
64
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
Ptaki uwa¿ane s¹ za dobry wskaŸnik stanu œro-
dowiska przyrodniczego. Zmiany liczebnoœci
populacji ptaków s¹ najczêœciej wynikiem prze-
kszta³cania siê siedlisk, a wiêc dostatecznie wcze-
sne rozpoznanie tych zmian i iloœciowa ocena ich
zakresu mo¿e wnieœæ istotny wk³ad w tworzenie
ogólnych programów ochrony przyrody. Poza
tym spotykamy wœród nich wiele gatunków skraj-
nie zagro¿onych i gin¹cych (endangered), nara-
¿onych na wyginiêcie (vulnerable) lub rzadkich
(rare) (Tucker&Heath 1995), które nale¿y chro-
niæ w sposób skuteczny, a wiêc wynikaj¹cy ze
znajomoœci istoty tego zagro¿enia. Szczególn¹
rolê mog¹ tu odegraæ parki narodowe, które przy
istnieniu odpowiednich, precyzyjnych i stale ak-
tualizowanych, danych o zagro¿onych gatunkach
powinny staæ siê nie tylko ich ostoj¹, ale œrodo-
wiskami Ÿród³owymi (source habitats), zapewnia-
j¹cymi im nie tylko odpowiedni¹ liczebnoœæ, ale
tak¿e skuteczny rozród i mo¿liwie nisk¹ œmier-
telnoœæ. Dziêki temu bêd¹ one mog³y zasilaæ tak-
¿e inne tereny, nawet te funkcjonuj¹ce jako œro-
dowiska ujœciowe (sink habitats).
Spoœród 22 lêgowych gatunków ptaków, wy-
stêpuj¹cych w Kampinoskim Parku Narodowym
i jego otulinie obecnie lub w minionym piêædzie-
siêcioleciu, a umieszczonych w Polskiej Czerwo-
nej Ksiêdze Zwierz¹t (G³owaciñski 2001), a¿ sie-
dem to ptaki drapie¿ne i sowy, które z wielu
wzglêdów wymagaj¹ szczególnej ochrony – osi¹-
gaj¹ zazwyczaj niewielkie zagêszczenia, maj¹
szczególne wymagania gniazdowe, póŸno osi¹-
gaj¹ wiek rozrodczy i charakteryzuj¹ siê na ogó³
nisk¹ rozrodczoœci¹, a przy tym czêsto bywaj¹
przeœladowane przez cz³owieka. Ponadto stano-
wi¹ ostatnie ogniwo w ³añcuchu troficznym,
a wiêc nara¿one s¹ na akumulowanie zwi¹zków
toksycznych ze œrodowiska, co u d³ugo ¿yj¹cych
gatunków przez wiele lat mo¿e hamowaæ rozród.
Tak wiêc bez dok³adnej znajomoœci aktualnej
sytuacji gatunku na konkretnym terenie jego
ochrona mo¿e okazaæ siê nieskuteczna.
Monitoring sk³adu gatunkowego, liczebnoœci
i rozmieszczenia ptaków drapie¿nych w KPN roz-
poczêto w 1980 r. i prowadzony jest do chwili
obecnej. Jego celem jest ustalenie aktualnego sta-
nu gatunkowego (w tym tak¿e stwierdzenie z du-
¿ym prawdopodobieñstwem, ¿e dany gatunek ju¿
nie wystêpuje) oraz ocena rozmieszczenia i pa-
rametrów rozrodu ptaków. Sta³a metodyka pra-
cy to wyszukiwanie w okresie bezlistnym wszyst-
kich du¿ych gniazd, a w sezonie lêgowym
ustalanie zajêtych rewirów i w miarê mo¿liwoœci
dwukrotna kontrola „z ziemi” gniazd zasiedlo-
nych. W ten sposób mo¿na stwierdziæ wysiady-
wanie, a wiêc obecnoœæ jaj, oraz brak lub obec-
noœæ m³odych w gniazdach. W próbie 20 – 40%
zajêtych gniazd kontrolowana jest bezpoœrednio
wielkoœæ zniesienia i liczba wychowanych m³o-
dych, czyli sukces lêgowy, parametry niezbêd-
ne, by stworzyæ modele funkcjonowania popu-
lacji poszczególnych gatunków na badanym
terenie.
