
1
WYKŁAD 1
Kwiaty i zalążki
pąk – rozwija się z merystemów wierzchołkowych na szczycie pędu w tzw.: pachwinach liści,
następuje przekształcenie wegetatywnych pąków w merystem kwiatostanowy, zamiast
łodygi i liści rozwija się kwiat i jego poszczególne części
Schemat rozwoju kwiatu
merystem apikalny przekształca się, rozszerza się i spłaszcza
pęd przestaje się wydłużać
powstają zaczątki kielicha (działek kielicha), następnie płatków korony, pręcików,
owocolistków. Proces ten zachodzi od podstawy do wierzchołka (rozwój akropetalny)
pąki powstają pod koniec okresu wegetacji w postaci tzw.: pąków zimowych, z grubą okrywą,
otoczone powłoką podobną do śluzu
Kwiat
organ generatywny wszystkich roślin okrytozalążkowych
u podstawy kwiatu występuje szypułka, dno kwiatowe, na których osadzony jest okwiat
okwiat złożony zawiera płatki korony i działki kielicha (czyli zmodyfikowane liście)
część generatywna zawiera słupek z makrosporofilami, czyli owocolistkami (zalążnia, szyjka,
znamię), pręciki zbudowane z nitki i główki. Wszystkie te elementy osadzone są na dnie
kwiatowym
w kwiecie wyróżniamy zatem część wegetatywną i płodną
Ewolucja kwiatu
ze skróconego pędu ze zredukowanymi międzywęźlami i zmodyfikowanymi liśćmi
tendencje ewolucyjne
o skracanie osi kwiatowej i przejście ze spiralnego do okółkowego ułożenia w kwiecie
o zmniejszenie liczby okółków
o słupek dolny zamiast górnego
o zalążnia na wypukłym dnie kwiatowym
o symetria grzbiecista zamiast promienistej
o zalążnia zrośnięta z dnem kwiatowym i znajdujące się w niej części kwiatu
Rodzaje symetrii kwiatu
dwuboczna
promienista
grzbiecista
asymetria (np. u storczyka)
Rodzaje kwiatów
obupłciowe (lilia)
rozdzielnopłciowe
o jednopienne (kwiaty męskie i żeńskie na tej samej roślinie – kukurydza)
o dwupienne (kwiaty męskie i żeńskie na innej roślinie – topola)
Kwiaty rozdzielnopłciowe
kwiaty tylko z pręcikowiem – tylko męskie
kwiaty tylko z słupkowiem – tylko żeńskie
kwiaty płonne – pierwsze obwodowe kwiaty Hortensji

2
Budowa okwiatu
zbudowany z kielicha i korony
o kielich – z liści przykwiatowych, najczęściej zielony
o korona – z liści przykwiatowych lub pręcików, najczęściej barwna
typy koron
o korona wolnopłatkowa – jabłoń
o korona zrosłopłatkowa – petunia
o korona z przykoronkiem – żonkil
funkcje okwiatu
o chroni słupki i pręciki
o wabi zapylacze
może być pojedynczy lub podwójny
Okwiat
pojedynczy – jednoliścienne
podwójny – dwuliścienne = kielich K + działki kielicha + korona C + płatki korony
brak okwiatu – kwiaty bezokwiatowe
Pręciki
budowa: nitka + główka
główka zbudowana z dwóch pylników
w każdym z pylników są dwa woreczki pyłkowe (łącznie 4)
pylniki złączone są za pomocą łącznika w którym zatopiona jest wiązka przewodząca
nie wszystkie pręciki są płodne – tzw.: prątniczki u powojnika, nie zawierają pyłku
Pyłek
dojrzałe ziarno pyłku stanowi gametofit męski (i tylko on!)
ziarna pyłku niektórych roślin zawierają worki powietrzne, np.: sosna
Słupek
powstaje w wyniku zrośnięcia owocolistków
typy słupków
o jednoowocolistkowe – jeden szew – u grochu
o wieloowocolistkowe – tworzą tyle jednolistkowych słupków ile owocolistków lub
słupek wielokrotny z wielu owocolistków
Ewolucja słupka
w ewolucji liść z którego powstał słupek początkowo miał klasyczny kształt; zalążki położone
były obwodowo, tendencją było jego zwijanie i tworzenie tuby, gdzie zalążki położone były w
miejscu połączenia, czyli szwu
położenie zalążni względem dna kwiatowego
o słupek wystaje poza dno kwiatowe – słupek górny (hypogeiczny) – kwiat dolny
o słupek trochę wystaje poza dno kwiatowe – słupek górny (peigeiczny) – kwiat
okołozalążkowy
o słupek zapadnięty w dno kwiatowe – słupek dolny (epigeiczny) – kwiat górny
Budowa słupka
zawiera łożysko (placenta) z której wyrastają zalążki
często stigma (znamię) połączona jest z zalążnią szyjką, ale u traw jest inaczej

3
Funkcje słupka
znamię – przyjmuje ziarno pyłku, które kiełkuje
szyjka – odżywianie rosnącego gametofitu żeńskiego
zalążnia – rozwój zalążków (ovaria)
Zalążek
żeński organ rozmnażania z którego rozwija się gametofit żeński w postaci woreczka
zalążkowego
centrum zalążka tworzy nucellus, w którym występuje komórka archesporialna z której
powstają makrospory
nucellus osłonięty jest integumentem wewnętrznym i zewnętrznym, pomiędzy nimi tworzy
się tzw.: mikropyle
wyróżniamy dwa bieguny: chalazalny i mikropylarny
Powstawanie zalążka
miejscem powstania makrospory jest megasporangium
zalążki powstają w postaci wzgórków merystematycznych czyli primordiów
Etapy powstania zalążka
młody zalążek powstaje z komórek merystematycznych
z komórek pośrodku tworzy się nucellus
wielkość zalążka zależy od liczby warstw dzielącej się placenty (cecha gatunkowa)
ośrodek stanowi centralną część
podział protodermy prowadzi do powstania osłonki (integumentu) u podstawy wzgórka
nie u wszystkich roślin występują dwa integumenty
o u większości roślin – 2
o Santalales – 0
o Solanaceae i Acanthaceae – 1
osłonka zbudowana jest z miękiszu, który przekształca się w tkankę mechaniczną –
sklerenchymę łupiny nasiennej, czasem część z nich odżywia zarodek
w zalążku wyróżnia się bieguny – mikropylarny i chalazalny, gdzie zalążek łączy się ze
sznureczkiem
sznureczek – funicullus, tworzy się z tkanki miękiszowej w której zatopiona jest tkanka
przewodząca, a u Compositae, Convolvulaceae występują też w pozostałych elementach
endothelium – tapetum integumentalne – powstaje w osłonce, skórce przylegającej do
ośrodka (nucellusa), okryta grubą kutikulą, może zawierać więcej niż 2C DNA. Odżywia
woreczek.
o kutikula jest wewnętrzna
o u niektórych roślin stanowi bazową warstwę okrywy nasiennej (Ricinus communis)
o komórki wierzchołka endothelium mogą tworzyć czapeczkę – kalotkę, która może
wyrastać na zewnątrz – w wyniku podziałów periklinalnych
operculum – u Eukaliptusa
o wewnętrzne – w kolorze zalążka
o zewnętrzne – czerwono – żółte
u niektórych roślin w części chalazalnej występują komórki ulegające lignifikacji – jest to
hipostaza, która zapobiega rozrostowi woreczka, ułatwia też transport substancji z wiązki
przewodzącej do woreczka zalążkowego
podium (postament) – w kształcie miseczki o którą opiera się woreczek
epistaza – w ośrodku nielicznych gatunków zapobiega wysychaniu zapłodnionego zalążka,
infekcji patogenem
4
u niektórych roślin peryferyjne warstwy komórek ośrodka pozostają z nasieniu jako tkanki
spichrzowe w postaci obielma (perysperma)
u części gatunków tworzy się obturator – ze sznureczka lub z placenty, tworzy wachlarz i jest
drogą po której rośnie łagiewka (u lilii)
Typy zalążków
ortotropowy – mikropyle naprzeciw sznureczka – pokrzywowate, rdestowate
hemitropowy – okienko prostopadle do sznureczka – jaskrowate
kampylotropowy – okienko zagięte – strączkowe
amfitropowy – podkowiaste zagięcie zalążka i woreczka – żabieńcowate
anatropowy – odwrócony, okienko przy sznureczku – liliowate
Układ zalążków w słupku
placenta brzeżna (ścienna)
placenta osiowa (środkowa)
placenta swobodna (kolumnowa)
Ustawienia megasporocytu
pod epidermą – subepidermalny
pod warstwą subepidermalną
głęboko w ośrodku
Powstawanie makrospory
z komórki macierzystej, 2n chromosomów i 4C DNA
następuje pierwszy podział mejotyczny – redukcyjny 1n, 2C
powstają dzięki temu dwie duże komórki – diada
następuje drugi podział redukcyjny – zachowawczy, gdzie zmniejsza się zawartość DNA z 2C
na 1C
2n
1n
1n
4C
2C
1C
z tych 4 makrospor 3 są nieaktywne, jedna rozwija się dalej
zaczyna powstawać gametofit
Powstawanie gametofitu
po kariokinezie nie ma cytokinezy
pojawia się 8 jąder, pomiędzy którymi jest duża wakuola, część pozostaje na biegunie
chalazalnym, część na mikropylarnym
dwa z jąder wywędrowują do środka, może tam powstawać komórka zlana z dwóch jąder
biegunowych, która jest diploidalna
powstają dwie synergidy i jedna komórka jajowa
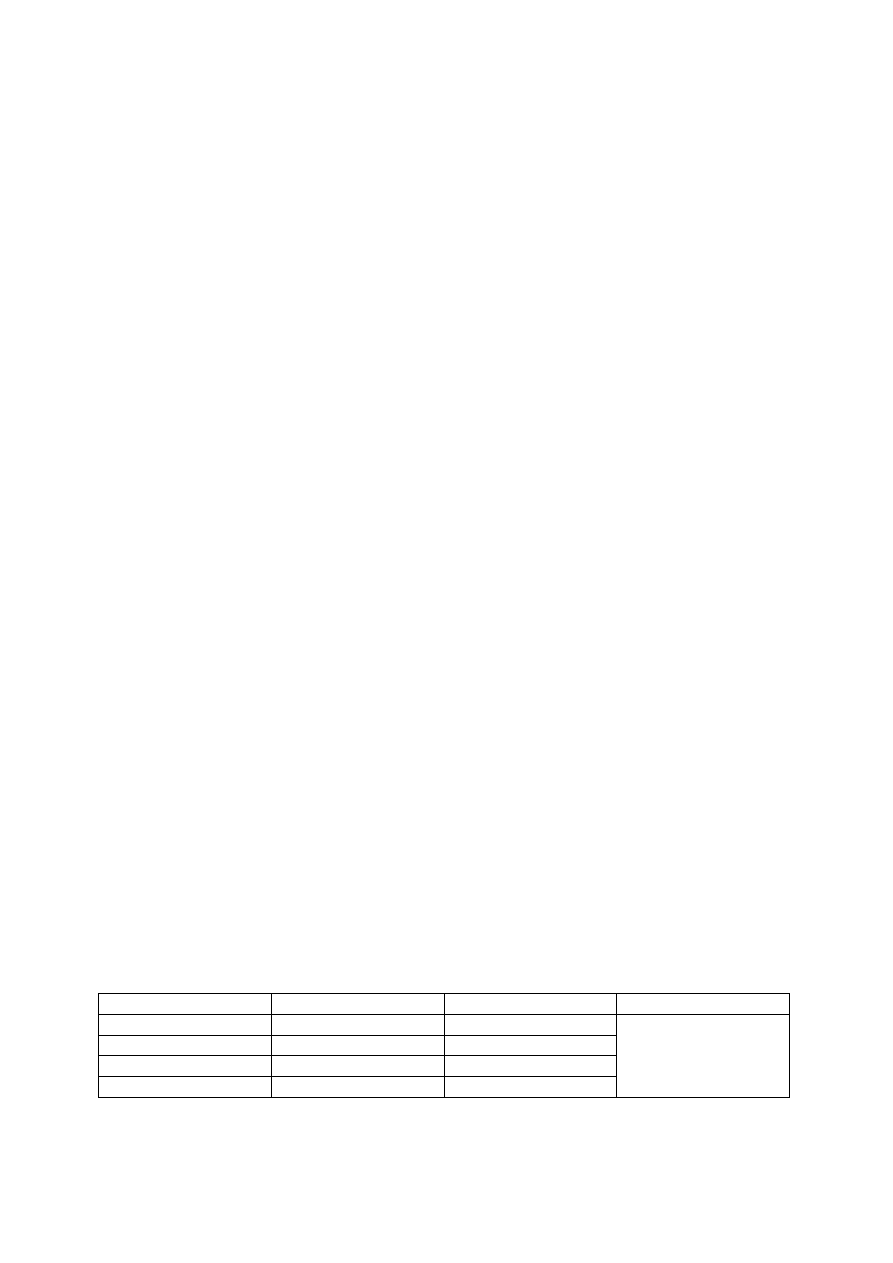
Ilość
Typ
Chromosomy
Łącznie
1
Komórka jajowa
1n
7 komórek
1
Komórka centralna
2n
2
Synergidy
1n
3
Antypody
1n

