNAPS¢
Dzi´ki eksperymentom autora okaza∏o si´, ˝e hipotetyczna czàsteczka niosàca informacj´ z synapsy do jàdra nie jest potrzeb-
na. Silne pobudzenie, b´dàce efektem powtarzanych stymulacji jednej synapsy lub równoczesnà aktywacjà wielu synaps jed-
nej komórki, depolaryzuje b∏on´ i generuje potencja∏ czynnoÊciowy w neuronie. Potencja∏ ten otwiera kana∏y wapniowe, a jony
wapnia za poÊrednictwem kaskady enzymów aktywujà czynnik transkrypcyjny CREB. Pobudza on ekspresj´ genów kodujàcych
bia∏ka wzmacniajàce synaps´. W efekcie jàdro komórkowe decyduje o trwa∏ym wzmocnieniu synapsy na podstawie „nas∏uchu”
sygna∏ów na wyjÊciu z komórki – potencja∏ów czynnoÊciowych. W ten sposób powstaje pami´ç d∏ugotrwa∏a.
1
Silna stymulacja
depolaryzuje
b∏on´ komórkowà
6
Bia∏ka dyfundujà
w komórce, wp∏ywajàc
tylko na te synapsy, które
zosta∏y tymczasowo
wzmocnione
2
Depolaryzacja
generuje potencja∏
czynnoÊciowy
3
Nast´puje otwarcie
kana∏ów wapniowych
zale˝nych od napi´cia
4
Jony wapnia
aktywujà enzymy
pobudzajàce CREB
5
CREB aktywuje
geny bia∏ek
wzmacniajàcych synapsy
Depolaryzacja
b∏ony
Bia∏ka
wzmacniajàce
synaps´
Kana∏
wapniowy
Potencja∏
czynnoÊciowy
Enzymy
Nap∏yw
wapnia
CREB
Gen
Jàdro
komórkowe
Nast´pnie sprawdziliÊmy aktywnoÊç genu zif268, który jest
zaanga˝owany w powstawanie LTP i pami´ci. Okaza∏o si´, ˝e
równie˝ on aktywowany jest przez pobudzenie komórek hipo-
kampa bez udzia∏u synaps. PowtórzyliÊmy ten sam ekspery-
ment z innà substancjà, blokujàcà dzia∏anie kana∏ów wapnio-
wych zale˝nych od napi´cia, które w naszej opinii by∏y
odpowiedzialne za wygenerowanie sygna∏u przekazywanego
z b∏ony do jàdra. W tym przypadku zarówno CREB, zif268, jak
i inne bia∏ko zwiàzane z póênym LTP, kinaza bia∏kowa MAPK,
nie zosta∏y pobudzone przez potencja∏y czynnoÊciowe.
Te wyniki wyraênie pokazujà, ˝e wcale nie jest potrzebne
bia∏ko wysy∏ane z konkretnej pobudzonej synapsy do jàdra.
Okaza∏o si´, ˝e – podobnie jak w naszych badaniach nad roz-
wojem mózgu – dla jàdra informacjà wystarczajàcà do zapoczàt-
kowania ekspresji konkretnego genu jest sygna∏ generowany
nap∏ywem wapnia po depolaryzacji b∏ony komórkowej przez po-
tencja∏ czynnoÊciowy. Wydaje si´, ˝e jest to du˝o prostsze roz-
wiàzanie ni˝ wysy∏anie do jàdra „prywatnej” wiadomoÊci przez
ka˝dà z tysi´cy synaps aktywowanych na neuronie. Urucho-
mienie ekspresji genów pami´ci zale˝y od tego, czy neuron jest
aktywny, czy przekazuje dalej informacje w postaci potencja-
∏ów czynnoÊciowych przebiegajàcych od dendrytów przez cia-
∏o komórki do aksonu.
Molekularne memento
W PROCESACH ZAPAMI
¢TYWANIA
byç mo˝e biorà udzia∏ nieod-
kryte dotàd czàsteczki niosàce sygna∏ od synapsy do jàdra,
jednak nasze eksperymenty dowiod∏y, ˝e nie sà one niezb´dne.
Jak okreÊli∏ to Hebb w swojej teorii uczenia si´, aktywnoÊç
neuronu b´dàca wynikiem równoczesnego pobudzenia wszyst-
kich wejÊç synaptycznych danej komórki jest niezb´dna
do konsolidacji pami´ci.
