
1
EWOLUCJONIZM
Dr Tadeusz Namiotko
Wykład 1.
Program wykładów:
1. Definicja ewolucjonizmu i zarys historii myśli ewolucyjnej;
2. Czynniki kształtujące częstość genów w populacjach;
3. Mechanizmy izolujące i modele specjacji;
4. Ograniczenia ewolucji i ich przekraczanie- ewolucja ponadgatunkowa;
5. Filogeneza a systematyka;
6. Ewolucyjne podstawy zachowań zwierząt.
Ewolucja biologiczna- (łac. evolvere- rozwijać), narastająca w linii czasu odmienność pokoleń
potomków obserwowana na dowolnym poziomie organizacji.
Teoria ewolucji jest podstawą teoretyczną współczesnej biologii, najszerzej integrującą jej
działy.
Teoria o bardzo szerokiej podbudowie empirycznej, korzysta z danych wszystkich dyscyplin
biologicznych.
Na teorię ewolucji składają się liczne hipotezy, które mogą być falsyfikowane poprzez
eksperymenty i obserwacje.
Odrzucenie niektórych hipotez lub zastąpienie ich innymi nie prowadzi do odrzucenia całej
teorii.
Nie zaproponowano jak dotąd takiej teorii alternatywnej, która lepiej tłumaczyłaby
zróżnicowanie świata ożywionego.
Historia myśli ewolucyjnej
Starożytne korzenie współczesnej biologii ewolucyjnej.
Próby wprowadzenia systematyki (zaprowadzenia ładu w obserwowanej różnorodności organizmów)-
podstawa późniejszej biologii ewolucyjnej.
Arystoteles
Teokrast
Karol Linneusz
Systemy sztuczne, kryteria podziału ustalone arbitralnie
Najważniejsze nurty starożytne istotne z punktu widzenia rozwoju teorii ewolucji
HYLOIZM- powszechność zjawiska życia
Cały świat jest ożywiony (skały, kamienie, a nawet ciała kosmiczne), a wszystko w przyrodzie
jest ze sobą powiązane, niczym w jednym wielkim organizmie;
Cykliczność i odwieczność Kosmosu- Ziemia i Kosmos są wieczne, a ich dzieje to nieustannie
powtarzająca się sekwencja takich samych zdarzeń.
Drabina jestestw Arystotelesa (Scala naturae)- wszystkie organizmy, które istnieją od początku
stworzenia świata, dają się uszeregować w nieprzerwalny ciąg o rosnącej doskonałości budowy
(człowiek jest najdoskonalszą i najmłodszą z form).
Wszystkie z tych starożytnych motywów odcisnęły swe piętno na rodzącym się ewolucjonizmie
dopiero na początku XIX w. ze względu na panujące wcześniej i ówcześnie poglądy kreacjonizmu.
KREACJONIZM- pierwotny pogląd we wszystkich kręgach kulturowych. Świat został stworzony w
wyniku zamierzonego aktu Stwórcy.
Uczelnia bli
żej Ciebie

2
W teologii chrześcijańskiej wypływa z filozofii św. Augustyna z Hippony (354- 430 n.e.):
• Świat został stworzony przez Boga z niczego ( creatio ex nihilo) w jednorazowym
akcie stworzenia;
• Stwórca ciągle podtrzymuje istnienie świata (creatio continua) i kieruje nim ku dobru;
• Prawa rządzące światem można poznać jedynie przez nadprzyrodzony akt stworzenia
(illuminatio).
Zmodyfikowany w chrześcijaństwie przez św. Tomasza z Akwinu (1225-1274):
• Podstawą poznania jest empiryzm i dowód logiczny;
• Jednorazowy akt kreacji, w którym świat został ukształtowany od razu w obecnej
postaci;
• Niezmienność gatunków i ich liczby w czasie oraz całkowita negacja zmian
ewolucyjnych wśród organizmów ( szczególnie powstawania jednego gatunku z
drugiego).
PRZED DARWINEM
LAMARKIZM
Jean Babtiste Antoine Pierre de Monet de Lamarck- (1744-1829), pierwszy nowożytny
ewolucjonista;
Różnorodność organizmów jest wynikiem cząstkowych modyfikacji i odstępstw od
podstawowego planu budowy;
Drobne początkowo modyfikacje powodują z czasem stopniowe transformacje jednych
organizmów w drugie, od form najprostszych do najbardziej złożonych („drabina jestestw
organicznych”- zmiany jednokierunkowe bez rozgałęziania);
Transformacje powstają na skutek oddziaływań środowiska, wywołujących zmiany w
organizmie, który musi dostosować się do nowych warunków;
Nabyte modyfikacje są przekazywane potomstwu i stopniowo ulegają utrwaleniu;
Intensywne używanie narządu powoduje jego rozwój, nieużywanie zaś- redukcję bądź
całkowity zanik;
Cechy nabyte przez organizm w ciągu życia są przekazywane potomstwu.
Mechanizm powstawania zmian w organizmie:
• Rośliny i zwierzęta niższe („bez czucia”)- środowisko oddziałuje bezpośrednio;
• Zwierzęta wyższe (posiadające „czucie wewnętrzne”)- zmieniają się w sposób celowy
pod wpływem „wewnętrznej potrzeby doskonalenia się” (fluid przekazujący sygnały
do narządu docelowego)
Drabina jestestw- skoro organizmy nieuchronnie się doskonalą i przesuwają po szczeblach, to
najprostsze organizmy występujące współcześnie musiały powstać stosunkowo niedawno ( na drodze
samorództwa).
Poglądy na zmienność organizmów głosili w tym czasie również:
• Georges Buffor
• Erasmus Darwin
• Johann Wolfgang Goethe
• Jędrzej Śniadecki
• Linneusz
KATASTROFIZM
Georges Cuvier (1769- 1832)
Krytyka poglądów ewolucyjnych i powrót do linneuszowskiej koncepcji stałości gatunku i
idei kreacjonizmu;
W historii Ziemi zdarzyło się kilka katastrof, które całkowicie zniszczyły żywe organizmy;
Po każdym kataklizmie organizmy pojawiały się na nowo w doskonalszej formie. Po ostatniej
katastrofie (5-6 tys. lat temu) pojawił się człowiek w swej obecnej postaci;

3
Nie istnieje jednolity plan budowy organizmów, a raczej cztery niezależne, zgodne z
założeniami aktu stworzenia: Vertebrata (kręgowce), Mollusca (mięczaki, osłonice i
ramienionogi), Articulata (stawonogi i pierścienice), Radiata (jamochłony i cała reszta).
Charles Robert Darwin (1809- 1882)
1825- 1831- studia
Studiował początkowo medycynę, później teologię przygotowując się do stanu duchownego, zgłębiał
też botanikę, zoologię i geologię na uniwersytetach w Edynburgu i Cambridge.
1831-1836- podróż dookoła świata na „HMS Beagle”
Kopalne szczątki szczerbaków, mastodontów i małży w Ameryce Południowej
Różnice między endemicznymi gatunkami na Wyspach Galapagos
Bogate zbiory zoologiczne, botaniczne i geologiczne
Teoria powstania wysp koralowych
1836-1841- pobyt w Londynie
Opis podróży, kontakty z wieloma znanymi uczonymi ( m.in.: Ch. Lyell)
1842- 1882- posiadłość w Down
1842 (1844)- streszczenie poglądów na powstawanie gatunków;
1858- krótkie doniesienia wraz z pracą A.R.Wallace’a („O dążności odmian do nieograniczonego
odbiegania od typu pierwotnego”)
1859- „O powstawaniu gatunku drogą doboru naturalnego, czyli o utrzymaniu się doskonalszych ras w
walce o byt”
• Badał pnącza, ruchy roślin, rośliny owadożerne, polimorfizm kwiatów, owadopylność
storczyków i skutki samozapylenia i zapylenia krzyżowego;
• Obserwował prawidłowości dziedziczenia w doświadczeniach z wyżlinem i pierwiosnkami
(dopuszczał dziedziczenie cech nabytych).
Inne ważne dzieła:
1868- „O zmienności roślin i zwierząt w stanie udomowionym”
1871- „O pochodzeniu człowieka i doborze płciowym”
1872- „O wyrazie uczuć u człowieka i zwierząt”
TEORIA DARWINA- WALLACE’A
1. W obrębie gatunku występuje zmienność osobnicza ( zmienność ma charakter losowy, różnice
mogą być korzystne lub nie);
2. Zmienność jest dziedziczna (pangeneza- gemule produkowane we wszystkich komórkach
ciała przechodzą do gamet i mogą odtworzyć wszystkie cechy całego organizmu, również te
nabyte za życia);
3. Liczba osobników młodych w każdym pokoleniu przewyższa liczbę osobników pokolenia
rodziców i pojemność środowiska;
4. Dysproporcja między liczbą rodzących się i przeżywających osobników stwarza konkurencję,
przejawiającą się w postaci walki o byt;
5. Największe szanse przeżycia mają osobniki najlepiej dostosowane do danych warunków
(środowisko prowadzi naturalną selekcję);
6. Dostosowanie gatunków stale rośnie- korzystne cechy się kumulują, gdyż tylko przeżywające
osobniki przekazują swoje cechy potomstwu. Gatunek nie jest statyczny- zmienia się w czasie;
7. Na różne grupy osobników w obrębie gatunku mogą działać różne czynniki, prowadząc do
powstania odmian, ras, podgatunków i odrębnych gatunków. Ze względu na losowy charakter
zmienności i zróżnicowanie nacisków selekcyjnych, graficzny obraz zmian ewolucyjnych
przypomina raczej rozgałęzione drzewo rodowe, a nie linię, czy drabinę;
8. Taksony wyższego rzędu powstają w ten sam sposób co gatunki, poprzez stopniowe
pogłębianie się różnic w czasie;
9. Podział gatunków na formy różniące się sposobem życia zwiększa pojemność środowiska;
10. Istnieje tendencja do stałego wzrostu liczby gatunków.
„Jakże jesteśmy głupi, że sami na to nie wpadliśmy.”- Thomas Huxley, 1859
„Módlmy się, żeby to nie była prawda, ale jeśli to jest prawda, prośmy Boga, żeby się nie rozniosło”

4
„Niczego w biologii nie da się zrozumieć bez odwołania do teorii ewolucji”- Theodosius Dobzhansky,
1859
„Teoria ewolucji to kłamstwo! To luźna koncepcja niewierzącego starszego pana, który tak widział
świat. Może dlatego ,że był wegetarianinem i zabrakło mu ognia wewnętrznego”
SYNTETYCZNA TEORIA EWOLUCJI
J.B.S. Haldane (1892- 1964)
Ronald A. Fisher (1890- 1960)
Julian Huxley (1887- 1975)
T. Dobzhansky (1900- 1975)
Sewall Wright (1889- 1988)
G.G. Simpson (1902- 1984)
G. Ledyard Stebbins (1906- 2000)
Ernst Mayr (1904- 2005)
1. Podstawą procesów ewolucyjnych jest losowa zmiennośc mutacyjna;
2. Zmienność rekombinacyjna wpływa na wzrost i utrzymywanie dużego zróżnicowania
genetycznego w populacjach;
3. Różne genotypy (mutacje i układy rekombinacyjne) mają różną wartość przystosowawczą do
danych warunków środowiskowych;
4. Środowisko naturalne powoduje eliminowanie osobników o cechach niekorzystnych- to
znaczy o relatywnie niższej rozrodczości i wyższej śmiertelności (dobór naturalny);
5. Dobór naturalny zmienia strukturę genetyczną populacji- geny determinujące cechy korzystne
zwiększają swoją frekwencję w populacji (nosiciele tych genów pozostawiają po sobie więcej
potomstwa), geny odpowiedzialne za cechy niekorzystne są eliminowane;
6. Zróżnicowane działania doboru prowadzi do powstawania różnic genetycznych między
populacjami;
7. Na skutek izolacji przepływu genów między populacjami różnice między nimi mogą się
pogłębiać z czasem aż do powstawania odrębnych gatunków (specjacja);
8. Wyższe taksony powstają na skutek kumulacji różnic genetycznych między grupami
organizmów w dłuższych zazwyczaj przedziałach czasowych.
Ewolucja zatem to proces zachodzący na poziomie populacyjnym, polegający na przebudowie
genetycznej populacji.
Podstawowa zmiana ewolucyjna to zmiana częstości alleli dowolnego genu w puli genowej populacji.
CZYNNIKI KSZTAŁTUJĄCE CZĘSTOŚĆ GENÓW W POPULACJACH
Struktura genetyczna populacji- określenie frekwencji genotypów i genów
Frekwencje genotypów
Załóżmy:
40 AA 47Aa 13aa = 100 osobników
N
AA
+ N
Aa
+ N
aa
= N (liczebność ogólna)
p
AA
= 40/100= 0,40 N
AA
/N
p
Aa
= 47/100= 0,47 N
Aa
/N
p
aa
= 13/100= 0,13 N
aa
/N
p
AA
+ p
Aa
+ p
aa
= 1
Frekwencje genów (alleli)
5
AA Aa aa ogółem
Liczba osobników
40
47 13
100
Liczba alleli A
80
47
0
127
Liczba alleli a
0
47 26
73
Ogólna liczba alleli
200
Frekwencja allela A
(p) = 127/200 = 0,635
p = (2 N
AA
+ N
Aa
)/ 2N
frekwencja allela a
(q) = 73/200 = 0,365
q = (2 N
aa
+ N
Aa
)/ 2N
p + q = 1 => q = 1 - p
Jak określić frekwencje alleli na podstawie frekwencji genotypów?
p= (2 N
AA
+ N
Aa
)/ 2N = (2 N
AA
/ 2N) + (N
Aa
/ 2N) = p
AA
+ 1/2 p
Aa
analogicznie:
q= (2 N
aa
+ N
Aa
)/ 2N = (2 N
aa
/ 2N) + (N
Aa
/ 2N) = p
aa
+ 1/2 p
Aa
Częstość kojarzeń w populacji panmiktycznej ( o losowym kojarzeniu)
kojarzenie
frekwencje
potomstwo
AA
Aa
aa
A×A
0,635×0,635 0,403
A×a
0,635×0,365
0,232
a×A
0,365×0,635
0,232
a×a
0,365×0,365
0,133
0,403 0,464 0,133
Wykład 2.
Struktura genetyczna populacji (c.d.)
kojarzenie frekwencje
potomstwo
AA
Aa
aa
A×A
p
2
1
A×a
pq
1
a×A
qp
1
a×a
q
2
1
p
2
2pq q
2
kojarzenie frekwencje kojarzeń
potomstwo
AA
Aa
aa
6
AA×AA
p
AA
2
1
AA×Aa
2p
AA
p
Aa *
½
½
AA×aa
2p
AA
p
aa *
1
Aa×Aa
p
Aa
2
¼
½
¼
Aa×aa
2p
Aa
p
aa *
½
½
aa×aa
p
aa
2
1
p
2
2pq q
2
*- 2 jest dlatego, że zarówno matka może być Aa, ojciec AA, jak i matka AA , ojciec Aa
║
▼
p
AA
2
+ ½ × 2p
AA
p
Aa
+ ¼ p
Aa
2
= ( p
AA
+ ½ p
Aa
)
2
= p
2
A (o frekwencji p) a (o frekwencji q=1-p)
A
AA (p
2
)
Aa (pq)
a
Aa (pq)
aa (q
2
)
CZYNNIKI KSZTAŁTUJĄCE CZĘSTOŚĆ GENÓW W POPULACJACH
PRAWO HARDY’EGO- WEINBERGA
Prawidłowości opisane w 1908 r., niezależnie przez angielskiego matematyka G.H. Hardy’ego (1877-
1947) i niemieckiego przyrodnika W. Weiberga (1862-1937).
Jeśli spełnione są określone założenia, to:
Frekwencje alleli w populacjach nie zmieniają się z pokolenia na pokolenie;
Frekwencje genotypów zależą jedynie od frekwencji alleli, ustalają się w następnym
pokoleniu i przy dwóch allelach w jednym locus autosomalnym wynoszą odpowiednio:
p
2
- dla homozygot dominujących
2pq- dla heterozygot
q
2
- dla homozygot recesywnych
1. Ograniczenie do dwóch alleli- uproszczenie
Jeśli alleli więcej niż dwa, to:
- rozpatrujemy jeden z nich, a wszystkie pozostałe traktujemy jako drugi alternatywny
- posługujemy się modelami dla większej liczby alleli ( np. dla trzech alleli frekwencje sześciu
możliwych genotypów w stanie równowagi określone są przez rozwinięcie trójmianu do
kwadratu).
2. Ograniczenie do jednego locus- uproszczenie
Modele dla większej liczby loci powodują większe komplikacje.
3. Ograniczenie do genów autosomalnych- uproszczenie
Frekwencje genów autosomalnych kształtują się nieco inaczej niż tych umieszczonych w
autosomach, co komplikuje modele.
Założenia prawa Hardy’ego – Weinberga
1. organizm diploidalny rozmnażający się płciowo
2. pokolenia nie zachodzą na siebie
Prawie wszystkie modele genetyki populacji uwzględniają to założenie. Jeśli frekwencje alleli
zmieniają się, to nowy stan równowagi może być osiągnięty dopiero, gdy wymże ostatni
osobnik poprzedniego pokolenia.
3. kojarzenie się osobników jest losowe (panmiksja)
Skutki odrzucenia założenia panmiksji opisują modele kojarzenia w pokrewieństwie
(INBRED)
4. populacja jest liczna (co najmniej kilkaset osobników)
7
Duża liczebność populacji zmniejsza wpływ przypadku na kształtowanie się frekwencji alleli.
Skutki odrzucenia tego założenia opisują modele DRYFU GENETYCZNEGO.
5. nie istnieją mutacje
Skutki odrzucenia założenia braku MUTACJI przedstawiają osobne modele genetyczno-
populacyjne.
6. nie występują migracje
Populacyjne skutki przepływu genów opisują modele MIGRACJI.
7. nie działa dobór naturalny
Ewolucyjne konsekwencje zróżnicowanego przeżywania i rozrodczości opisują liczne modele
DOBORU.
Jeżeli efekty mutacji, migracji i doboru równoważą się, to populacja także może pozostawać w stanie
równowagi genetycznej.
Prawo Hardy’ego- Weinberga stanowi podstawę teoretyczną, na której oparto większość modeli
genetyki populacyjnej, dlatego jego zasady powinny być w pełni zrozumiałe przed studiowaniem
innych modeli, które są w zasadzie rozszerzeniem tego prawa i powstały w wyniku uwalniania się od
jego założeń.
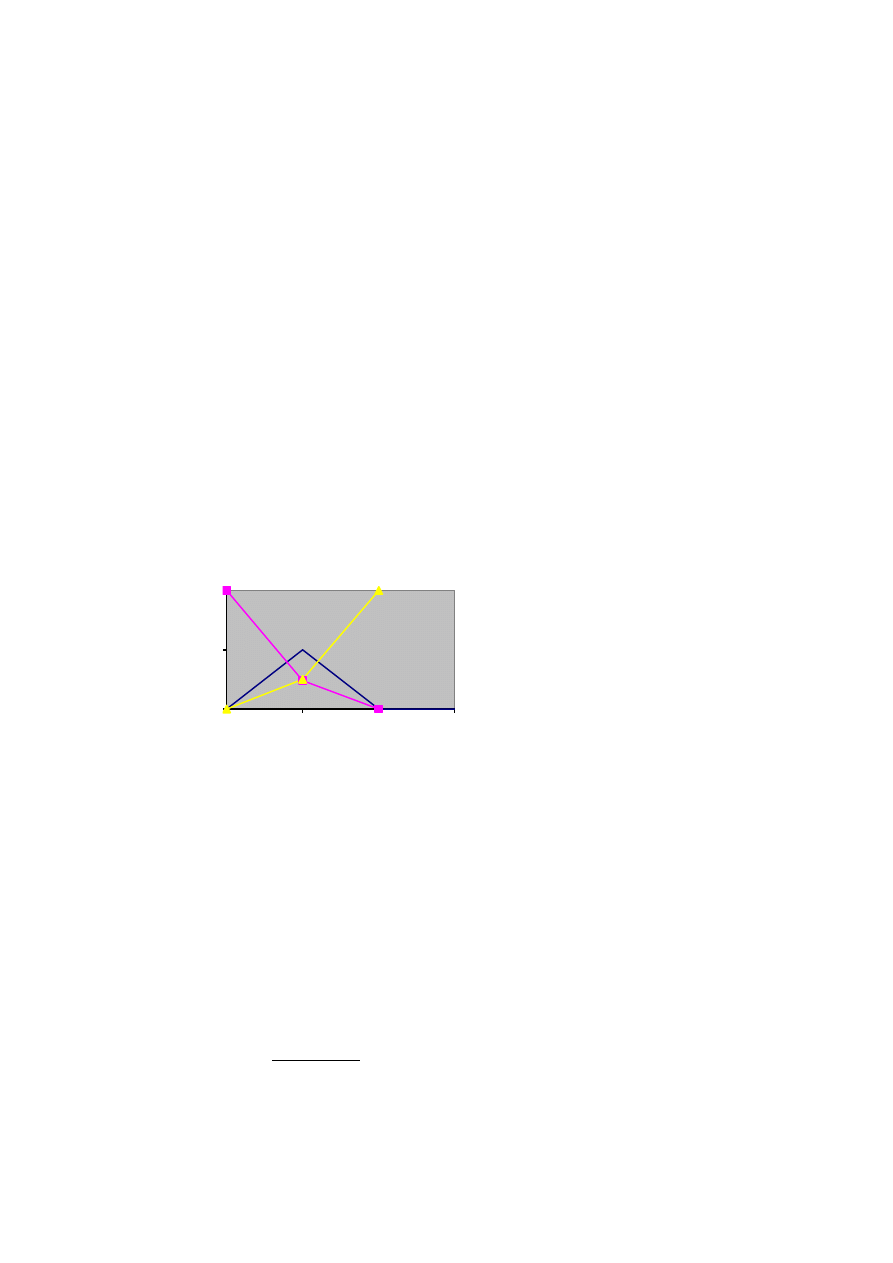
Częstość genotypów AA, Aa i aa jako funkcja częstości allela A(p)
Dla każdego p częstości genotypów sumują się do jedności, a frekwencja heterozygot nie przekracza
50% i jest najwyższa gdy p= q= 0,5.
0
0,5
1
0
0,5
1
frekwencja allela A (p)
fre
kwe
ncja
genot
ypów
Zadanie:
Frekwencja allela dominującego 0,2 (p)
Frekwencja allela recesywnego 0,8 (q)
q
2
= 0,64 – frekwencja homozygot recesywnych
Przykład:
Krasopani poziomkówka Callimorpha dominula (= Panaxia dominula)
(niedźwiedziówki Arctidae)
AA- jasne przednie skrzydła (dużo białych plam)
AA’- średnio białych plam
A’A’- mało białych plam (po dwie na skrzydle)
E.B. Ford 1939- 1970: 17062 AA
1295 AA’
28 A’A’
18385
Frekwencja allela A:
p= [(2×17062)+1295]/(2×18385)=0,963
frekwencja allela A’:
8
q= 1-0,963
Oczekiwane frekwencje genotypów Frekwencje obserwowane Liczebności oczekiwane
p
2
=(0,963)
2
= 0,9274
17062/18385= 0,9280
18385×0,9274= 17050
2pq= 2(0,963)(0,037)= 0,0713
1295/18385= 0,0704
18385×0,0713= 1311
q
2
=(0,037)
2
= 0,0013
28/18385= 0,0015
18385×0,0013= 24
Brak statystycznie istotnych różnic.
W rzeczywistości w ciągu 32 lat badań p wzrosło od 0,91 do 0,97
1955
1956
Genotyp
AA
AA’
A’A’
AA
AA’ A’A’
Obserwowana liczebność
308
7
0
1231
76
1
Obserwowane frekwencje genotypów 0,978 0,022 0,000
0,941
0,058 0,001
Frekwencje alleli
p= 0,989 q= 0,011
p= 0,970 q= 0,030
Oczekiwane frekwencje genotypów
0,978 0,022 0,0001
0,941
0,058 0,001
Oczekiwana liczebność
308,1
6,9
0,03
1230,8
75,9
1,31
Porównanie lat 1955 i 1956- brak statystycznie istotnych różnic między obserwowanymi i
oczekiwanymi frekwencjami alleli i genotypów w poszczególnych latach, ale istotne różnice między
porównywanymi latami.
Zachowując taki trend zmian, po 52 latach allel A niemalże zniknąłby z populacji.
Podział czynników kształtujących frekwencje genów i genotypów w populacjach
1. efekty systematyczne (można przewidzieć zarówno kierunek, jak i wielkość zmian)
a) selekcja (= dobór)
b) migracje
c) mutacje
d) dryf mejotyczny
2. efekty dyspersyjne (można przewidzieć wielkość zmian, ale kierunek jest nieprzewidywalny)
a) dryf genetyczny
b) inbred
3. zdarzenia nieokresowe (nie można przewidzieć ani kierunku, ani wielkości zmiany)
a) efekt założyciela
b) efekt szyjki butelki (wąskiego gardła liczebności)
Dryf genetyczny
Definicja- nieregularne (losowe) fluktuacje frekwencji alleli zachodzące z pokolenia na pokolenie,
ujawniające się najsilniej podczas stale lub okresowo ograniczonej liczebności.
Pokolenie 1:
RR, Rr, RR, rr, Rr, RR, RR, RR, Rr, Rr
p =0,7 (frekwencja R)
tylko 5 pozostawia potomstwo!
q =0,3 (frekwencja r)
Pokolenie 2:
rr, RR, Rr, RR, rr, Rr, RR, rr, Rr, Rr
p =0,5
tylko 2 pozostawiają potomstwo!
q = 0,5
Pokolenie 3:
RR (wszystkie)
p =1,0
q =1,0
9
Przykład hipotetyczny:
Populacja 4 osobników:
1 homozygota AA
2 heterozygoty Aa
p = q = 0,5
1 homozygota aa
Zalożenia: nie działa dobór, migracje, mutacje i wielkość populacji ta sama z pokolenia na pokolenie
Pokolenie rodzicielskie: 1AA 2Aa 1aa
Gamety: A A a a
Możliwości i częstości połączenia gamet:
A
a
A AA (p
2
) Aa (pq)
a
Aa (pq) Aa (q
2
)
Pierwsze pokolenie potomne:
( możliwe kombinacje genotypów przy zachowanej wielkości populacji N=4)
AA AA AA AA AA AA aa
AA AA AA Aa Aa aa aa
AA AA Aa Aa aa aa aa
AA Aa aa aa aa aa aa
Frekwencja allela recesywnego q
1
:
0 0,125 0,375 0,5 0,625 0,75 1
Na pytanie, jak zmieni się q nie potrafimy dokładnie odpowiedzieć i nie potrafimy tej zmiany
przewidzieć, gdyż wynika ona ze zjawisk losowych.
Symulacje dryfu genetycznego
Im mniej osobników w populacji, tym wpływ przypadku jest większy!
Prawdopodobieństwo wystąpienia określonej frekwencji q (czyli liczby alleli a=k) w następnym
pokoleniu liczącym 4 osobniki (czyli 8 alleli= N), przy początkowej frekwencji q= 0,5 odczytać
można z rozkładu dwumianowego.
P
(q, N, k)
= [N
!
/ (N- k)
!
k
!
][ p
k
(1- p)
N-k
]
gdzie: N- całkowita liczba alleli (tu= 8)
k- liczba alleli recesywnych
q- początkowa frekwencja allela recesywnego (tu= 0,5)
Ponieważ w przykładzie p= q= 0,5 natomiast N= 8, wartość [ p
k
(1- p)
N-k
] jest stała i wynosi 0,5
8
=
0,0039.
Przy możliwych 9 kombinacjach alleli w następnym pokoleniu:
8 alleli A 7 6 5 4 … 0
0 alleli a
1 2 3 4 … 8
I odpowiednich frekwencjach allela recesywnego q
i
:
0 0,125 0,25 0,375 0,5 … 1
Prawdopodobieństwo wystąpienia poszczególnych kombinacji wynosi:
0,00391 0,03125 0,10938 0,21875 0,27344 … 0,00391