5
Typ woreczka zalążkowego a ilość jąder
monosporowy – z 1 komórki powstają 4 komórki jednojądrowe
bisporowy – z 1 komórki powstają 2 komórki dwujądrowe
tetrasporowy – nie ma cytokinezy, zawsze są 4 jądra
Synergidy
tworzą aparat włókienkowy czyli filiformowy, który ułatwia przepływ substancji odżywczej
odpowiada za funkcję troficzną i oddziaływanie na łagiewkę
musi mieć plastydy ze skrobią i mitochondria
w pobliżu bieguna chalazalnego jedna lub kilka wakuoli
Tamaryszek
tworzy jeden lub kilka typów woreczków
zależy to od temperatury
mogą być aż cztery typy.: np. chryzantemum, adoxa lub dhrusa
Podwójne zapłodnienie
tylko u okrytozalążkowych
występują dwa jądra komórek plemnikowych
jedno zapładnia komórką jajową, drugie komórkę centralną, tworzy się zarodek (2n) i bielmo
(3n)
z osłony zalążka tworzy się łupina nasienna
zalążek przekształca się zatem w nasiono

6
WYKŁAD 2
Budowa pręcika
pylnik
filament
Kwiaty
obupłciowe – hermafrodytyczne – 90% gatunków
o rośliny jednopienne – 1% gatunków, modrzew, leszczyna, brzoza, dąb, orzech włoski,
sosna, tuja, jodła, ogórek, kukurydza, melon, figowiec
o rośliny dwupienne – 3% gatunków, cis, klon jesionolistny, topola, wierzba, szparag,
szpinak, chmiel, konopie, kiwi, daktylowiec, pistacja
jednopłciowe – dwupienne + jednopienne
Płeć roślin i kwiatów jako umowny wskaźnik
kwiaty pręcikowe uważamy za męskie
kwiaty słupkowe uważamy za żeńskie
same kwiaty nie mają płci, gdyż są częścią sporofitu
męskość i żeńskość zależne jest od wytwarzania gamet
tylko gametofity posiadają płeć – woreczki zalążkowe i ziarna pyłku
u roślin dwupiennych sporofit żeński oznaczamy XX, a męski XY
u jednopiennych i hermafrodytów nie ma zróżnicowanych chromosomów płci, czyli nie ma X i
Y, a geny związane z płcią znajdują się na innych chromosomach
o płci decydują zatem wytwarzane hormony płciowe
Przemiana pokoleń
wyróżniamy dwie fazy
o faza wzrostu, organogenezy (wegetatywna, diploidalna) – po zapłodnieniu powstaje
sporofit, kończy się w momencie powstania tetrad (mejocytów)
o faza rozmnażania się (generatywna, haploidalna) – w kwiatach, rozpoczyna się
powstaniem zarodników – mikro i makrosporocytów, kończy się wytworzeniem
gamet w gametofitach
Budowa pręcika
zbudowany z nitki i główki, każda główka zbudowana jest z dwóch pylników, a każdy pylnik
zawiera dwa woreczki, inaczej mikrosporangium
funkcją jest produkcja ziaren pyłku, jego wytwarzanie i uwalnianie
w przekroju poprzecznym widoczna jest epiderma, endotecjum, dwie warstwy komórek
pośrednich, tapetum i pokład ziaren pyłku
rozwój pręcika – powstaje z wierzchołka kwiatowego w młodym pędzie z merystemów
o komórki merystematyczne otoczone są epidermą
o następuje różnicowanie w czterech narożach główki – tkanka pramacierzysta
mikrospor
o dookoła nich różnicuje się tapetum, wokół tapetum okrywa
tworzenie się woreczka pyłkowego
o endotecjum – otwieranie pylników i specyficzna ściana komórkowa
o komórki pośrednie – zawierają skrobię i tłuszcze
o tapetum
o komórki pyłkowe
komórka macierzysta mikrospor – różnicuje się w mikrosporocyty