Powy¿sza metodyka dotyczy leœnych ptaków
drapie¿nych. O gatunkach terenów otwartych
zbierane s¹ ogólne dane, dotycz¹ce jedynie liczeb-
noœci i rozmieszczenia par lêgowych, bez wyszu-
kiwania i kontroli gniazd.
Porównanie uzyskanych danych z wczeœniej-
szymi danymi z literatury pozwoli³o na stwier-
dzenie, ¿e w ostatnim æwieræwieczu ust¹pi³y
z Parku 2 gatunki lêgowe – gado¿er Circaetus
gallicus i sokó³ wêdrowny Falco peregrinus, na-
tomiast pojawi³ siê nienotowany dotychczas orze³
bielik Haliaetus albicilla. Obecnie wystêpuje
w KPN 9 gatunków lêgowych i 5 zalatuj¹cych,
których sytuacja obecna i historia s¹ znane. Wie-
loletnie dane o rozrodzie tych niezbyt licznych
gatunków pozwalaj¹ zgromadziæ dostateczny ma-
Bogumi³a Olech
Kampinoski Park Narodowy
Dzia³ Nauki i Monitoringu Przyrody
MONITORING PTAKÓW
W KAMPINOSKIM PARKU
NARODOWYM W LATACH 1980
–2002

65
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
teria³, by rozwa¿aæ tak¿e specjalistyczne zagad-
nienia ich biologii czy behawioru (Olech 1997,
Olech, Pruszyñski 2000).
Jednym z lepiej poznanych gatunków jest ja-
strz¹b Accipiter gentilis. Na pocz¹tku badañ, kie-
dy jeszcze zagêszczenie jastrzêbi w Polsce uwa-
¿ane by³o za niskie, stwierdzono tu 14 par/100
km
2
powierzchni ogólnej, czyli jedno z najwy¿-
szych zagêszczeñ w Europie. W ci¹gu 22 lat spa-
d³o ono o prawie 30%, zapewne z powodu
zmniejszania siê antropogennej bazy pokarmo-
wej, zwi¹zanej z prowadzonymi w parku wyku-
pami i likwidacjami wsi. Wieloletnie dane pozwo-
li³y porównaæ sukces rozrodczy i œmiertelnoœæ
gniazdow¹ w okresach o ró¿nym zagêszczeniu
jastrzêbi, a tak¿e zanalizowaæ skutecznoœæ prze-
pisów ochronnych dla tego gatunku. Nieoczeki-
wanie wykryto stopniowe zmniejszanie siê objê-
toœci jaj jastrzêbi, a tak¿e zwiêkszenia udzia³u
niezalê¿onych jaj lub/i zamar³ych zarodków. Jest
to zjawisko wymagaj¹ce badañ naukowych w ce-
lu jego wyjaœnienia. (Olech 1986, 1996, Olech,
Pruszyñski – dane niepublikowane).
W póŸnych latach 90. stwierdzono wzrost li-
czebnoœci krogulca Accipiter nisus, trwaj¹cy do
2001 r. U innych gatunków nie stwierdzono wy-
raŸnych zmian liczebnoœci poza myszo³owem
Buteo buteo, którego niezbyt wysoka tu liczeb-
noϾ Р44,2 pary/100 km
2
(Pruszyñski – praca
doktorska, dane niepublikowane) waha siê
z 1–2-letnim opóŸnieniem wraz ze zmieniaj¹c¹
siê baz¹ pokarmow¹, któr¹ stanowi¹ g³ównie
drobne gryzonie.
Innym du¿ym gatunkiem leœnym, monitorowa-
nym w Kampinoskim Parku Narodowym od po-
nad 20 lat, jest bocian czarny Ciconia nigra.