10
Gdy N wzrasta, rozkład binominalny zbliża się do rozkładu ciągłego i może zostać zastąpiony przez
rozkład normalny opisywany przez średnią i wariancję.
Frekwencja allela recesywnego w następnym pokoleniu może zostać wtedy określona wzorem:
q
1
= q
0
±√[ q
0
( 1 - q
0
)/2N]
odchylenie standardowe
N= 10, a q
0
= 0,5 => q
1
= 0,5± 0,11( 0,39- 0,61)
zakres zmian
N= 10000, a q
0
= 0,5 => q
1
= 0,5± 0,0035
Z reguł statystycznych wynika, że prawdopodobieństwo uzyskania:
q
1
= q
0
± SD (jedno odchylenie standardowe) wynosi 68%, natomiast:
q
1
= q
0
± 2SD= 95%
Przy stałym utrzymywaniu się dryfu, w kolejnych pokoleniach rozkład frekwencji alleli odbiega od
rozkładu normalnego, zaś wariancja rośnie w każdym pokoleniu tym szybciej, im mniejsze N.
Wykład 3.
Średnia liczba pokoleń do czasu zniknięcia allela dana jest wzorem:
ñ= -(4N/ 1- q
0
) q
0
ln q
0
q
0
= 0,5 => n zniknięcia allela a = 2,773N
czyli np. średnio ok. 28 pokoleń dla N= 10, a 277 pokoleń dla N= 100
q
0
= 0,1 => n zniknięcia allela a = 1,023N
czyli np. średnio ok. 10 pokoleń dla N= 10, a 102 pokolenia dla N= 100
q= 0,1 => n utrwalenia lub zniknięcia allela a:
n= 0,1× 3,793N + 0,9× 1,023N= 1,295N
Jeżeli allele występują w różnej frekwencji, tak że frekwencja jednego z nich jest bliska 0, utrata
któregokolwiek z nich trwa krócej, niż gdy q
0
= p
0
= 0,5
DRYF GENETYCZNY- EKSPERYMENT (BURI, 1956)
Drosophila melanogaster
107 populacji
Locus bw:
bw bw- oczy brązowe
bw bw
75
- oczy czerwone
bw
75
bw
75
- oczy jasnobrązowe
początkowa frekwencja bw
75
= 0,5 (16 heterozygot)
stała wielkość populacji N= 16
za każdym razem losowo 8♀♀ i 8♂♂ jako rodzice następnego pokolenia
określenie genotypów następnego pokolenia i frekwencji alleli
19 pokoleń
W 19 pokoleniu:
11
28 populacji q= 0
30 populacji q= 1
Średnia wartość q we wszystkich populacjach ok. 0,5
Konsekwencje dryfu genetycznego:
1. powstawanie różnic genetycznych między subpopulacjami
2. zmniejszenie się zmienności genetycznej w obrębie subpopulacji
3. wzrost częstości homozygot kosztem heterozygot
Przykład 1
Zmiana frekwencji alleli locus MN (allele L
M
i L
N
) w sekcie religijnej Dunkersów
Sekta powstała w XVIII w. w Niemczech
27 par założyło grupę wyznaniową w USA
Nie zawierali małżeństw z przedstawicielami innych wyznań/ religii
Grupa wiekowa Frekwencja L
M
3- 27 lat
0,73
28- 54 lata
0,66
> 54 lata
0,55
W Niemczech i USA frekwencja L
M
wśród nie- Dunkersów wynosi ok. 0,54.
Wniosek:
W ciągu ok. 200 lat izolacji frekwencja allela L
M
w wyniku zjawisk losowych wzrosła u Dunkersów o
ok. 0,2.
Przykład 2
Frekwencja alleli grup krwi w układzie ABO w plemieniu Czarnych Stóp z południowej Kanady: I
A1
=
0,8
U Indian Ameryki Północnej- prawie 100% grupa krwi O
Efektywna wielkość populacji
1. niejednakowa liczba samic i samców:
Ne = (4Nm×Nf)/ (Nm+ Nf)
gdzie: Nm- liczba samców
Nf- liczba samic
Przykład:
Dla stada złożonego z 200 krów i 2 byków : Ne= 8
2. wielkość populacji zmienia się z pokolenia na pokolenie:
1/Ne= 1/n(1/N
1
+1/ N
2
+…+1/ N
n
)
gdzie: n- liczba pokoleń
Przykład:
Dla populacji o cyklicznych zmianach liczebności N
1
= 10, N
2
= 100, N
3
= 1000 i N
4
= 100, Ne
wynosi 33.
INBRED
Definicja- kojarzenie osobników spokrewnionych ze sobą przez wspólnego przodka
Paradoks genealogiczny:
W populacji biseksualnych organizmów, każdy osobnik ma 2 rodziców, 4 dziadków, 8 pradziadków,
a n pokoleń wstecz- 2
n
przodków. Nawet w niezbyt odległym pokoleniu, liczba potrzebnych przodków
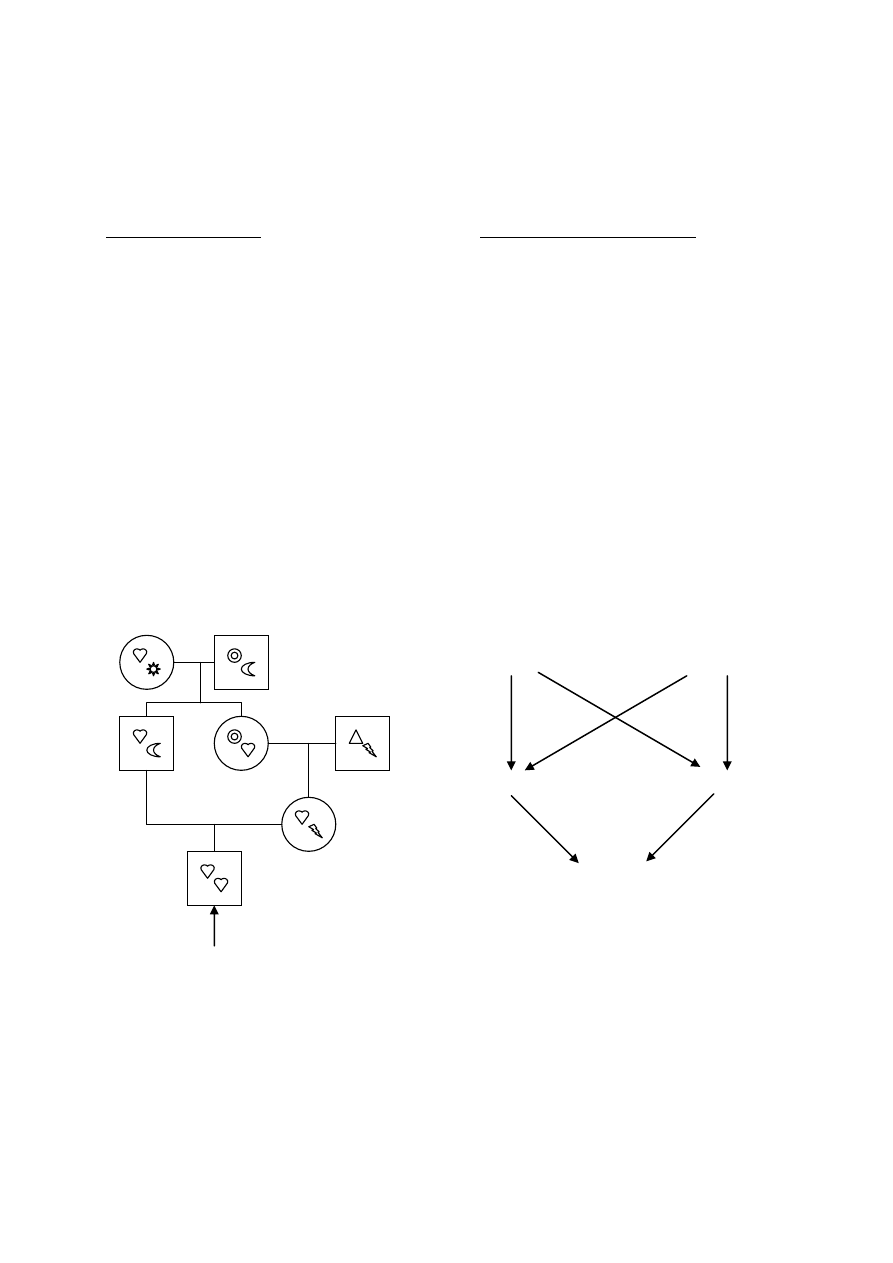
12
dla zabezpieczenia oddzielnych przodków każdemu żyjącemu osobnikowi staje się większa, niż
mogłaby zawierać jakakolwiek faktycznie istniejąca populacja.
Z tego powodu osobniki w populacji muszą być między sobą spokrewnione przez jednego lub więcej,
bliższych lub dalszych przodków. Im w poprzednich pokoleniach mniejsza liczebność populacji, tym
mniej odległe w rodowodach osobniki i większa liczba wspólnych przodków.
Stopień pokrewieństwa między osobnikami w populacji zależy od wielkości tej populacji.
Przykład:
Człowiek: czas trwania jednego pokolenia 25 lat
W ciągu 1000 lat istnienia państwa polskiego było ok. 40 pokoleń
Każdy z nas ma w swej historii 2
40
przodków, czyli ok. 1,1×10
12
(bilion).
Liczba ludności na Ziemi w roku 1000- ok. 1 miliard (10
9
)
Cofając się do początków naszej ery (ok. 80 pokoleń wstecz), każdy z nas miał 2
80
przodków (czyli
1,2×10
24
)
Inbred jest czynnikiem, którego wpływu na ewolucję gatunku nie można lekceważyć.
Osobniki mogą być homozygotami z dwóch powodów:
1. dwa allele danego locus mogą być kopiami tego samego allela pochodzącego od wspólnego
przodka, będą więc identyczne przez wspólne pochodzenie
- AUTOZYGOTYCZNE
2. dwa allele danego locus mogą być identyczne, ale nie dzięki wspólnemu pochodzeniu
-ALLOZYGOTYCZNE
Heterozygoty z definicji są allozygotyczne.
A
1
/ A
2
♂
A
3
/A
4
♀
p= 1/2
A
1
/ A
3
A
1
/ A
4
p= 1/2
A
1
/ A
1
autozygota
autozygota
Przykłady inbredu w rodowodach:
osobnik 1 osobnik 2 osobnik 3
13
A
1
A
1
A
1
A
2
A
3
A
3
a
1
a
1
a
3
a
4
a
5
a
6
osobnik 4
osobnik 5
osobnik O
A
i
oznacza konkretny allel w genotypie
a
1…
a
6
- symbole identyfikujące poszczególne allele i określające ich ewentualną identyczność
przez wspólne pochodzenie ( nie oznaczają genotypów)
Możliwe stany osobnika O:
1. a
3
a
3
autozygota
A
1
A
1
homozygota
2. a
4
a
4
autozygota
A
2
A
2
homozygota
3. a
3
a
4
allozygota
A
1
A
2
heterozygota
4. a
1
a
3
allozygota
A
1
A
1
homozygota
Jak określić prawdopodobieństwo autozygotyczności osobnika O?
Współczynnik inbredu (wsobności) F:
prawdopodobieństwo z jakim dwa allele tego samego locus u jednego osobnika są
autozygotyczne
prawdopodobieństwo, że dany osobnik z populacji jest autozygotą (prawdopodobieństwo, że
dany osobnik jest allozygotą = 1-F)
populacja zinbredowana- to taka, w której prawdopodobieństwo, że dany osobnik jest autozygotą
jest większe wskutek kojarzenia w pokrewieństwie niż wynikałoby to z losowego kojarzenia w
populacji panmiktycznej
F <0,1>
F= 0 => populacja panmiktyczna
F= 1 => populacja całkowicie zinbredowana (np. samozapłodnienie)
Sposób obliczania wartości F na podstawie znanego rodowodu:
osobnik 1
osobnik 2
osobnik 3
osobnik 4
osobnik 5
osobnik O
Ile wynosi F osobnika O?
zaznaczamy na rodowodzie wszystkie ścieżki spokrewnienia poprzez wszystkich wspólnych
przodków- tu osobniki 4-2-5
ustalamy liczbę osobników (n) w każdej ścieżce (nie licząc osobnika O)
obliczamy wartość F dla każdej ścieżki osobno jako (1/2)
n
14
sumujemy wartości obliczone dla każdej ścieżki otrzymując ogólną wartość F
F= (1/2)
3
= 1/8
1/2
= 1/2 1/2
1/2 1/2
F
(I)
= (1/2)
5
=
+
F
(I)
= (1/2)
5
+ (1/2)
5
Współczynnik inbredu- wzór ogólny (Sewall Wright 1922)
F
X
= Σ[(1/2)
n1+ n2+ 1
(1+ F
A
)]
gdzie: F
X
– współczynnik inbredu osobnika X
n
1(2)
- liczba pokoleń od ojca (matki) do wspólnego przodka
F
A
- współczynnik inbredu wspólnego przodka A
genotyp
AA
Aa
aa
Frekwencje w populacji panmiksji F=O
p
2
2pq
q
2
Frekwencje w populacji zinbredowanej (0<F<1) p
2
(1-F)+ pF 2pq(1-F)+ 0 q
2
(1-F)+ qF
Frekwencje przy całkowitym inbredzie (F=1)
p
0
q
Pogrubione- allozygoty
Normalne- autozygoty
Czy inbred zmienia frekwencje genotypów?
Czy p
2
(1-F)+ pF> p
2
?
p
2
(1-F)+ pF = p
2
- p
2
F+pF = p
2
+pF(-p+1) = p
2
+ Fpq
czyli jeżeli q>0 i F>0 to w populacji będzie nadmiar homozygot
Inbred zmienia frekwencje genotypów
Genotypy
AA
Aa
aa
Frekwencje genotypów
x
y
Z

15
samozapłodnienie
AA×AA Aa×Aa aa×aa
Stałe kojarzenie w pokrewieństwie powoduje wzrost autozygotyczności
Np. pszenica (Triticum) samozapłodnienie, prawie wszystkie kwiaty zamknięte
½ allozygot ubywa w każdym pokoleniu
F wzrasta gdy inbred utrzymuje się w populacji przez szereg pokoleń
n→∞ => F→1 (czyli F
e
= 1)
Ostatecznie populacja złożona będzie jedynie z autozygot (homozygot).
Wykład 4.
Czy inbred zmienia frekwencje alleli?
p= P
AA
+ ½ P
Aa
P
AA
= p
2
(1-F) + pF = p
2
+ Fpq P
Aa
= 2pq (1-F)
p= p
2
+ Fpq + ½ [2pq (1-F)] = p
2
+ Fpq + ½ (2pq – 2Fpq) = p
2
+ Fpq + pq – Fpq = p
2
+ pq= p(p+q)= p
Inbred nie zmienia frekwencji alleli
Wartość F może być obliczona:
bezpośrednio, jeśli znamy genealogię osobników obejmującą kilka pokoleń
pośrednio, jeśli znamy frekwencje genotypów i alleli w jednym lub większej liczbie loci
H
E
– heterozygotyczność oczekiwana = 2pq
H
F
– heterozygotyczność obserwowana = 2pq(1-F)
H
F
= H
E
(1-F) => F= (H
E
– H
F
)/ H
E
lub
F= 1- (H
F
/ H
E
)
F wyraża o jaką częstość obniża się heterozygotyczność populacji kojarzącej się systemem
krewniaczym przy zachowaniu frekwencji alleli w stosunku do oczekiwanej przy kojarzeniu losowym.
Heterozygotyczność:
1. frekwencja układów (osobników) heterozygotycznych w populacji (H
L
)
2. frekwencja loci heterozygotycznych na osobnika (H
I
)
Genetyczno- ewolucyjne konsekwencje inbredu:
1. zmiana frekwencji genotypów (wzrost frekwencji homozygot, spadek frekwencji heterozygot)
AA
Aa
aa
Frekwencja genotypów w następnym pokoleniu x+ y/4 y/2 z+ y/4
pokolenie
F
1
Tylko A
1
A
2
0
2
¼ A
1
A
1
, ½ A
1
A
2
, ¼ A
2
A
2
,
0,500
3
¼ A
1
A
1
,1/8 A
1
A
1
, ¼ A
1
A
2
,1/8 A
2
A
2
, 1/4 A
2
A
2
0,750
4
0,875
5
0,938
…
…
n
1
16
2. wzrost wariancji genetycznej w populacji
Jeśli F=0 (populacja zrównoważona) i p= q= 0,5, to fenotyp determinowany przez Aa o
frekwencji 2pq= 0,5 jest równy wartości średniej dla całej populacji, a jedynie homozygoty
odbiegają od średniej wpływają na wariancję
Jeśli F>0, to pojawia się więcej homozygot i wzrasta wariancja (zróżnicowanie genetyczne
populacji)
Przy kompletnej dominacji inbred zmniejsza średnią wartość cechy proporcjonalnie do
wzrostu F
Przy niekompletnej dominacji średnia wartość cechy nie zmienia się
3. depresja inbredowa- obniżenie płodności i przeżywania wskutek wzrostu frekwencji
homozygot recesywnych
4. nierównowaga sprzężeniowa- obniżenie prawdopodobieństwa rekombinacji
depresja inbredowa przy 10% wzroście F:
gatunek
cecha
Depresja inbredowa
kura
Sukces wykluwania
4,36%
Roczna produkcja jaj
9,26 jaj
człowiek Wysokość w wieku 10 lat 2,0 cm
IQ
4,4%
świnia
Masa ciała (154 dni)
2,6 kg
Wielkość miotu
0,24 młodego
krowa
Roczna produkcja mleka
135 kg
Wpływ inbredu na wzrost frekwencji autosomalnych recesywnych chorób genetycznych u człowieka
(fenyloketonuria, albinizm, alkaptonuria)
choroba
q
Kojarzenie losowe
Frekwencja homozygot aa przy inbredzie
q
2
+Fq(1-q) *
Frekwencja
nosicieli Aa
2q(1-q) *
Frekwencja
homozygot aa
q
2
Kojarzenie kuzynów
I stopnia
(F=1/16)
Kojarzenie
rodzeństwa
(F= ¼ )
fenyloketonuria
1/100
1/50
1/10000
1/1380
1/385
albinizm
1/141
1/70
1/20000
1/2000
1/550
alkaptonuria
1/1000
1/500
1/1000000
1/16000
1/4000
* q→0 => 2q(1-q)= 2q
q→0 => Fq(1-q)=Fq
Depresja inbredowa u szczurów (ok. 30 pokoleń kojarzeń w bliskim pokrewieństwie)
rok
Nieudane kojarzenia
%
Średnia liczba młodych w
miocie
Śmiertelność od urodzenia do 4 tyg.
życia %
5
lat
Od 0% do 41,2%
Od 7,5% do 3,2%
Od 3,9% do 45,5%
Zależność między inbredem a dryfem genetycznym:
dryf genetyczny zmienia frekwencje alleli ( i genotypów)
inbred zmienia frekwencje genotypów, nie zmieniając frekwencji alleli
oba czynniki (mimo różnic) są podobne do siebie w różnych skutkach:
Pod kontrolą inbredu populacja rozsegregowuje się na subpopulacje różniące się od
siebie genetycznie.
Prowadzi to do demowej struktury populacji dużej zmienności między populacjami
(efekt dyspersyjny)
17
Inbred w populacjach naturalnych
Owies Avena sativa- 95-98% potomstwa pochodzi z samozapylenia
Badania wielu lokalnych populacji w Kalifornii- frekwencje genotypów 3 loci, każdy w dwoma
allelami determinującymi cechy kwiatów (niepełna dominacja)
Wnioski:
1. frekwencje heterozygot oczekiwane są większe od obserwowanych
2. wartość F zbliżona dla każdego z 3 loci
Nadmiar homozygot może wynikać również z EFEKTU WAHLUNDA
Definicja:
W dużej populacji jako całości z reguły jest mniej homozygot niż średnio w jej subpopulacjach.
Rozważmy dwie subpopulacje:
AA
Aa
aa
Subpopulacja 1
0,5625 0,3750 0,0625 p
1
=0,75; q
1
=0,25
Subpopulacja 2
0,0625 0,3750 0,5625 p
2
=0,25; q
2
=0,75
Wartości średnie 0,3125 0,3750 0,3125 p=0,50; q=0,50
_
p
2
__
2pq
_
q
2
Subpopulacje łączą się w jedną dużą populację.
Wartości zgodne z regułą H-W
Różnica między średnią frekwencją homozygot aa w subpopulacjach przed ich połączeniem a
frekwencją tych homozygot w jednej populacji powstałej z połączenia subpopulacji wynosi:
_ _
q
2
- q
2
= 0,3125- 0,2500= 0,0625
Wariancja:
_
_
σ
2
= (∑(x
i
- x )
2
)/N lub σ
2
=(∑ x
i
2
/N)- x
2
Wariancja frekwencji allela recesywnego w całej populacji:
_ _ _
σ
2
= (∑ q
i
2
/ k)- q
2
= q
2
- q
2
gdzie k- liczba subpopulacji, na które podzielona jest cała populacja
Podział populacji na szereg subpopulacji doprowadza do tego samego wyniku co wsobność w obrębie
populacji. Są to dwa różne sposoby rozpatrywania tego samego zjawiska.
genotypy
AA
Aa
aa
Populacje zinbredowane (rozkład Wrighta)
p
2
+ pqF
2pq(1-F) q
2
+pqF
0,25 0,50 0,25
p
2
2pq
q
2

18
Populacje podzielone (rozkład Wahlunda)
p
2
+ σ
2
2pq- 2 σ
2
q
2
+ σ
2
↓
2pq (1-F)= 2pq- 2 σ
2
↓ _ _
F= σ
2
/ p q
__
F= σ
2
/ pq = 0,0625/ (0,5×0,5)= 0,25
Zinbredowanie subpopulacji w stosunku do całej populacji:
F
ST
= (H
T
– H
S
)/ H
T
= 0,500- 0,375/ 0,500= 0,25
H
S
– oczekiwana heterozygotyczność osobnika w subpopulacji panmiktycznej
H
T
– oczekiwana heterozygotyczność osobnika w całej populacji panmiktycznej
Zinbredowanie między subpopulacjami jest tym mniejsze, im bardziej nasilona jest migracja
osobników między tymi subpopulacjami ( im bardziej intensywny przepływ genów).
F
ST
określa natężenie przepływu genów między subpopulacjami (natężenie migracji) lub inaczej
stopień izolacji genetycznej między subpopulacjami.
Wnioski i konsekwencje:
1. przed wykorzystaniem reguły Hardy’ego- Weinberga powinna być poznana struktura
populacji (nie uwzględnienie demowej struktury populacji może skutkować błędną
interpretacją sytuacji genetycznej populacji- np. nadmiar homozygot jako skutek selekcji
faworyzującej homozygoty)
2. jeśli wcześniej oddzielone subpopulacje połączą się w jedną dużą- frekwencja homozygot
spada (u człowieka może to oznaczać spadek występowania autosomalnych recesywnych
chorób genetycznych gdy np. wcześniej izolowane mniejsze subpopulacje zlewają się w duże
populacje).
Zdarzenia nieokresowe
(nieprzewidywalne, często o charakterze katastroficznym)
EFEKT ZAŁOŻYCIELA
Konsekwencje genetyczno- populacyjne sytuacji, kiedy osobniki pewnego gatunku dobrowolnie lub
nie znajdą się w miejscu nie zajętym jeszcze przez własny gatunek, ale nadającym się do życia dla
tego gatunku.
Efekt założyciela ujawnia się przez porównanie puli genowej populacji macierzystej i nowo założonej.
Jeśli nowo założona populacja jest mała, to:
może nie zawierać genów, które są rzadkie w populacji macierzystej
jeśli jednak rzadkie geny zostaną wniesione do nowej populacji, to mogą stać się tam częste.
W takiej nowo utworzonej, małej populacji wolno wzrastającej, będzie działał silnie:
• dryf genetyczny (ponowne zubożenie o pewne allele)
• inbred wraz z selekcją (kolejne odrzucenie alleli i połączenie specyfiki genu
populacji)
Spadek heterozygotyczności
H
1
= [1- (1/2 Ne)] H
O
Ne – efektywna liczebność założycieli
Np.
H
O
=0,5 i N=2
H
1
=0,75 H
O
czyli zmniejszenie o 25%