7
intensywny wzrost komór pyłkowych – rozdział mikrosporocytów zawieszonych w płynie
odżywczym pochodzącym z tapetum, które dostarcza polisacharydy, białka, lipidy
powstają mikrospory (w młodych, nierozwiniętych pąkach)
zanika tapetum
mikrospory rozwijają się w ziarna pyłku
Synchronizacja rozwoju
mikrospory połączone są z tapetum plasmodesmami
połączenia zanikają po rozpoczęciu mejozy
zanik ściany celulozowo-pektynowej, utworzenie ściany kalozowej mikrosporocytów
mejoza rozpoczyna się w sezonie poprzedzającym kwitnienie u roślin drzewiastych i
zatrzymuje się na etapie mejozy w fazie profazy tetrady lub mikrospor
na wiosnę proces ulega dokończeniu (np.: u wiśni)
Synchronizacja mikrosporocytów
sąsiadujące komórki połączone są kanałami cytomiktycznymi
synchronizację warunkują dyfundujące substancje
Tapetum
pojedyncza warstwa komórek otaczająca komorę pylnikową
rozwija się równolegle z mikrosporocytami
dochodzi do utraty plazmodesmy z przyległymi tkankami (mejocyty i warstwa pośrednia)
cytoplazma zawiera skrobię i orbikule (otoczone sporopolleniną, tzw. ciałka Ubisha)
jądra stają się poliploidalne, komórki stają się poliploidalne
dostarcza kalazę, która trawi ścianę kalozową (ta zastępowana jest sporopolleninową)
rozpuszcza własną ścianę komórkową i staje się protoplastem
komórki rozpadając się (PCD) przed otwarciem okwiatu, kaloza zostaje wymieniona na
egzynę (sporopollenina)
wydziela płyn do komory sporangium
brak tapetum powoduje problemy w rozwoju komórek pyłku
są dwa rodzaje tapetum:
o ameboidalne (plazmodium)
o wydzielnicze (sekrecyjne, parietalne)
Rodzaje tapetum
wydzielnicze – u dwuliściennych – na obrzeżu komory pylnika
ameboidalne – u jednoliściennych – komórczaki
Endotecjum
pojedyncza warstwa komórek tuż pod epidermą główki pręcika
ściany styczne i promieniste – z zwiększoną zawartością celulozy, ściany styczne zewnętrzne
są znacznie cieńsze, warunkuje to otwarcie komory i wysyp
warstwa zewnętrzna wysycha, warunkuje to otwarcie komory, a miejsce otwarcia to stomia
Różnicowanie tkanki mikrosporialnej
komórki pramacierzyste mikrospor (tkanka archesporialna) różnicuje się
powstają komórki macierzyste mikrospor (mikrosporocyty)
następuje mikrogametogeneza w wyniku której powstają mikrospory
mikrospory dojrzewają dając ziarna pyłku
8
tkanka archesporialna mikrosporocyty mikrospory ziarna pyłku
Mikrosporogeneza
od etapu mikrosporocytu do etapu mikrospory
Stadium
Ilość chromosomów
Ilość DNA
Mikrosporocyt
2n
2c
Mikrosporocyt
2n
4c
Diada
1n
2c
Tetrada
1n
1c
Mikrospory
1n
1c
Dojrzała mikrospora
1n
1c
Główne układy tetrad
u większości dwuliściennych – tetraedr
u jednoliściennych – jedna, liniowa płaszczyzna
Podziały
komórka merystematyczna (mitoza) komórka pramacierzysta mikrospor (mitoza)
komórka macierzysta mikrospor (mejoza I) diada (mejoza II) tetrada (różnicowanie)
ziarno pyłku (mikrospora)
Mikrogametogeneza
od mikrospory do ziarna pyłku
w tetradzie znajdują się 4 haploidalne mikrospory (1c, 1n(
DNA ulega replikacji (2c, 1n)
jądro przesuwa się w stronę ściany, powstaje mikrospora spolaryzowana
podział mitotyczny, powstaje niedojrzałe ziarno pyłku, komórka generatywna i wegetatywna
(ta druga w fazie G₀) 2c, 1n
komórka generatywna dzieli się na dwie komórki
ziarno pyłku składa się zatem z komórki wegetatywne i dwóch komórek generatywnych
każda komórka ziarna pyłku jest haploidalna
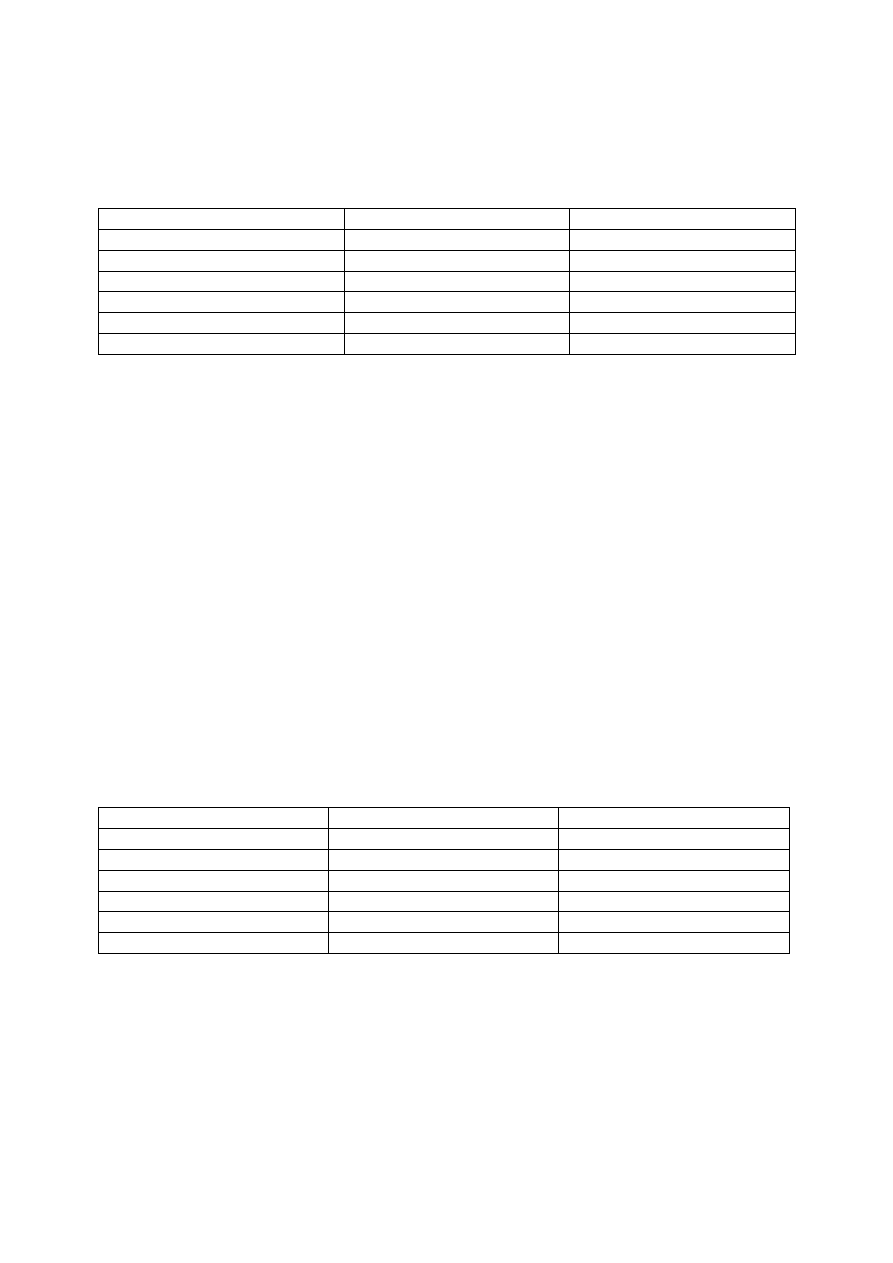
Etap
Ilość DNA
Ilość chromosomów
Mikrosporocyt
4c
2n
Diada
2c
1n
Tetrada
1c
1n
4 mikrospory
1c
1n
4 ziarna pyłku
1c
1n
8 komórek plemnikowych
1c
1n
Podział jądra generatywnego
dwujądrowe ziarno pyłku może dzielić się
o jeszcze w pylniku
o w łagiewce pyłkowej

9
Dojrzałe ziarno pyłku
duża komórka generatywna
okryta sporodermą, która jest dwuwarstwowa (egzyna z kutyny oraz intyna z celulozy i
pektyn)
wewnątrz komórki wegetatywnej dwie wakuole i jądro
dwa jądra generatywne
Sporoderma - budowa
Tectum
Segzyna
Egzyna
Kolumelle
Negzyna
Intyna
egzyna – tworzy się jako primegzyna, z włókien celulozowych i glikoprotein, syntetyzowana
przez mikrospory
negzyna – sporopollenina, syntetyzowana w mikrosporach, osadza się pod primegzyną,
powstaje z orbikul
intyna – odkłada się na końcu, zbudowana z celulozy, pektyn, białek
wolna przestrzeń między kolumellami to przestrzeń dla kiełkującej łagiewki
sporopolleniny mają różną barwę
Ściany komórek szeregu mikrosporogenezy
komórki pramacierzyste mikrospor
- celuloza + pektyna
mikrosporocyt
- kaloza
tetrada
- egzyna + sporopolleniny
mikrospory
- sporopollenina
ziarno pyłku
- sporopollenina (intyna + egzyna)
Powstawanie sporodermy
powstaje ściana kalozaowa na zewnętrznej błonie komórkowej
pomiędzy kalozą, a błoną powstaje primegzyna
w primegzynie powstają kolumelle
powstaje negzyna i teptum, zanika kaloza
odkładają się sporopolleniny z orbikul
zanika primegzyna
tworzy się intyna
Sporoderma
odpowiada za ochronę ziarna pyłku, gdyż sporopollenina jest odporna na stres
urzeźbienie ułatwia przyleganie do zapylacza i do znamienia słupka
szpiczasta egzyna charakterystyczna jest dla roślin owadopylnych
pyłek wiatropylny jest gładki
Pollenkit (kit pyłkowy)
lepka mieszanina zawierająca białka i lipidy, pozostałość po tapetum
podnosi efektywność zapylenia ze względu na dużą lepkość
chroni pyłek przed warunkami środowiska
odpowiada za sporofitową samoniezgodność zapylenia
występuje w porach egzyny

10
Łagiewka pyłkowa
transporter komórek plemnikowych
jest konieczna, gdyż u roślin wyższych komórki plemnikowe nie mają witek
Ziarno pyłku przed zapyleniem
dojrzewanie w woreczkach polega na
o odwadnianiu
o wejściu w spoczynek (1 dzień – 1 rok), staje się wtedy nieaktywne fizjologicznie, jest
gotowe do rozsiewania, zdolne do kiełkowania

11
WYKŁAD 3
Zapylenie –
proces polegający na przemieszczeniu ziarna pyłku na znamię słupka u roślin
kwiatowych. Jest warunkiem wstępnym i niezbędnym do zapłodnienia
Pylnik otwiera się
pod wpływem dotyku owada
pod wpływem podmuchu wiatru
samoczynnie
Mechanizm otwarcia
wysychanie tkanek
niszczenie komórek ścian pylnika
pęcznienie celulozy w endotecjum
duże naprężenie, ściana nie wytrzymuje bodźca mechanicznego i pęka
Wektory pyłku
wiatr
zwierzęta – owady, ptaki, nietoperze, inne (koewolucja)
siła ciężkości
woda
Entomofilia (entomogamia) – owadopylność
pyłek jest bogatym substratem energetycznym
wabikiem jest nektar, który jest wydzielany do momentu zapłodnienia
najskuteczniejszym zapylaczem jest pszczoła, przenosi do 5 mln ziaren pyłku dziennie, widzi
w zakresie światła UV, nie widzi światła czerwonego
egzyna szpiczasta warunkuje przyczepianie do owada (wspomaga to lepki kit pyłkowy)
rośliny owadopylne produkują ok. 100 razy mniej pyłku niż rośliny wiatropylne.
Pollenkit
pochodzi z tapetum
zlepia ziarna pyłku w agregaty
odpowiada za samoniezgodność
Anemofilia (anemogamia) – wiatropylność
pyłek lekki, suchy, małe ziarna, ale produkowane w dużej ilości
znamiona słupków są bardzo rozbudowane
w zalążni zaledwie 1-2 zalążków
egzyna ziarna pyłku jest gładka, bez płaszcza pollenkitu
pylniki wiszą na długich nitkach pręcikowych
Ornitofilia (ornitogamia) – ptakopylność
głównie w tropikach
gatunki: kolibry, nektarynki
kwiaty są duże, barwne, bezzapachowe, dużo nektaru
Chiropterofilia (chiropterogamia) – nietoperzopylność
kwiaty zapylane są przez nietoperze, typ ten jest popularny w tropikach
kwiaty są słabo zabarwione