Gniazda wynajdywane s¹ podczas kontroli gniazd
ptaków drapie¿nych, niekiedy zreszt¹ zajmowane
s¹ przez te gatunki zamiennie. Dziêki wieloletnim
materia³om przeœledzono zmiany liczebnoœci
czarnych bocianów na tle jego wystêpowania
na Mazowszu (Kot, Olech 1995), zwi¹zane ze
zmianami poziomu wód gruntowych oraz z tren-
dem ogólnym u tego gatunku. Przez wiele lat
utrzymuj¹ca siê liczebnoœæ oko³o 10 par lêgo-
wych spad³a w latach dziewiêædziesi¹tych do 3
– 4 par, by w ci¹gu ostatnich kilku lat osi¹gn¹æ
liczbê 15–17. Prawdopodobnie czêœæ z nich to
jednak ptaki niedojrza³e, gdy¿ np. w 2002 r. na
17 par poprawiaj¹cych lub buduj¹cych gniazda
z³o¿y³o jaja tylko 9 par (Zawadzka, Olech, Za-
wadzki 1989, Olech, Pruszyñski – dane niepu-
blikowane).
Inn¹ grup¹ gatunków monitorowanych na te-
renie ca³ego parku s¹ chruœciele – ptaki dolin
rzecznych, szczególnie, jak siê wydaje, zagro¿o-
ne tutaj obni¿aj¹cym siê poziomem wód grunto-
wych i zwiêkszaniem powierzchni leœnej w KPN.
Najd³u¿ej, bo od 1996 r. kontrolowana jest liczeb-
noϾ i rozmieszczenie derkacza Crex crex. Ten
euroazjatycki gatunek nie znalaz³ siê wprawdzie
w Polskiej Czerwonej Ksiêdze, ale uwa¿any jest
za nara¿ony na wyginiêcie w skali globalnej, gdy¿
jego liczebnoœæ w ca³ym areale wystêpowania,
a szczególnie w Europie, gwa³townie spada (Tuc-
ker&Heath 1995). Liczenie derkaczy prowadzo-
ne jest metod¹ nocnego nas³uchu na wszystkich
otwartych terenach Parku i czêœciowo jego otuli-
ny na prze³omie maja i czerwca oraz czerwca i lip-
ca, czyli przed I i przed II lêgiem. Bierze w nich
udzia³ oko³o 30 osób, które z wyznaczonych
punktów okreœlaj¹ azymuty do odzywaj¹cych siê
samców. Uzyskane w ten sposób dane naniesio-
ne na mapê pozwalaj¹ zlokalizowaæ poszczegól-
ne samce, a wiêc i oceniæ ich zagêszczenie, które
okaza³o siê jednym z najwy¿szych w Europie,
w latach 1996 –97 osi¹gaj¹c lokalnie do 12 sam-
ców/km
2
(Juszczak, Olech 1997). LiczebnoϾ der-
kaczy w Kampinoskim Parku Narodowym waha
siê: od maksymalnej dotychczas wykrytej – 290
samców, do najni¿szej w suchym 2000 r., kiedy
by³o ich zaledwie oko³o 90. Odbywaj¹ tu 2 lêgi
rocznie, co przy faktycznym braku koszenia ³¹k,
stanowi¹cym, szczególnie dla gniazd i piskl¹t,
du¿e zagro¿enie, pozwala uznaæ KPN za teren
Ÿród³owy dla tego gatunku. Sytuacja zmieni siê,
jeœli poprzez podniesienie poziomu wody lub
przez koszenie nie powstrzymamy sukcesji ro-

66
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
œlinnoœci drzewiastej na obecnych terenach otwar-
tych.
W podobny sposób od 2000 r. monitorowana
jest w KPN liczebnoœæ i rozmieszczenie par lê-
gowych ¿urawi Grus grus. Nas³uch prowadzi siê
w ci¹gu godziny przed i po wschodzie s³oñca
w marcu i na pocz¹tku kwietnia, bezpoœrednio po
przylocie ptaków. W tym okresie wydawane
o œwicie g³osy nawo³uj¹cych siê par pochodz¹
z miejsc, w których bêd¹ budowane gniazda. Me-
toda azymutowa, podobnie jak u derkaczy, po-
zwala przy du¿ym nawet zagêszczeniu ¿urawi
oceniæ ich liczebnoœæ. W badanym okresie utrzy-
mywa³a siê ona w Kampinoskim Parku Narodo-
wym na wysokim poziomie 35 par.