19
Przykład:
Majka żałobna Acridotheres tristis (Indie)
pod koniec XIX w. przeniesiona do Australii, Nowej Zelandii, na Fidżi, Hawaje i do
południowej Afryki
liczba introdukowanych ptaków- od kilkuset do kilku par w południowej Afryce
niektóre z nowych kolonii- niska liczebność poprzez szereg lat
Porównanie zmienności elektroforetycznej białek:
Populacje macierzyste z Indii i populacje introdukowane
mniejsza liczba alleli na locus w populacji introdukowanej
mniejszy udział loci polimorficznych
niższa heterozygotyczność w populacjach introdukowanych
wszystkie allele, których brakowało poza Indiami- w Indiach są rzadkie
frekwencje niektórych alleli poza Indiami znacznie wyższe niż w Indiach
w południowej Afryce zmienność najmniejsza
dystans genetyczny między populacjami introdukowanymi większy niż między macierzystymi
i porównywalny do wartości charakterystycznych
Amisze w Pensylwanii:
Ok. 100 osób z polidaktylią (6 palców), wszystkie pochodzą od jednego z założycieli tej społeczności
z 1744 r.
Stosunkowo często występuje zespół Ellis- van Crevelda (bardzo rzadka wada genetyczna)
Muzułmanie w południowej Afryce:
Dysplazja obojczykowo- czaszkowa- rzadka choroba dziedziczona autosomalnie dominująca, od
jednego założyciela Chińczyka, który przeszedł na Islam.
Wykład 5.
Efekt „szyjki butelki” („wąskiego gardła liczebności”)
Poważna, okresowa, często nagła redukcja wielkości populacji prowadząca do obniżenia zmienności
genetycznej. Skutki są podobne do tych wywołanych efektem założyciela, ale sytuacja dotyczy tej
samej populacji. Po katastrofie pozostająca mała populacja zakłada nową.
W obu efektach można prognozować, jak będzie się zmieniała heterozygotyczność. Tu też spada
według podanej wcześniej zależności.
Przykład:
słoń morski północny (Mirounga angustirostis)
prawie doszczętnie wytępiony przez myśliwych
w 1890 r. – 20 szt.
Dziś – ok. 30000 szt.
Niemal całkowity brak zmienności elektroforetycznej w 24 loci
chomik syryjski (Mesocricetus auratus)
wszystkie dziś żyjące pochodzą od trzech osobników z jednego miotu znalezionych w 1930 r.
na pustyni w Syrii (kolejne 19 dzikich osobników znaleziono pod koniec XX w.)
gepard (Acinonyx jubatus)
drastyczny spadek liczebności w plejstocenie
przeszczepy skóry między niespokrewnionymi osobnikami nie są odrzucane
żubr (Bison bonasus)
gatunek odtworzony z 12 osobników (wyjątkowo niska zmienność genetyczna)
bizon (Bison bison)
przed 1492r. ok. 60000000, 1890r. - 750, 2000r. – 360000
Garnizon, który strzegł Napoleona skazanego na wygnanie na Św. Helenę- grupka żołnierzy
osiadła na wyspie Tristan da Chuna na południowym Atlantyku.
20
Kryzys- część osób opuściła wyspę, część utopiła się podczas połowu ryb.
Dzisiaj- kilkaset mieszkańców odrodzonych z tej garstki.
Stosunkowo często dziedziczona ślepota po jednej z żon założycieli.
Erupcja superwulkanu Toba (Indonezja) 67,5- 75,5 tys. lat temu, redukcja populacji ludzkiej
do kilku tysięcy (? 1 tys.)
↓ ↓ ↓
Homogenizacja genetyczna, odróżnicowywanie się lokalnych populacji, wymarcie innych
gatunków Homo poza H. sapiens i H. neanderthalensis.
EFEKTY SYSTEMATYCZNE
Migracje
Przyjmijmy:
N- wielkość analizowanej populacji
q- frekwencja wyjściowa allela a w analizowanej populacji
q’- frekwencja tego allela w populacji, z której pochodzą imigranci (q≠q’)
m- tempo migracji
N= constans
Model przepływu genów
Z analizowanej populacji w każdym pokoleniu emigruje mN osobników, a na ich miejsce imigruje ta
sama liczba osobników z innej populacji o frekwencji allela a równej q’, przy czym w kolejnych
pokoleniach ani m, ani q’ nie zmieniają się.
Jak zmieni się frekwencja allela a?
q
1
= (1-m)q + mq’ = q – mq + mq’
↓ ↓
Autochtoni; Przybysze
Tempo zmiany częstości allela a:
Δq= q
1
– q= (q- mq + mq’)- q= mq’- mq= m(q’- q)
Δq= 0 m(q’- q)= 0 czyli jeśli m= 0 lub q’= q
Migracja (w podanym modelu) jest procesem nierównowagowym i prowadzi do homogenizacji
genetycznej populacji.
q
q’
m

21
Tempo wyrównania różnicy między q i q’:
q
1
- q’= (q – mq+ mq’)- q’= q- q’- m(q- q’)= (1- m)(q- q’)
analogicznie:
q
2
- q’= (1- m)( q
1
- q’)= (1- m)
2
(q- q’)
Ogólnie po n pokoleniach imigracji z tym samym tempem m, różnice między frekwencją allela a w
analizowanej populacji w stosunku do frekwencji tego allela w populacji, z której pochodzą imigranci:
q
n
- q’= (1- m)
n
(q- q’)
(q
n
- q’)/ (q- q’)= (1- m)
n
Np.
m= 0,2
q
0
= 0,5
q
n
= 0,8
q’= 0,9
(0,8- 0,9)/(0,5- 0,9)= (1- 0,2)
n
0,25= 0,8
n
n= log
0,8
0,25= ok. 6 pokoleń
Migracje mogą mieć bardzo duży wpływ na kształtowanie częstości alleli.
Zadanie:
W populacji składającej się z 1000 osobników frekwencja pewnego allela wynosiła 80%
A) ile wyniesie frekwencja tego genu, jeśli populację opuści 400 osobników, a na ich miejsce
przybędzie 400 osobników u których frekwencja tego genu wynosi 10%?
Odp. A
q
1
= q- mq+ mq’= 0,8- (0,4×0,8)+ (0,4×0,1)= 0,52
B) ile wyniesie frekwencja tego genu bez emigracji 400 osobników tubylczych?
Odp. B
q
1
= (q+ mq’)/ (1+ m)= [0,8+ (0,4×0,1)]/ (1+ 0,4)= 0,60
(imigracja bez emigracji)
Mutacje
Załóżmy, że nie działa selekcja, czyli dostosowanie wszystkich genotypów jest identyczne (brak
różnic w przeżywalności i rozrodczości)
Allel, który nie powoduje zmian w dostosowaniu genotypów (zarówno w układzie homo- ,jak i
heterozygotycznym) uważany jest za neutralny z punktu widzenia selekcji (allel neutralny, mutacja
neutralna).
Jeśli A mutuje w kierunku a z częstością u, to nowa frekwencja allela recesywnego:
q
1
= q+ (1- q) u
Zmiana tej frekwencji w ciągu jednego pokolenia:
Δq= q
1
- q= q+ (1- q) u- q= (1- q) u
Zmiana frekwencji allela a zależy od frekwencji allela A (1- q= p)
p= 1 => Δq= u
p= 0 => Δq= 0
Im rzadszy allel a (a częstszy allel A), tym więcej mutacji A→a.
Jeśli również a mutuje w kierunku A z pewną częstością v, to nowa frekwencja allela dominującego:
p
1
= p+ (1- p) v
natomiast Δp= (1- p) v
Ogólnie:
u
A
----→
a
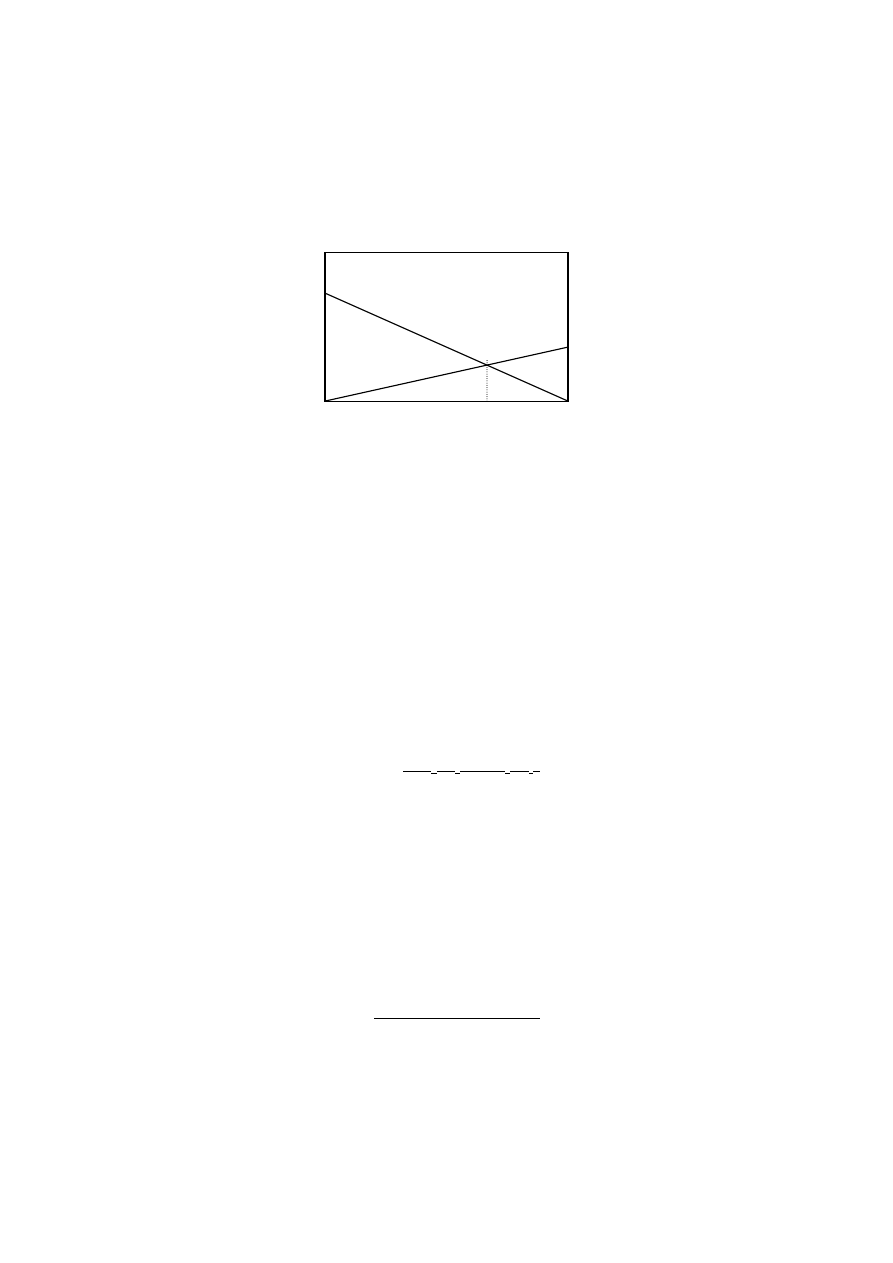
22
←----
v
Zmiana częstości allela a przy mutacjach w obu kierunkach:
q
1
= q
0
+ pu- qv
Czy istnieje taka frekwencja allela a (q
e
), przy której zmiany frekwencji w jednym kierunku (A→a) są
równoważone przez zmiany frekwencji w drugim kierunku (a→A)?
Δq= q
1
- q= (q+ pu- qv)- q= pu- qv
Δq= 0 pu- qv= 0
(1- q) u- qv= 0
u- qu- qv= 0
qu+ qv= u
q
e
= u/ (u+ v)
analogicznie:
p
e
= v/ (u+ v)
Wniosek:
Mutacje (w przedstawionym modelu) są procesem równowagowym, a frekwencja równowagowa
(punkt równowagi trwałej) nie zależy od początkowej częstości alleli, lecz tylko od częstości mutacji.
Jak kształtuje się w czasie osiąganie wyznaczonego punktu równowagi frekwencji alleli neutralnych?
n- liczba pokoleń
Muszą być spełnione warunki: q
0
>q
e
i q
n
>q
e
(logarytm ma wtedy sens)
Np. u= v= 10
-6
q
0
= 0,9
q
n
= 0,6
q
e
= u/ (u+ v)= 10
-6
/ (10
-6
+10
-6
)= 0,5
Zmiana z q
0
= 0,6 do q
n
= 0,51 wymaga 1151293 pokolenia!
Wnioski:
Liczba pokoleń potrzebna do ustalenia się równowagi określonej jedynie przez mutacje jest bardzo
duża biorąc nawet pod uwagę długi czas przebiegu procesów ewolucyjnych.
v
Δp
Δq
u
q
e
ln(
q
0
-
q
e
)- ln(
q
n
-
q
e
)
u+ v
n=
ln(
0,9
-
0,5
)- ln(
0,6
-
0,5
)
10
-6
+
10
-6
n=
= 693147

23
Sama presja mutacyjna jest zwykle bardzo słabą siłą ewolucyjną, która może powodować tylko
niewielkie zmiany w częstości alleli, a opisany stan równowagi jest raczej konsekwencją teoretyczną
niż zjawiskiem, którego należałoby oczekiwać w naturze.
Zadanie:
W populacji o liczebności 1 mln osobników częstość allela a wynosi 10%, zaś mutacja allela A w allel
a przebiega z częstością 10
-4
na allel na pokolenie.
Określ liczbą alleli A, które zmieniły się w allel a w jednym pokoleniu.
Odp.
2N(1- q) u= 2× 1000000× 0,9× 10
-4
= 180
Dryf mejotyczny (Meiotic drive)
Mechanizmy, które powodują, że określony czynnik (chromosom, gen) przechodzi do funkcjonalnych
komórek płciowych częściej niż można tego oczekiwać zgodnie z prawami przypadkowości
(nielosowa segregacja chromosomów w mejozie)
Przykład 1.
Czynniki zakłócające segregację (SD- segregation-distorter) u Drosophila melanogaster w sektorze
heterochromatynowym chromosomu II.
Heterozygotyczne samce SD/+ przekazują chromosom SD z częstością większą niż 50% (nieraz
prawie 100%).
Plemniki bez czynnika SD nie rozwijają się prawidłowo wskutek hamujących oddziaływań
chromosomu z czynnikiem SD.
Budowa kompleksu genów czynnika SD- patrz podręcznik!
Przykład 2.
Locus T (Brachyury) u Mus musculus- supergen (szereg loci położonych blisko siebie o ograniczonej
rekombinacji) zajmujący ok. 1% genomu.
W populacjach naturalnych allel t osiąga duże częstości, mimo że w stanie homozygotycznym jest
letalny bądź powoduje sterylność samców. Układy heterozygotyczne wywołują zmiany kręgosłupa i
okolicy ogonowej.
Mimo niekorzystnych efektów allel t przekazywany jest do 80- 90% gamet.
Mechanizm podobny jak u samców Drosophila z czynnikiem SD.
W naturalnych populacjach frekwencja tego allela nie przekracza 0,4 (według modelu powinna
wynosić ok. 0,7).
Wniosek:
Występuje równowaga między siłami dryfu mejotycznego i selekcją eliminującą allele recesywne.
Mechanizmy dryfu mejotycznego stanowią kierunkową siłę, która wbrew selekcji może istotnie
zwiększyć częstość określonych alleli w populacji.
Często na skutek dryfu mejotycznego dochodzi do zwiększenia frekwencji i rozprzestrzeniania się
alleli niekorzystnych aż do ustalenia określonej frekwencji równowagowej (lub wymarcie całej
populacji).
Dryf mejotyczny jest mechanizmem trudnym do modelowania.
Wykład 6.
Selekcja (dobór naturalny)
Przesłanki (K. Darwin):
osobniki każdego gatunku różnią się między sobą i potomkowie są zwykle podobni do
rodziców
24
liczba gamet i osobników młodych w każdym pokoleniu przewyższa liczebność pokolenia
rodziców i pojemność środowiska
Wniosek:
Tylko niektóre osobniki (najlepiej dostosowane do danych warunków życia) dożywają do okresu
rozrodu i pozostawiają potomstwo (przeżycie najstosowniejszego).
Zróżnicowana transmisja genów do kolejnego pokolenia przez nosicieli różnych genotypów
Selekcja- warunki
występowanie wariantów danej właściwości (zróżnicowanie)
osobniki reprezentujące poszczególne warianty cech odznaczają się zróżnicowanym
prawdopodobieństwem przeżycia i reprodukcji (różnice w przeżywalności i rozrodczości)
zmienność przynajmniej częściowo determinowana genetycznie (zmienność dziedziczna)
wynik:
osobniki o właściwościach warunkujących większe szanse przeżycia i rozrodu stają się
częstsze w populacji.
Selekcja naturalna w toku
1. Heliothis virescens, Noctuidae
Występowanie: zachodnia półkula, od północnej Kanady po Argentynę
Rośliny żywicielskie: uprawiane (głównie bawełna, soja, pomidor, tytoń), dziko
rosnące (trawy Muhlenbergia regens i babkowate)
Cykl życiowy: zimują jako poczwarki, Imagines w kwietniu- maju (USA), jaja na
liściach i pąkach kwiatowych, larwy żerują od 3 do 4 tygodni, 4 pokolenia w roku,
poczwarki w ziemi od września.
Na początku XX w. występował na żywopłotach i w lasach.
Plantatorzy bawełny od 1940r. stosują DDT.
Nieliczne osobniki H. virescens tolerowały DDT i zauważono je żerujące na bawełnie.
Większe dawki DDT, nowe insektycydy (szczyt lata 60-70 XX w.)
Krótkotrwały sukces- „wyścig zbrojeń”
Pyretroidy (nowe środki owadobójcze)- początkowo podniesienie plonów o ¼ - 1/3.
1980- osobniki o zwiększonej 50× oporności na pyretroidy
Maj 1987- przeżywa 6% osobników
koniec września 1987r.- tę samą dawkę przeżywa 61% (jeden sezon!)
lata 90. XX w.- osobniki o 200× zwiększonej oporności, są obecnie niemal zupełnie oporne na
działanie wszystkich produkowanych insektycydów!
Insektycyd- wywiera presję selekcyjną tzn. jest czynnikiem selekcyjnym skierowanym
przeciw cechom, które stanowią, że osobnik jest podatny na truciznę. Trucizna dokonuje
selekcji na rzecz cechy, która czyni osobnika mniej podatnym, ponieważ najbardziej oporne
osobniki mają większe szanse przeżycia i zostawiają najwięcej potomstwa.
2. krępak brzozowy (Biston betularia, Geometridae)
forma typica- mm
forma carbonaria- M_

25
Początkowo większość populacji stanowiła odmiana jasna (f. typica)- kamuflaż na drzewach z
porostami. Odmiana melanistyczna, czarna (f. carbonaria)- łatwiej widoczna dla drapieżników.
Forma typica- prawie 100% w 1848 r.
Rozwój przemysłu i zanieczyszczenie środowiska- zmniejszenie występowania porostów.
Forma ciemna maskowała się lepiej od jasnej i takie osobniki zaczęły występować częściej
(Anglia- przełom XIX i XX w.).
Forma carbonaria- 98% w 1895 r.
Od 1962 r. na skutek zmniejszenia stopnia zanieczyszczenia obserwuje się przeciwny proces.
Przy ilościowym prognozowaniu selekcji podstawową sprawą jest określenie różnic w przeżywalności
i rozrodczości.
Współczynnik reprodukcji netto (R)
Definicja: średnia liczba potomków przeżywających do następnej reprodukcji przypadających na
jednego osobnika o danym genotypie.
R= B × S
Gdzie: B- liczba potomków
S- prawdopodobieństwo ich przeżycia
R=1 => liczebność nie zmienia się
R<1 => liczebność maleje
R>1 => liczebność rośnie
Genotypy różniące się rozrodczością i prawdopodobieństwem przeżycia różnią się zwykle wartością
współczynnika reprodukcji netto (R).
Różne genotypy mogą mieć równe wartości R mimo różnic w wartościach B i S.
Liczebnośc będzie wzrastała osiem razy.
W modelach genetyczno- populacyjnych istotne są różnice między genotypami, a nie rzeczywiste
wartości R.
Dostosowanie (wartość przystosowawcza, fitness)- W
Definicja: stosunek wartości R danego genotypu do (zwykle) najwyższej wartości R dla danego
zestawu genotypów.
Np.
AA Aa
aa
R
4
4
3
W
1
1
0,75
W określa w jakiej mierze dany genotyp jest faworyzowany przez selekcję.
Współczynnik selekcji (s)
Definicja: miara intensywności eliminacji genotypu (genu).
s= 1- W
czyli w powyższym przykładzie:
s
AA
= 0
s
Aa
= 0
s
aa
= 0,25
Genotyp A B= 20 S= 0,4 R= 8
Genotyp B B= 10 S= 0,8 R= 8