12
Hydrofilia (hydrogamia) – wodopylność
rośliny przystosowane są do zapylenia w wodzie
pyłek przenoszony jest przez fale lub prąd wody
czasami rośliny wodne zapylane są jednak przez owady (niski poziom wód)
Kleistofilia (kleistogamia) – samopylność
czasami pylniki pozostają zamknięte i otwierają się w nierozwiniętym kwiecie (kwiaty
kleistogamiczne)
samopylne
rzadko otwierają się, stając się owadopylne
Samopylność
pyłek pochodzi z tego samego kwiatu na którym znajduje się zapylany słupek (obupłciowe)
lub pyłek pochodzi z tego samego osobnika na którym znajduje się słupek (rozdzielnopłciowe,
jednopienne)
samozapylenie jest procesem mało efektywnym, ogranicza zróżnicowanie puli genetycznej
potomstwa
co jakiś czas rośliny autogamiczne muszą się krzyżować, aby zapobiegać zwyrodnieniu i
chowowi wsobnemu
Zapobieganie samopylności
dwupienność – na każdym osobniku występują tylko kwiaty żeńskie lub tylko kwiaty męskie
nierównomierne w czasie dojrzewanie słupków i pręcików
specjalna budowa kwiatów, tzw. heterostylia
o krótkie słupki, długie pręciki
o krótkie pręciki, długie słupki
zapylenie krzyżowe
Obcopylność
przenoszenie pyłku z jednej rośliny (osobnika) na inną roślinę (osobnika) tego samego
gatunku
dotyczy wszystkich roślin kwiatowych
tylko u dwupiennych musi być tego typu mechanizm, nie mogą być fakultatywnie obcopylne
zalety
o przyspiesza ewolucję
o zapewnia zmienność
o zwiększa żywotność
Samoniezgodność zapylenia
inaczej nazywana samobezpłodnością
najskuteczniejszy i najprostszy sposób zabezpieczenia
geny ulegające ekspresji w pręcikach i słupkach mogą być jednakowe lub różne, jeśli są
jednakowe, to ziarno pyłku zostaje odrzucone
w przypadku, gdy białka które uległy ekspresji są różne, pyłek kiełkuje na znamieniu
ekspresja obu alleli na słupku i pręciku powodują zaburzenia związane z
o obecnością pyłku na znamieniu
o wnikaniu łagiewki do szyjki słupka
Geny
gen samoniezgodności – wieloalleliczny, zarówno pyłku jak i słupka, oznaczony jako „S”, a
allele „S1”, „S2”, „S3” itd. są wariantami tego genu

13
są poliformiczne i dziedziczone jako jedna jednostka genetyczna
ziarna pyłku mają genotypy haploidalne, ale od tej samej rośliny pochodzą allele S1 i S2
genotypy słupka są diploidalne
jeśli mamy słupek Sx i słupek Sx, to nie dochodzi do kiełkowania pyłku, jeśli słupek ma
genotyp Sx, a ziarno pyłku Sy, to może kiełkować łagiewka pyłkowa
Podział alleli samoniezgodności
diploidalny mikrosporocyt ulega podziałowi
powstaje diada, a następnie tetrada, genetycznie mamy więc 2 komórki jednego rodzica i
dwie drugiego rodzica
rozwija się ziarno pyłku, początkowo dwukomórkowe, później trójkomórkowe
ważna zasada: w jednym locus S występują przynajmniej dwa wieloalleliczne geny. jeden z
nich jest odpowiedzialny za genotyp słupka i pręcika. Pojawiają się zatem fenotypy: Sl i Pl
w diploidzie genotyp jest jakby podwójnym haplotypem, a z ziarnach pułku mamy pojedynczy
haplotyp
wszystkie geny S są dziedziczone jako jedna jednostka genetyczna
Fenotyp słupka
genotyp – zestaw genów; fenotyp – zdeterminowany wygląd
allelem jest wariant danego genu, w tym przypadkiem Sx
haplotyp – genom haploidalny; diplotyp – genom diploidalny
fenotyp słupka – zdeterminowany podwójnym haplotypem
geny ulegają ekspresji w komórkach
w pyłku o fenotypie decyduje
o diploidalny genotyp komórki macierzystej – zależy od tapetum
o haploidalny genotyp ziarna pyłku – zależy od pyłku
Fenotyp pyłku
ze względu na determinację fenotypu ziarna pyłku wyróżniamy
o samoniezgodność gametofitową – u gatunków rośli, u których fenotyp jest wynikiem
ekspresji genu S występującego w pojedynczym haplotypie pyłku
o samoniezgodność sporofitową – fenotyp pyłku jest wynikiem ekspresji genu S obu
alleli tkanek sporofitu, głównie pylnika (tapetum)
Samoniezgodność gametofitowa
jako samoniezgodny rozpoznawany jest ten wariant, który ma allel S identyczny z jednym z
dwóch alleli genotypu słupka (np.: pyłek S1 i słupek S1 i S2)
wyróżniamy dwa typy samoniezgodności gametofitowej
o typ Solanaceae – zahamowanie rozwoju męskiego gametofitu ma miejsce dopiero w
górnej części szyjki słupka
o typ Papaveraceae – zahamowanie rozwoju gametofitu męskiego ma miejsce już na
znamieniu słupka
s słupek wrastają tylko te łagiewki, które mają różne od słupka haplotypy
Typ Solanaceae
ziarno od początku jest niezgodne, ale kiełkuje
zahamowanie następuje w podznamieniowej części słupka
szlak transmisyjny – komórki szyjki uczestniczą w reakcji samoniezgodności
opisano u trzech rodzin okrytonasiennych
o Rosaceae (różowate)
o Solanaceae (psiankowate)

14
o Scrophulariaceae (trędownikowate)
Budowa locus S i jego produktów w typie Solanaceae
w locus S zidentyfikowano dwa geny
o gen S-RNA-azy ulegający ekspresji w słupku
o gen SFB ulega ekspresji w pyłku
produkty genów
o S-RNA-azy – białko z domenami PS (sekwencja sygnałowa), C1-C5 (domeny
konserwatywne), C2-C3 (sekwencje katalityczne), HVa i HVb (regiony wysoce
zróżnicowane, wykazują alleliczny polimorfizm genu)
o SFB – białko z domenami F-box (sekwencje konserwatywne), regiony wysoce
zróżnicowane (uczestniczą w rozpoznawaniu RNA-azy)
Mechanizm molekularny typu Solanaceae
fenotyp pyłku S2 i słupka (S1 S3) są odmienne
wniknięcie słupkowej RNA-azy do łagiewki
rozpoznanie S1-RNA-azy przez białko SFB2 jako odmiennej, obcej
białko SFB2 jest inhibitorem S1-RNA-azy
następuje ubikwitynacja RNA-azy
enzym ulega degradacji w proteasomach 26s (brzmi znajomo)
ocalony RNA pyłku powoduje wzrost łagiewki
Mechanizm u Papaveraceae
rozpoznanie i odrzucenie niezgodnego ziarna pyłku odbywa się już na znamieniu słupka,
łagiewka nie wrasta
reakcje te oparte są na oddziaływaniu sygnał-receptor
dzieje się to na drodze przekazywania sygnału wapniowego
układ sygnalizacyjny to
o cząsteczki sygnałowe – ligandy – białka S znamienia
o receptory (białka typu S)
o kaskada wewnątrzkomórkowych procesów prowadzi do zahamowania wzrostu
łagiewki niezgodnych ziaren pyłku
występuje u Papaveraceae (makowatych)
Mechanizm molekularny typu Papaveraceae
ligand S1 jest rozpoznawany przez receptor S1
uruchomienie kaskady zdarzeń
o wzrost stężenia jonów wapnia (zasoby wewnątrz i zewnątrzkomórkowe)
o aktywacja kinaz białkowych (zależnych od CAM i wapnia), które regulują aktywność
białek regulatorowych
efektem jest
o spadek gradientu wapnia
o depolimeryzacja aktyny
o zahamowanie syntezy składników niezbędnych do wzrostu łagiewki’
konsekwencją jest zahamowanie wzrostu łagiewki pyłkowej
ostatecznie dochodzi do PCD i fragmentacji jąder pyłkowych

15
Samoniezgodność sporofitowa
jako niezgodny rozpoznawany jest ten pyłek, który wytworzyła roślina posiadająca
przynajmniej jeden haplotyp S1 S2 wspólny z jednym z haplotypów S2 S3
zahamowanie rozwoju ziarna pyłku ma miejsce na znamieniu słupka, a łagiewka nie wrasta
wrasta łagiewka ziaren pyłku wytworzonych przez roślinę, której obydwa haplotypy S3 S4 są
różne od haplotypów S1 S2
gatunkami z samo niezgodnością sporofitową są Brassicaceae (kapustowate)
Determinanty samoniezgodności sporofitowej
rozpoznawanie i odrzucanie niezgodnego ziarna pyłku oparte jest o interakcję receptor-
ligand
determinanty
o żeński – białko obecne w papilli znamienia (receptor)
o męski – białko obecne w płaszczu pyłku (ligand)
Geny niezgodności sporofitowej w locus S
wyróżniamy 3 geny odpowiedzialne za ten proces
o SLG – białko wydzielnicze o wysokim stopniu polimorfizmu, wiąże ligand (ekspresja
na znamieniu słupka)
o SRK – białko receptorowe, kinaza serynowo-treoninowa (ekspresja na znamieniu
słupka)
o SP11/SPR – białko sygnałowe, wysoce polimorficzne (ekspresja w komórkach
tapetum)
Struktura i funkcja białek SRK
białko wydzielnicze, receptorowe, transbłonowe, o aktywności kinazy receptorowej (Ser/Tre)
w znamieniu słupka
zawiera domeny
o PS – sekwencja sygnałowa
o DS - zewnątrzkomórkowa domena z trzema wysoce polimorficznymi regionami
decydującymi o rozróżnieniu allelicznym, rozpoznaje i wiąże ligand pyłkowy, gdy
haplotypy są takie same
o DT – domena transbłonowa
o DK – konserwatywna domena kinazy Ser/Tre
Struktura i funkcja białek SLG
glikoproteina wydzielnicza o wysokim polimorfizmie
wydzielana do ściany papilli znamienia
wysoce homologiczne do domeny DS. białka SRK
posiada sekwencję sygnałową PS
posiada trzy regiony wysoce zróżnicowane
przypuszczalna funkcja to
o tworzenie połączenia z białkiem SRK umożliwiając wiązanie liganda pyłkowego
o uczestniczy w dojrzewaniu białka SRK
o jest przenośnikiem ligandów
Struktura i funkcja białka SCR
białko sygnałowe (ligand)
zlokalizowane w płaszczu lipidowym pyłku
ulega ekspresji w sporoficie, w tapetum, stąd eksportowane do sporodermy
zawiera sekwencję sygnałową PS