Taka koncepcja daje nam doÊç atrakcyjnà komórkowà analo-
gi´ naszych codziennych doÊwiadczeƒ z pami´cià. Tak jak Le-
onard z filmu Memento lub jakikolwiek Êwiadek przest´pstwa
nikt zawczasu nie wie, które wydarzenia powinny na sta∏e tra-
fiç do naszych wspomnieƒ. Dostrajanie si∏y po∏àczeƒ synap-
tycznych idealnie nadaje si´ do przechowywania informacji na
krótki okres, co jest niezb´dne do normalnego funkcjonowania
w teraêniejszoÊci. Je˝eli jednak jakieÊ wydarzenia majà dla nas
du˝e znaczenie lub wielokrotnie czegoÊ doÊwiadczamy, wzmo-
˝ona, powtarzana aktywacja synaps daje znaç sieci neuronów,
˝e to wydarzenie nale˝y trwale zapami´taç. W∏àczane sà odpo-
wiednie geny, a kodowane przez nie bia∏ka docierajà do synaps
oczekujàcych na decyzj´: zapami´taç lub zapomnieç. Tam od-
ciskajà pi´tno tak trwa∏e jak tatua˝ na skórze Leonarda.
n
KOMENTARZ
R. Douglas Fields opisuje wyniki badaƒ swojego zespo∏u i in-
nych uczonych, dotyczàce mechanizmów powstawania d∏u-
gotrwa∏ych Êladów pami´ciowych w mózgu. Nie kwestionu-
jàc sposobu rozumowania autora, warto zwróciç uwag´, ˝e
przekonanie o roli bia∏ka CREB w tych procesach opiera si´
na kontrowersyjnych danych. Najwa˝niejsze z nich pochodzà
z 1994 roku, kiedy to wspó∏pracujàce ze sobà zespo∏y badaczy
pod kierunkiem A. J. Silvy, wówczas z Cold Spring Harbor
Laboratory w Nowym Jorku, i G. Schuetza z Niemieckiego Cen-
trum Badaƒ nad Rakiem w Heidelbergu opublikowa∏y w Cell
opisy doÊwiadczeƒ, w których u myszy z uszkodzeniem genu
kodujàcego CREB stwierdzi∏y zaburzenia pami´ci d∏ugotrwa-
∏ej i LTP. Wydawa∏o si´ to bardzo mocnym dowodem doÊwiad-
czalnym na krytycznà rol´ CREB w mechanizmie zapami´ty-
wania. Defekt CREB dotyczy∏ jednak wszystkich komórek
organizmu, a nie tylko mózgu. Nie mo˝na zatem by∏o wyklu-
czyç, ˝e obserwowane zaburzenia mia∏y inne pod∏o˝e, nawet
zupe∏nie niezwiàzane z mózgiem. Na naszà zdolnoÊç zapami´-
tywania majà przecie˝ wp∏yw bardzo ró˝ne zjawiska, w tym
m.in. ogólne samopoczucie – nawet banalne przezi´bienie upo-
Êledza pami´ç!
Dlatego te˝ grupa Schuetza stworzy∏a kolejne szczepy myszy,
u których defekt CREB (a tak˝e jego homologu CREM) przeja-
wia∏ si´ wy∏àcznie w mózgu – w jednym przypadku w tzw. neu-
ronach pobudzajàcych (to o nich pisze Fields) przodomózgo-
wia, w tym i hipokampa, a w drugim przypadku we wszystkich
komórkach nerwowych i glejowych. Wyniki badaƒ tych myszy
opublikowano w 2003 roku w Journal of Neuroscience – oka-
za∏o si´, ˝e ani jedne, ani drugie nie majà zaburzeƒ LTP ani pa-
mi´ci zale˝nej od hipokampa!