26
Sytuacja szczególna- zupełna eliminacja homozygot recesywnych
genotypy
AA
Aa
aa
frekwencje
p
2
2pq
q
2
liczebności
Np
2
N2pq Nq
2
Dostosowanie (W)
1
1
0
Współczynnik selekcji (s)
0
0
1
Całkowita liczebność populacji (N) po eliminacji homozygot recesywnych:
aa= Np
2
+ N2pq + 0 = Np (p + 2q) = Np (p + q + q) = Np (1 + q)
Jeżeli ogólnie: q= (N
Aa
+ 2N
aa
)/ 2N, to:
Frekwencja allela recesywnego w następnym pokoleniu:
q
1
= (2Np
0
q
0
+ 0)/ 2N p
0
(1 + q
0
) = (p
0
q
0
)/ p
0
(1+ q
0
) = q
0
/(1 + q
0
)
w drugim pokoleniu:
q
2
= q
0
/(1 + 2 q
0
)
w pokoleniu n-tym:
q
n
= q
0
/(1 + n q
0
)
Ile pokoleń potrzeba na zmniejszenie frekwencji allela a z q
0
do q
n
?
n= (1/ q
n
) – (1/ q
0
)
jeżeli q
0
= 0,5 to:
q
1
0,333
q
2
0,250
q
3
0,200
q
4
0,167
q
5
0,143
…
…
q
10
0,084
…
…
q
100
0,010
…
…
q
1000
0,001
Przykład hipotetyczny:
MUKOWISCYDOZA- choroba autosomalna recesywna, uszkodzenie białka CFTR regulującego
działanie kanałów chlorkowych w błonie komórkowej (ok. 500 typów mutacji)
Częstość- 1:2000 czyli p
aa
= 0,0005 (nosiciele- 4%)
Frekwencja alleli recesywnych q= 0,022
Załóżmy, że W
aa
= 0
Ilu pokoleń potrzeba, aby zmniejszyć frekwencję allela a o połowę?
n= (1/ 0,011) – (1/ 0,022) = 91 – 45 = 46 czyli około 1150 lat!!!
Przy q=0,011 nadal 1 heterozygota na 50 osób!!!
Wniosek:
Przy zupełnej eliminacji homozygot recesywnych frekwencja allela recesywnego spada początkowo
bardzo szybko, a następnie tym wolniej, im ten allel jest rzadszy w populacji (jest utrzymywany
głównie w heterozygotach).
Zupełna eliminacja przy dużej liczebności populacji wymaga nieskończenie wielu pokoleń!
Selekcja może dotyczyć każdego stadium rozwojowego.
Zasadne jest, aby rozpatrywać 3 poziomy:
Poziom gamet
Poziom zygot
27
Etap zalotów (selekcja płciowa)
Poza tym:
Poziom grupy osobników (selekcja grupowa)
Poziom gatunkowy
Poziom genów
Selekcja gametyczna- model formalny
100 gamet A- do zapłodnienia przeżywa 90
100 gamet a- do zapłodnienia przeżywa 80
p
0
(A)= 0,5 W
1
(A)= 0,9 lub W
1
(A)= 1,0
q
0
(a)= 0,5 W
2
(a)= 0,8
lub W
2
(a)= 0,89 (0,8/0,9)
Nową frekwencję gamet a po selekcji obliczymy jako 80/(80+90)= 0,47
Lub jeśli W
1
= 1 to:
W celu określenia tempa selekcji określamy zmiany w częstości allela a na pokolenie:
∆q= q
1
- q
0
Kiedy ∆q= 0?
∆q= 0 q= 0 lub q= 1
Lub
brak selekcji
∆q= 0 W= 1
Wniosek:
Selekcja gametyczna jest procesem nierównowagowym i prowadzi do utraty któregoś z alleli.
Wykład 7.
Ogólny model selekcji
genotyp
AA
Aa
aa
frekwencja
p
2
2pq
q
2
wartość przystosowawcza
W
1
W
2
W
3
wartość przystosowawcza wyrażona współczynnikiem selekcji
1- s
1
1- s
2
1- s
3
q
1
=
W
2
q
0
W
2
q
0
+
W
1
p
0
q
1
=
W
q
0
Wq
0
+ p
0
-
p
0
q
0
(1 – W)
p
0
+ W q
0
=
=
=
=
=
W
q
0
p
0
- p
0
q
0
Wq
0
+ p
0
W
q
0
(1-
q
0
) -
p
0
q
0
Wq
0
+p
0
W
q
0
- q
0
(W q
0
+ p
0
)
Wq
0
+ p
0
W
q
0
- W
q
0
2
-
p
0
q
0
W
q
0
+
p
0
W
q
0
Wq
0
+ p
0
∆q=
=
28
↓
↓
↓
selekcja
↓
↓
↓
frekwencje genotypów
_
p
2
(1- s
1
)+ 2pq(1- s
2
)+ q
2
(1- s
3
)= W
_
p
2
W
1
+ 2pqW
2
+ q
2
W
3
= W
W- średnia wartość przystosowawcza populacji
Określa stosunek liczby wszystkich osobników po selekcji do liczby wszystkich osobników przed
selekcją:
W→ 1 słaba selekcja
W→ 0 silna selekcja
_
W = p
2
(1- s
1
)+ 2pq(1- s
2
)+ q
2
(1- s
3
) = p
2
- p
2
s
1
+ 2pq – 2pqs
2
+ q
2
- q
2
s
3
= (p
2
+ 2pq + q
2
) - p
2
s
1
- 2pqs
2
-
q
2
s
3
= 1 - p
2
s
1
- 2pqs
2
- q
2
s
3
Frekwencja allela a po selekcji:
_
W≠ 0
pq
(1- s
2
) – połowa frekwencji heterozygot po selekcji
q
2
(1- s
3
) – frekwencja homozygot recesywnych po selekcji
Zmiana częstości allela a w ciągu jednego pokolenia: ∆q =
q
1
-
q
0
↓
∆q =
pq [ p(s
1
- s
2
) – q (s
3
- s
2
) ]
Kiedy ∆q = 0?
(czyli w jakich warunkach nie występują zmiany frekwencji alleli?)
∆q = 0 1. p= 0
jeśli nie ma zróżnicowania alleli- nie ma selekcji
2. q= 0
3. p(s
1
- s
2
) – q(s
3
- s
2
)= 0
p(s
1
- s
2
) – q(s
3
- s
2
)= 0
↓
Np
2
W
1
+ N2pqW
2
+ Nq
2
W
3
N
_
W =
q
1
=
pq
(1- s
2
) + q
2
(1- s
3
)
_
W
- q
0
2pq
(1- s
2
) + q
2
(1- s
3
)
_
W
∆q =
1
_
W
s
2
-
s
1__
2s
2
- s
1
- s
3
29
q=
s
2
= 0 =>
q
e
=
s
1
= 1
s
1
= 0,5
s
2
= 0,5 → zakładamy, że s
2
= 0 → s
2
= 0
s
3
= 0,2
s
3
= -0,3
s
1
= 1
s
1
= 1,5
lub s
2
= 0,5 → zakładamy, że s
2
= 1 → s
2
= 1
s
3
= 0,2
s
3
= 0,7
Trzy możliwe warianty sytuacji:
q
e
=
s
1
s
2
s
3
I
>0 0 >0
selekcja stabilizująca
MOCNA
stabilizuje polimorfizm, faworyzuje heterozygoty, naddominacja
heterozygot = selekcja skierowana przeciwko obu homozygotom
II a
>0 0 <0
selekcja kierunkowa
eliminacja homozygot dominujących, faworyzacja recesywnych
II b <0 0 >0
eliminacja homozygot recesywnych, faworyzacja dominujących
III
<0 0 <0
selekcja stabilizująca
SŁABA
heterozygoty eliminowane, faworyzacja homozygot, wstęp do selekcji
rozrywającej
Przykładowe wartości:
s
1
s
2
s
3
I
0,3
0
0,2
q = 0,6 czyli 0<q<1
0,2
0
0,3
q = 0,4 czyli 0<q<1
II a 0,2
0
-0,1 q = 3,0 czyli q>1
brak fizycznego sensu
II b -0,1 0
0,2
q = -2,0 czyli q<0
III
-0,3 0
-0,2 q = 0,6 czyli 0<q<1 - bardzo chwiejna równowaga
- zaburzenie powoduje zmierzanie do p=0 lub p=1
-0,2 0
-0,3 q = 0,4 czyli 0<q<1
I – proces równowagowy (równowaga trwała)
III – proces równowagowy (równowaga nietrwała)
II a, II b – proces nierównowagowy prowadzący do wyeliminowania jednego z alleli
Selekcja „twarda” (= „sztywna”) i „miękka” (= „giętka”)
Selekcja „twarda”
Zakłada stałość wartości współczynników selekcji (s)
Eliminuje pewną proporcję osobników w każdych warunkach, zależnie jedynie od wartości „s”
poszczególnych genotypów, bez względu na działanie ogółu innych czynników.
Selekcja „miękka”
Nie zakłada stałości wartości współczynnika selekcji (s)
Eliminuje określoną proporcję osobników, która przewyższa pojemność siedliska i chociaż działa
wybiórczo na poszczególne genotypy, to ich wartości „s” mogą zależeć np. od zagęszczenia populacji
czy dostępności zasobów.
Selekcja typu r i typu K
s
1___
s
1
+ s
3
s
1___
s
1
+ s
3
30
dN/dt = rN - nieograniczony wzrost liczebności populacji
gdzie: N – liczebność populacji
r – miara przyrostu naturalnego (= wewnętrzna miara wzrostu populacji)
t – czas
Interpretacja:
Przyrost liczby osobników na jednostkę czasu jest liniowo proporcjonalny do liczby osobników N, a
jego tempo zależy od wewnętrznej miary wzrostu populacji r.
r = (1/N)(dN/dt)
miara przyrostu naturalnego przy nieograniczonym wzroście liczebności populacji (można stosować
do poszczególnych genotypów)
Selekcja będzie faworyzowała takie genotypy, które odznaczają się wyższą wartością „r”, czyli w
jednostce czasu będą pozostawiały więcej potomstwa.
(dN/dt) = rN
- ograniczony wzrost liczebności populacji wynikający z ograniczonej
ilości zasobów (prawa Verhulsta)
gdzie: K – pojemność siedliska
(1 – N)(dN/dt) = r
- tempo wzrostu populacji na osobnika (miara dostosowania przy
wzroście ograniczonym)
1. populacja często niszczona z powodu niesprzyjających warunków, tak że jej liczebność (N)
rzadko osiąga pojemność siedliska (K)
N<<K => K – N → K => (K – N)/K → 1 => (1/N)(dN/dt) → r
Aby zmaksymalizować wzrost liczebności populacji należy maksymalizować miarę przyrostu
(r) → selekcja typu r
2. populacja żyje w siedlisku stabilnym, a jej wielkość (N) jest bliska pojemności siedliska (K)
N≈K => K – N → 0 => (K – N)/K → 0 => (1/N)(dN/dt) → 0
Nawet przy bardzo wysokich wartościach miary przyrostu (r), tempo wzrostu liczebności
populacji zbliża się do 0, a jedynym sposobem jego maksymalizacji jest zwiększenie wartości
K, czyli zwiększenie zdolności do wykorzystania zasobów siedliska (K) → selekcja typu K
Selekcja typu r
Występuje gdy:
warunki zmienne i nieprzewidywalne
śmiertelność często katastroficzna i niezależna od zagęszczenia
zbiorowiska nienasycone
często rekolonizacja
słaba lub zmienna konkurencja wewnątrz- i międzygatunkowa
Prowadzi do:
szybki rozwój osobniczy
wczesny rozród
jednorazowa reprodukcja
mała wielkość ciała
wysokie tempo wzrostu liczebności populacji
wysoka produktywność
Selekcja typu K
Występuje gdy:
warunki stałe i przewidywalne
śmiertelność zależna od zagęszczenia
zbiorowisko nasycone
K – N
K
K – N
K
31
silna konkurencja
Prowadzi do:
powolne dojrzewanie
duża zdolność do konkurencji
duża wielkość ciała
długie życie
duża zdolność do wykorzystania zasobów
Selekcja apostatyczna (zależna od częstości)
Selekcja faworyzująca „odstępców”- rzadsze formy, odmienne genetycznie, mogą mieć większe
szanse przeżycia.
W stanie równowagi dostosowanie wszystkich genotypów jest identyczne
Przykłady:
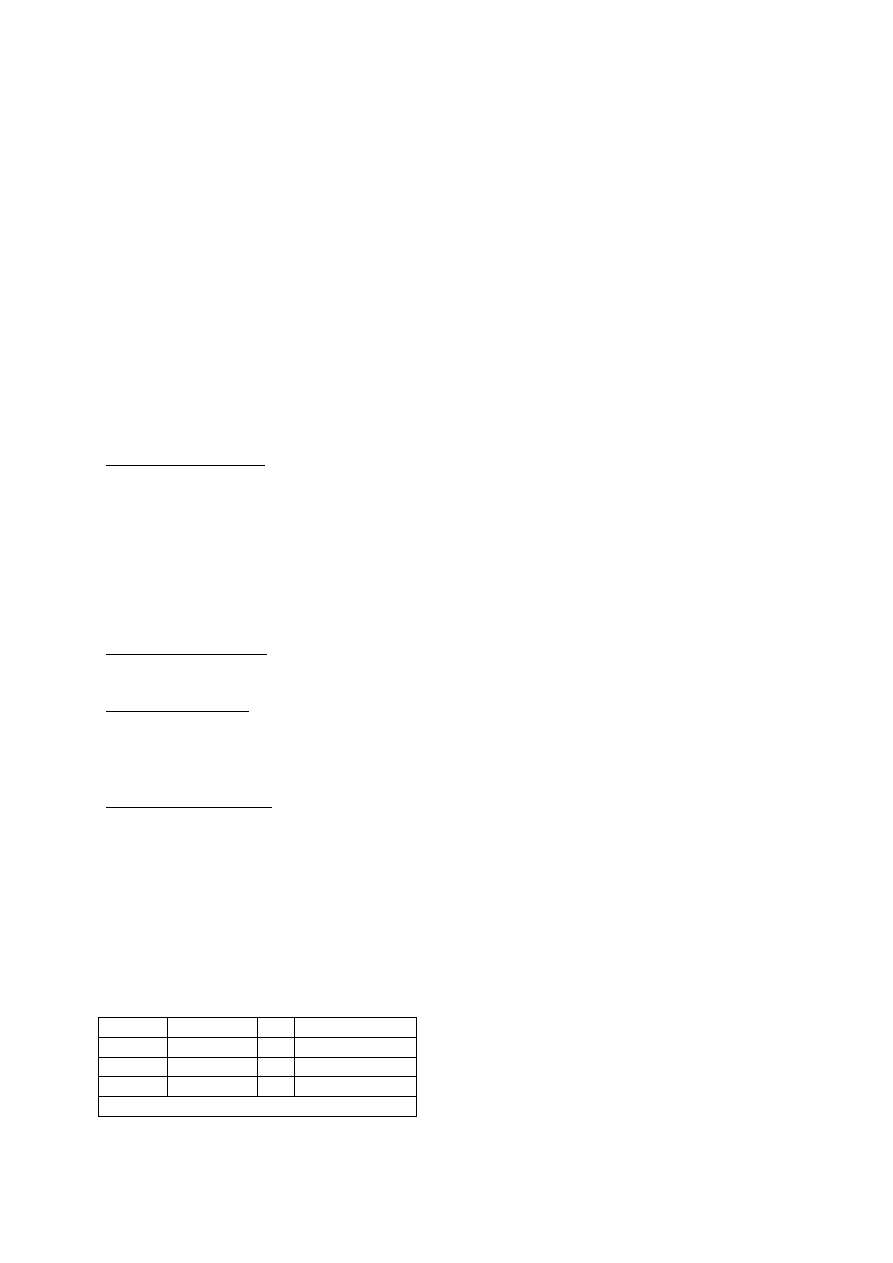
1. Perissodus microlepis (Cichlidae)
Ryby zjadające łuski- usta w lewą lub w prawą stronę
2. Cepaea nemoralis
Osobne loci decydujące o tle muszli, ilości pasków i ich grubości
Czynnikiem selekcyjnym są ptaki np. drozdy zjadające ślimaki
Ubarwienie- charakter kryptyczny
Najczęściej wyjadana typowa, przeciętna forma- rzadsze są faworyzowane (nie są
zjadane)
3. Paracerceis sculpta (Crustacea; Isopoda)- równonóg (wesz gąbkowa)
Samce α- duże, wyraźnie odróżniające się
Samce β- wyglądają jak samice
Samce γ- wyglądają jak samice, ale są dużo mniejsze
samiec α X
A
X
A
81%
samiec β X
B
X
_
4%
samiec γ X
A
X
G
lub X
G
X
G
15%
Teoretycznie selekcja zależna od częstości może również faworyzować formy częstsze (selekcja
aposematyczna), ale są to sytuacje znacznie rzadsze w przyrodzie
Polimorfizm genetyczny
Jednoczesne występowanie w populacji więcej niż jednego allela w tym samym locus, przy czym
frekwencja najrzadszego z nich jest na tyle duże, że nie wynika jedynie z powtarzających się mutacji
(czyli zwykle frekwencja ta > 1%)
prawouste
lewouste
32
Polimorfizm przejściowy- przy zmieniających się warunkach ekologicznych jeden allel jest
eliminowany przez inny w wyniku doboru
Polimorfizm zrównoważony- utrzymywanie się w populacji przez długi czas więcej niż jednego allela
danego locus
Mechanizmy chroniące zmiennośc genetyczną i utrzymujące w populacji zrównoważony polimorfizm:
Naddominacja heterozygot (selekcja faworyzująca heterozygoty)
Selekcja apostatyczna
Selekcja „miękka”
Przepływ genów
Przeciwny nacisk selekcyjny na różne genotypy
Heterogamia (kojarzenie się partnerów niepodobnych)
Supergeny i bloki genów koadaptowanych, supresory crossing- over
Wykład 8.
Obciążenie genetyczne
Obniżenie wartości przystosowawczej danego osobnika lub populacji w stosunku do teoretycznej
wartości optymalnej na skutek występowania genów o niekorzystnych oddziaływaniach.
Jeżeli w populacji istnieją genotypy o różnym dostosowaniu, to jedno z nich jest najwyższe (W
max
), a
średnie dostosowanie osobnika W< W
max
.
Polimorfizm obniża średnie dostosowanie W osobnika w populacji i powoduje obciążenie genetyczne
L, dane wzorem:
L= (W
max
– W)/ W
max
W
max
= 1 => L= 1- W
obciążenie segregacyjne- powodowane wysokim dostosowaniem heterozygot (określone jest tylko
przez współczynnik selekcji)
obciążenie mutacyjne- powodowane przez niekorzystne mutacje; zależy jedynie od częstości mutacji,
a nie od dostosowania genotypu recesywnego, ponieważ przy wyższych wartościach współczynników
selekcji mutacje te są skuteczniej eliminowane przez dobór i dlatego dają takie same obciążenie
mutacyjne, jak mutacje mniej szkodliwe.
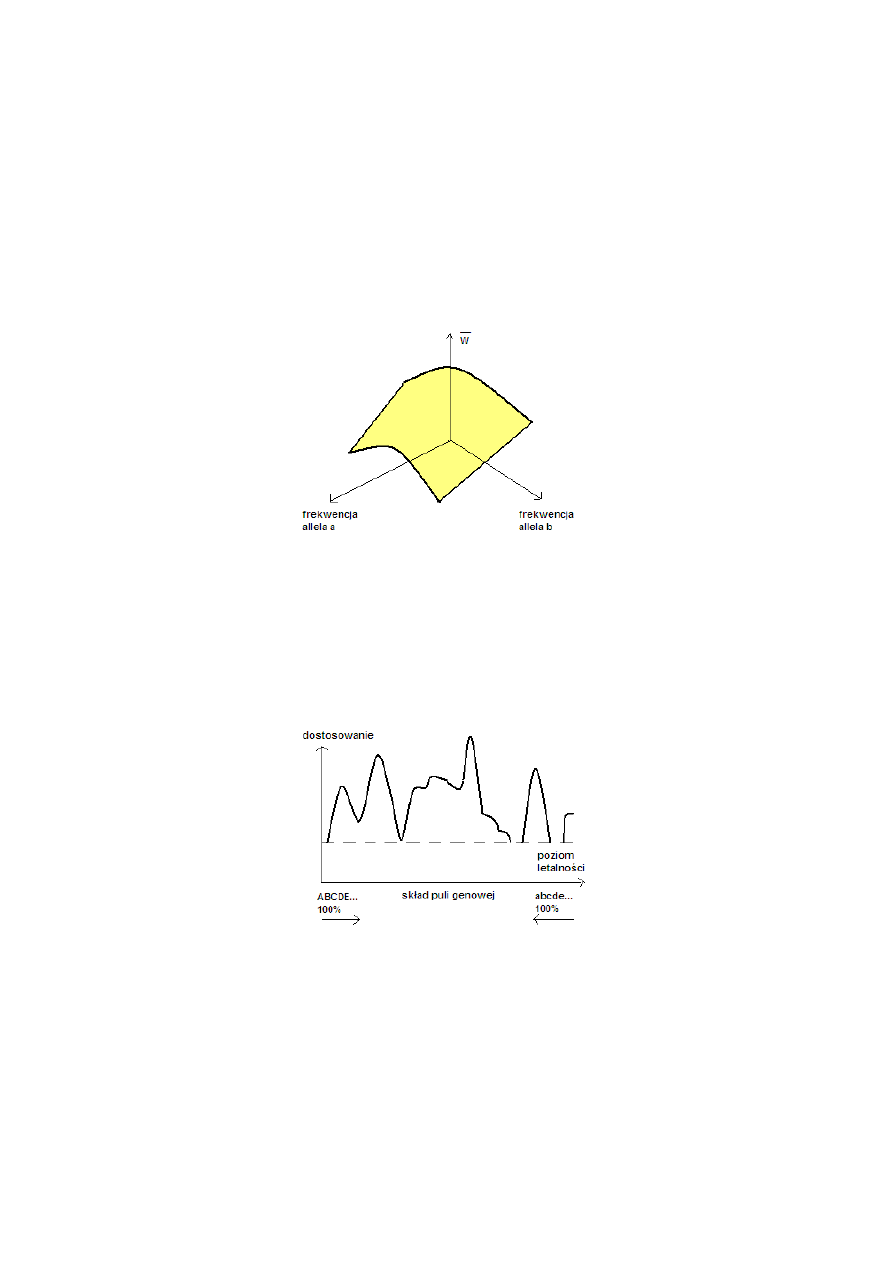
Krajobraz adaptacyjny
Średnie dostosowanie populacji zależy od składu jej puli genowej (S. Wright)
Załóżmy, że:
W
1
= 1
W
2
= 0,8
W
3
= 0,2
Przy różnych frekwencjach alleli można wyliczyć średnią wartość przystosowawczą dla całej
populacji:
Np. przy p= q= 0,5
genotyp frekwencja W
frekwencja × W
AA
p
2
= 0,25
1,0 0,25
Aa
2pq= 0,5
0,8 0,40
aa
q
2
= 0,25
0,2 0,05
Średnia wartość przystosowawcza 0,70
Np. przy p= 0,4 i q= 0,6
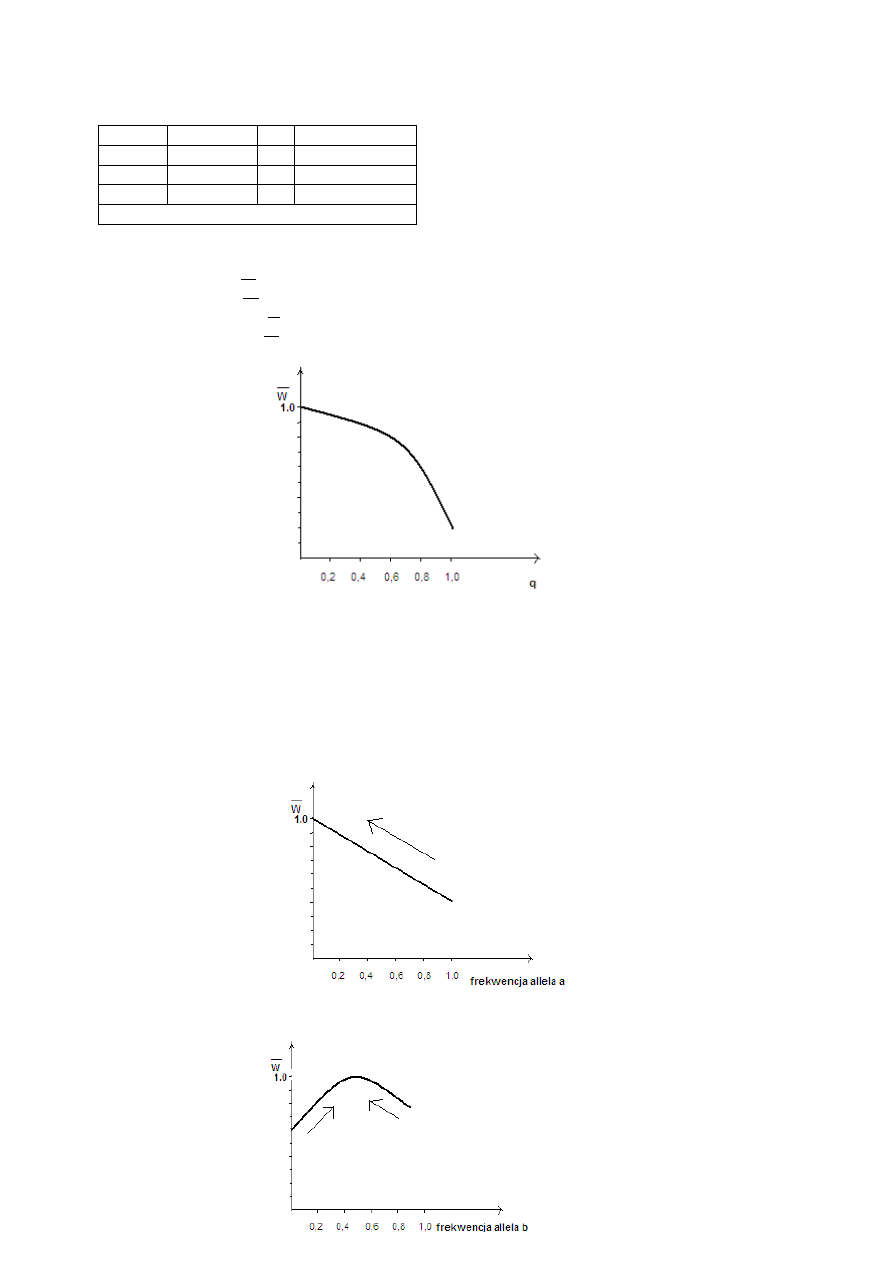
33
genotyp frekwencja W
frekwencja × W
AA
p
2
= 0,16
1,0 0,160
Aa
2pq= 0,48
0,8 0,384
aa
q
2
= 0,36
0,2 0,072
Średnia wartość przystosowawcza 0,616
Ponieważ ta sytuacja opisuje selekcję kierunkową faworyzującą homozygoty dominujące, to przy
wyższych frekwencjach allela dominującego W średnie będzie rosło.
przy p= 1, q= 0 => W= 1,00
przy p= 0, q= 1 => W= 0,20
przy p= 0,5, q= 0,5 => W= 0,70
przy p= 0,4, q= 0,6 => W= 0,616
W średnie dla drugiego i następnych loci obliczamy analogicznie.
Średnia zbiorowa wartość przystosowawcza uwzględniająca wszystkie analizowane loci jest średnią
arytmetyczną średnich W dla każdego z nich.
Selekcja kierunkowa (eliminacja aa)
Selekcja stabilizująca
(eliminacja BB i bb)
34
Biorąc dwa w/w pod uwagę:
Populacja może być postrzegana jako wspinająca się po powierzchni adaptacyjnej w kierunku
szczytów adaptacyjnych do osiągnięcia punktu równowagi.
Krajobrazy adaptacyjne można przedstawić w postaci map, na których punkty o jednakowym
dostosowaniu łączy się w tzw. izolinie.
Można też dokonać przekroju przez diagram (mapę) w dowolnej płaszczyźnie pionowej i wykreślić
profil, podobnie jak z mapy.
Hipotetyczny profil przekrojowy mapy wartości przystosowawczej złożonego systemu genetycznego.
Można oczekiwać, że wszystkie populacje powinny zmierzać do najwyższego szczytu adaptacyjnego.
W niektórych przypadkach jest to jednak niemożliwe, ponieważ populacja nie może zejść poniżej
poziomu letalności.
Przykład:
Krajobraz adaptacyjny na podstawie polimorfizmu inwersyjnego chromosomów Moraba scurra
(Orthoptera)
metacentryk →→ inwersja pericentryczna→→ akrocentryk
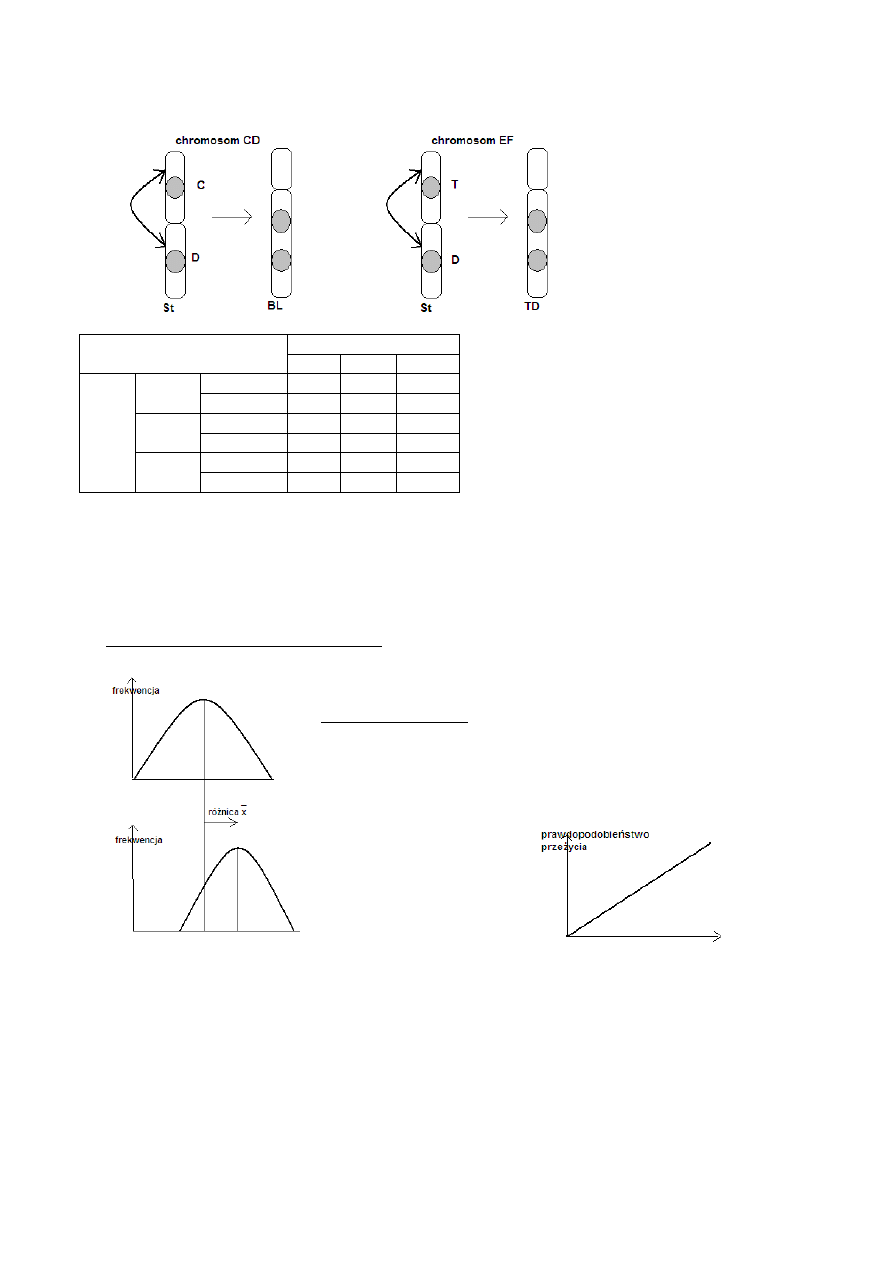
35
chromosom CD
St/St
St/BL BL/BL
chr
om
osom
EF
St/St
liczebność
7
100
324
żywotność 0,970 1,000 0,917
St/TD
liczebność
3
22
118
żywotność 1,282 0,672 1,029
TD/TD liczebność
0
4
6
żywotność 0,000 1,506 0,645
S- położenie naturalnej populacji (siodło między dwoma szczytami adaptacyjnymi)
Populacja powinna zatem zmierzać do jednego ze szczytów adaptacyjnych.
Powtórne badania- stan stabilny
Dodatkowy czynnik- inbred (gatunek osiadły)
Położenie populacji naturalnej odpowiada przyjętym założeniom działania selekcji i inbredu (F= 0,25)
Model zgodny z rzeczywistością!
Selekcja dotycząca cech ilościowych
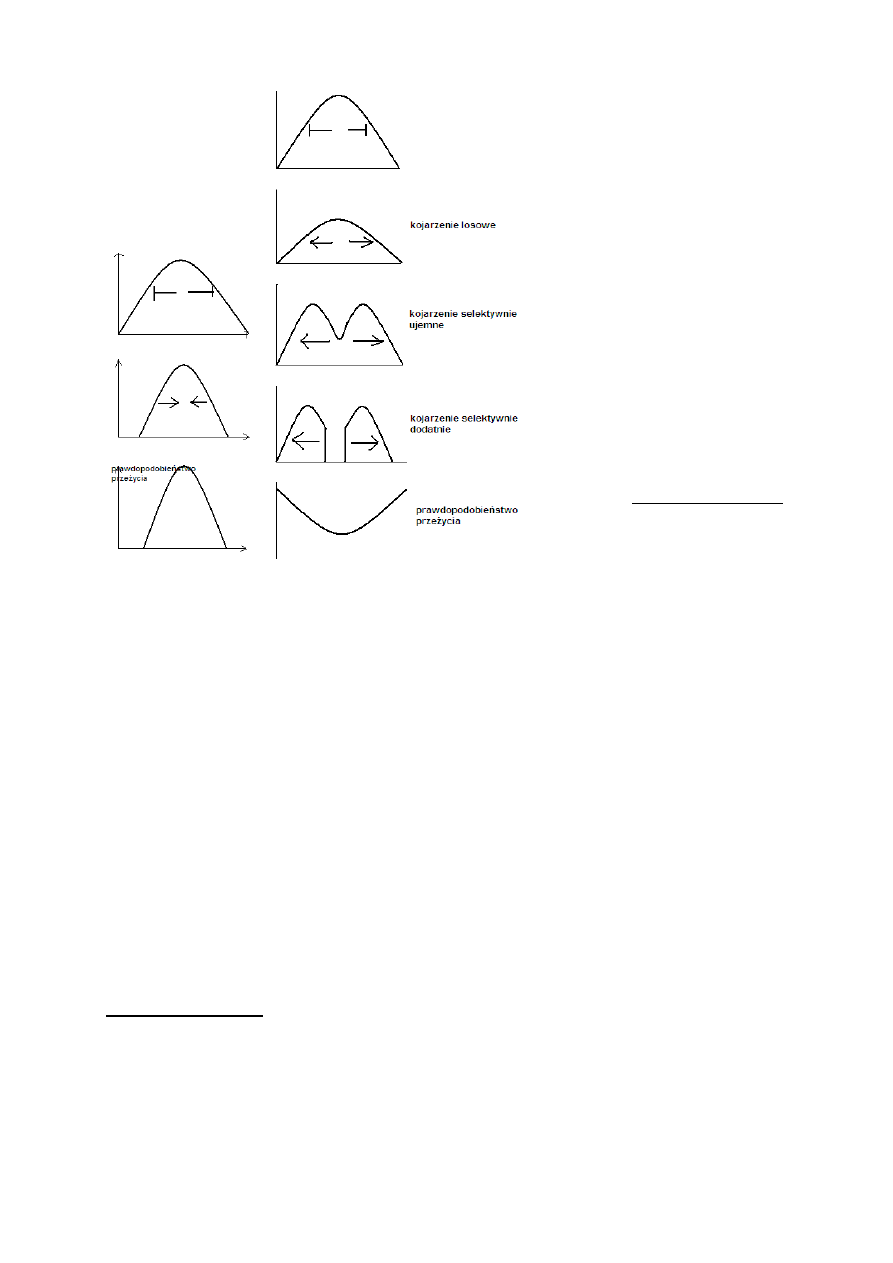
Selekcja kierunkowa
Eliminowane są osobniki o skrajnej wartości cechy.
Następuje przesunięcie średniej wartości In plus (pozytywna) lub In
minus (negatywna).
→→→A więc →→→
Przykład:
Geospiza fortis
Korelacja- większy dziób to większy ptak
Wielkość dzioba- cecha dziedziczna
1976- 1978- susza
Spadek liczebności populacji
Mało pokarmu- duże suche nasiona dostępne tylko dla osobników o dużych dziobach
Suszę przeżyły tylko duże osobniki
Po 1978- wzrost liczebności dużych osobników w populacji
36
Susze 1980,
1982- czynnik
selekcyjny
1984, 1986- rok
mokry i wilgotny;
dominują
osobniki małych
dziobach
10- 20 epizodów suszy bez
selekcji pomiędzy
spowodowałoby upodobnienie się
gatunku G. fortis do większej G.
magnirostis.
Selekcja stabilizująca
Eliminowane są osobniki o obu
skrajnych wartościach cechy.
Następuje zmniejszenie zakresu zmienności cechy bez zmiany wartości średniej.
Przykład:
Eurosta solidaginis
Galasówka (jaja głównie na nawłoci)
Geny determinują wielkość galasa
Pośrednie galasy są najczęstsze
Małe galasy- pasożytnictwo; Eurytoma obtusiventris(błonkówka) składa jaja do
galasa
Duże galasy- ptaki odżywiające się galasami (są dobrze widoczne)
człowiek
masa urodzeniowa noworodka
najczęściej ok. 7 funtów (ok. 3,178 kg)- mają najmniejszą śmiertelność)
mniejsza masa- większa śmiertelność
duża masa- komplikacje przy porodzie
Selekcja rozdzielająca
Eliminowane są osobniki o średniej wartości cechy.
Następuje zwiększenie zakresu zmienności.

