
Prawo Hardy’ego - Weinberga
dr in
ż
. Dariusz Kaczmarczyk

Prawo Hardy’ego - Weinberga
Godfrey Harold Hardy (matematyk, Anglia)
Wilhelm Weinberg (fizjolog , Niemcy)
Obydwaj panowie pracuj
ą
c niezale
ż
nie od siebie
sformułowali w roku 1908 tez
ę
głosz
ą
ca
ż
e:
aa
Aa
AA
2pq
H
q
2
p
2
f
a
= q
f
A
= p
Godfrey Harold Hardy
Wilhelm Weinberg
Je
ż
eli w danym locus genowym wyst
ę
puj
ą
dwa allele
(A i a) i je
ż
eli frekwencj
ę
wyst
ę
powania allelu
dominuj
ą
cego (A) oznaczymy jako p, a recesywnego
(a) jako q, to cz
ę
sto
ść
wyst
ę
powania
poszczególnych genotypów w populacji b
ę
dzie
nast
ę
puj
ą
ca:
Zale
ż
no
ść
ta nazywana jest Prawem Hardy’ego - Weinberga

p
2
+2pq+q
2
gdzie:
p- frekwencja cz
ęś
ciej wyst
ę
puj
ą
cego allelu
q- frekwencja rzadziej wyst
ę
puj
ą
cego allelu
Model Hardy’ego-Weinberga
Zało
ż
enia modelu Hardy’ego-Weinberga:
1. liczebno
ść
populacji jest wielka i stała w kolejnych pokoleniach,
2. kojarzenie osobników jest losowe,
3. organizmy s
ą
diploidalne,
4. pokolenia nie zaz
ę
biaj
ą
si
ę
,
5. rozród odbywa si
ę
drog
ą
płciow
ą
,
6. wpływ mutacji i selekcji jest niewielki i mo
ż
na go zaniedba
ć
.