Przy okazji nocnych nas³uchów derkaczy i ¿u-
rawi lokalizowane s¹ tak¿e miejsca odzywania
siê b¹ka Botaurus stellaris, gatunku œciœle zwi¹-
zanego z biocenozami wodno-bagiennymi, naj-
bardziej zagro¿onymi w Europie. Dotychczas
wykryto 6 stanowisk, z których nie wszystkie by³y
zajmowane w kolejnych latach.
Oprócz w³asnych programów monitoringu
ptaków pracownicy KPN uczestnicz¹ w dwóch
wieloletnich programach ogólnopolskich, koor-
dynowanych przez Zak³ad Ornitologii PAN
w Gdañsku. Jeden z nich, to rozpoczêty w 2001 r.
Monitoring Flagowych Gatunków Ptaków
(MFGP), finansowany przez G³ówny Inspekto-
rat Ochrony Œrodowiska. Jego celem jest uzyska-
nie wskaŸników liczebnoœci populacji lêgowych
4 gatunków o zasiêgu ogólnokrajowym: bociana
bia³ego Ciconia ciconia, ³abêdzia niemego Cy-
gnus olor, gawrona Corvus frugilegus i ¿urawia
Grus grus, a od 2002 r. dodatkowo b¹ka Botau-
rus stellaris i b³otniaka stawowego Circus aeru-
ginosus. Ocena odbywa siê na 30 losowo wybra-
nych powierzchniach 100 km
2
, z których
powierzchnia nr MW2 znalaz³a siê czêœciowo na
terenie gmin Czerwiñsk i Brochów, a wiêc w za-
chodniej czêœci KPN i jego otuliny. Dotychczas
zebrane dane wskazuj¹, ¿e 19 z 21 gniazd bocia-
nów bia³ych i wszystkie 3 pary lêgowe ¿urawi
znajduj¹ siê po kampinoskiej stronie Wis³y.
Drugi z programów to krajowy wariant Con-
stant Effort Sites (CES) – programu rozpoczête-
go w Wielkiej Brytanii w 1983 r., a w Polsce
w 2000 r. pod nazw¹ GIOŒ Sta³e Powierzchnie
Od³owu. G³ównym celem programu jest wielo-
letnie monitorowanie zmian liczebnoœci oraz
struktury gatunkowej i wiekowej drobnych lêgo-
wych ptaków wróblowatych. Dane pochodziæ
maj¹ z 8–12 od³owów ptaków, wykonanych
w okresie od maja do sierpnia w wyznaczonych
godzinach, w sta³ych miejscach i w sta³¹ liczbê
sieci. Udzia³ w programie mog¹ braæ wy³¹cznie
osoby o uprawnieniach obr¹czkarskich. W Kam-
pinoskim Parku Narodowym punkt taki funkcjo-
nuje od 2002 r. w okolicy Górek Kampinoskich,
w sezonie 2002 podczas serii 10 od³owów zebra-
no dane o 113 osobnikach z 17 gatunków.
Do wszystkich prezentowanych tu tematów
monitoringu ptaków sporz¹dzane s¹ odpowied-
nie komputerowe bazy danych. Mamy tak¿e na-
dziejê, ¿e rozmaite przejœciowe trudnoœci nie
zmusz¹ nas do przerwania badañ.
Literatura
G³owaciñski Z. (ed.) 2001. Polska Czerwona
Ksiêga Zwierz¹t – Krêgowce. Pañstwowe Wy-
dawnictwo Rolnicze i Leœne. Warszawa.
Juszczak K., Olech B. 1997. LiczebnoϾ i roz-
mieszczenie derkacza Crex crex na terenach
otwartych Kampinoskiego Parku Narodowego
i jego okolic w latach 1996–1997. Not.Orn. 38,3:
197–213.