16
białko wysoce polimorficzne (50-66aa), konserwatywne jest jedynie 8 reszt Cys i jedna Gly na
C-końcu, pozostałe aa są zmienne
Inicjacja zapylenia
bezpośredni kontakt między sporodermą a ścianą znamienia
sporoderma uwalnia lipidowo-białkowe opłaszczenie
między pyłkiem a papillą wytwarza się stopka odpowiedzialna za adhezję i uwolnienie pyłku
białka SP11/SPR przez kanały w kutikuli wchodzą do ściany papilli i docierają do kompleksu
receptorowego znamienia.
Molekularny mechanizm samoniezgodności sporofitowej
uwolnienie białka SP11/RSC ze sporodermy
transport SP11/SCR przez glikoproteinę SLG
interakcja z domeną receptora SRK (gdy haplotypy są takie same)
dysocjacja SRK od inhibitora fosforylacji
autofosforylacja SRK
aktywacja funkcji kinazowej SRK
fosforylacja białek uczestniczących w reakcji samoniezgodności
fosforylacja białka ARC (przenoszące ubikwitynę)
ARC1 wiąże się do białek niezbędnych w rozwoju pyłku
następuje ubikwitynacja
degradacja w proteasomach 26s
efekt – odrzucenie pyłku
Uwagi do samoniezgodności
proces trwa ok. 45 minut

17
WYKŁAD 4
Faza progamiczna
wydarzenia w okresie od zapylenia do zapłodnienia
łagiewka przerasta przez słupek i przedostaje się do woreczka zalążkowego
następuje wzajemne oddziaływanie między łagiewką i komórkami słupka
Procesy związane z aktywacją ziarna pyłku
ziarno pyłku jest uśpione i nieaktywne
po upadku na znamię słupka następuje adhezja (adhezja)
ziarno pyłku zostaje aktywowane (aktywacja)
tkanki słupka przejmują rolę wprowadzenia łagiewki
w wyniku uwodnienia dochodzi do aktywacji genów łagiewki pyłkowej (cer, rtg) (uwodnienie)
następuje kiełkowanie i aktywacja genów (tip1, seth 1 i 2) (kiełkowanie)
przerost tkanek słupka (penetracja słupka)
aktywny wzrost łagiewki (ffd34, ffd42, mad, pgd, anx 1 i 2) (wzrost łagiewki)
wzrost ukierunkowany do woreczka (wzrost ukierunkowany)
przebicie ściany woreczka zalążkowego
zapłodnienie
Kiełkowanie ziarna pyłku
ziarno pyłku pada na znamię słupka
ziarno zwiększa swoją objętość na skutek pobierania wody z powierzchni znamienia
intyna tworzy uwypuklenie i tworzy ujście łagiewki w egzynie wydostając się na zewnątrz w
postaci łagiewki pyłkowej
łagiewka składa się z protoplastu pochodzącego z komórki wegetatywnej otoczonej ścianą
komórkową (celulozową)
Droga łagiewki pyłkowej
znamię
szyjka
przegroda zalążni
komora woreczka zalążkowego
mikropyle (czasami od strony chalazalnej lub integumentu)
Przerastane tkanki
tkanka znamienia
tkanka szyjki
tkanka transmisyjna
tkanka ściany zalążka
tkanka sznureczka
tkanka osłonek
komórki synergid
Oddziaływania wzajemne pyłku i słupka
pyłek - wydziela kutynazę i inne enzymy, rozkładają one ścianę komórkową znamienia i szyjki,
ułatwiają penetrację łagiewki pyłkowej
łagiewka – wydziela hormony i enzymy z klasy pektynaz: poligalakturonazy, esterazy, liazy,
które degradują pektyny na drodze łagiewki pyłkowej, później stanowią materiał budulcowy
dla ściany łagiewki

18
słupek – wydziela wodę umozliwiająca kiełkowanie, substancje odżywcze i wzrostowe, które
mogą być bardzo zróżnicowane (np.: flawonoidy), enzymy degradujące inhibitory kiełkowania
i wzrostu, raz chemocyjaniny kierunkujące wzrost łagiewki w słupku
w przypadku niezgodności:
o słupek nie wytwarza czynnika kiełkowania łagiewki
o słupek wytwarza substancje hamujące wzrost łagiewki
o słupek nie wydziela wody umożliwiającej prawidłowy wzrost lagiewki
Tkanka transmisyjna
komórki słupka wytwarzają specyficzne warunki
o wydłużone komórki transmisyjne tworzą szlak dla kiełkującej łagiewki
o komórki luźno ułożone bez blaszki poprzecznej umożliwiają wzrost
o komórki wydzielnicze odżywiają łagiewkę
o napęczniałe ściany komórkowe są łatwiejsze w penetracji
o przestrzenie międzykomórkowe wypełnione są maziowatą substancją – macierzą
zewnątrzkomórkową (ECM), która składa się z wody, substancji pektynowych,
polisacharydów, białek i czynniki kierunkujące wzrost. Umożliwia to wzrost i rozwój
łagiewki, oraz odżywia ją
Wyjście łagiewki na przegrodę
łagiewka wychodzi z tkanki transmisyjnej szyjki słupka i dalej rośnie na przegrodzie
nie jest dokładnie znany mechanizm przejścia z tkanki transmisyjnej na przegrodę
prawdopodobne jest, że warunkuje to lokalna degradacja kutikuli
Sygnały w obrębie zalążni
w zalążniach wielozalążkowych gametofity niekoniecznie są zapładniane w kolejności
ułożenia zalążków , łagiewki często omijają płodne zalążki, być może jest to kwestia
przypadku, ale niewykluczone, że jest to mechanizm preferencyjnego zapłodnienia. Im więcej
kiełkujących ziaren pyłku, ty większe prawdopodobieństwo, że zalążki zostaną zapłodnione
po kolei (Arabidopsis thaliana)
tkanki słupka stanowią nie tylko trakt dla łagiewki, ale i umożliwiają jej odbiór sygnału
wysyłanych przez zalążki, które kierunkują wzrost
każdy płodny zalążek wydziela na powierzchnię łożyska substancje chemotaktyczne, czyli
atraktanty, które dyfundują i rozpraszają się wyznaczając drogę wzrostu. Są to tak zwane
czynniki długodystansowe
Droga łagiewki pyłkowej po powierzchni sznureczka
łagiewka rośnie przylegając do sznureczka, kierunek wyznaczają diploidalne tkanki zalążka
łagiewka nagle skręca i kieruje się w stronę mikropyle. Kierunek wyznaczają sygnały z
gametofitu żeńskiego, głównie z synergid
czynniki krótkodystansowe występujące na powierzchni synergid, sznureczku i kanale
mikropylarnym zalążków kierunkują wzrost łagiewki
Tkanka mikropylarna
przygotowuje środowisko wzrostu łagiewki do receptywnych (płodnych) zalążków
cechuje się nasiloną aktywnością wydzielniczą
występuje powiększenie przestrzeni międzykomórkowych
przestrzenie międzykomórkowe wypełnione są wydzieliną zawierającą
o substancje odżywcze i budulcowe
o pektyny
o substancje kierunkujące wzrost łagiewki