Przebadano populacj
ę
pewnego gatunku
motyla i wykryto ze składa si
ę
ona z
osobników o nast
ę
puj
ą
cych fenotypach i
genotypach
BB - 226 osobników
Bb - 43 osobniki
bb - 1 osobnik
N – 270 osobników
BB
Bb
Bb
bb
N
N
N
p
Bb
BB
2
)
*
2
(
+
=
N
N
N
q
Bb
bb
2
)
*
2
(
+
=
08
,
0
270
*
2
43
)
1
*
2
(
92
,
0
270
*
2
43
)
226
*
2
(
≈
+
=
≈
+
=
q
p
Obliczanie frekwencji alleli
gdzie:
B – p
b - q
p+p=1
N
BB
– liczba osobników
o genotypie BB
N
Bb
– liczba osobników
o genotypie BB
N
bb
– liczba osobników
o genotypie BB
cz
ę
sto
ść
allelu a
c
z
ę
s
to
ś
ć
g
e
n
o
ty
p
u
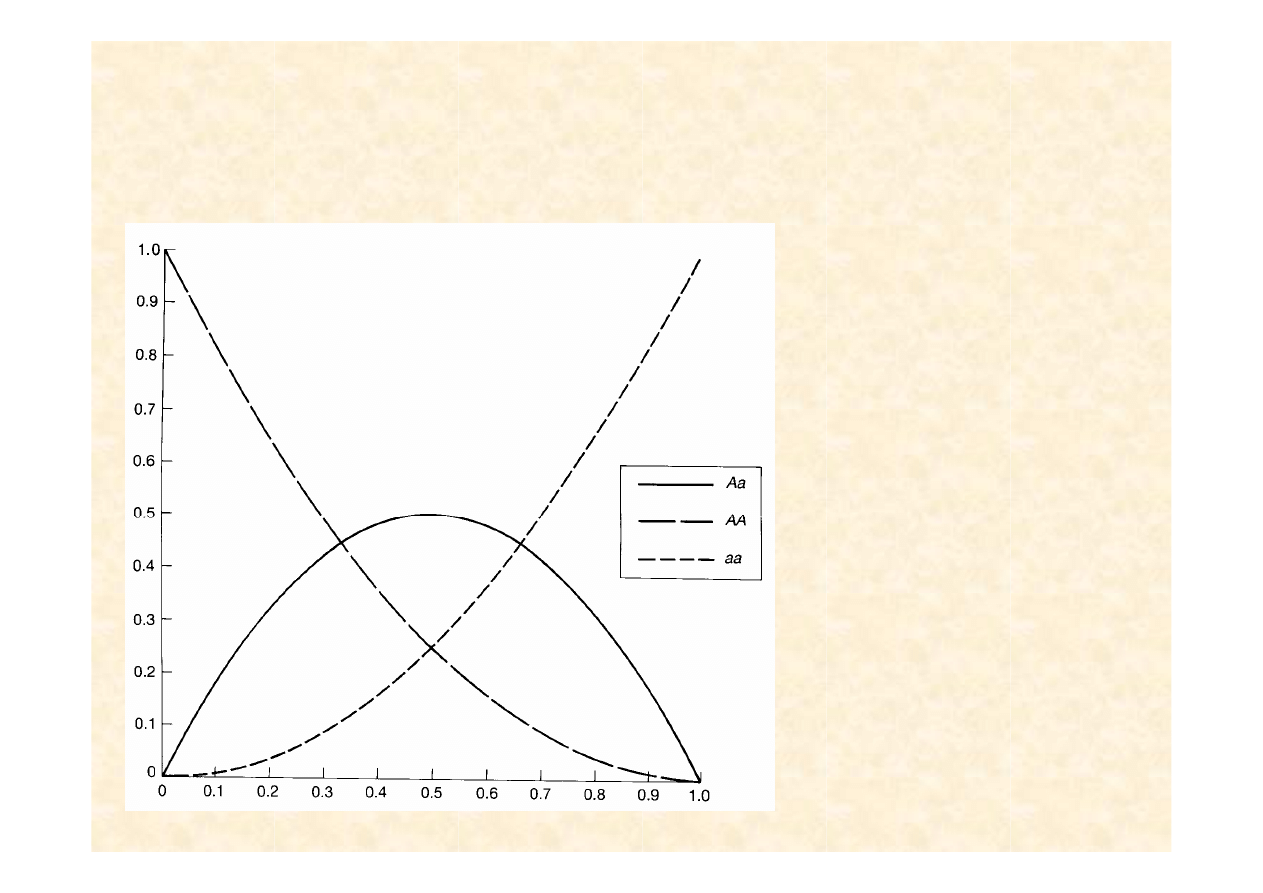
Frekwencja genotypów wynikaj
ą
ca z prawa Hardy’ego i Weinberga
Wnioski:
Gdy allel rzadki, prawie
wyłącznie związany jest w
heterozygotach (które
zazwyczaj nie różnią się
fenotypowo od homozygot
dominujących)
Eliminacja rzadkich alleli
recesywnych jest nieskuteczna
Plan Hitlera eliminacji chorób
genetycznych (zazwyczaj
warunkowanych przez
recesywne układy
homozygotyczne) był zatem nie
tylko zbrodniczy lecz i
nonsensowny
Nawet po eliminacji homozygot
recesywne allele są
dziedziczone za pośrednictwem
układów heterozygotycznych;
nie oddawajmy więc władzy
szaleńcom

Dlaczego tak trudno jest zmieni
ć
frekwencj
ę
alleli
recesywnych, gdy prowadzimy selekcj
ę
na podstawie cech
fenotypowych i nie mo
ż
emy odró
ż
ni
ć
homozygot
dominuj
ą
cych od heterozygot?