37
Do kojarzenia selektywnie dodatniego może dojść ponieważ kojarzenie losowe prowadzi do niższego
dostosowania części potomstwa.
Przykład:
Drosophila melanogaster
Liczba szczecin na tułowiu
Selekcja sztuczna w laboratorium
Brano samca i samicę o największej liczbie szczecin oraz samca i samicę o najmniejszej
liczbie szczecin
Po 12 pokoleniach rozdzielono
Selekcja jest tym skuteczniejsza, im wyższa jest odziedziczalność cechy.
Odziedziczalność
Udział wariancji genetycznej w całkowitej wariancji genotypowej (w szerokim znaczeniu).
lub
Udział addytywnej wariancji genetycznej w całkowitej wariancji genotypowej (w wąskim znaczeniu)
h
2
= V
G
/ V
P
lub h
2
= V
A
/ V
P
V
P
– całkowita zmienność fenotypowa (wariancja fenotypu) = V
G
+ V
E
V
E
– wariancja środowiskowa
V
G
– wariancja genetyczna = V
A
+ V
D
+ V
I
V
A
– addytywna wariancja genetyczna (sumujące się efekty alleli poszczególnych loci; przy założeniu,
że heterozygoty mają fenotyp pośredni w stosunku do alternatywnych homozygot, a efekty wszystkich
loci są takie same)
V
D
– wariancja wynikająca ze współdziałania allelicznego (dominacja alleli)
V
I
– wariancja wynikająca ze współdziałania nieallelicznego (np. epistaza)
h
2
= 0
M - średnia wartość cechy w populacji przed działaniem selekcji
M
S
– średnia wartość cechy w populacji po działaniu selekcji
M
1
– średnia wartość cechy u osobników faworyzowanych przez selekcję
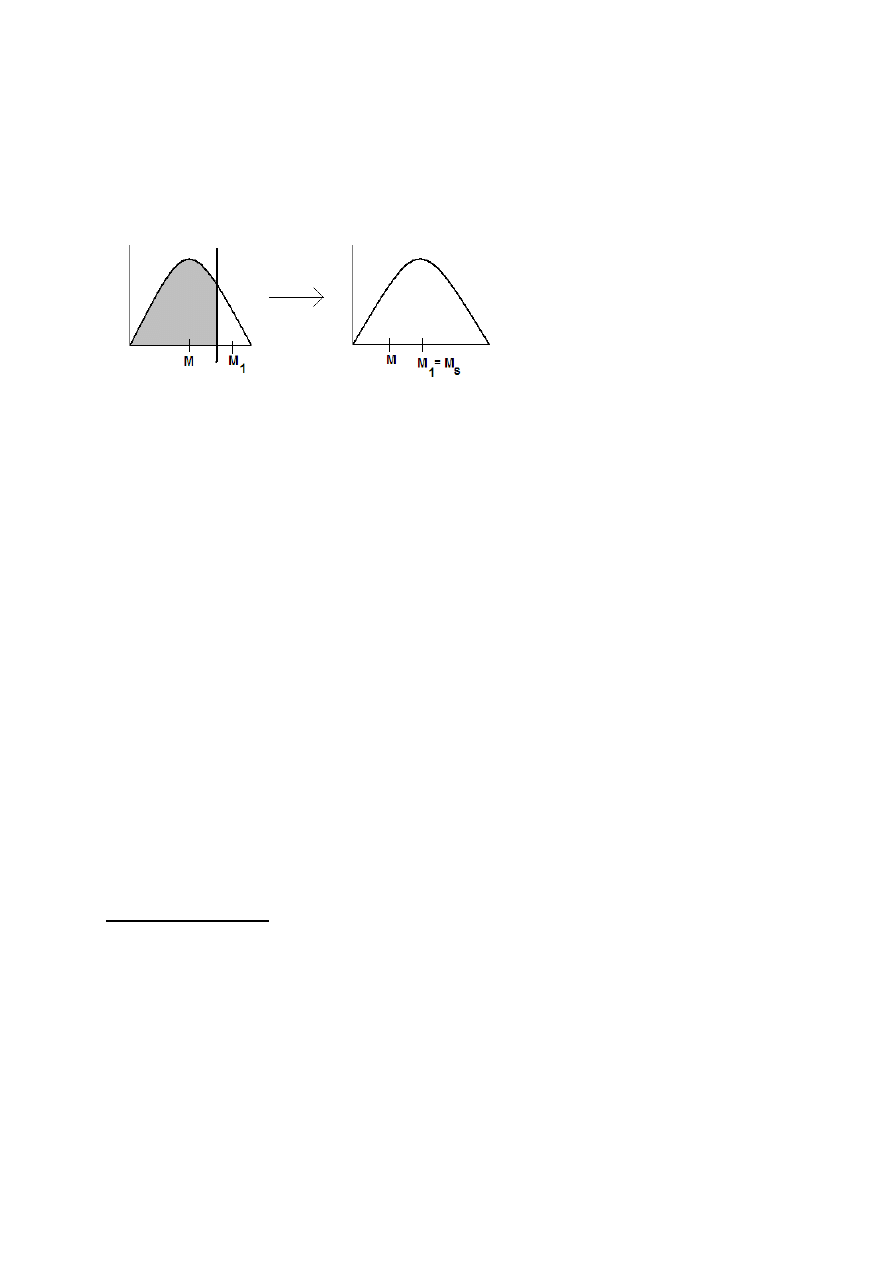
38
h
2
= 1
h
2
= ( M
S
– M ) / ( M
1
– M )
Wykład 9.
Pojęcia gatunku:
1. pojęcie gatunku nomenklatorycznego:
Okaz typowy oraz inne okazy, które specjalista odnosi do okazu typowego. Okaz typowy
wyznacza nazwę gatunku (kodeksy nomenklatury).
2. pojęcie biologiczne gatunku:
Grupa naturalnych populacji realnie lub potencjalnie się krzyżujących, która jest genetycznie
odizolowana od innych grup zdefiniowanych w ten sposób (Mayr, 1969)
3. pojęcie ewolucyjne gatunku:
Pojedyncza linia populacji utworzona przez ciąg potomków i przodków, która podtrzymuje
swoją odrębność od innych linii i wykazuje własne tendencje ewolucyjne i własne
przeznaczenie historyczne (Wiley, 1978)
4. pojęcie filogenetyczne gatunku:
Najmniejsza grupa monofiletyczna o wspólnym pochodzeniu (de Queiroz i Donoghue, 1990)
Gatunki to izolowane względem siebie pule genowe. Tylko w obrębie gatunku geny podlegają
swobodnej rekombinacji w wyniku rozrodu i mogą „przepływać” z jednej populacji do drugiej w
wyniku migracji osobników lub gamet. Wymiana taka nie zachodzi między populacjami różnych
gatunków.
Mechanizmy izolujące:
Zapobiegające wymianie genów pomiędzy populacjami różnych gatunków)
1. Mechanizmy prezygotyczne (działają przed połączeniem gamet):
a) Izolacja ekologiczna lub środowiskowa (potencjalni partnerzy mimo zasiedlania
wspólnego obszaru nie spotykają się, gdyż zajmują odmienne siedliska)
b) Izolacja sezonowa (okresy kwitnienia lub kojarzenia się są przesunięte względem
siebie w rytmie dobowym lub w większych okresach czasu)
c) Izolacja etologiczna lub płciowa (potencjalni partnerzy się spotykają ale wykazują
odmienne zachowanie przy kojarzeniu)
d) Izolacja mechaniczna (mimo prób odbycia kopulacji nasienie nie zostaje
wprowadzone ze względu na odmienność genitaliów)
e) Izolacja gametyczna (gamety są przekazywane, ale nie dochodzi do ich połączenia)
2. Mechanizmy postzygotyczne (działające po połączeniu gamet):

39
a) Śmiertelność zygot
b) Letalność lub obniżona żywotność mieszańców
c) Całkowita lub częściowa sterylność mieszańców
d) Depresja mieszańcowa F2 (mieszańce produkują upośledzone potomstwo)
Izolacja ekologiczna
Np. Epilachna yasutomi i Epilachna niponica (Coccinellidae)
Epilachna yasutomi- całe życie na Caulophylum robustum (Berberidaceae)
Epilachna niponica- żyje na Cirsium japonicum (Asteraceae)
Jedyny mechanizm izolacyjny!
Eksperymenty krzyżówkowe nie wykazały żadnej innej izolacji- po umieszczeniu owadów w
jednym miejscu mogą się swobodnie krzyżować
Izolacja sezonowa
Np. cykle życiowe siedmiu gatunków świerszczy Gryllus (Gryllidae, Orthoptera)
G. veletis- dojrzałe osobniki występują latem
G. pennsylvanicus- dojrzałe osobniki występują jesienią
Blisko spokrewnione gatunki uzyskują dojrzałość płciową w różnych porach roku
Izolacja etologiczna
Np. sygnały akustyczne nieodróżnialnych morfologicznie gatunków złotooków Chrysoperla
(Neuroptera, Chrysopidae)
Ch. plorabunda P1
Ch. plorabunda P2 = Ch. adamsi różne widma akustyczne (oscyllodramy)
Ch. plorabunda P3 = Ch. johnsoni
Sygnały akustyczne u dwóch gatunków północnoamerykańskich cykad Tibicen (Hemiptera,
Cicidodae)
Tibicen linnei
Tibicen pruinosa
Izolacja mechaniczna
Np. różnice w morfologii kwiatów podkolanów białego (Platanthera bifolia) i zielonawego (P.
chlarantha) (Orchidaceae) warunkują zapylanie przez inne gatunki motyli
P. biforia- Sphingidae
P. chlorantha- Noctuidae
Izolacja gametyczna
Np. niezgodność gametyczna u ślimaków morskich o zapłodnieniu zewnętrznym- uchowce Haliotis
(Archeogastropoda, Haliotidae)
Plemniki zawierają gatunkowo specyficzne enzymy trawiące osłony komórek jajowych samic jedynie
własnego gatunku.
Duże międzygatunkowe zróżnicowanie sekwencji DNA kodujących białka enzymatyczne plemników
u blisko spokrewnionych gatunków.
Śmiertelność zygot
Np. tzw. „odczyn inseminacyjny”- plemniki obcego gatunku w drogach rodnych samicy wywołują
reakcję antygenową prowadzącą do unieruchomienia i zniszczenia gamet (szereg gatunków
Drosophila)
Śmiertelność i/lub sterylność mieszańców (F1, F2)
Np. muł (63 chromosomy)- mieszaniec międzygatunkowy klaczy konia domowego (64 chromosomy)
i ogiera osła (62 chromosomy). Niemal bezpłodny, tylko 5% mulic jest płodnych, samce całkowicie
bezpłodne.
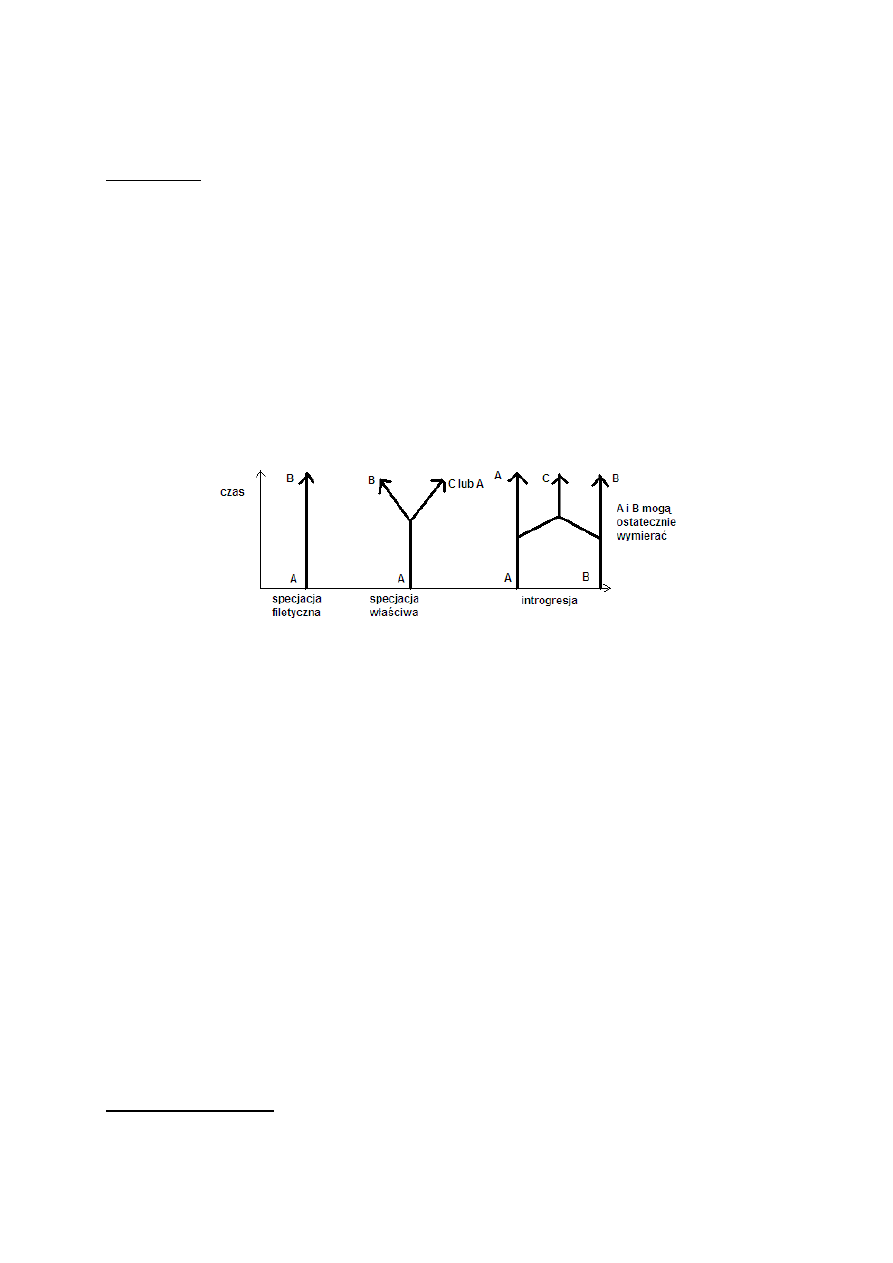
40
Osłomuł- mieszaniec międzygatunkowy klaczy osła z ogierem konia domowego. Całkowicie
bezpłodne.
SPECJACJA- proces powstawania gatunków
Klasyfikacja procesów specjacyjnych:
Podział ze względu na liczbę powstających gatunków:
Specjacja filetyczna- stopniowe, całościowe przekształcanie się jednego gatunku w
drugi- liczba gatunków się nie zmienia
W większej skali ewolucyjnej ciąg specjacji filetycznej bez rozszczepienia danej linii
rozwojowej- anageneza
Gatunki wydzielane w szeregu anagenetycznym- Chronospecies
Specjacja właściwa (radiacyjna)- rozdzielenie gatunku macierzystego na dwa lub więcej
gatunki potomne (promieniste rozejście się linii rozwojowej)- liczba gatunków wzrasta
W większej skali ewolucyjnej ciąg specjacji radiacyjnych- kladogeneza
Specjacja introgresywna- zlanie się dwóch gatunków- liczba gatunków może się
zmniejszać
Wykształcenie linii ewolucyjnych układających się w kształt „sieci”- specjacja siatkowa
Podział ze względu na tempo zachodzenia:
Specjacja skokowa (nagła)- krótka seria pokoleń na poziomie osobników
Specjacja stopniowa- seria pokoleń w funkcji czasu na poziomie populacji, kumulowanie
drobnych różnic w pulach genowych zwykle izolowanych populacji
Podział ze względu na stosunki przestrzenne między populacjami:
Specjacja allopatryczna (= geograficzna)- zachodzi w warunkach izolacji geograficznej
a)
Specjacja dychopatryczna (vicariant speciation)
Przez:
o
Rozdzielenie dwóch szeroko rozprzestrzenionych populacji barierą
geograficzną
o
Zanikanie pośrednich populacji w ciągłym łańcuchu populacji (skrajne
populacje mogły wcześniej już uzyskać izolację rozrodczą)
o
Przez migrację osobników na odrębne obszary
b)
Specjacja perypatryczna- przez izolację niewielkiej grupy osobników na
skraju zasięgu szeroko rozprzestrzenionej populacji macierzystej,
wykształcającej następnie mechanizmy izolujące.
Specjacja parapatryczna (semigeograficzna)- wykształcenie izolacji rozrodczej na
brzegach zasięgów między populacjami, które geograficznie stanowią pewien ciąg,
zwykle poprzez adaptację do odmiennego środowiska- szczególne podkreślenie różnic na
brzegach zasięgów.
Specjacja sympatryczna- bez izolacji geograficznej
Specjacja allopatryczna
Ewolucja izolacji reprodukcyjnej między populacjami, które są przestrzennie rozdzielone przez
bariery fizyczne