19
o jony wapnia
Rozwój gametofitu żeńskiego
aparat jajowy – 2 synergidy + 1 komórka jajowa – biegun mikropylarny
1 komórka centralna (2 jądra biegunowe) – po środku
3 antypody - biegun chalazalny
Zmiany w zalążku
przed zapłodnieniem – fuzja jąder komórki centralnej, degeneracja antypod
po wniknięciu łagiewki pyłkowej – migracja łagiewki w kierunku jednej z synergid, wzrost
łagiewki w synergidzie, wniknięcie do synergidy, zatrzymanie wzrostu, degeneracja tejże
synergidy, pęknięcie łagiewki, wylanie cytoplazmy z uwolnionymi jądrami plemnikowymi
Wybór synergidy
synergida wybierana (receptywna) wydziela atraktanty, ale również jest częściowo
zdegenerowana, ma uszkodzoną błonę
degeneracja zaczyna się tuż po zapyleniu, kiedy łagiewka wzrasta w szyjce słupka
osłabienie i sygnały chemotaktyczne kierują łagiewkę do receptywnej synergidy
druga synergida pozostaje nienaruszona do chwili pomyślnego zapłodnienia obu komórek
Funkcje synergid
działanie chemotropowe na łagiewkę poprzez wydzielanie białek wabiących i atraktantów
umożliwiają pękanie wierzchołka łagiewki
uwalniają plemniki do jednej z synergid
Uwolnienie zawartości łagiewki
następuje zahamowanie wzrostu łagiewki w synergidzie
wierzchołek łagiewki ulega powiększeniu
dochodzi do pęknięcia łagiewki
o sygnałem ku temu jest białka z synergidy – ZmES4 (DEFL)
o odbiorcą jest kanał jonów potasowych – KZM1
o następuje depolaryzacja błony
o wierzchołek łagiewki pęka
o dochodzi do energicznego wyrzutu zawartości
pozostałe czynniki biorące udział w pękaniu
o ANX1 i ANX2 – zapobiega pękaniu łagiewki zanim ta dotrze do synergidy
o ACA9 – pompa wapniowa, hamuje wzrost łagiewki i warunkuje pękanie
Po wejściu łagiewki do synergidy, zachodzi degeneracja tejże synergidy (PCD), jest to proces
niezbędny do zahamowania wzrostu łagiewki
Komórki plemnikowe w woreczku zalążkowym
obie komórki plemnikowe są ze sobą połączone poprzecznie błoną komórkową, a syncytium
to powstało już w wyniku podziału mitotycznego komórki generatywnej, ponadto tacza je
błona komórki wegetatywnej
takie połączenia umożliwia synchronizację docierania do synergidy (Arabidopsis thaliana)
z synergidy jądra wydostają się do przestrzeni między komórkami woreczka zalążkowego i
układają się między komórką jajową, a komórką centralną
20
Izomorfia komórek plemnikowych
u większości roślin kwitnących, w tym u Arabidopsis, obie komórki nie są morfologicznie
zróżnicowane, i są równocenne kompetentne w zapładnianiu zarówno komórki jajowej jak i
centralnej
prawdopodobieństwo zapłodnienia komórki jajowej i centralnej jest takie same, zarówno
przez jedną jak i drugą komórkę plemnikową
tylko u niektórych gatunków, np.: Plumbo zeylanica ma miejsce asymetryczny podział
komórki generatywnej prowadzący do dymorfizmu komórek plemnikowych. Większe jądra
plemnikowe z większą liczbą mitochondriów preferencyjnie zapładniają komórki centralne
Łączenie się gamet – podwójne zapłodnienie
zlanie się cytoplazmy nazywamy plazmogamią, zlanie się jąder to kariogamia
komórki żeńskie podlegające zapłodnieniu (komórka jajowa i centralna) nie mają kompletnej
ściany komórkowej
plemniki wcale nie posiadają ściany komórkowej (są zatem naturalnymi protoplastami)
w miejscu syngamii (połączenia gamet) stykają się błony obu gamet
ścisła kontrola tego procesu zapewnia najpierw adhezję (przyleganie), a następnie fuzję
(łączenie) komórki jajowej i plemnikowej, oraz komórki centralnej i plemnikowej – te dwa
etapy są krytyczne
Adhezja i fuzja gamet
adhezja komórek i fuzja gamet to procesy niezbędne i konieczne do zapłodnienia
białko GEX2 na powierzchni męskich gamet jest niezbędne do adhezji
białko HAP2/GCS1 (fuzogen) ulokowane są po wewnętrznej stronie błony i pęcherzykach
cytoplazmatycznych w gametach męskich, jest to niezbędne do fuzji gamet
białka EC1 (kumulowane w pęcherzykach wydzielniczych cytoplazmy komórki jajowej) są
wydzielane na zewnątrz
następuje aktywacja komórki plemnikowej reorganizującej swoją błonę komórkową,
następuje powierzchniowa ekspozycja białek HAP2/GCS1
ponadto białka TET zlokalizowane w błonach komórek plemnikowych (TET 1 i TET 2), oraz
komórek jajowej i centralnej (TET 9) prawdopodobnie wspomagają fuzję komórek
Komórki podwójnego zapłodnienia (amfimiksji)
Ziarno pyłku
1n - komórka plemnikowa
1n - komórka plemnikowa
Woreczek zalążkowy
1n – komórka jajowa
2n – komórka centralna
Efekt
2n – zygota
3n – jądro bielmowe
początkowo łagiewka niesie jedną komórkę generatywną i jedną wegetatywną
komórka generatywna dzieli się, tworzą się dwie komórki plemnikowe
łagiewka z dwoma komórkami plemnikowymi wchodzi do synergidy, zanika jądro
wegetatywne
synergida rozpada się
jedno jądro plemnikowe zapładnia komórkę centralną, a jedno komórkę jajową
Interakcje łagiewka – woreczek u Arabidopsis
łagiewka wrasta do receptywnej synergidy – jądro wegetatywne ulokowane jest przed
komórkami plemnikowymi. jądro komórki wegetatywnej jest zasocjowane z komórką
plemnikową leżącą za nią (męska jednostka generatywna)

21
pękniecie łagiewki pyłkowej i gwałtowne wyrzucenie zawartości – dochodzi do zmieszania
zawartości łagiewki z zawartością synergidy, zawartość rozlewa się poza synergidę
(dezintegracja błony)
w ciągu jednej minuty komórki plemnikowe docierają do miejsca fuzji, pomiędzy komórkami
jest tylko wąska szczelina
po kilku minutach (ok. 7) bezruchu (w wyniku komunikacji gamet męskich i żeńskich)
następuje plazmogamia jednej komórki plemnikowej z komórką jajową, zapłodnienie
wyzwala natomiast mechanizm zabezpieczający przed fuzją drugiej komórki plemnikowej z
powstałą zygotą (blok przeciw polispermii). Taki sam mechanizm występuje w komórce
centralnej
nieco później ma miejsce plazmogamia drugiej komórki plemnikowej z komórką centralną, od
razu odbywa się kariogamia między jądrami tych komórek. Szybka dekondensacja
chromatyny jądra plemnikowego w j komórce centralnej
po zsynchronizowaniu cykli komórkowych obu komórek następuje kariogamia jadra
pierwszej komórki jajowej. Dekondensacja chromatyny zapłodnionego jądra komórki
centralnej , a następnie aktywacja cykli komórkowych (szybciej niż w zapłodnionej komórce
jajowej)
Ile ziaren pyłku kiełkuje na jednym znamieniu?
w słupku wielozalążkowym kiełkuje wiele ziaren pułku, więcej niż jest zalążków
kiełkuje tylko pyłek zgodny
łagiewka pyłku niezgodnego o ile wykiełkuje, degeneruje w części podznamieniowej
niekiedy pyłek niezgodny pokona bariery i zapładnia komórki zalążka. Powoduje to:
o powstawanie mieszańców niezgodnych roślin
o powstawanie mieszańców międzygatunkowych
Ile zalążków penetruje jedna łagiewka?
zazwyczaj jedna łagiewka wydostaje się z przegrody i zmierza do jednego mikropyle
zdarza się, że wiele łagiewek zmierza do jednego zalążka, ale dociera tylko jedna
sporadycznie zdarza się, że kilka łagiewek dotrze do mikropyle, ale wchodzi tylko jedna
łagiewki nie wchodzą do zapłodnionego zalążka
Mechanizmy zabezpieczenia
pomyślne zapłodnienie inicjuje degradację drugiej synergidy, która zaprzestaje wysyłania
atraktantów w celu zwabienia łagiewek
sygnały repulsywne (odpychania) pochodzą z różnych źródeł i mają na celu zahamowanie
wzrostu innych łagiewek
o od zalążka – komórek sporofitowych, komórek gametofitu
o od łagiewki – łagiewki zapładniającej, konkurujących łagiewek
Tylko dwie szanse na zapłodnienie zalążka
łagiewki pyłkowe są przetrzymywane w słupku dość długo (do 28h), dzieje się tak na
wypadek niepowodzenia w zapłodnieniu pierwszą łagiewką
system awaryjny zostaje uruchomiony, gdy pierwsza łagiewka dostarcza niezdolne do
zapłodnienia komórki plemnikowe
druga i ostatnia szansa do pomyślnego zapłodnienia zalążka to wniknięcie drugiej łagiewki,
trzecia nie wniknie ze względu na brak trzeciej synergidy – wyjątkiem jest Amborella
trichopoda, która posiada 3 synergidy

22
Awaryjny system zapładniania u Arabidopsis
pierwsza łagiewka wabiona jest przez atraktanty receptywnej synergidy, łagiewka dostaje się
do niej, ginie synergida, która tworzyła atraktant, następnie są dwie opcje
o sytuacja normalna – pomyślna fuzja gamet wyzwala blokadę innych łagiewek,
degeneracja drugiej synergidy , zaprzestanie wabienia innych łagiewek, brak
atraktantów
o sytuacja awaryjna – jądra plemnikowe posiadają defekt, nie dochodzi do fuzji
komórek, pierwsza synergida zostaje zniszczona, druga jest nadal nienaruszona. Po
kilku godzinach (z powodu natychmiastowej blokady ze strony pierwszej łagiewki –
czas latencji) druga synergida zaczyna produkować atraktanty, które wabią drugą
łagiewkę. Po fuzji atraktanty przestają być produkowane
Zdarzenia po wniknięciu łagiewki do zalążni
wkiełkowanie łagiewki do zalążni
wzrost łagiewki na sznureczku
wyjście łagiewki do mikropyle
kierowanie łagiewki do synergidy
wejście łagiewki do synergidy
zatrzymanie wzrostu łagiewki z synergidzie
pęknięcie i wyrzut zawartości łagiewki
degeneracja synergidy
podwójne zapłodnienie
Efekt podwójnego zapłodnienia
Zalążek
Zygota
Zarodek
Nasienie
Owoc
Jądro bielmowe
Bielmo
Osłonki
Łupina nasienna
Ściany zalążni
Owocnia
Strefy w łagiewce pyłkowej
strefa wierzchołkowa – strefa wzrostu (zawiera materiały budulcowe)
strefa podwierzchołkowa
strefa trzonowa
strefa zamykania (kaloza)
Udział cytoszkieletu we wzroście łagiewki
ściana komórkowa zbudowana jest z kalozy i celulozy
pęcherzyki budulcowe niesione są przez filamenty aktynowe
wzrost zależy zatem od aktyny
ruch cytoplazmy jest odwrotny do fontannowego
pęcherzyki uwalniają się w wierzchołku, a w części podwierzchołkowej aktyna depolaryzuje
część pęcherzyków jest wykorzystywana, a nadmiar zawraca i krąży
Szybkość wzrostu łagiewki i czas trwania fazy progamicznej
sposoby i tempo wzrostu zależą od gatunku, liczby ziaren na znamieniu, kiełkujących
łagiewek, dojrzałości tkanek słupka, temperatury, wilgotności
szybkość wzrostu – 1 – 3,5mm na godzinę, u kukurydzy ponad 30cm, czas potrzebny do
zapylenia waha się od kilku godzin do kilku dni lub miesięcy