Kot H., Olech, B. 1995. Raport z realizacji pro-
gramu „Ochrona bociana czarnego i jego siedlisk
w œrodkowo-wschodniej Polsce” w 1995 r. Wo-
jewództwo sto³eczne warszawskie. Siedlce 1995.
Olech B. 1991. Ochrona ptaków drapie¿nych
w Kampinoskim Parku Narodowym – stan
i wskazania. Ochrona Przyrody. 49,1 : 65–79.
Olech B. 1992. O ptakach drapie¿nych w Kam-
pinoskim Parku Narodowym. Puszcza Kampino-
ska 3(92) : 12–13.
Olech B. 1997. Diet of the Goshawk Accipiter
gentilis in Kampinoski National Park (Central

67
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
Poland) in 1982–1993. Acta Orn. 32,2 :191–200.
Olech B. 1998. Population dynamics and bre-
eding performance of the Goshawk Accipiter
gentilis in Central Poland in 1982–1994. In:
Chancellor R. D., Meyburg B. U., Ferrero J. J.
(eds.): Holarctic Birds of Prey. ADENEX-
WWGBP: 101–110.
Olech B., Pruszyñski M. 2000. Food caching
or surplus killing in the Common Buzzard Buteo
buteo ?Acta Orn. 35,2 : 215–216.
Olech B. 2001. Orze³ bielik w Puszczy Kam-
pinoskiej. Puszcza Kampinoska 4(32) : 1–2.
Tucker G. M., Heath M. F. 1995. BirdLife Con-
servation Series No. 3.
Zawadzka D., Olech B., Zawadzki J. 1990. Za-
gêszczenie, rozród i pokarm bociana czarnego Ci-
conia nigra w Kampinoskim Parku Narodowym
w latach 1979–1987. Not. Orn.31, 1– 4: 5–20.
Anna Andrzejewska
Kampinoski Park Narodowy
MONITORING WÓD
W KAMPINOSKIM PARKU
NARODOWYM – TRZY LATA
OBSERWACJI
Wed³ug podzia³u administracyjnego Polski re-
zerwat le¿y w województwie mazowieckim, na
terenie 6 powiatów oraz 17 gmin.
W porównaniu do s¹siednich obszarów rów-
ninnych teren ten charakteryzuje siê wyj¹tkowo
zró¿nicowan¹ rzeŸb¹. Ró¿nice wysokoœci
wzglêdnej dochodz¹ tu do 30 m. Najwy¿ej po³o-
¿ony punkt na terenie rezerwatu znajduje siê
w pobli¿u jego wschodniej granicy (107,4 m
n.p.m.), a najni¿szy – przy ujœciu Bzury do Wi-
s³y (66,7 m n.p.m.)
Ca³y obszar ma wyraŸn¹ budowê pasow¹.
Wyró¿nia siê tu nastêpuj¹ce obszary: wspó³cze-
sne koryto Wis³y ograniczone wa³ami przeciw-
powodziowymi, dwa poziomy tarasu zalewowe-
go Wis³y, pó³nocny pas wydmowy, pó³nocny
(œrodkowy) pas bagienny, stanowi¹cy dolinê
£asicy, po³udniowy pas wydmowy, po³udniowy
pas bagienny, skarpê pradoliny Wis³y, Równinê
B³oñsk¹.
Obszar rezerwatu niemal w ca³oœci po³o¿o-
ny jest w zlewni £asicy, która jest najwa¿niej-
szym ciekiem wodnym Puszczy Kampinoskiej.
W znacznej czêœci swojego biegu jest obecnie
sztucznie przekopanym kana³em. Prace meliora-
cyjne, maj¹ce na celu osuszenie bagien puszczañ-
skich, rozpoczê³y siê w XIX wieku, a zakoñczy-
³y w latach 1968–1972 wybudowaniem szeregu
jazów i innych budowli piêtrz¹cych. Dwa z nich,
wybudowane w latach 1997–1998, posiadaj¹ sze-
rokie przep³awki w œrodku nurtu rzeki, dziêki
czemu nie stanowi¹ bariery ekologicznej. £asica
ma kilka dop³ywów, z których najwiêkszymi s¹:
prawobrze¿ny kana³ £-9 i lewobrze¿ne: Kana³
Olszowiecki oraz Kana³ Zaborowski.