Udzia∏ bia∏ka CREB w zapami´tywaniu jest zatem wàtpli-
wy. Czy oznacza to, ˝e wàtpliwy jest sam poglàd o roli zmian ak-
tywnoÊci genów w procesach pami´ciowych? Zdecydowanie
nie! Ma on bowiem du˝o solidniejsze i bogatsze podstawy. Nikt
nie kwestionuje wyników bardzo licznych badaƒ z u˝yciem in-
hibitorów biosyntezy bia∏ka i RNA, które blokujà tworzenie
trwa∏ych Êladów pami´ciowych. Ponadto zjawiskom uczenia
si´ i LTP rzeczywiÊcie towarzyszà zmiany aktywnoÊci genów w
mózgu. DowiedliÊmy tego pierwsi (równolegle z K. W. Ano-
chinem z Moskwy), prowadzàc w 1988 roku wspólne doÊwiadcze-
nia z zespo∏em H. Matthiesa z Magdeburga i stwiedzajàc wzrost
ekspresji c-fos podczas uczenia si´. Co ciekawe, bia∏ko c-Fos
kodowane przez ten gen jest tak˝e (jak CREB) regulatorem trans-
krypcji! Zgromadzono bardzo wiele danych doÊwiadczalnych,
pokazujàcych zwi´kszonà aktywnoÊç c-Fos podczas zapami´-
tywania. Na pewno tak wielu informacji nie mamy w przypad-
ku ˝adnego innego bia∏ka. Gdy zak∏ócano funkcjonowanie tego
bia∏ka w ca∏ym organizmie lub selektywnie w mózgu, za ka˝dym
razem obserwowano tak˝e zaburzenia uczenia si´.
Trzeba jednak dodaç, ˝e i te wyniki sà krytykowane z podob-
nego powodu co badania nad CREB: bia∏ko c-Fos odgrywa
ogromnà rol´ w ró˝nych komórkach (nie tylko nerwowych) i
jego uszkodzenie ma znaczàce skutki uboczne. WàtpliwoÊci roz-
wia∏oby selektywne zahamowanie czynnoÊci c-Fos w ÊciÊle okre-
Êlonym miejscu i czasie w mózgu. Niestety, tego nauka jeszcze
nie potrafi.
prof. dr hab. LESZEK KACZMAREK
Instytut Biologii DoÊwiadczalnej im. M. Nenckiego PAN
http://neurogene.nencki.gov.pl
MARZEC 2005 ÂWIAT NAUKI
73
Ekspresja genów w procesach d∏ugotrwa∏ych zmian czynnoÊci oÊrodko-
wego uk∏adu nerwowego. Leszek Kaczmarek; Mózg, wyd. Libramed,
s. 20-32, 1997.
Memory Systems of the Brain: A Brief History and Current Perspe-
ctive. Larry R. Squire; Neurobiology of Learning and Memory, tom 82,
s. 171-177, XI/2004.
Somatic Action Potentials Are Sufficient for Late-Phase LTP-Related Cell
Signaling. Serena M. Dudek i R. Douglas Fields; Proceedings of the
National Academy of Sciences USA, tom 99, nr 6, s. 3962-3967;
19 III 2002.
Synaptic Tagging and Long-Term Potentiation. Uwe Frey i Richard G. M.
Morris; Nature, tom 385, s. 533-536; 6 II 1997.
Regulated Expression of the Neural Cell Adhesion Molecule L1 by Spe-
cific Patterns of Neural Impulses. Kouichi Itoh, B. Stevens, M. Schach-
ner i R. D. Fields; Science, tom 270, s. 1369-1372, 24 XI 1995.
JEÂLI CHCESZ WIEDZIEå WI¢CEJ
Wyszukiwarka
Podobne podstrony:
2005 Bid 25351 Nieznany (2)
chemia maj 2005 id 112453 Nieznany
2005 Sprzezone rozwiazywanieid Nieznany (2)
prawo bud 2005 id 339439 Nieznany
45 Nature 438 197200 2005 id 38 Nieznany (2)
auto cad 2005 nauka VWUQLMDEZTB Nieznany (2)
1 Pobudliwosc komorkiid 9556 Nieznany (2)
2005 Vid 25393 Nieznany (2)
ADWOKACI pyt testowe 2005 id 52 Nieznany (2)
2005 rokid 25389 Nieznany (2)
Aspiracje nr 3 2005 [pdf] Aspir Nieznany (2)
1 Pojecia wstepne komorka, tka Nieznany (2)
03 2005 niezwykle materialyid 4 Nieznany (2)
2005 GPWid 25363 Nieznany (2)
1 cw BLONY KOMORKOWEid 9083 Nieznany (2)
alfik 2005 3 id 56893 Nieznany (2)
2005 Bid 25351 Nieznany (2)
więcej podobnych podstron