41
Dowody:
Geograficzne prawidłowości występowania i zgodności z obecnym lub dawnym
występowaniem barier
Eksperymenty laboratoryjne
Przykład:
Czarnowron (Corvus corone)- zachodnia część Europy
Wrona siwa (Corvus cornix)- wschodnia część Europy, Włochy, północna Szkocja
Mieszańce- żywotne, ale bezpłodne, np. wzdłuż Łaby, rzeki Jenisej (ale tu już nie z czarnowronem)
Nadgatunek (superspecies)- monofiletyczna grupa całkowicie (lub w znacznej części)
allopatrycznych gatunków, które są zbyt różne morfologicznie aby zaliczyć je do jednego gatunku,
albo wykazują izolację rozrodczą na styku zasięgów. Nadgatunek nie jest kategorią systematyczną i
nie nadaje mu się nazwy!
Półgatunek (semispecies)- gatunek wchodzący w skład kategorii „nadgatunek” lub populacja, która
nabyła pewne, ale nie wszystkie atrybuty rangi gatunku (na pograniczu gatunku i podgatunku)
Przykład:
Wrona i czarnowron
Ok. 53 nadgatunki wśród ptaków północnoamerykańskich
Nur pacyficzny (Gavia pacyfica)
Nur czarnoszyi (Gavia arctica)
Inne przykłady par gatunków powstałych na drodze specjacji allopatrycznej:
Jeż zachodni i wschodni (Erinaceus europaeus i Erinaceus concolor)
Pełzacz ogrodowy i leśny
Dzięcioł zielony i zielonosiwy
Kumak nizinny i górski
Rodentia, Sciuridae
Scurius aberti aberti- północ Wielkiego Kanionu
Scurius aberti kaibabensis- południe Wielkiego Kanionu
Gambusia affinis affinis i Gambusia affinis holbrooki- macierzysty teren to północny Meksyk
W. Missisipi SE USA
G. a. affinis
G. a. holbrooki w strefie sympatrii
liczba promieni w płetwie grzbietowej 7
8
7/8
liczba promieni w płetwie odbytowej
9
10
9/10
ząb na gonopodium
brak
obecny
+/-
G. a. affinis- ♀ (ZW), ♂ (0)- chromosomy płci nie zaznaczone u samców
G. a. holbrooki- ♀ (0), ♂ (0)- chromosomy płci nie zaznaczone dla obu płci
W strefie sympatrii brak chromosomów płci:
♀ G. a. affinis (ZW) × ♂ G. a. holbrooki (0)- zarodki i larwy nieżywotne
♀ G. a. holbrooki (0) × ♂ G. a. affinis (0)- potomstwo płodne
Wniosek- zaczątkowy mechanizm bariery postzygotycznej!
Platan zachodni (Platanus occidentalis) i platan wschodni (P. orientalis)- platanowate
(Platanaceae)
Rozpowszechniony w Palearktyce, przez lodowiec zepchnięte do wschodniej Ameryki
Północnej i regionu Europy północno- wschodniej
Oddzielone od siebie od ponad 20 mln lat dają nadal płodne mieszańce- platany klonolistne
(P. x hybrida= P. x hispanica)
Tulipanowiec amerykański (Liriodendron tulipifera)- Ameryka Północna
Tulipanowiec azjatycki (L. chinense)- Chiny, Wietnam
Magnoliowate (Magnoliaceae)
Rodzaj o pierwotnym rozmieszczeniu okołobiegunowym podczas zlodowaceń plejstoceńskich
wymarł.
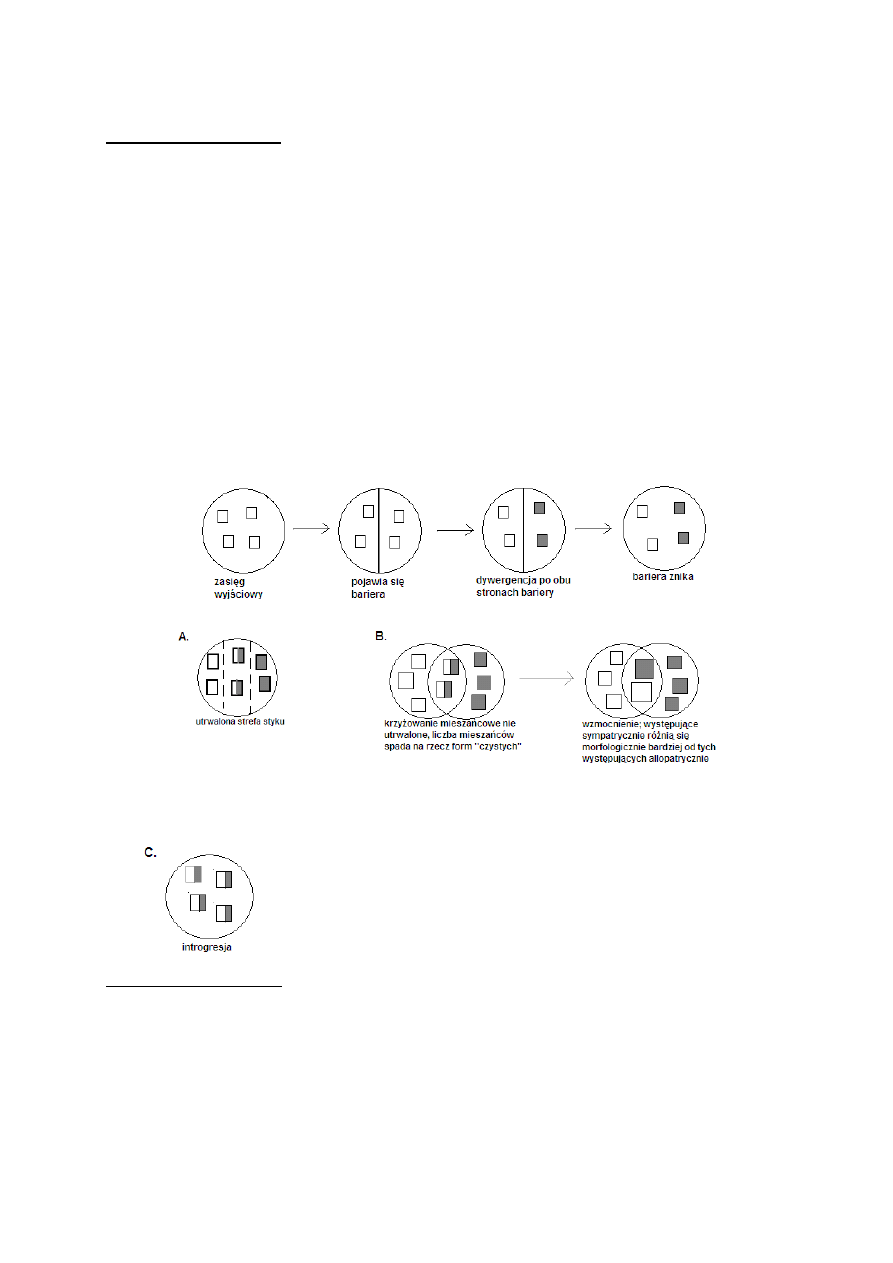
42
Specjacja perypatryczna
Nowy gatunek powstaje jako niewielka populacja odizolowana od dużej macierzystej. Dywergencja
genetyczna w stosunku do populacji macierzystej:
Początkowo na skutek efektu założyciela i dryfu genetycznego
Z czasem pogłębiona przez odmienne w sile i kierunku naciski selekcyjne działające w
odmiennych środowiskach
Przykład:
Szereg endemitów wyspowych:
Sterowik białobrzuchy (Tanisiptera galatea)- kakaburowate (Dacelonidae) na Nowej Gwinei i
okolicznych wyspach
Zięby Darwina (Geospizinae) na Galapagos i Wyspie Kokosowej
500 opisanych (+ 500) gatunków Drosophila na Wyspach Hawajskich
Buszówka brązowa (Acanthiza pusilla) i rdzawoczarna (A. ewingi)- Pardalotidae, Australia i
Tasmania
Szereg karłowatych gatunków słoni i mamutów zamieszkujących w plejstocenie wyspy na
Morzu Śródziemnym
Specjacja parapatryczna
Ewolucja izolacji reprodukcyjnej między populacjami, które są tylko częściowo izolowane
geograficznie (ich zasięgi częściowo się pokrywają)
Zwykle populacje gatunku szeroko rozprzestrzenionego, stanowiące pewien ciąg, różnicują się
poprzez adaptację do odmiennego środowiska (różne upodobania pokarmowe, różne okresy
aktywności, dojrzałości płciowej) w wąskim geograficznym pasie.
Przykład:
tomka wonna Anthoxanthum odoratum (Poaceae)
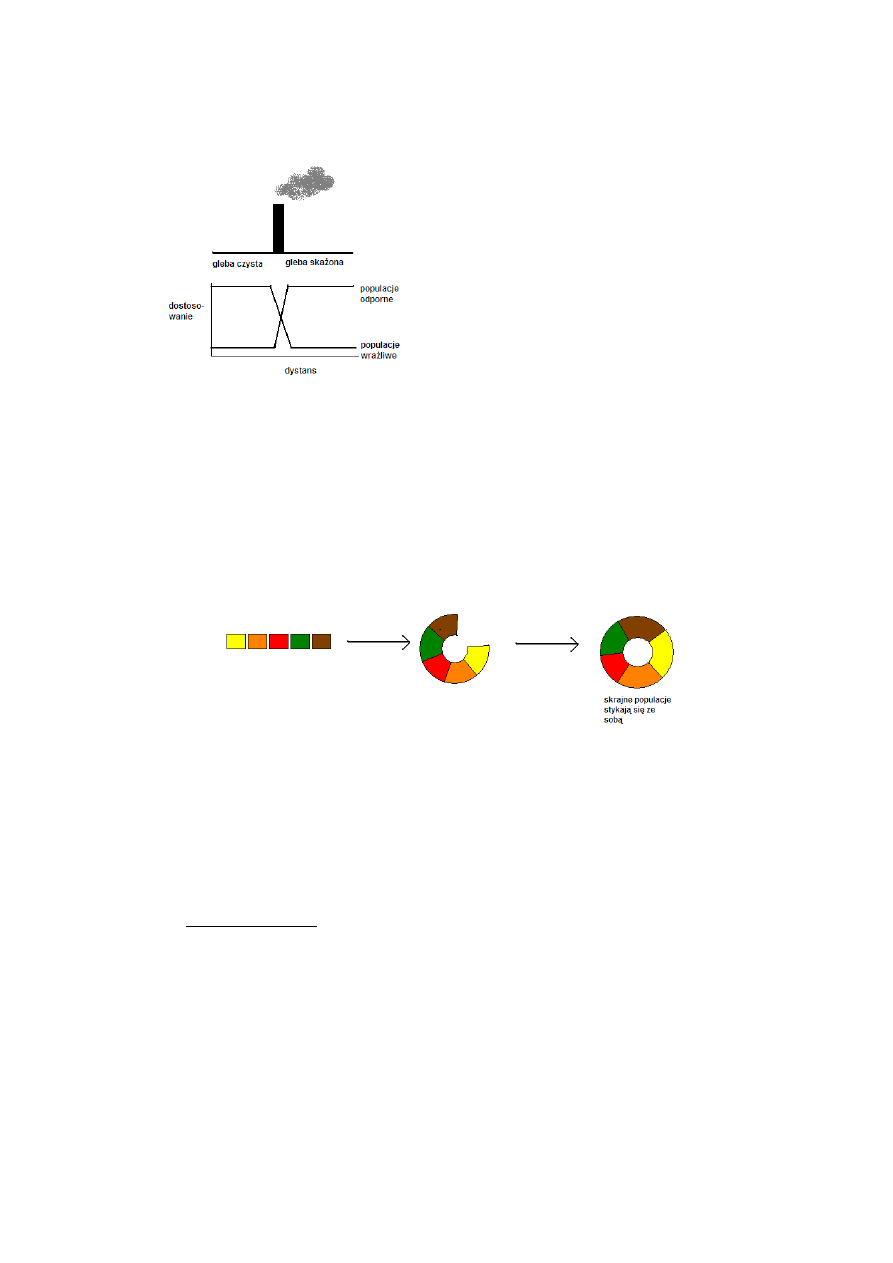
43
jeden z gatunków traw, który w ciągu ostatnich paru stuleci wykształcił odporność na skażenie
metalami ciężkimi Pb i Zn w pobliżu kopalni
Populacje odporne i wrażliwe odróżnicowały się także pod względem:
a) czasu kwitnienia (później zakwitają te rosnące na glebie skażonej)
b) zdolność do produkcji nasion pochodzących z samozapylenia (samozapylenie częstsze u roślin
odpornych)
Wniosek: znaczna izolacja reprodukcyjna między sąsiadującymi populacjami.
Wykład 10.
Gatunki pierścieniowe- przykład specjacji parapatrycznej
przepływ genów między populacjami sąsiadującymi
brak przepływu między populacjami skrajnymi
Przykład:
Salamandry Ensantina spp. (Plethodontidae) wokół Central Valley w Kaliforni
klauberi i eschsantii- nie krzyżują się
Kompleks gatunków mew Larus spp. (Laridae) wokół Bieguna Północnego
migracje z refugium M. Kaspijskiego
mewa żółtonoga i srebrzysta- nie krzyżują się
model alternatywny- dwa refugia (atlantycki i kaspijski)
według modelu gatunki Larus spp. nie są gatunkami pierścieniowymi
model pierwotny zakładający pojedyncze aralsko- kaspijskie refugium i stosunkowo
niedawną inwazję Larus argentatus z Ameryki Północnej do Europy, gdzie spotyka się z
Larus fuscus (izolacja reprodukcyjna)
model alternatywny na podstawie mtDNA (cyt b) zakładający:
a)
dwa refugia- aralsko- kaspijskie i atlantyckie
b)
brak inwazji L. argentatus z Ameryki Północnej do Europy
jeśli L. fuscus greallsii skolonizuje Amerykę Północną i będzie izolowana reprodukcyjnie
z L. smithsoniatus- pierścień się zamknie
świstunka zielonawa (Phylloscopus trochiloides)
wokół Himalajów

44
P. t. viridanus i P. t. plumbeitarsus- stykają się, ale nie krzyżują
Dwa szlaki migracji- potwierdzone analizą śpiewu (dwie linie rozwojowe z jednego
gatunku)
Rozgałęzienia wschodnie i zachodnie dzielą się dodatkowo na północne i południowe
(północne- osobniki mniejsze; południowe- osobniki większe, bo rzadsze zarośla)
10000 lat- migracja!
Przykład par gatunków powstałych na drodze specjacji allopatrycznej w Europie:
Kumak nizinny (Bombina bombina)- czerwone plamy + białe plamki;
Kumak górski (B. variegata)- żółte plamy zlewające się
Strefy mieszańcowe- Pescenica, Chorwacja; ok. 30 km- gwałtowna zmiana frekwencji
alleli
Jeż zachodni (Erinaceus europaeus)- brązowe włosy na brzuchu;
Jeż wschodnioeuropejski (E. concolor)- jasne włosy na brzuchu
Są zróżnicowane pod względem genetycznym (zróżnicowanie między- i
wewnątrzgatunkowe)
Zróżnicowanie intraspecyficzne wskazuje na:
a) Okresy ograniczonej wielkości populacji i ograniczonego zasięgu występowania w
refugiach glacjalnych
b) Odmienne szlaki postglacjalnej ekspansji
Wyniki badań mtDNA wyraźnie dzielą oba gatunki na parę monofiletycznych grup
populacji, wyniki badań nad MHC wskazują na taki podział jedynie u E. concolor
Wniosek- historia i lokalizacja refugiów oraz postglacjalnej ekspansji u obu gatunków
różniły się, a E. concolor najprawdopodobniej nie reprezentuje jednego gatunku
Wniosek- populacje ze środkowej i wschodniej Europy reprezentują dwa odrębne gatunki
Jeż wschodni (E. roumanicus)- początkowo jako podgatunek E. europaeus, później
podgatunek E. concolor; obecnie na podstawie badań genetycznych i morfologicznych
traktowany jako odrębny gatunek o wschodniej granicy zasięgu wzdłuż rzeki Ob w
zachodniej Syberii
Specjacja sympatryczna (stopniowa)
Ewolucja izolacji reprodukcyjnej bez izolacji geograficznej w obrębie (początkowo) losowo
kojarzącej się populacji (poddawana w wątpliwość)
Przykład 1.
Enchenopa binatata (Homoptera, Membracidae, zgarbkowate)
9 gatunków bliźniaczych związanych z innym gatunkiem żywicielskim rośliny
Izolacja reprodukcyjna zainicjowana przez czynnik zewnętrzny (roślinę żywicielską)
Jaja składane do wnętrza gałązek, stąd rozwój larw warunkowany jest uruchomieniem
przepływu substancji odżywczych w rurkach sitowych roślin
Skoczki związane z dławiszem amerykańskim (Celastrum scandens) wcześniej wykluwają się
z jaj i wcześniej dojrzewają płciowo (o ok. 10 dni) od tych związanych z wiązowcem (Celtis
canadensis), a samce wytwarzają dźwięki o niższej częstotliwości
Przykład 2.
Nasionnica jabłoniowa Rhagoletis pomonella (Diptera, Tripedidae, nasiennicowate)
Imagines w VII- VIII kojarzące się na określonym gatunku żywicielskim rośliny (pierwotnie
w USA na głogu Crateagus)
Larwy rozwijają się w dojrzałych owocach, później wraz z nimi spadają na ziemię i zimują
jako poczwarki
Po wprowadzeniu do Ameryki upraw drzew owocowych, w 1860 po raz pierwszy zauważona
jako szkodnik jabłoni w NE USA (później na gruszach, wiśniach i różach)
Niepełna izolacja sezonowa!

45
Imagines wychodzą z poczwarek i rozmnażają się zanim dojrzeją owoce, owady związane z
jabłoniami rozwijają się o ok. 1,0- 1,5 miesiąca wcześniej od tych związanych z głogiem
Frekwencja alleli w szeregu loci znacznie się różni pomiędzy rasami związanymi z różnymi
roślinami żywicielskimi; i to w ten sam sposób zarówno u larw, jak i imagines, które łapane
były na drzewach po wyjściu z poczwarek (preferencje do gatunku żywicielskiego)
W larwach nasionnicy odżywiających się owocami jabłoni stwierdza się o 70% mniej jaj
pasożytniczych os niż w larwach rozwijających się w owocach głogu (owoce jabłoni są
większe od owocu głogu)
Nasionnica składa więcej jaj w owocach jabłoni niż w owocach głogu (większe zasoby
pokarmowe na jabłoniach), ale prawdopodobieństwo rozwoju obu „ras” nasionnicy jest
większe w owocach głogu (57% vs 27% w owocach jabłoni); większa wartość odżywcza
głogu
Obecnie specjalizacja ta jest na tyle posunięta, że kojarzenia mieszane stanowią jedynie 4- 6%
tzn. przepływ genów jest silnie zredukowany (specjacja w toku)
Specjacja sympatryczna (skokowa)
Specjacja przez hybrydyzację i poliploidalność
Jedyny nie budzący większych wątpliwości mechanizm specjacji bez bariery rozdzielającej populacje i
stosunkowo krótkotrwały, gdyż forma poliploidalna uzyskuje natychmiastową izolację rozrodczą w
stosunku do formy wyjściowej, ponieważ mieszańce między 2n i 4n są zwykle sterylne lub
nieżywotne
Poliploidalność odgrywa bardzo dużą rolę w specjacji u roślin
Ok. 50% okrytozalążkowych
1,5% nagozalążkowych (!?)
poliploidy
95% paprotników
Poliploidy odkrywamy poprzez obserwację zmienności liczby chromosomów u gatunków
spokrewnionych (wielokrotności lub multiwalenty w mejozie)
Np. poszczególne gatunki złocieni Chrysanthemum (Asteraceae) stanowiące serię wielokrotności
wyjściowej liczby 9: 2n= 18, 36, 54, 72, 94
Przykłady ze świata roślin:
Wiesiołek Oenothera (Onagraceae)- de Vries (1905)
Wśród osobników wiesiołka czerwonokielichowego Lamarcka (Oenothera lamarckiana)
o 2n= 14 zaobserwował osobniki o 2n= 28, które nie krzyżowały się z populacją
wyjściową (mutant nazwany O. gigas)
pierwiosnek Primula (Primulaceae)
W wyniku eksperymentalnej hybrydyzacji P. verticillata i P. floribunda w londyńskim
ogrodzie botanicznym Kew w 1929 powstała niewielka liczba sterylnych mieszańców a
następnie, po poliploidyzacji pojawiło się kilka płodnych tetraploidów 2n= 36 nazwanych
P.kewensis.
Kilkakrotnie (1905, 1923, 1926) obserwowano spontanicznie powstałe mieszańce P.
verticillata i P. floribunda dające tetraploidalne nasiona
Zawilec Anemone (Ranunculaceae)
A. rivularis i inne gatunki rodzaju 2n= 16
A. quinquefolia 2n= 32
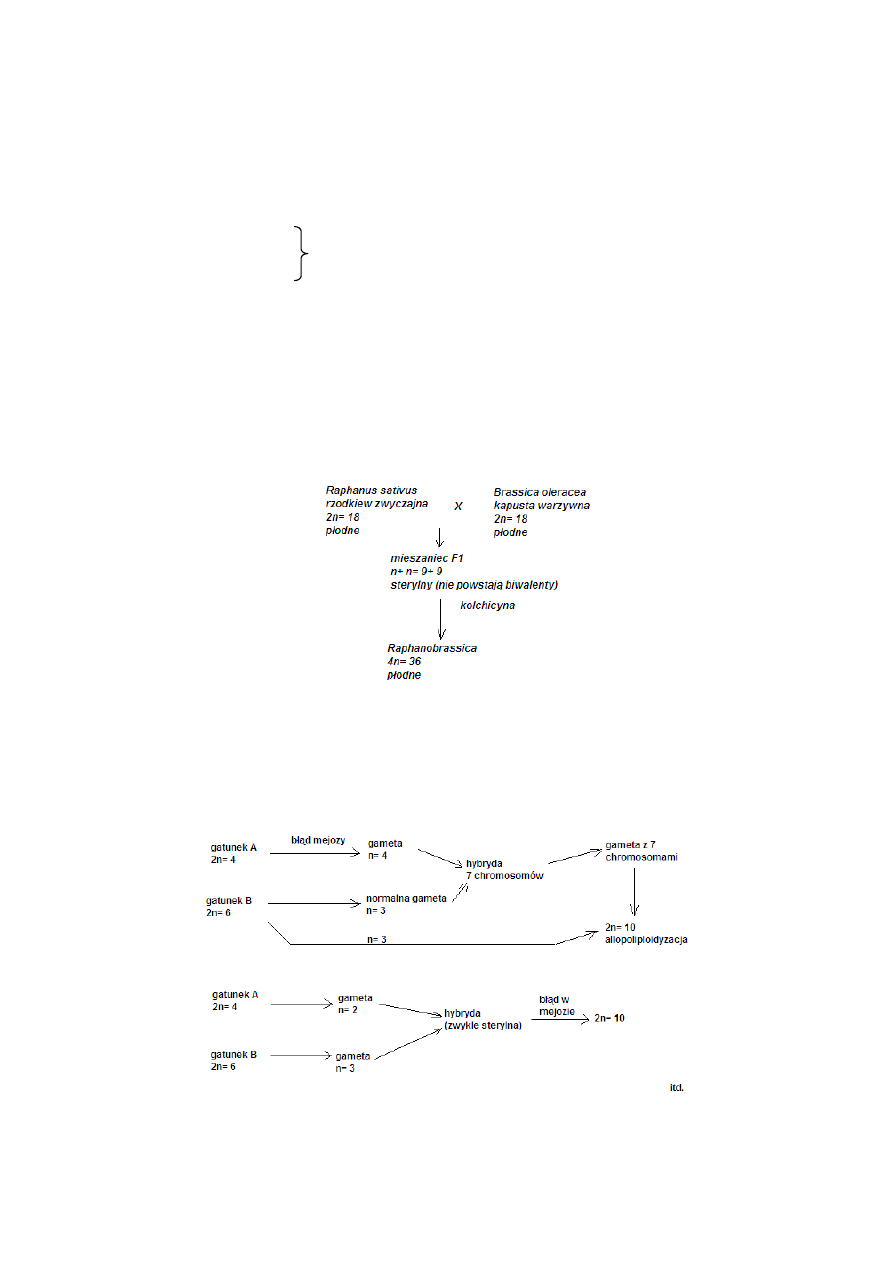
46
Wniosek- A. quinquefolia powstał przez poliploidyzację
Gilia (Polemoniaceae)
Mutacje chromosomowe po hybrydyzacji i poliploidyzacji różnicujące skład
chromosomowy
Kozibród Tragopogon (Asteraceae)
Gatunki indukowane do USA na początku XIX w.
T. dubius
T. porrifolius
2n
T. prathensis
gatunki mieszańcowe 4n
T. mirus= T. porrifolius (♀♂) + T. dubius
T. miscellus= T. prathensis (♀) + T. dubius (♂)
T. mirus i T. miscellus powstały wskutek poliploidyzacji mieszańców
Analizy morfologiczne, cytologiczne, izozymów, chlDNA i rDNA potwierdza 9
niezależnych gatunków względem pochodzenia linii T. mirus i 21 T. miscellus
Raphanobrassica (Brasicaceae)- Karpechenko 1927
Krzyżówka międzyrodzajowa!
Niestety uzyskany tetraploid miał liście rzodkwi i korzeń kapusty ;)
Krzyżówki tego typu:
Raphanus sativus × Brassica rapa (rzepa)
Raphanus caudatus × Brassica tourneflori
Możliwe
drogi
powstania
płodnych
gatunków
mieszańcowych w wyniku allopoliploidalności:

47
Ewolucja heksaploidalnej pszenicy Triticum aestivum
14 AA × 14 BB → 14 AB (wielokrotna poliploidyzacja i hybrydyzacja)
28 AABB × 14 DD → 48 AABBDD
Specjacja przez hybrydyzację u zwierząt
Przykład:
Żaby zielone Rana esculenta complex (specjacja w toku)
Rana ridibunda- żaba śmieszka
Rana lessonae- żaba jeziorkowa
Rana esculenta- żaba wodna
Bardzo trudno odróżnialne zarówno pod względem ubarwienia, jak i cech morfometrycznych
(np. długość podudzia, wielkość modzeli piętowych)
Odmienne preferencje środowiskowe:
Rana ridibunda- zbiorniki o stałym przepływie wody, zimuje na dnie
Rana lessonae- płytkie stawy, zimuje na lądzie
Rana esculenta- zróżnicowane zbiorniki wodne, zimuje na lądzie lub w wodzie
Spotykają się populacje wyłącznie R. ridibunda i wyłącznie R. lessonae, ale niemal nigdy nie
występują sympatrycznie
Rana esculenta jest mieszańcem R. ridibunda i R. lessonae. Nie powstaje jednak stale z
krzyżówek międzygatunkowych
Jeśli R. esculenta żyje z R. lessonae, u R. esculenta jeszcze przed mejozą usuwane są
chromosomy (hybrydogeneza) R. lessonae, tak że w gametach występują jedynie chromosomy
R. ridibunda.
Genotyp mieszańcowy odtwarzany z pokolenia na pokolenie
R. esculenta obejmuje 3 formy osobnicze, które różnią się pomiędzy sobą składem
chromosomowym:
RL (2n= 26)- RL× RL (F1 letalne)
LLR (3n= 39)
RRL (3n= 39)
Triploidy są płodne, produkują zwykle jaja 2n, które łączą się z plemnikiem n
Granica przebiega wzdłuż rzeki Świny, na zachodzie i północy (Niemcy, Dania) R. esculenta
jest płodna, na wschodzie (Polska)- niepłodna.
Specjacja filetyczna
Filogeneza i tempo specjacji 7 gatunków otwornic z rodzaju Globorotalia w południowo- zachodnim
Pacyfiku w ciągu ostatnich 7 mln lat:
5 zastąpień filetycznych (anageneza)
2 specjacje radiacyjne (kladogeneza)

48
Sekwencja gatunków otwornic w równych interwałach czasowych (co 500 tys. lat) w jednym profilu
osadów morskich.
Najważniejszą cechą procesu specjacji jest gromadzenie się takich genetycznych różnic, które
prowadzą do izolacji rozrodczej rozszczepiających się gatunków.
Mechanizmy wyjaśniające ewolucję międzygatunkowych barier rozrodczych:
1. dryf genetyczny
Drosophila pseudoobscura- wielokrotna redukcja liczebności (Powell 1978, Dodd i
Powell 1986)
Drosophila mercantonum- ekstremalne obniżanie liczebności (Templeton 1979, 1983)
2. selekcja naturalna
selekcja działająca pośrednio na cechy determinujące izolację rozrodczą (Hipoteza
Müllera- Mayra)
bariery reprodukcyjne to efekt uboczny działania doboru naturalnego w czasie
przestrzennej izolacji. Genetyczna dywergencja to jedynie wynik dopasowania różnic
genetycznych do odmiennych warunków środowiskowych, a to dopiero pociąga za sobą
powstanie zarówno pre- jak i postzygotycznych barier rozrodczych. Nie wyklucza się
wzmocnienia barier prezygotycznych po zniknięciu izolacji przestrzennej, ale jest to tylko
nasilenie barier już istniejących.
o D. pseudoobscura- nietypowe rodzaje pożywki (Dodd 1989)
Czy mogła wykształcić się izolacja rozrodcza?
Policzono liczbę 4 rodzajów kojarzeń między muszkami hodowanymi na maltozie
i skrobi i porównano z kontrolą.
Wskaźnik izolacji:
I= (H
S
– H
O
)/ N
H
S
- liczba kojarzeń między osobnikami z tych samych warunków
H
O
- liczba kojarzeń mieszanych
N- liczba wszystkich kojarzeń
♀
♂
skrobia maltoza
skrobia
22
9
maltoza
8
20
Przykładowy eksperyment: I= (42 – 17)/ 59= 0,42
ta sama linia różne klatki
ta sama linia 18
15
różne klatki 12
15
Przykładowa kontrola: I= (33 – 27)/ 66= 0,09
Średnia wartość wskaźnika izolacji po wszystkich krzyżówkach:
Krzyżówka skrobia × maltoza: I= 0,33
Krzyżówka kontrolna: I= 0,01
Wniosek- prezygotyczna izolacja reprodukcyjna powstała na skutek selekcji do odmiennych
warunków środowiska.
Przykłady innych doświadczeń tego typu:
o D. melanogaster- różne temperatury i wilgotność