Na omawianym terenie znajduje siê te¿ szereg
ró¿nej wielkoœci jezior i mniejszych zbiorników
wodnych. Najwiêksze z nich to starorzecza le¿¹-
ce w obrêbie tarasów zalewowych Wis³y. S¹ to:
jeziora Kie³piñskie i Dziekanowskie w rejonie
£omianek oraz jeziora Dolne i Górne w rejonie
Kazunia.
Niemal na ca³ym obszarze rezerwatu (oprócz
Równiny B³oñskiej, gdzie notuje siê kilka pozio-
1. Wstêp
Rezerwat Biosfery UNESCO MaB „Puszcza
Kampinoska” rozci¹ga siê miêdzy 52°26` a 52°
14`N oraz 20°12` a 20°59`E. Po³o¿ony jest na
Nizinie Œrodkowomazowieckiej w po³udniowo-
zachodniej czêœci Kotliny Warszawskiej i pó³noc-
nej czêœci Równiny £owicko-B³oñskiej (Kon-
dracki 1981), na pó³nocny zachód od Warszawy.
Obszar rezerwatu wyznacza granica otuliny Kam-
pinoskiego Parku Narodowego. Od pó³nocy
i pó³nocnego wschodu stanowi j¹ prawy brzeg
Wis³y, od wschodu granica przebiega przedmie-
œciami Warszawy, od po³udnia biegnie rolniczy-
mi obszarami Równiny-£owicko-B³oñskiej, od
zachodu granicê stanowi lewy brzeg Bzury. Re-
zerwat le¿y na obszarze wêz³a hydrologicznego
Wis³y, Bugu, Narwi, Wkry i Bzury.

68
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
mów wodonoœnych) mamy do czynienia z jed-
nym ci¹g³ym poziomem wodonoœnym, obejmu-
j¹cym ca³y pakiet utworów czwartorzêdowych,
o mi¹¿szoœci od 10 do 50 m i znacznych mo¿li-
woœciach przewodzenia wody.
Zasilanie wód podziemnych pochodzi g³ównie
z przesi¹kania wód opadowych, przede wszyst-
kim w obrêbie wystêpuj¹cych tu pasów wydmo-
wych. Po³udniowa czêœæ tarasu zasilana jest wo-
dami dop³ywaj¹cymi z warstw wodonoœnych
poziomu b³oñskiego.
Odp³yw wód odbywa siê g³ównie na obszarach
bagiennych systemem rowów i kana³ów melio-
racyjnych oraz w wyniku ewapotranspiracji,
zw³aszcza w okresie wegetacyjnym.
2. Cele i program monitoringu wód
Na terenie Kampinoskiego Parku Narodowe-
go od wielu lat prowadzone by³y badania hydro-
logiczne. Mia³y one najczêœciej zakres lokalny
(Kazimierski 1980, Sikorska-Maykowska 1981,
Krogulec 1995).
W latach 1985–1990 prowadzony by³ monito-
ring w sieci, któr¹ tworzy³o ponad 100 punktów
obserwacyjnych wód podziemnych (studni i pie-
zometrów) i 25 punktów wodowskazowych. Po-
miary prowadzone by³y przez okres 5 lat, tj. od
1985 do 1990 r. raz na dwa tygodnie (Kazimier-
ski i in. 1984, 86, 87, 88, 89, 90, Sikorska-May-
kowska, Kazimierski 1989). Wyniki monitorin-
gu oraz kompleksowe badania w zakresie oceny
warunków hydrogeologicznych na terenie Parku
pos³u¿y³y do przygotowania Planu Ochrony
Kampinoskiego Parku Narodowego (Kazimier-
ski i in. 1995).