)
(
1
0
0
Nq
q
q
n
+
=
Do szacowania skuteczno
ś
ci prób zmiany frekwencji allelu recesywnego na drodze
selekcji stosuje si
ę
nast
ę
puj
ą
cy wzór:
Ocena skuteczno
ś
ci prób wyeliminowania allelu recesywnego ze stada
gdzie:
q
0
- pocz
ą
tkowa frekwencja allelu recesywnego
q
n
– docelowa frekwencja recesywnego allelu po n
pokoleniach
N – liczba pokole
ń
Warto
ść
N informuje nas o liczbie pokole
ń
potrzebnych do osi
ą
gni
ę
cia zamierzonego celu
hodowlanego obliczamy korzystaj
ą
c ze wzoru:
n
n
f
q
f
q
=
=
0
0
0
1
1
q
q
N
n
−
=
gdzie:
f
0
– pocz
ą
tkowa frekwencja osobników b
ę
d
ą
cych
homozygotamu recesywnymi
f
n
– docelowa frekwencja osobników b
ę
d
ą
cych
homozygotamu recesywnymi
Warto
ś
ci q
0
i q
n
obliczamy korzystaj
ą
c ze wzoru:
W stadzie suma kanałowego jest 9% ryb albinotycznych, hodowca chce zredukowa
ć
liczb
ę
albinosów do 0,01% stada, czyli uzyska
ć
stado w którym odsetek sumików
normalnie ubarwionych wynosi 99,99%. Przez ile pokole
ń
suma kanałowego (N)
b
ę
dzie musiał prowadzi
ć
selekcj
ę
aby osi
ą
gn
ąć
zamierzony cel? Ile lat mu to zajmie
wiedz
ą
c ze sum kanałowy dojrzewa płciowo w wieku dwóch lat?
Ocena skuteczno
ś
ci prób eliminacji allelu recesywnego ze stada - Zadanie przykładowe
W pierwszej kolejno
ś
ci obliczamy warto
ś
ci q
0
i q
n
a nast
ę
pnie uzyskane warto
ś
ci
podstawiamy do wzoru na N
01
,
0
0001
,
0
3
,
0
09
,
0
0
0
0
=
=
=
=
=
=
n
n
n
q
f
q
q
f
q
97
67
,
96
3
,
0
1
01
,
0
1
1
1
0
≈
=
−
=
−
=
N
N
q
q
N
n
Odpowied
ź
– hodowca b
ę
dzie musiał prowadzi
ć
selekcj
ę
przez 97 pokole
ń
suma
kanałowego, czyli 194 lata!

Testowanie czy populacja pozostaje w równowadze H-W
Je
ś
li analizujemy populacj
ę
w której mo
ż
emy obliczy
ć
liczb
ę
osobników o
poszczególnych genotypach to jeste
ś
my w stanie okre
ś
li
ć
czy
pozostaje ona w równowadze H-W
(szczegółowy opis ka
ż
dego etapu oblicze
ń
znajduje si
ę
w przewodniku do
ć
wicze
ń
)