49
o D. melanogaster- foto- i geotaksja
o Musca domestica- ujemna i dodatnia geotaksja
Wykład 11.
c.d. do selekcji naturalnej
Selekcja działająca bezpośrednio na cechy determinujące izolację rozrodczą (Hipoteza
Fishera- Dobzhansky’ego)
Bariery rozrodcze to skutek bezpośredniego działania na nie doboru naturalnego w czasie
ponownego zetknięcia się gatunków siostrzanych. Bariery postzygotyczne nabyte w
okresie allelopatrii poprzedzają powstanie barier prezygotycznych. Selekcja eliminuje
krzyżujące się genotypy rodzicielskie ze względu na obniżone dostosowanie mieszańców.
o Drosophila mojavensis i D. arizonae- gatunki bliźniacze o częściowo
pokrywających się zasięgach.
Jeśli hipoteza Fishera- Dobzhansky’ego jest prawdziwa, to należy oczekiwać
większego nasilenia izolacji prezygotycznej w strefie sympatrii (okolice Sonory w
Meksyku) niż między populacjami allopatrycznymi.
Testy wyboru:
♀ jednego gatunku umieszczona z ♂ własnego i innego gatunku
♂ jednego gatunku umieszczony z ♀ własnego i innego gatunku
Wskaźnik izolacji prezygotycznej:
I= (H
S
– H
O
)/ N
H
S
- liczba kojarzeń między tymi samymi gatunkami
H
O
- liczba kojarzeń mieszanych
N- liczba wszystkich kojarzeń
♀ D. mojavensis × ♂ D. arizonae ♀ D. arizonae × ♂ D. mojavensis
♀ D. mojavensis
♀ D. arizonae
allopatria I= 0,30
allopatria I= 0,90
sympatria I= 0,94
sympatria I= 0,80
♂ D. arizonae
♂ D. mojavensis
allopatria I= 0,54
allopatria I= 0.78
sympatria I= 0,60
sympatria I= 0,92
Wniosek- kojarzenia mieszane występują znacznie rzadziej między gatunkami w strefie sympatrii.
Przesunięcie cechy, wzmocnienie (character displacement, reinforcement)- większe różnice w
sympatrii w porównaniu z allopatrią.
Krytyka- nierównomierna fuzja i wymieranie gatunków
Szereg allopatrycznych gatunków siostrzanych. U jednych silniejsza izolacja prezygotyczna, u innych
słabsza. Gatunki wchodzą w ponowny konflikt. Pary, które w allopatrii wytworzyły słabą izolację, w
sympatrii będą się zlewały w jeden gatunek. Rozdzielone zostaną tylko te, które wcześniej wytworzyły
silne mechanizmy izolujące.
Drosophila spp.- 119 gatunków
1. zróżnicowanie genetyczne
dystans genetyczny Neia (D)- miara proporcjonalna do liczby podstawień aminokwasów w
badanych elektroforetycznie białkach oraz do czasu, który upłynął od momentu rozszczepienia
się dwóch gatunków.
2. rozmieszczenie geograficzne (allo- i sympatria)
3. pre- i postzygotyczna izolacja
izolacja prezygotyczna= 1-(H
O
/ H
S
)
H
O
- częstość kojarzeń międzygatunkowych
H
S
- częstość kojarzeń wewnątrzgatunkowych
Izolacja postzygotyczna- stopień obniżenia żywotności lub płodności mieszańców F1
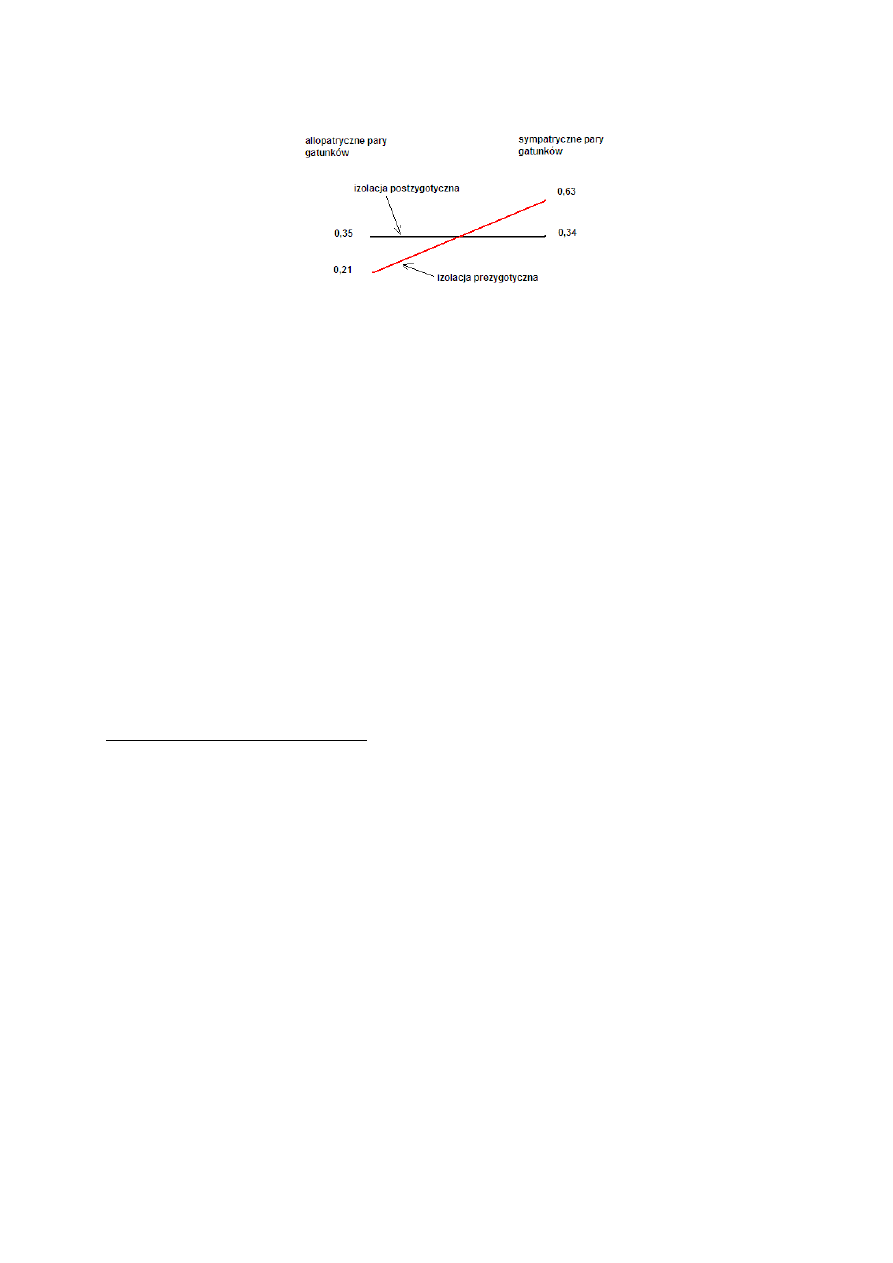
50
Wnioski-
dla gatunków sympatrycznych wartość wskaźnika izolacji prezygotycznej jest około trzy razy
wyższa niż dla gatunków allpatrycznych, podczas gdy wartość wskaźnika izolacji
postzygotycznej są takie same
można odrzucić krytykę o fuzji gatunków ze słabo wykształconymi mechanizmami izolującymi,
bo dotyczy ona w równym stopniu izolacji pre- i postzygotycznej.
Stopień izolacji pre- i postzygotycznej jest wprostproporcjonalna do wartości dystansu
genetycznego
Wartość wskaźnika izolacji prezygotycznej jest istotnie wyższa od wartości wskaźnika izolacji
postzygotycznej u par gatunków, które rozdzieliły się niedawno
Sterylność mieszańców wykształca się w tym samym czasie, co ich śmiertelność
Izolacja postzygotyczna pojawia się prawie zawsze wcześniej u samców (u płci
heterogametycznej) niż u samic (reguła Haldane’a)
Minimalny stopień izolacji pre- i postzygotycznej pozwalający gatunkom utrzymać odrębność
wynosi 0,85
Gatunki allopatryczne uzyskują I= 0,85 przy D= 0,66
Gatunki sympatryczne uzyskują I= 0,85 przy D= 0,31
W rodzaju Drosophila dystans genetyczny D= 1 uzyskiwany jest po około 5 mln lat.
D= 0,66 => 3,3 mln lat
D= 0,31 => 1,5 mln lat
Genetyczne podłoże izolacji rozrodczej
Zmiany liczby i struktury chromosomów bez zmian w strukturze genów
Zmiany w strukturze genów i ich częstościach (jak duże zmiany są konieczne, aby zapewnić
odrębność gatunkową)
o Kumaki (Bombina)- około 55 genów
o
Kroplik żółty (Mimillus guttatus)- trędownikowate, Scrophulariaceae
Powstanie bariery reprodukcyjnej pod wpływem naturalnej selekcji na tolerancję wysokiego
stężenia jonów Cu w glebie. Różnice dotyczą 2 genów odporności. Ich plejotropowym
efektem jest bardzo niska żywotność mieszańców pomiędzy rasą odporną a typem dzikim.
o Drosophila ex. gr. melanogaster
Izolacja rozrodcza ma charakter behawioralny i dotyczy różnicy w rytmie wibracji skrzydeł w
trakcie tańca godowego. Odpowiedzialny jest jeden gen per na chromosomie X.
o Wolbachia- pasożyty reprodukcyjne Arthopoda dziedziczone cytoplazmatycznie, obniżają
intensywność przepływu genów
a)
Indukcja partenogenezy- Encarsia formosa (Hymenoptera)
Samce można otrzymać jeśli samicy podawać antybiotyk (samce nie są jednak zdolne
do kojarzeń)
b)
Niezgodność cytoplazmatyczna dwukierunkowa- Nasonia spp. complex
(Hymenoptera)
Krzyżówki między osobnikami zainfekowanymi dwoma odmiennymi szczepami
Wolbachia nie dają potomstwa.

51
c)
Niezgodność cytoplazmatyczna jednokierunkowa- Drosophila recens i D.
subquinaria (związane z grzybami, północne USA- południowa Kanada)
Asymetryczny behawioralny mechanizm izolacyjny
♀R × ♂S – NIE!
♂R × ♀S – TAK! (brak istotnych różnic w porównaniu z krzyżówkami
wewnątrzgatunkowymi)
D. recens (W)
D. subquinaria (brak W)
D. recens ♂ (W) × D. subquinaria ♀ = F1 nieliczne
D. recens ♂ (W, antybiotyk) × D. subquinaria ♀ = F1 liczne
Postzygotyczny mechanizm izolacyjny
Mieszaniec F1 ♂ × R lub S = brak potomstwa (mieszańce F1 ♂ = sterylne, reguła Haldane’a)
Mieszaniec F1 ♀ × R lub S = liczne potomstwo (mieszańce F1 ♀ = płodne)
Równoległości między rozwojem ontogenetycznym i filogenetycznym
Prawo biogenetyczne Haeckla
Rozwój osobniczy (ontogeneza) jest skróconym i przyspieszonym powtórzeniem (rekapitulacją)
rozwoju rodowego (filogenezy).
Osobnik w swym rozwoju od jaja do postaci dojrzałej przechodzi przez podobne stadia, jakie
występowały w rozwoju rodowym jego przodków.
Rekapitulacja nie jest wierna!
Procesy rozwojowe, które oddają istotę filogenezy (uwarunkowane przebiegiem filogenezy)-
palingenetyczne
Procesy stanowiące odchylenia od rozwoju rodowego- cenogenetyczne
Heterochronizmy- niezgodności dotyczące czasu i kolejności zawiązywania się poszczególnych
narządów (akceleracje, retardacje, odwrócenie kolejności)
Heterotopizmy- niezgodności dotyczące miejsca (zmiany lokalizacji, sąsiedztwa zawiązków
narządów)
Specyficzne cechy przystosowawcze zarodkowe i larwalne (np. kiedy larwa jest planktoniczna, a
osobnik dojrzały bentoniczny)
„Nowości filogenetyczne” (cechy, których brak w ontogenezie przodka)- mogą pojawiać się na
różnych etapach rozwoju ontogenetycznego.
Siewiercow zaproponował trzy terminy dla różnych sytuacji:
Anabolia (nadkładka)- nowość filogenetyczna pojawiające się pod koniec rozwoju. Po pojawieniu
się kolejnych nowości może dochodzić do wstecznego wypierania cech pojawiających się w
filogenezie
Dewiacja- w środkowym etapie ontogenezy
Archalacja- w początkowych etapach ontogenezy
Jeśli w czasie ontogenezy stosunek rozmiarów dwóch części ciała organizmu nie zmienia się- wzrost
izometryczny
Wzrost allometryczny: y= b x
k
gdzie: y- wielkość I (np. części twarzowej czaszki)
x- wielkość II (np. puszki mózgowej)
b- stała wzrostu (wielkość narządu przy wielkości ciała= 1)
k- współczynnik alometrii (k= 1 => wzrost izometryczny)
Klasyfikacja związków między filogenezą i ontogenezą (według Goulda)
Trzy względnie niezależne procesy (zmienne) przebiegające w tempie charakterystycznym dla danego
taksonu:
1. wzrost (izometryczny)
2. rozwój (allometryczne zmiany kształtu ciała)
52
3. dojrzewanie płciowe
Model zegarowy uwzględniający powyższe trzy procesy wyskalowany na czas osiągnięcia dojrzałości
rozrodczej przodka przy umownych wartościach 1 dla wzrostu i rozwoju:
wzrost rozwój
przodek osiągał dojrzałość płciową przy: 1
1
potomek osiąga dojrzałość płciową przy: <1
<1
PROGENEZA
≥1
<1
NEOTENIA
>1
>1
HIPERMORFOZA
1
>1
AKCELERACJA
<1
1
KARŁOWATOŚĆ
>1
1
GIGANTYZM
1. hipomorfoza (= pedomorfoza = fetalizacja)
a) neotenia
b) progeneza (= pedogeneza)
2. hipermorfoza s. l. (= gerontomorfoza = anabolia)
a) hipermorfoza s. str.
b) akceleracja
3. karłowatość
4. gigantyzm
→ ONTOGENEZA →
←
F
ILO
G
EN
EZ
A
←
przodek
e1
e2
e3
m1
m2
m3
potomek e1
e2
e3
m1
NEOTENIA
(rozciągnięcie w czasie)
e1
e2
e3
m1
m2
PROGENEZA
e1
e2
e3
m1
m2 m3
m4
HIPERMORFOZA
(wydłużony rozwój)
e1
e2
e3
m1 m2
m3
m4
AKCELERACJA
(w
czasie jak u przodka, więcej
cech)
e1 e2 e3 m1 m2
m3
KARŁOWATOŚĆ
(szybsza ontogeneza)
e1
e2
e3
m1 m2
m3
GIGANTYZM
(dłuższa
ontogeneza)
e1, e2, e3- cechy embrionalne
m1, m2, m3, m4- cechy dojrzałe (postembrionalne)
podkreślone- moment uzyskania dojrzałości płciowej
Neotenia
Spowolnienie rozwoju somatycznego, dojrzałość płciowa osiągana w stadium rozwoju typowym dla
zarodka lub postaci młodocianej przodków.
Występuje przy takiej zmianie środowiska, że któreś z cech młodzieńczych stają się bardziej
przydatne od cech dojrzałego organizmu; prowadzi do opóźnienia rozwoju somatycznego zmiana
specjalizacji przez zmiany w rozwoju osobniczym)
Np.
Ambystoma mabeei- aksolotl (larwy w wodzie, dojrzałe na lądzie)
Proteus anguinus- odmieniec jaskiniowy
człowiek- płaska twarz, redukcja lub brak owłosienia, duża masa mózgu, rozjaśnienie
pigmentacji ciała, brak wałów nadoczodołowych i grzebieni czaszkowych, drobne uzębienie,
późne wyżynanie zębów itp.
Progeneza
Skrócenie ontogenezy i przyspieszenie dojrzewania płciowego (młodociana lub zarodkowa cecha
przodka zostaje zachowana przez dojrzałego potomka)

53
Występuje w warunkach częstych i nieprzewidywalnych wobec zasobnego środowiska i prowadzi do:
a) skrócenia cyklu życiowego
b) wczesnego osiągania dojrzałości płciowej
c) zmniejszenia rozmiarów ciała
d) dużej produkcji potomstwa (ucieczka od specjalizacji poprzez zmiany ewolucyjne w rozwoju
osobniczym)
Np.
Siteroptes graminum- roztocz bytujący na grzybach, Mycophila (Diptera)
Przy obfitości pokarmu- kolejne pokolenia partenogenetycznie w ciele matki, mając w swych
ciałach kolejne pokolenie partenogenetyczne (do 250 takich pokoleń)
W warunkach głodu- wśród larw pojawiają się samce i samice, poszukują nowych źródeł
pokarmu
Hipermorfoza s. l.
Opóźnienie dojrzałości płciowej i najczęściej wydłużenie ontogenezy (hipermorfoza s. str.) przez
wzbogacenie jej o nowe stadia, niespotykane u przodków.
Występuje w warunkach stabilnych i przewidywalnych, w wysycanym organizmami i gatunkami
środowisku i prowadzi (w typowej postaci) do wydłużenia cyklu życiowego, opóźnienia w osiąganiu
dojrzałości płciowej, wzrostu rozwoju ciała i obniżenia rozrodu (pogłębienie specjalizacji poprzez
zmiany ewolucyjne w rozwoju osobniczym)
Np.
Megaloceros- jeleniowate (poroże o rozpiętości około 3,5 m)
y = 0,034 x
1,85
Zmiany proporcji czaszki koniowatych
Eocen- formy liściożerne
Dziś- formy trawożerne
Gigantyzm i hipermorfoza (długość trzewioczaszki i głębokość żuchwy), następnie akceleracja
(pogłębienie żuchwy)
Akceleracja
Exocoetidae
Hemiramphidae akceleracja dotyczy wydłużenia szczęki dolnej, górnej lub obu
Belonidae
Genetyczna kontrola morfogenezy
Znaczenie ewolucyjne polimorfizmu liczby powtórzeń tandemowych (variable number tandem repeats
VNTR)
142 psy z 92 ras, 37 genów i sekwencji regionów powtórzeniowych
Istotne różnice w liczbie powtórzeń w różnych genach regulujących rozwój u różnych
ras
Istotne różnice w liczbie powtórzeń w czasie rozwoju danej rasy
Korelacja między kształtem czaszki i liczbą powtórzeń
Więcej powtórzeń tandemowych kodonów glutaminowych w genie Runx- 2 => kufa dłuższa i bardziej
prosta (np. bulterier)
Gen Alx- 4 (aristaless- 4)- u wielkich psów pirenejskich (6 palców w tylnej łapie) gen krótszy o 51
zasad w porównaniu do innych psów (5 palców)
Wykład 12.
Ewolucja cech altruistycznych i rytualizacja zachowań konfliktowych u zwierząt (wybrane teorie
socjobiologiczne)
Socjobiologia

54
E. Wilson (1975): „Systematyczne studium biologicznych podstaw wszelkich zachowań społecznych,
czyli interakcji wewnątrzgatunkowych”.
Czynniki biologiczne odgrywają decydującą rolę w determinowaniu zachowań społecznych u zwierząt
(również u człowieka).
Warunki ewolucji zachowań na drodze doboru:
1. W populacji muszą istnieć różne zachowania
2. Różnice muszą być przynajmniej częściowo determinowane genetycznie
3. Niektóre spośród alternatywnych zachowań muszą prowadzić do większego sukcesu
reprodukcyjnego
Genetyczne podłoże zachowań (na przykładzie Drosophila melanogaster)
1. zorientowanie
2. uderzanie w odwłok
3. śpiew godowy
4. lizanie genitaliów
5. samiec wspina się na samicę
6. kopulacja
Analiza sekwencji zachowania godowego u D. melanogaster oraz położenie regionów CUN, które
kontrolują poszczególne etapy zalotów samców wskazuje na skomplikowane podłoże genetyczne.
Nie dojdzie do kopulacji dopóki samiec nie odprawi całego rytuału i dopóki samica nie przyjmie
zalotów samca.
Zidentyfikowano już kilkanaście genów odpowiedzialnych za zachowanie godowe. Większość genów
ma efekty plejotropowe i odmienną ekspresję u samców i samic.
Wydawanie pieśni godowej przez samców kontrolowane jest przez trzy geny.
Gen period steruje rytmiką trzepotu skrzydeł.
Samce z dzikim genem wykonują normalną pieśń (zwiększanie przerw między pulsami przez 27s ,
następnie zmniejszanie przerw), która uwrażliwia samice.
Mutacje powodują nienormalne rytmy lub utratę rytmu- obniżenie chęci samic do przyjęcia samca.
Gen period wpływa również na rytm okołodobowy (ekspresja w wielu częściach CUN).
Gen fruitless- odpowiedzialny za preferencje seksualne (samce mutanty zalecają się zarówno do
samic, jak i do samców) i niezbędny na dalszych etapach zalotów (mutanty nigdy nie kopulują z
samicami).
Gen gen kinazy białkowej II zależnej od kalmoduliny i wapnia CaMKII bierze udział w
indukowaniu procesów biochemicznych, które niezbędne są w uczeniu się.
Dzikie samce tracą zainteresowanie samicą, która miała kontakt z innym samcem. Mutanty
niezmordowanie podążają za pokrytymi już samicami.
1. samica, która już kopulowała wydziela feromon „antyafrodyzjak”
2. u samca wzrost stężenia Ca
2+
w rejonie mózgu odpowiedzialnym za pobudzenie seksualne
3. aktywacja CaMKII
4. fosforylacja białka EAG (produkt genu eag) w kanałach potasowych
5. otwarcie kanałów umożliwiające wypływ jonów K
+
z neuronów
6. zniesienie pobudliwości i obniżenie zdolności uwalniania neurotransmiterów
7. samiec nie podejmuje zalotów
Klasyczna teoria selekcji naturalnej sformalizowana przez genetykę populacyjną nie wyjaśnia
ewolucji zachowań altruistycznych u zwierząt.
Altruizm biologiczny
Takie zachowanie osobnika, które zmniejsza jego dostosowanie, a zwiększa dostosowanie innego
osobnika.
Najbardziej znanym przykładem altruizmu jest altruizm między krewnymi.
DOBÓR KREWNIACZY