Obecnie w Rezerwacie Biosfery „Puszcza
Kampinoska” monitoring elementów abiotycz-
nych i biotycznych prowadzony jest w dwóch
skalach przestrzennych: bardziej szczegó³owej –
w ramach Zintegrowanego Monitoringu Œrodo-
wiska Przyrodniczego
1
w Stacji Bazowej „Po¿a-
ry”, której zlewnia obejmuje obszar 20,17 km
2
oraz bardziej ogólnej na ca³ym obszarze rezer-
watu o powierzchni 762 km
2
.
Stacja Bazowa ZMŒP „Po¿ary” dzia³a od roku
1994. Na terenie górnej czêœci zlewni kana³u Ol-
szowieckiego w tzw. kotlinie „Po¿ary” zainsta-
lowane s¹ 22 piezometry s³u¿¹ce do pomiaru
po³o¿enia zwierciad³a oraz badañ chemizmu wód
podziemnych (Fic, Wierzbicki 1994).
W roku 1998 rozpoczêto monitoring wód grun-
towych i powierzchniowych w sieci piezometrów
i punktów wodowskazowych na obszarze ca³ego
Parku wraz z otulin¹. W zwi¹zku z obni¿aniem
siê poziomu lustra wód powierzchniowych i pod-
ziemnych w Planie Ochrony KPN zaproponowa-
no wprowadzenie zmian w gospodarce wodnej
tego obszaru, która pierwotnie by³a ukierunko-
wana na potrzeby rolnictwa, a obecnie musi s³u-
¿yæ ochronie i restytucji siedlisk roœlinnych.
Wskazano wiele dzia³añ zmierzaj¹cych do pod-
niesienia poziomu wód na obszarze ca³ego Par-
ku. Aby podj¹æ to zadanie, nale¿a³o stworzyæ
system obserwacji stanu i jakoœci wód.
Monitoring zmian poziomu lustra wód po-
wierzchniowych i podziemnych, a tak¿e bilansu
wodnego na terenie Puszczy Kampinoskiej, ma
kilka celów:
– dostarczanie bie¿¹cych danych o stanie
i zmianach poziomu wód, co umo¿liwia podej-
mowanie szybkich decyzji;
– zbieranie danych, które umo¿liwiaj¹ analizê
obliczanie bilansu obiegu wody i jego zmian
w d³ugich okresach czasu;
– dostarczanie danych przed, w czasie i po
wprowadzeniu zmian zmierzaj¹cych do renatu-
ralizacji zasobów wodnych Parku;
– dostarczanie danych dla badañ naukowych
z dziedzin hydrologii, hydrogeologii, a tak¿e z in-
nych dotycz¹cych przyrody Parku;
– dostarczanie danych technicznych umo¿li-
wiaj¹cych rzeczow¹ dyskusjê z przedstawiciela-
mi lobby hydrotechnicznego;
– umo¿liwienie modelowania i prognozowa-
nia zmian obiegu wody.
1
Zintegrowany Monitoring Œrodowiska Przyrodnicze-
go jest jednym z podsystemów Pañstwowego Monitorin-
gu Œrodowiska

69
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
Zakres monitoringu wód oraz sieæ punktów
obserwacyjnych zosta³y zaprojektowane przez
hydrogeologów (E. Krogulec, M. Sikorska-May-
kowska 1996). Podstawowym celem projektowa-
nej sieci monitoringu lokalnego by³o stworzenie
bazy piezometrów i punktów wodowskazowych,
przystosowanych do prowadzenia obserwacji
aktualnego stanu stosunków wodnych. Ogólna
koncepcja lokalizacji punktów obserwacyjnych
zosta³a podporz¹dkowana nastêpuj¹cym uwarun-
kowaniom:
– objêcie obserwacjami wszystkich wystêpu-
j¹cych na terenie KPN stref hydro-dynamicznych,
– uwzglêdnienie wp³ywu czynników antropo-
presyjnych na chemizm wód podziemnych i po-
wierzchniowych,
– nawi¹zanie do poprzednio istniej¹cej sieci,
tak by jej wyniki mog³y byæ wykorzystywane
przy analizie pomiarów projektowanej sieci mo-
nitoringu,
– nawi¹zanie do zakresu proponowanych dzia-
³añ hydrotechnicznych zawartych w „Operacie
ochrony i kszta³towania stosunków i ekosyste-
mów wodnych w obrêbie Kampinoskiego Parku
Narodowego”,
– techniczna mo¿liwoœæ prowadzenia obserwa-
cji hydrogeologicznych i hydrologicznych
(³atwoœæ dojazdu, bezproblemowa lokalizacja
punktu itp.).