Testowanie czy populacja pozostaje w równowadze H-W-
metodyka rozwi
ą
zania zada
ń
1. Obliczenie liczby osobników o ka
ż
dym z analizowanych
genotypów a nast
ę
pnie obliczenie zaobserwowanej frekwencji
genotypów.
2. Okre
ś
lenie frekwencji alleli, wykorzystuj
ą
c zaobserwowane
frekwencje genotypów.
3. Kalkulacja oczekiwanej frekwencji genotypów .
Warno
ś
ci frekwencji alleli nale
ż
y podstawi
ć
do równania H-W a
nast
ę
pnie pomno
ż
y
ć
przez liczebno
ść
stada/populacji,
4. Oznaczenie warto
ść
x
2
wstawiaj
ą
c do wzoru obserwowane i
oczekiwane ilo
ś
ci osobników o danym genotypie. Obliczenia te
prowadzi si
ę
dla jednego stopnia swobody (2 allele).
Obliczanie obserwowanej frekwencji genotypów i frekwencji alleli
Analizuj
ą
c zmienno
ść
genetyczn
ą
w locus koduj
ą
cym białko hemoglobiny
stwierdzono nast
ę
puj
ą
c
ą
liczb
ę
osobników o poszczególnych
genotypach:
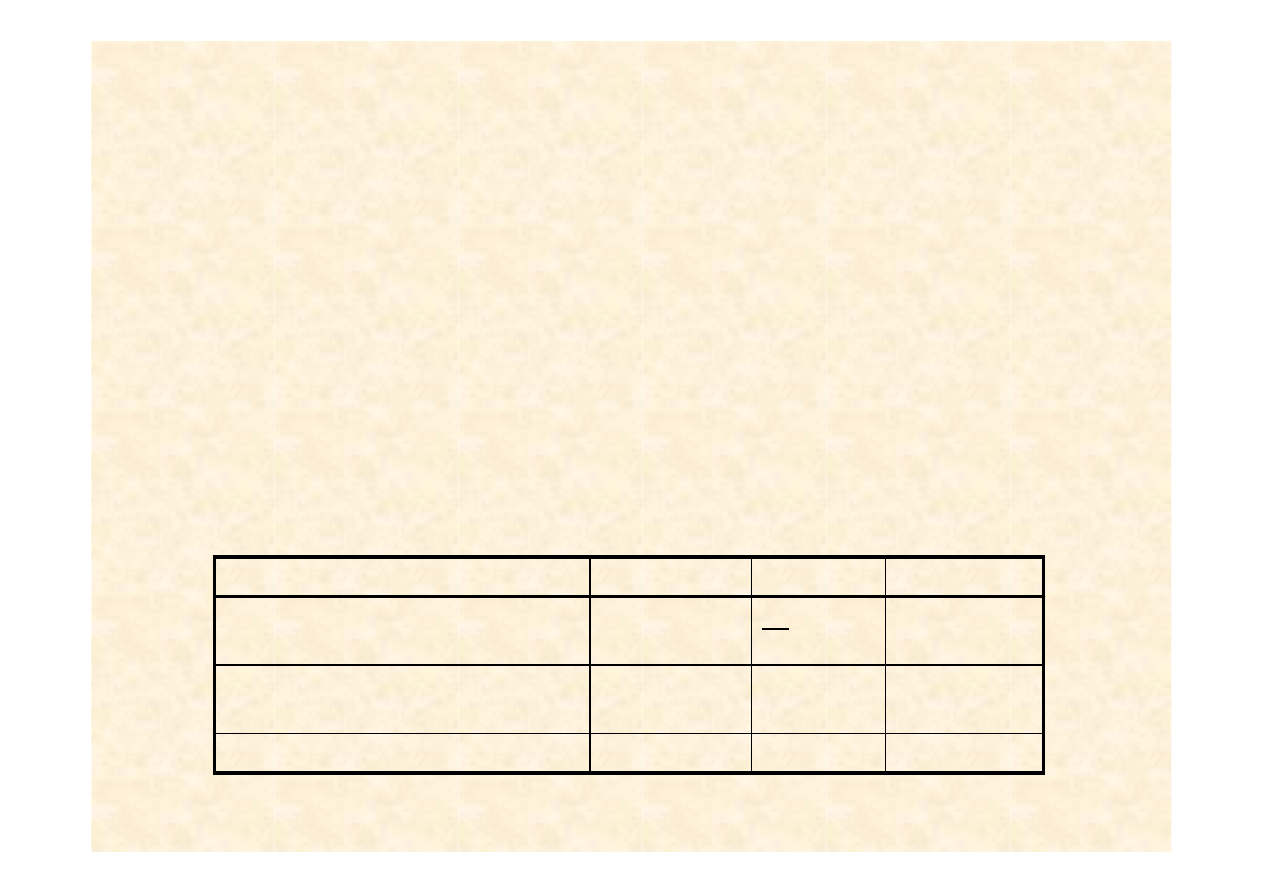
AA - 46/100 = 0,46
Aa – 50/100 = 0,50
aa – 4/100 = 0,04
p = 0,71
q=0,29
Frekwencja alleli
Obserwowana frekwencja
genotypów
Obserwowana liczba
genotypów
genotyp
q = 0,29
p = 0,71
0,04
0,50
0,46
4
50
46
aa
Aa
AA