23
WYKŁAD 5
Owoc
u okrytozalążkowych
organ powstający z zalążni słupka i czasami innych części, głównie dna kwiatowego
składa się z owocni i nasion
znamię przekształca się w twór na końcu owocu, zalążki w nasiona, a ściana zalążni w
owocnię
Owocnia (perykarp) składa się z trzech warstw
egzokarp – powstaje z zewnętrznej części zalążni słupka
mezokarp – powstaje z miękiszu ściany zalążni słupka
endokarp – powstaje ze skórki wewnętrznej części zalążni słupka (owocolistka)
Właściwości owocu
chroni nasiona przed warunkami środowiska w czasie rozsiewania nasion
pełni funkcję powabni, usprawniając rozsiewanie nasion
niektóre owoce nie mają jednak nasion i powstają w wyniku partenokaprii (np.: banan)
Klasyfikacja owoców
ze względu na sposób powstawania
o owoc właściwy – powstaje wyłącznie ze ścian zalążni słupka
o owoc rzekomy (szupinkowy) – powstaje nie tylko z zalążni, ale również innych części
kwiatu, najczęściej wykształca się z rozrośniętego, mięsistego dna kwiatowego
ze względu na liczbę zalążni
o owoce pojedyncze – kwiat pojedynczy, jeden słupek
o owoc zbiorowy – kwiat pojedynczy, słupkowie
o owocostan – kwiatostan
ze względu na postać owocni
o suche – skórzasta lub zdrewniała owocnia
pękające – łuszczyna, torebka, mieszek, strąk
niepękające – orzech, ziarniak, niełupka, rozłupnia
o mięsiste – soczyste, z dużą zawartością wody
pojedyncze – jagoda, pestkowiec
zbiorowe – wielopestkowiec, wieloorzeszkowiec
Owoce pojedyncze suche
pękające – dojrzewają i pękają wyrzucając nasiona
o mieszek – powstaje z jednego owocolistka, otwiera się wzdłuż szwu zrośnięcia
(brzusznego) – piwonia, kaczeniec
o strąk – powstaje z jednego owocolistka, ale otwiera się zarówno wzdłuż szwu
zrośnięcia i wzdłuż żyłki środkowej owocolistka, nasiona ułożone są wzdłuż szwu
brzusznego – groch, fasola
o torebka – powstaje z zalążni zrośniętej z kilku owocolistków, może pękać w różny
sposób: wzdłuż (całkowicie lub częściowo), okrężnie, lub przez powstanie otworów w
ścianie owocni – mak
o łuszczyna – powstaje z dwóch owocolistków, pęka dwiema klapami, pomiędzy
którymi znajduje się tzw. fałszywa przegroda, na której umieszczone są nasiona –
rzepak
o łuszczynka – podobna do łuszczyny, ale krótka i szeroka, o owalnym kształcie –
tasznik

24
niepękające – gdy dojrzewają nie otwierają się, ale wraz z nasionami odpadają od rośliny
o orzech – ma owocnię zdrewniałą, zawiera jedno nasiono, z którym owocnia się nie
zrasta – leszczyna
o ziarniak – ma skórzastą owocnię, która zrasta się z łupiną nasienną pojedynczego
nasiona – kukurydza, zboża
o niełupka – ma owocnię skórzastą, niezrastającą się z pojedynczym nasieniem –
słonecznik
o rozłupnia – owoc wielonasienny, rozpada się przy dojrzewaniu na suche, niepękające
owocki zwane rozłupkami, odpowiadające poszczególnym owocolistkom i
zawierające po jednym nasieniu – kminek
Owoce pojedyncze mięsiste
jagoda – owoc o mięsistej, niepękającej owocni, zbudowanej z zewnętrznego egzokarpu, oraz
mięsistego mezokarpu, który wypełnia całe wnętrze owocu. Cechą charakterystyczną jagody
jest brak endokarpu. bezpośrednio w mezokarpie znajdują się liczne (rzadko jedno –
awokado) nasiona – porzeczka, kiwi, winorośl, pomidor
pestkowiec – tylko środkowa część owocni jest mięsista – mezokarp. Egzokarp tworzy skórkę,
a wewnętrzny endokarp jest twardy, silnie zdrewniały, zbudowany ze sklerenchymatycznej
tkanki – komórek kamiennych. Ta część owocni wraz z zawartymi w niej nasieniem nosi
nazwę pestki – śliwa, brzoskwinia
Owoce zbiorowe
owoc wielopestkowy – z poszczególnych zalążni wielosłupkowego kwiatu powstają drobne
pestkowce, które zrastają się ze sobą mięsistymi częściami owocni i osadzone są na jednej
wspólnej osi powstałej z dna kwiatowego – malina
owoc wieloorzeszkowy – liczne zalążnie kwiatu przekształcają się w drobne orzeszki,
osadzone pojedynczo na silnie rozwiniętym, mięsistym i soczystym, wypukłym dnie
kwiatowym – truskawka
wielomieszek – z poszczególnych zalążni powstają mieszki osadzone na wspólnym dnie
kwiatowym – jaskier
szupinka – jabłoń
Owocostany
powstają z przekształcenia całych kwiatostanów
w ich utworzeniu oprócz zalążni wielu kwiatostanów biorą udział także dno kwiatowe,
okwiat, liście kwiatostanowe, oś kwiatostanu
przykłady – owocostan pestkowcowy figi, jagodostan ananasa
Budowa ziarniaka pszenicy
okrywa owocowo-nasienna
bielmo
o warstwa aleuronowa
o bielmo skrobiowe
zarodek (położenie biegunowe)
o liścień (tarczka)
o koleoptyl
o zawiązki liści
o koleoryza
o korzeń zarodkowy
o czapeczka korzenia

25
Szupinka jabłoni - powstawanie
grubiejące dno kwiatowe tak mocno inwazyjnie otacza słupek i pręciki, że pochłania je,
zostają zalane
płatki korony i działki kielicha więdną, wystają poza owoc tworząc charakterystyczną
strukturę
czasami poza owoc wystaje zaschnięta szyjka słupka
Owoc pozorny szupinkowy – jabłoń
skórka - pokryta jest grubą kutikulą, która chroni przed nadmiernym parowaniem, a w
miejscu aparatów szparkowych tworzą się przetchlinki
owocnia rzekoma - jest jadalną, miękiszową częścią jabłka dojrzałego, zbudowana jest z
bardzo dużych komórek, dosyć luźno ułożonych, mających cienką, przyścienną warstwę
cytoplazmy, oraz bardzo duże wakuole zawierające sok komórkowy. Część jadalna jabłka, od
skórki do wiązek, powstaje z dna kwiatowego, pozostała część położona głębiej pod wiązkami
powstaje z owocni i jest owocem właściwym.
wiązka przewodząca – jest ich 10, należą do działek kielicha i płatków korony
egzokarp i mezokarp – stanowi zewnętrzną, mięsistą część perykarpu, nie różnią się od siebie
endokarp – tworzy niejadalną, łuskowatą, pergaminową wyściółkę komór nasiennych,
zbudowaną z wydłużonych w kierunku osi jabłka sklereidów, o bardzo grubych ścianach.
komora nasienna - zawiera po dwa wykształcone nasiona
nasiona – podobnie jak u pestkowców pokryte są dwiema łupinami: wewnątrz biała, trudno
przepuszczalna dla gazów i regulująca ich wymianę, oraz zewnętrzna brązowa, chroniąca
nasienie przed urazami mechanicznymi
szypułka – stanowi dolną część owocu
wgłębienie kielichowe – zawiera zaschnięte pręciki i znamię słupka, stanowi górną część
owocu
Zalążek, a nasienie
osłonki łupina nasienna
ośrodek obielmo
jądra biegunowe bielmo
komórka jajowa zarodek (liścienie, merystem pędu, hipokotyl, korzeń zarodkowy)
Rozwój bielma
może powstać bez zapłodnienia (w wyniku zapylenia napromieniowanym pyłkiem, lub
autonomicznie, czyli naturalnie). Dzieje się tak u jabłoni, kakaowca, ogórka, kiwi
autonomiczny rozwój bielma zachodzi u bawełny, jęczmienia, ryżu, Melandrium album,
Helloborus niger, Lupinus luteus, Brassica napus
Typy bielma
nuklearne – inaczej jądrowe, Leguminaceae, Graminaceae, Cucurbitaceae, powstaje w
postaci skupień w części zewnętrznej, początkowo nie obserwuje się procesu cytokinezy
celularne – inaczej komórkowe, Plantaginaceae, Slantaginaceae, po każdym podziale jądra
dochodzi do cytokinezy i wytworzenia ściany
helobialne – inaczej pośrednie, Helobiaceae, Trisium, Junkus, powstaje jedna ściana,
naprzemiennie występują cykle z cytokinezą i formowaniem ściany i cykle bez tych procesów.
Rodzaje nasion
bielmowe – Ricinus communis (rącznik)
obielmowe – Beta vulgaris (burak)
bezbielmowe – Pisum sativum (groch)