55
Altruizm ma sens jeśli zyski w dostosowaniu odbiorcy aktu altruistycznego (b) przewyższają straty w
dostosowaniu altruistycznego dawcy (c).
b> c
Częstość alleli determinujących zachowania altruistyczne będzie wzrastać, jeśli jest spełniona tzw.
reguła Hamiltona (1964)
c/ b < r (lub rb> c)
gdzie: r- współczynnik pokrewieństwa dawcy i biorcy
Współczynnik pokrewieństwa r:
Prawdopodobieństwo, że gen u jednego osobnika jest dzięki wspólnemu dziedzictwu, identyczną
kopią genu innego osobnika.
r
w linii prostej w linii bocznej
0,5
potomstwo
rodzeństwo
0,25
wnuki
rodzeństwo przyrodnie, siostrzeńcy, siostrzenice
0,125 prawnuki
kuzyni
Hamilton wprowadził koncepcję dostosowania łącznego (inclusive fitness) która mówi, że
dostosowanie danego organizmu jest sumą sukcesu reprodukcyjnego i wpływu tego organizmu na
sukces reprodukcyjny jego krewniaków
rb> c
Interpretacja:
Jeśli krewny jest nosicielem genu odpowiedzialnego za zachowania altruistyczne z
prawdopodobieństwem np. r= 0,5, to tracąc jednostkę dostosowania i zwiększając dostosowanie
krewnego o więcej niż dwie jednostki można przyczynić się do rozpowszechnienia tego genu bardziej,
niż wówczas, gdy zachowanie altruistyczne zostanie powstrzymane.
Przykłady altruizmu między krewnymi:
kooperacja i okrzyki alarmowe u susłów Beldinga Spermophilus beldingi (Sciuridae)
spokrewnione samice nie walczą o nory, nie przepędzają się ze swych terenów, współpracują
ze sobą w obronie młodych przed dzieciobójczymi skłonnościami innych osobników
okrzyki alarmowe wydawane są wtedy, gdy w pobliżu są krewniacy (oprócz potomstwa,
również krewni w linii bocznej)
okrzyki alarmowe u nieświszczuków (piesków preriowych) Cynomys ludovicianus
okrzyki alarmowe wydawane są niemal równie często w obecności potomstwa, jak i innych
krewnych
samce wydają okrzyki rzadziej niż samice (opuszczają samice po porodzie)
wspólne samice u kokoszki tasmańskiej Tribonyx (= Gallinula) mortierii (Rallidae)
bracia mają wspólną samicę, oba z nią kopulują, a następnie pomagają przy wychowie
młodych (jeden z braci jest dominujący, drugi podporządkowany)
szacunki sukcesu reprodukcyjnego w różnych warunkach wskazują, że zarówno samiec
podporządkowany, jak i dominujący korzysta dzięki wzrostowi dostosowania łącznego
Dobór krewniaczy został oryginalnie zaproponowany przez Karola Darwina jako wytłumaczenie
istnienia sterylnych kast u owadów, lecz ogólną akceptację zyskał dopiero dzięki przełomowym
pracom W. D. Hamiltona.
Altruizm ekstremalny
Sterylne kasty u owadów eusocjalnych
Predyspozycje genetyczne- haploidalny (haplodiploidalny?) system determinacji płci!
Stosunki pokrewieństwa u owadów z haplodiploidalnym systemem determinacji płci
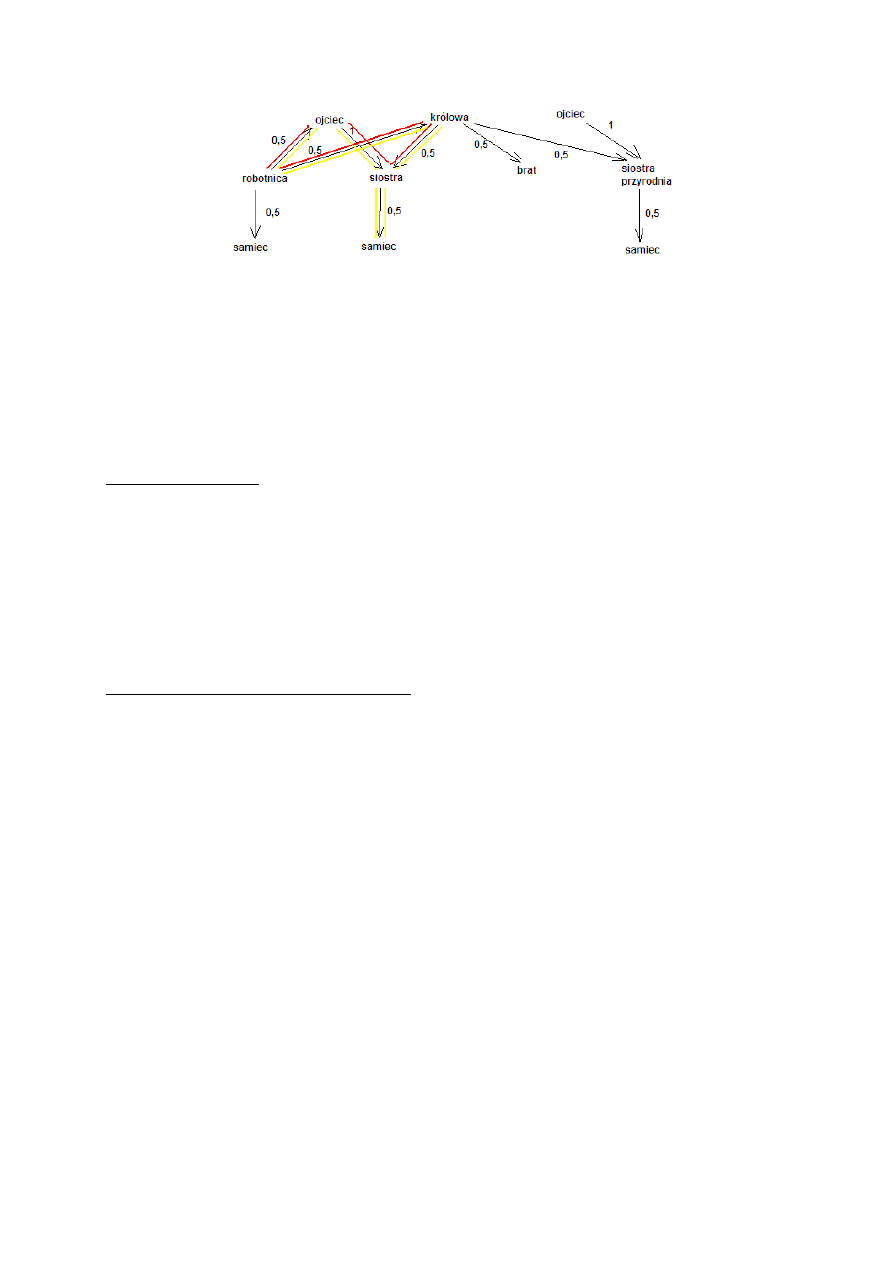
56
Przykłady obliczeń wartości r:
r (robotnica- siostra) = (0,5 × 1) + ( 0,5 × 0,5) = 0,75
(czerwone)
r (robotnica- syn siostry) = (0,5 × 1 × 0,5) + (0,5 × 0,5 × 0,5) = 0,375
(żółte)
Wniosek- sterylna robotnica może pod względem genetycznym więcej zyskać wychowując
rozmnażającą się siostrę, niż gdyby sama stała się płodna i miała własne córki.
Haplodiploidalność może stanowić ważną predyspozycję do wykształcenia eusocjalności u
Hymenoptera, nie jest to jednak ani warunek konieczny, ani wystarczający.
Eusocjalność u ssaków
golce Heterocephalus glaber (Batyergidae)
eusocjalność- opieka kooperatywna
bezpłodne kasty
zachodzące na siebie pokolenia
zespołowe kopanie tuneli
podział pracy związany z rozmiarami ciała (polietyzm)
jedna samica reproduktorska
85% kopulacji między bliskimi krewnymi
Średnie pokrewieństwo między członkami kolonii- 81%
Wzajemność (altruizm wobec krewniaków)
Dzielenie się krwią u nietoperzy wampirów Desmodus rotundus (Desmodontidae)
Gatunek pospolity na przeobrażonych w pastwiska obszarach tropikalnych Ameryki
Odżywiają się nocą krwią ssaków (głównie koni)
Lokalizacja ofiary- kombinacja węchu, słuchu i echolokalizacji.
Organizacja społeczna:
o kolonia- 8- 12 dojrzałych samic z młodymi przebywających za dnia w pustych pniach lub
jaskiniach
o kolonie odizolowane od siebie, ale skład kolonii zmienny- migracje osobników między koloniami
o
dorastające samce opuszczają matkę i gniazdują samotnie lub w małych grupach broniąc teren z
kolonią samic
o 50% potomstwa w kolonii pochodzi z tego samego ojca
Nietoperz co 60 godzin musi wypić 20- 30 ml krwi (50- 100% masy ciała)
30% młodych i 7% dorosłych nie zdobywa pokarmu danej nocy (w danej grupie wiekowej
niepowodzenie jest losowe)
Po 60 godzin postu traci 25% masy ciała i ginie nie mogąc utrzymać krytycznej temperatury ciała.
Osobniki, którym nie udało się zdobyć pokarmu danej nocy, żebrzą o krew u tych, którym się
powiodło.
Zwracanie krwi następuje między bliskimi krewnymi lub między niespokrewnionymi, ale często
przebywającymi blisko siebie w tej samej kolonii
o 70%- matka- dziecko
o 30%- samica- obce dziecko
Samica- samica
Samiec- dziecko
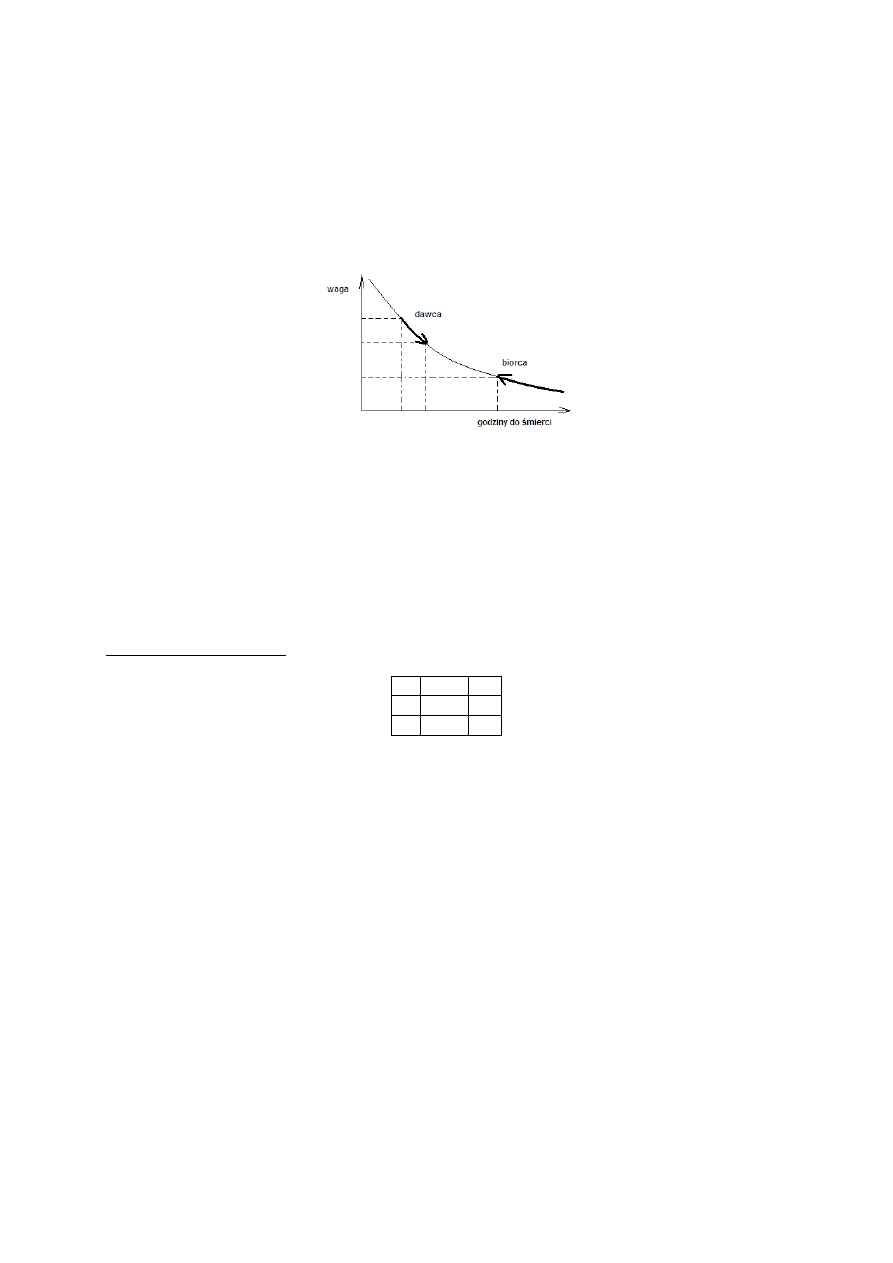
57
Dzielenie się krwią nie odbywa się losowo. Najedzone przekazują krew osobnikom znajdującym się
na krawędzi głodu (< 24h życia) i z którymi miały wcześniej kontakt w ramach tej samej kolonii
(populacji).
Czasem dwa niespokrewnione osobniki zawierają „koleżeński układ”
Też: hieny i ludzie
Strata masy po posiłku przebiega wzdłuż ujemnej krzywej wykładniczej
Posiłek z przekazanej krwi wiąże się z niewielkim kosztem dla dawcy i wielką korzyścią dla biorcy
(możliwość przeżycia kolejnej nocy- szansa na żerowanie)
o
Śmiertelność roczna rzeczywista- 24%
o
Śmiertelność roczna potencjalna (bez odwzajemnienia)- 82%
Głodne nietoperze iskają potencjalnych dawców (rozpoznają po objętości żołądka) częściej niż
wynikałoby to z przypadku (wstęp do przekazania pokarmu)
Zachowania pielęgnacyjne (5% czasu każdego dnia)- częściej między osobnikami spokrewnionymi
lub współmieszkającymi.
Matka i młode rozpoznają się wzajemnie dzięki zróżnicowanym odgłosom.
Altruizm odwzajemniony
Macierz wzrostu (b) i spadku (c) dostosowania dla strategii altruisty (A) i niealtruisty (B):
A
B
A b – c - c
B
b
0
Warunki odwzajemniania u wampirów D. rodundus:
1. samice przebywają razem przez dłuższy czas, tak że mają duże szanse na ponowne spotkanie
2. prawdopodobieństwo podzielenia się krwią może być przewidziane na podstawie poprzednich
spotkań
3. role dawcy i biorcy często się zmieniają
4. krótkotrwały zysk odbiorcy przewyższa stratę dawcy
5. altruiści mają zdolność rozpoznawania i wyeliminowania oszusta z układu
Warunki ewolucji altruizmu zwrotnego:
1. b ≥ c (wzrost dostosowania w wyniku działania altruistycznego nie jest mniejszy od spadku
dostosowania, zwykle b >> c)
2. S → 1 (wysokie prawdopodobieństwo ponownego spotkania partnerów)
3. koniczność indywidualnego rozpoznania
4. pewna krytyczna frekwencja altruistów odwzajemniających się (powstaje w wyniku działania
doboru krewniaczego lub dryfu genetycznego w małych populacjach)
5. spełniona nierówność ( b – c/1 – S) > b
(b – c) + (b – c)S + (b – c)S
2
+ … = (b – c/1 – S)
Inne przykłady altruizmu zwrotnego:
1. przymierza u samców pawianów Papio Anubis (jeden samiec odgania inne, a drugi w tym
czasie kopuluje z samicą)
58
2. „handel jajami” u ryby Hypoplectrus nigricans
Strategia ewolucyjnie stabilna (ESS)- 1973
Wprowadzona do biologii po raz pierwszy w wyniku zastosowania teorii gier do wyjaśnienia
ograniczonej agresji między zwierzętami. Dziś ESS pozwala analizować również inne, bardziej
złożone zachowania organizmów.
Dylemat Więźnia
więzień A
oskarżenie milczenie
w
ię
zi
eń
B
oskarżenie
A. 5 lat
A. 0 lat
B. 5 lat
B. 20 lat
milczenie
A. 20 lat
A. 1 rok
B. 0 lat
B. 1 rok
Dylemat Więźnia- macierz wygranych
WW- nagroda
WZ- wypłata oszukanego
ZW- pokusa
ZZ- kara
Strategia „wet za wet” (Tit for tat)
Współpraca w pierwszym ruchu, potem- powtarzanie ostatniego posunięcia przeciwnika (altruizm
odwzajemniony)
Strategia Pawłowa
Powtarzanie poprzedniego ruchu jeśli był on zyskowny (nagroda za współpracę lub zdradę
współpracownika), natomiast zmiana posunięcia jeśli przyniosło straty (obopólna zdrada lub
współpraca ze zdradzającym).
Macierz „wygranych” osobników stosujących strategię agresora (H), ustępującego (D) i legalisty (L):
V- wzrost dostosowania
C- zmniejszenie dostosowania
Zmiana dostosowania osobnika stosującego strategię wymienioną w pierwszej kolumnie przy
spotkaniu z osobnikiem stosującym strategię wymienioną w główce tabeli.
Wykład 13.
1. V > C strategia H jest ESS
2. V < C
Przykłady poważnych walk:
Woły piżmowe (Ovibos moschatus)- rocznie 5- 10% dorosłych samców ginie w walce o
samice
Jelenie mulaki (Odocoileus hemionus)- ślady obrażeń nosi 10% samców starszych niż 1,5
roku
Narwale (Monodon monoceros)- nawet 60% samców ze złamanymi ciosami i innymi
obrażeniami głowy
V < C czyste strategie H i D nie są ESS
V < C strategią ESS może być strategia mieszana!
Gracz 2
współpraca (W)
zdrada (Z)
G
ra
cz
1
współpraca (W) 1.- 3 punkty
2.- 3 punkty
1.- 0 punktów
2.- 5 punktów
zdrada (Z)
1.- 5 punktów
2.- 0 punktów
1.- 1 punkt
2.- 1 punkt
H
D
L
H (V – C)/2
V
(V – C)/4 + V/2
D
0
V/2
V/4
L (V – C)/4 V/2 + V/4
V/2

59
H
D
H (V – C)/2
V
D
0
V/2
Średnie dostosowanie H:
W
H
= [(V – C)/2]P + V(1 – P)
gdzie: W- dostosowanie
P- proporcja osobników stosujących strategię agresora
Średnie dostosowanie D:
W
D
= 0 + (V/2)(1 – P)
Proporcja agresorów w stanie równowagi:
W
H
= W
D
P= V/C
Przykład:
Grzebaczowate Sphex ichneumoneus (Hymenoptera, Sphecidae, grzebaczowate)
o
Kopanie jamek („kopacz”) i przejmowanie jamek gotowych („włamywacz”) stabilizuje się na
określonym poziomie
o 68 samic, 1500 godzin obserwacji, 410 nor
59% decyzji- kopanie sukces= 9,6 jaj/ 42 doby
41% decyzji- przejmowanie sukces= 8,4 jaj/ 42 doby
o
Kopanie i włamywanie jest mieszaną ESS
Macierz „wygranych” osobników stosujących strategię legalisty (L) w porównaniu ze strategiami
agresora (H) i ustępującego (D):
legalista- wdaje się w konflikt tylko wtedy, gdy jest posiadaczem danego zasobu
Przykład:
Motyl osadnik egeria Pararge egeria (Satyridae)
o Strategia legalisty- samce respektują prawo własności: intruzi nie wdają się w walkę z
rezydentami
o
Motyle żyją na dnie lasu, bronią plam świetlnych
Strategia legalisty może być ESS.
Opisane trzy przykłady są niewątpliwie zbyt proste, żeby szczegółowo odzwierciedlić rzeczywiste
strategie przyjmowane przez zwierzęta. Stanowią jednak łatwo wyobrażalne skrajności, które możemy
uznać za uproszczone wersje strategii obserwowanych w przyrodzie.
EWOLUCJA ROZMNAŻANIA PŁCIOWEGO
Fenomen płciowości to „królowa” problemów biologii ewolucyjnej
Zadawanych jest wiele pytań, np.:
1. Jakie były przyczyny i drogi powstania płciowości?
2. Dlaczego płciowość jest tak rozpowszechniona, skoro wcale nie jest ona niezbędna do
trwałego rozmnażania?
Podstawowe terminy i istota problemu
1. rozmnażanie płciowe= biseksualne, seksualne
H
D
L
H (V – C)/2
V
(V – C)/4 + V/2
D
0
V/2
V/4
L (V – C)/4 V/2 + V/4
V/2
60
2. rozmnażanie bezpłciowe (= aseksualne)- brak mejozy
Tu: partenogeneza apomiktyczna ( telitokia- powstawanie samic z niezapłodnionych jaj na drodze
mitozy)
Na najbardziej podstawowym poziomie płeć (proces płciowy) i rozmnażanie to przeciwieństwo
o
Rozmnażanie= podział komórki na dwie potomne
o
Proces płciowy= fuzja dwóch komórek w jedną
Płciowość nie jest niezbędna do trwałego rozmnażania (wiele organizmów rozmnaża się przez szereg
pokoleń bez udziału procesów płciowych)
Rozmnażanie płciowe jest wysoce kosztowne!
Koszty rozmnażania płciowego:
1. rekombinacja „miesza genotypy” rozbijając adaptacyjnie korzystne kombinacje genów
2. rozmnażanie płciowe z naprzemienną mejozą i syngamią (fuzją gamet) wymaga więcej czasu
niż apomiktyczna partenogeneza i spowalnia proces reprodukcji
3. u organizmów „wyższych” kojarzenie może być ryzykowne i kosztowne: trudności i strata
czasu podczas poszukiwania partnera, narażenie na niebezpieczeństwo ze strony drapieżników
podczas kopulacji, ryzyko zarażenia chorobą przenoszoną drogą płciową, koszt nadprodukcji
gamet, koszt utrzymania dymorfizmu płciowego, podejmowanie konkurencji płciowej itp.
4. przy niskim zagęszczeniu populacji rozmnażanie się płciowo może być trudne do
koordynowania. Partenogeneza zapewnia reprodukcję potencjalnie w każdym miejscu i czasie.
5. każdy organizm bezpłciowy wydaje tylko swoje potomstwo, a samice rozmnażające się
płciowo muszą ustępować połowę miejsca w genotypie swych dzieci samcom. Twierdzi się,
że ponoszą one tzw. podwójne koszty samców.
Miara reprodukcji netto jako tzw. wskaźnik zastępowania, czyli liczba córek przypadających w
następnym pokoleniu na jedną matkę.
R = B S p
B- liczba potomków
S- prawdopodobieństwo ich przeżycia
p- frekwencja samic w potomstwie
61
Jeśli B i S pozostają niezmienne, to im większy udział samic w potomstwie, tym większe R.
Wniosek- osobniki rozmnażające się bezpłciowo rozmnażają się dwukrotnie szybciej, co oznacza, że
mają dwukrotnie wyższe dostosowanie.
Przykład hipotetyczny:
Każda samica wydaje w ciągu życia cztery osobniki potomne (proporcja płci w potomstwie samic
rozmnażających się płciowo wynosi 1:1)
pokolenie
rozmnażanie
płciowe
rozmnażanie
bezpłciowe
proporcja osobników aseksualnych w
populacji
1
♀ × ♂
♀
1/3
2
♀ × ♂, ♀ × ♂
♀ ♀ ♀ ♀
1/2
3
♀ × ♂, ♀ × ♂
♀ × ♂, ♀ × ♂
♀ ♀ ♀ ♀
♀ ♀ ♀ ♀
♀ ♀ ♀ ♀
♀ ♀ ♀ ♀
2/3
Wniosek- dostosowanie osobników rozmnażających się płciowo jest o połowę niższe niż tych
rozmnażających się partenogenetycznie (tzw. koszt samców)
Podstawowe rodzaje partenogenezy:
1. partenogeneza apomiktyczna
2. partenogeneza automiktyczna
3. gynogeneza
plemnik pobudza do rozwoju!
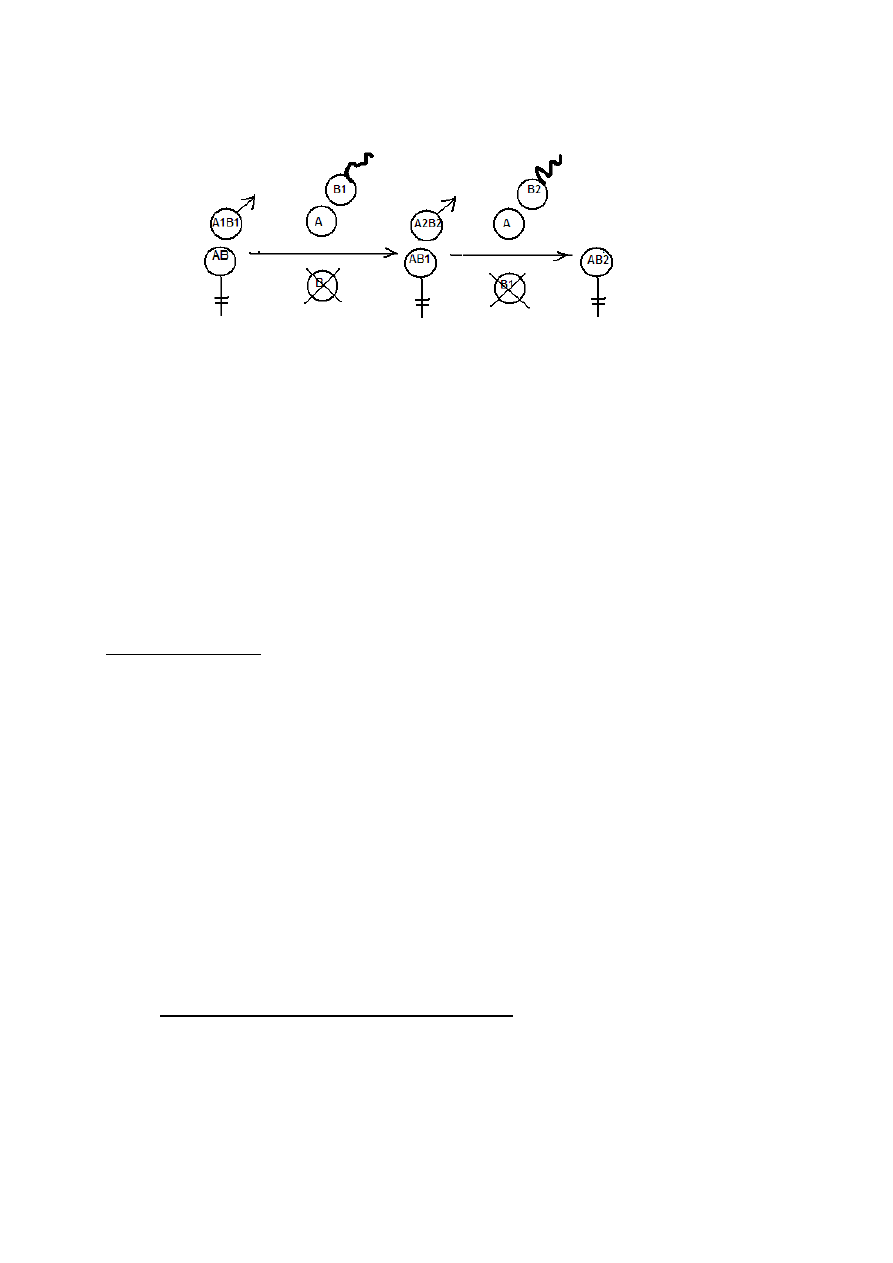
62
4. hybrydogeneza
Mimo zdecydowanej przewagi rozmnażania płciowego, w wielu grupach Eucaryota spotykamy formy
rozmnażające się obligatoryjnie bezpłciowo.
o Protista- np. Cryptophyceae, Haptophyta
o
Rośliny- < 1% nasiennych, 10% paprotników
o Grzyby- około 20%, szczególnie Endogonales w sprzężniakach, Zygomycota
o Zwierzęta- < 1% (wśród kręgowców 70 gatunków)
Grupy gatunków bez rozmnażania płciowego:
Filogenetycznie młode (np. jaszczurki Cnemidophorus, gyno- i hybrydogeniczne ryby
Peociliopsis)
Współwystępują w danym taksonie z blisko spokrewnionymi gatunkami płciowymi (np.
szereg roztoczy Oribatei)
Filogenetycznie stare, tzw. odwieczni aseksualiści tzn. grupy u których brak rozmnażania
płciowego od ponad 3 mln lat (lub występuje bardzo sporadycznie)
Odwieczni aseksualiści
Wrotki Bdelloidea
380 gatunków współcześnie, kosmopolityczne i niemal ubikwistyczne
Zamieszkują wody słodkie oraz wilgotne mchy i glebę
Gatunki lądowe w niekorzystnych warunkach przechodzą w stan anhydrobiozy.
85 mln lat bez seksu (1,5 mld pokoleń)
Małże Lasaea
Kosmopolityczne, wielkość około 2 mm
Od 3 do 6 mln lat bez seksu
Skorupiaki Darwinulidae (Ostracoda)
26 gatunków kosmopolitycznych, wielkość mniej niż 1 mm
200 mln (?) lat bez seksu
Darwinula stevensoni
Vestalenula cylindrica
Vestalenula cornelia
Alicenula leguminella
Eucypnis virens (Ostracoda)
Pospolity, często masowo w wodach okresowych, wielkość około 2,5 mm, organizm modelowy
1. unikanie tzw. Zapadki Müllera (Müller’s ratched)
U gatunków bezpłciowych z pokolenia na pokolenie przybywa mutacji nieznacznie obniżających
dostosowanie (nie eliminowane przez dobór w ciągu jednego pokolenia), których efekty się kumulują
Po pewnym czasie z populacji znikają osobniki wolne od mutacji (liczba mutacji nie może się
zmniejszyć), a średnie dostosowanie populacji spada (zapadka na kole zębatym)
Rozmnażanie płciowe poprzez rekombinację może zmniejszyć liczbę mutacji u potomstwa i zachować
osobniki o maksymalnym dostosowaniu- zapobiega zapadce.
Populacja bezpłciowa będzie obciążona większym ładunkiem szkodliwych mutacji.
63
Bez rozmnażania płciowego szansa na powrót do klasy optymalnej z liczbą mutacji równą 0 jest
znikoma.
Zapadka Müllera zmniejsza dostosowanie aseksualnie rozmnażających się szczepów Salmonella
typhimurium. Pięć linii (1%) miało znacząco mniejsze dostosowanie (obniżone tempo podziałów
komórkowych) niż linia wyjściowa.
Średni czas podziałów- 23,2 + 0,7 minut WT
Żadna z linii nie miała większego dostosowania niż linia ancestralna.
Wniosek- bez procesów płciowych i przy silnie działającym dryfie genetycznym mutacje powrotne nie
mogą kompensować skutków akumulacji szkodliwych mutacji.
Uwagi:
Zapadka Müllera działa jedynie jeśli:
genom jest duży (wydaje się, że drobne pierwotniaki są na granicy)
populacja jest mała
Mutacje mało szkodliwe obniżają wyraźnie dostosowanie jeśli ich liczba przekroczy pewien próg.
Tempo kumulowania takich mutacji zależy od:
wielkości genomu (+)
częstości zachodzenia mutacji (+)
wielkości populacji (-)
Wirusy RNA podlegają zapadce, bo tempo mutacji RNA jest większe niż DNA.
2. gilotynka Kondrashova
Jeśli:
a) tempo mutacji małoszkodliwych jest duże (>1/ genom/ pokolenie)
b) mutacje wykazują działanie synergiczne
to rozmnażanie płciowe i rekombinacja zmniejszają obciążenie tymi mutacjami nawet w dużych
populacjach (działanie ogólnie podobne do zapadki Müllera)
U jest pozytywnie skorelowane z czasem trwania jednego pokolenia
Gilotynka Kondrashova może działać w niektórych przypadkach, ale z pewnością nie jest
wytłumaczeniem uniwersalnym.
Potencjalna długoterminowa korzyść dla populacji płciowej, która nie kompensuje krótkoterminowej
korzyści jaką mają osobniki rozmnażające się bezpłciowo.
Inne przykłady (np. hipoteza Czerwonej Królowej)- podręcznik.
Wyszukiwarka
Podobne podstrony:
Bejnarowiczsprawozdanie2, do szkoły, mechanizmy ewolucji roślin
Mechanizmy ewolucji test
pytania ewolucjonizm, Studia UMCS, V semestr, Mechanizmy ewolucji
9 Mechanizmu ewolucji, ich konsekwencje i metody badania (2009)
ewolucjonizm wykłady + pytania, zagadnienia 1-4, Mechanizmy ewolucji
Mechanizmy ewolucji lic (spr przez kramarz), ochrona środowiska UJ, II semestr SUM, ewolucjonizm
Mechanizmy ewolucji Egzamin i Poprawa 15r
BŁĘDY JAKO MECHANIZM EWOLUCJI
Mechanizmy ewolucji Wykłady
Mechanizmy ewolucji egzamin test
ewolucjonizm wykłady + pytania, tematyewol, Mechanizmy ewolucji
Mechanizmy ewolucji skrypcik z wykładów prof Kozłowskiego
Mechanizmy ewolucji zadania tekstowe
Mechanizmy ewolucji egzamin test
Krzanowska, Łomnicki, Rafiński, Szarski, Szymura Zarys mechanizmów ewolucji str 142 147, 215 264
22 Gecow, Algorytmy ewolucyjne i genetyczne, ewolucja sieci zlozonych i modele regulacji genowej a m
Ewolucja Szokująco prosty mechanizm Kozłowski Jan
Antybiotykooporność sprzed 30 tys lat Lekoopornosc to mechanizm w ramach nowmy reakcji na srodowisko
więcej podobnych podstron