Genotypy
8
42
50
Oczekiwane liczebno
ś
ci genotypów
q
2
2pq
p
2
Frekwencje genotypów zgodne z
prawem H-W
BB
Bb
bb
p = 0,71
q = 0,29
Frekwencja alleli
8
100
*
29
,
0
42
100
*
29
,
0
*
71
,
0
*
2
50
100
*
71
,
0
1
29
,
0
29
,
0
*
71
,
0
*
2
71
,
0
1
2
2
2
2
2
2
2
≈
=
≈
=
≈
=
=
+
+
=
+
+
bb
Bb
BB
F
F
F
q
pq
p
Oczekiwana frekwencja genotypów jest ilorazem frekwencji genotypów
wynikaj
ą
cej z równania HW i liczebno
ś
ci populacji
Obliczanie oczekiwanej frekwencji genotypów

Warto
ść x
2
:
(Obs - Exp)
2
χ
2
=
Σ -------------------
Exp
Interpretacja:
Je
żeli uzyskana wartość testu x
2
( przy stopniu swobody 1), jest wi
ększa od
tabelarycznej. Hipotez
ę, że populacja pozostaje w równowadze H-W należy
odrzuci
ć.
Testowanie istotno
ś
ci odst
ę
pstw od równowagi H-W
H
o
Populacja jest w w stanie równowagi HW
H
1
populacja nie jest w stanie równowagi HW
gdzie:
Obs – obserwowana frekwencja genotypów
Exp – oczekiwana frekwencja genotypów

Zadanie przykładowe
W składzie tarłowym składaj
ą
cym si
ę
z 100 ryb stwierdzono:
50 ryb o normalnym ubarwieniu (genotyp AA)
40 ryb o ubarwieniu palomino (genotyp Aa)
10 ryb
ż
ółtych (genotyp aa)
Obliczono
ż
e w tym stadzie zgodnie z równowag
ą
H-W frekwencja
poszczególnych genotypów powinna wynosi
ć
Genotyp
n osobników
AA
49
Aa
42
aa
9
Oblicz istotno
ść
odst
ę
pstw od równowagi H-W
Testowanie istotno
ś
ci odst
ę
pstw od równowagi H-W

Obliczanie warto
ś
ci statystyki x
2
Testowanie istotno
ś
ci odst
ę
pstw od równowagi H-W
22
,
0
9
)
9
10
(
42
)
42
40
(
49
)
49
50
(
)
(
2
2
2
2
2
2
2
=
−
+
−
+
−
=
−
=
∑
χ
χ
χ
Exp
Exp
Obs
Obliczanie liczby stopni swobody
Dysponujemy trzema klasami danych (frekwencje trzech genotypów), oraz jednym
badanym parametrem zatem:
Liczba stopni swobody = 3 – 1 - 1 = 1
Liczba stopni swobody jest równa liczbie klas danych pomniejszonych o liczb
ę
badanych parametrów i pomniejszon
ą
o jedno
ść
Warto
ść
krytyczna statystyki x2 odczytanej z tabel dla jednego stopnia swobody i
prawdopodobie
ń
stwie 0,05 = 3,84
Wniosek:
Nie ma podstaw do odrzucenia hipotezy
ż
e ta populacja jest w stanie równowagi H-W

Wyszukiwarka
Podobne podstrony:
Model silnika pradu stalego id Nieznany
Model silnika pradu stalego id Nieznany
Bioreaktor Passavant Model id 8 Nieznany (2)
PKM Model Fenomenologicznyv3 id Nieznany
6 Mimezis model modelowanie id Nieznany (2)
Model odpowiedz II etap 2012 id Nieznany
pdf anuiaaaPoprawiona Model id Nieznany
IMW W04 Model mech podnosz id 2 Nieznany
Bioreaktor Passavant Model id 8 Nieznany (2)
PKM Model Fenomenologicznyv3 id Nieznany
cw 16 odpowiedzi do pytan id 1 Nieznany
Opracowanie FINAL miniaturka id Nieznany
How to read the equine ECG id 2 Nieznany
PNADD523 USAID SARi Report id 3 Nieznany
OPERAT STABLE VERSION ugoda id Nieznany
biuletyn katechetyczny pdf id 8 Nieznany
Finanse publiczne cw 4 E S id 1 Nieznany
więcej podobnych podstron