26
Zakładanie się ścian w bielmie
triploidalne bielmo pierwotne zlokalizowane jest na proksymalnym końcu komórki centralnej
powstaje wielojądrowe cenocytowe bielmo – peryferycznie zlokalizowane jądra otaczają
centralną wakuolę
dochodzi do cyklu mitoz zachodzących falami, w czasie ich trwania można dostrzec wrzeciona
cytokinetyczne między jądrami siostrzanymi i niesiostrzanymi
tworzy się radialny system mikrotubul (RMS)
tworzą się ściany komórkowe prostopadłe do warstwy komórek położonych poniżej,
prowadzi to do powstania alweoli
proces powtarza się, prowadzi to do powstania wielu pokładów warstw bielma
Endosperma – bielmo
u większości roślin jest triploidalne, występuje u roslin z woreczkiem zalążkowym typu
Polygonium, Alium, Drusa i Adoxa
u pozostałych typów woreczków waha się w granicach 2-15n
wiąże się to z występowaniem różnej liczby jąder biegunowych, lub pewnymi procesami
wtórnymi, które zachodzą w toku różnicowania się woreczka zalążkowego.
oenothera – gametofit żeński wytwarza tylko jedno jądro biegunowe, które łącząc się z
komórką plemnikową daje diploidalne (2n) jądro endospermy
penea, plumbago – gametofit żeński wytwarza cztery jądra biegunowe, stąd endosperma jest
pentoidalna (5n), podobnie u Frutillaria i Plumbagella
Haustoria bielmowe (endosporialne)
wypustki w kształcie ssawek przerastające tkanki zalążka
pobierają substancje odżywcze i przenikają do tkanki przewodzącej
bielmo jądrowe jest płynne i soczyste, bielmo komórkowe jest twarde i zmączniałe
u części roślin występują poliploidalne jądra haustoriów (96-384C DNA)
o niecierpek
o dynia
o tocia
Sposoby rozsiewania się owoców i nasion
samorzutne – samosiewne (autochoria)
o wyrzucanie
w skutek zmiany turgoru – niecierpek
w skutek działania włókien mechanicznych – motylkowe
o wysypywanie – mak
z udziałem czynników zewnętrznych – obcosiewne (allochoria)
o wiatr – nasiona małe, miękkie (orchidea), nasiona i owoce zaopatrzone w aparaty
lotne, skrzydełka lub puch (mniszek)
o woda – owoce i nasiona zawierają w tkankach dużo powietrza, dzięki czemu nie toną
(lilia wodna)
o zwierzęta – obecność haczyków, zadziorków, elementów czepnych (rzepak), zjedzone
nasiona mogą ulec strawieniu i być wydalane z kałem (borówka)
o człowiek – nieświadomie, jak zwierzęta, świadomie przez siew i handel, transport
Obcosiewność
zoochoria
o myrmekochoria – mrówki
o ornitochoria – ptaki
antropochoria

27
o etolochoria – świadome rozsiewanie
o speirochoria – chwasty w uprawach
o agochoria – rozprzestrzenianie z produktami spożywczymi, paszami
hydrochoria – grążel, palma kokosowa
anemochoria – mlecz, klon, jesion
Samosiewność
blastochoria – rdest ptasi, orzech ziemny, trzcina pospolita – długie organy wegetatywne
powodują rozprzestrzenienie nasion na kilka metrów, dotyczy to również roślin
geokarpicznych (nasiona dojrzewają pod ziemią)
ballochoria – niecierpek pospolity, tryskawiec – mechanizm eksplozyjny warunkuje rozrzut
nasion, rozrzut w granicach 3-16m.
barochoria – pod wpływem działania grawitacji rośliny zrzucają organy rozmnażania
o owoce lub nasiona – kasztan, dąb
o komórki wegetatywne – lilia, rdest
o zarodniki – mszaki, porosty
herpochoria – na skutek ruchów pełzających dochodzi do rozprzestrzeniania nasion – iglica,
szczotlicha, ostnica – nasiona są śrubowato wygięte, powoduje to samorzutne wkręcanie się
w glebę
Fazy kiełkowania
imbibicja – wzrost oddychania, bardzo duży pobór wody, pęcznienie
faza kataboliczna – mobilizacja substancji zapasowych, synteza fitohormonów, pękanie
łupiny nasiennej
faza anaboliczna – rozwój liści, fotosynteza, produkcja
Czynniki kiełkowania
światło – u roślin fotoblastycznych z fotoblastia dodatnią udział światła jest niezbędny do
zainicjowania procesu kiełkowania. Tylko 5% nasion nie jest wrażliwe na światło w procesie
kiełkowania, jest to więc fotoblastia ujemna (len, zboża, motylkowe)
fitochrom – jest receptorem światła niebieskiego – dla koneserów polecam wykłady z
Sygnalizacji
cytokiny – stymulują kiełkowanie
gibereliny – stymulują kiełkowanie
kwas abscysynowy – hamuje kiełkowanie
temperatura
woda
tlen
Rodzaje kiełkowania
epigeiczne – nadziemne – pojawia się korzeń zarodkowy, wydłuża się hypokotyl wynosząc
liścienie ponad ziemię
hipogeiczne – podziemne – pojawia się korzeń zarodkowy, wydłuża się epikotyl, liścienie
pozostają pod ziemią

28
WYKŁAD 6
Tworzenie się zarodka
bielmo rozwija się prawidłowo – powstaje bielmo jądrowe, następnie bielmo celularne
bielmo rozwija się nieprawidłowo – nie tworzy się bielmo komórkowe
Rozwój
początkowy podział
o prazarodek 2 komórkowy (komórka bazalna + apikalna)
o zarodek 4 komórkowy (kwadrant)
o zarodek 8 komórkowy (oktant)
o itd.
kolejnym stadium jest stadium globularne, zawieszone jest na suspensorze, które zakończone
jest tzw. haustorium (odpowiada za pobieranie pokarmu z otaczających tkanek)
u Arabidopsis początkowo powstaje zygota, następnie dzieli się na komórkę bazalną i
apikalną, tworzy się kwadrant, następnie stadium złożone z 6-12 komórek, kolejne stadium
globuli, w wyniku tych procesów z komórek powierzchniowych powstaje dermatogen
(skórka), a kolejnym stadium jest stadium torpedy
Suspensor
syntetyzuje hormony – auksyny i gibereliny
stanowi drogę transferową dla substancji z bielma lub innych tkanek (np.: obielma) do
zarodka
u większości gatunków zarodek zatopiony jest w bielmie
Budowa zarodka po stadium globuli
komórki inicjujące powstanie czapeczki (hypophysis)
komórki inicjujące korę pierwotną korzenia
dermatogen (praskórka)
peryblem (miękisz)
plerom (walec osiowy)
Typy embriogenezy
onagrad – komórka bazalna + komórka apikalna tworzą zarodek (Onagraceae, Crucifereae,
Ramunculaceae, Liliaceae)
astegrad – duża część zarodka powstaje z komórki bazalnej, reszta z komórki apikalnej
(Asteraceae, Vitaceae)
solarad – z komórki bazalnej powstaje tylko suspensor (Solanaceae, Papaveraceae,
Lineaceae)
henopodiad– komórka bazalna i komórka apikalna w równym stopniu tworzą zarodek
(Chenopodiaceae)
cariophylad – suspensor powstaje tylko z komórki apikalnej (Caryophylaceae)
piperad – podział zygoty może być skośny (nieliczne rodziny)
Schemat podziałów
tworzy się komórka bazalna i komórka apikalna
z komórki bazalnej powstaje suspensor, z komórki apikalnej dwukomórkowy zarodek
tworzy się kwadrant
tworzy się oktant
tworzy się stadium globularne (wytworzenie prakutikuli i domeny centralnej) o symetrii
promienistej

29
tworzy się stadium sercowate, ilość podziałów zostaje zmniejszona, dominuje rozrost
komórek, pojawia się symetria dwuboczna, tworzy się oś pędowo-korzeniowa
powstaje epikotyl, liścienie, stożek wzrostu łodygi, hypophysis (korzeń + czapeczka)
tworzy się stadium torpedy
Hormony embriogenezy
IAA - kwas indoilo-3-octowy – auksyna
o odpowiada za I etap embriogenezy
o warunkuje polarność zarodka
o ułatwia wzrost komórek
o umożliwia szybki wzrost ścian komórkowych
o bierze udział we wszystkich typach organogenezy
o umożliwia zjawisko dominacji wierzchołkowej
ABA – kwas abscysynowy
o odpowiada za środkowy etap embriogenezy
o wysoki poziom ABA w komórkach powoduje zahamowanie podziałów komórkowych i
rozpoczęcie procesów wzrostowych
o powoduje inicjację metabolizmu związanego syntezą i kumulowaniem materiału
zapasowego
o powoduje stopniowe odwodnienie zwiększając tolerancję protoplastu na suszę
o wprowadza nasienie w stan spoczynku
Embriogeneza, a regulacja genomu
komórki apikalne zawierają dużo rybosomów, gęstą cytoplazmę i słabo rozbudowane ER
suspensor zawiera mało rybosomów, rzadką cytoplazmę i rozbudowane SER
udział hormonów w tym procesie jest ważny, ale nie do końca poznany
w regulację włączają się czynniki transkrypcyjne i epigenetyka
wszystkie wymienione czynniki prowadzą do histogenezy
auksyny odpowiadają za różnicowanie zarodka, są jedynymi hormonami transportowanymi u
rośliny polarnie, dopiero w zarodku wnika pomiędzy poszczególne komórki, wynika to z
polarności błony, a więc różnicy w rozlokowaniu transporterów (dokomórkowe i
pozakomórkowe)
auksyny aktywują auksynozależne czynniki transkrypcyjne
inhibitory czynników transkrypcyjnych łączą się z auksynami i zostają naznaczone do
degradacji w proteasomach 26s
zróżnicowanie stężenia auksyn prowadzi do powstania lokalnych gradientów stężenia
Embriogeneza roślin, a zwierząt – różnice
u zwierząt mają miejsce ruchy morfogenetyczne
u roślin ściana uniemożliwia swobodną migrację komórek
kinazy map biorą u roślin udział w reorganizacji cytoszkieletu warunkując rozwój wrzeciona
podziałowego
Wyszukiwarka
Podobne podstrony:
kolokwium embrio-rośliny i zwierzęta-poprawa, studia-biologia, Licencjat, sem 5-6, embriologia-biolo
Embriologia roślin
ROS wykorzystanie roslin do unieszkodliwiania osadow
ROŚLINY ZAWSZE ZIELONE
embriologia rozrodczy meski
Znaczenie liści dla roślin
83 rośliny, mchy, widłaki, skrzypy, okryto i nagonasienne
rosliny GMO
Wykład8 morfogeneza roślin
TECHNIKA OCHRONY ROŚLIN
skrócony Wzrost i rozwój roślin
Metody pozyskiwania, konserwacji i przechowywania surowców roślinnych
Rośliny transgeniczne
komórka roślinna i tkanki roślinne
RoslinyModelowe
więcej podobnych podstron