Realizacja sieci monitoringu wód powierzch-
niowych i podziemnych by³a mo¿liwa dziêki
dofinansowaniu z Narodowego Funduszu Ochro-
ny Œrodowiska i Gospodarki Wodnej. £¹cznie
zainstalowano lub zaadaptowano 74 punkty ob-
serwacyjne, w tym 55 wód podziemnych i 19 wód
powierzchniowych (rys. 1).
Po zakoñczeniu prac terenowych zosta³a opra-
cowana „Dokumentacja hydrologiczna z wyko-
nanych piezometrów wraz z programem moni-
toringu na obszarze Kampinoskiego Parku
Narodowego” (M. Sikorska-Maykowska, E. Kro-
gulec, M. Fic 1997).
Punkty wodowskazowe maj¹ postaæ bolców
metalowych (reperów) umieszczonych na most-
kach lub budowlach hydrotechnicznych. Ponad-
to w punkcie zamykaj¹cym zlewnie kana³u £asi-
ca i Kana³u Olszowieckiego na granicy KPN za-
instalowane s¹ ³aty hydrometryczne. Wszystkie
punkty obserwacyjne zosta³y zaniwelowane.
Monitoring prowadzony jest od pocz¹tku roku
hydrologicznego 1998 (1.11.1997). Stany wód
podziemnych i powierzchniowych s¹ mierzone
we wszystkich punktach obserwacyjnych co dwa
tygodnie (w poniedzia³ki). W okresie tzw. sta-
nów ni¿ówkowych i wy¿ówek pomiary wyko-
nuje siê raz na tydzieñ.
Obserwacje prowadzone w ramach monitorin-
gu wód podziemnych i powierzchniowych uzu-
pe³niane s¹ danymi meteorologicznymi. Na tere-
nie Parku znajduj¹ siê 3 stacje meteorologiczne
i 6 posterunków opadowych. Dane te umo¿liwiaj¹
obliczanie bilansu wodnego tego obszaru.
W latach 1999–2002 prowadzone by³y kom-
pleksowe badania bilansu wodnego Zlewni £asi-
cy we wspó³pracy z Amerykañsk¹ S³u¿b¹ Parków
Narodowych (National Park Service), w ramach
grantu z II Polsko-Amerykañskiego Funduszu im.
M. Sk³odowskiej-Curie. W badaniach tych udzia³
wziê³y: Dzia³ Nauki i Monitoringu Przyrody oraz
zespo³y naukowe z Zak³adu Hydrologii Wydzia-
³u Geografii i Studiów Regionalnych oraz Zak³a-
du Hydrologii Wydzia³u Geologii Uniwersytetu
Warszawskiego. W badaniach tych obok wielo-
letnich danych uzyskanych z Instytutu Meteoro-
logii i Gospodarki Wodnej wykorzystano m.in.
dane z sieci monitoringu wód KPN.
4. Wyniki trzyletnich obserwacji
Opady na stacji meteorologicznej w Granicy
mierzone s¹ od roku 1985. W tym okresie œred-
nia roczna suma opadów wynios³a 546,9 mm.
Najwy¿sze opady notowane s¹ w lipcu 92,6 mm,
a najni¿sze w styczniu 23,9 i lutym 25,7 mm.
W ostatnich trzech latach pod wzglêdem opa-
dów lata 1999 i 2001 by³y przeciêtne (odpowied-
nio 569,6 mm i 565,6 mm), natomiast rok 2000 –
suchy (413,9 mm). W tym okresie najwy¿sze opa-
dy (162,8 mm) zanotowano w czerwcu 1999 roku,

70
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
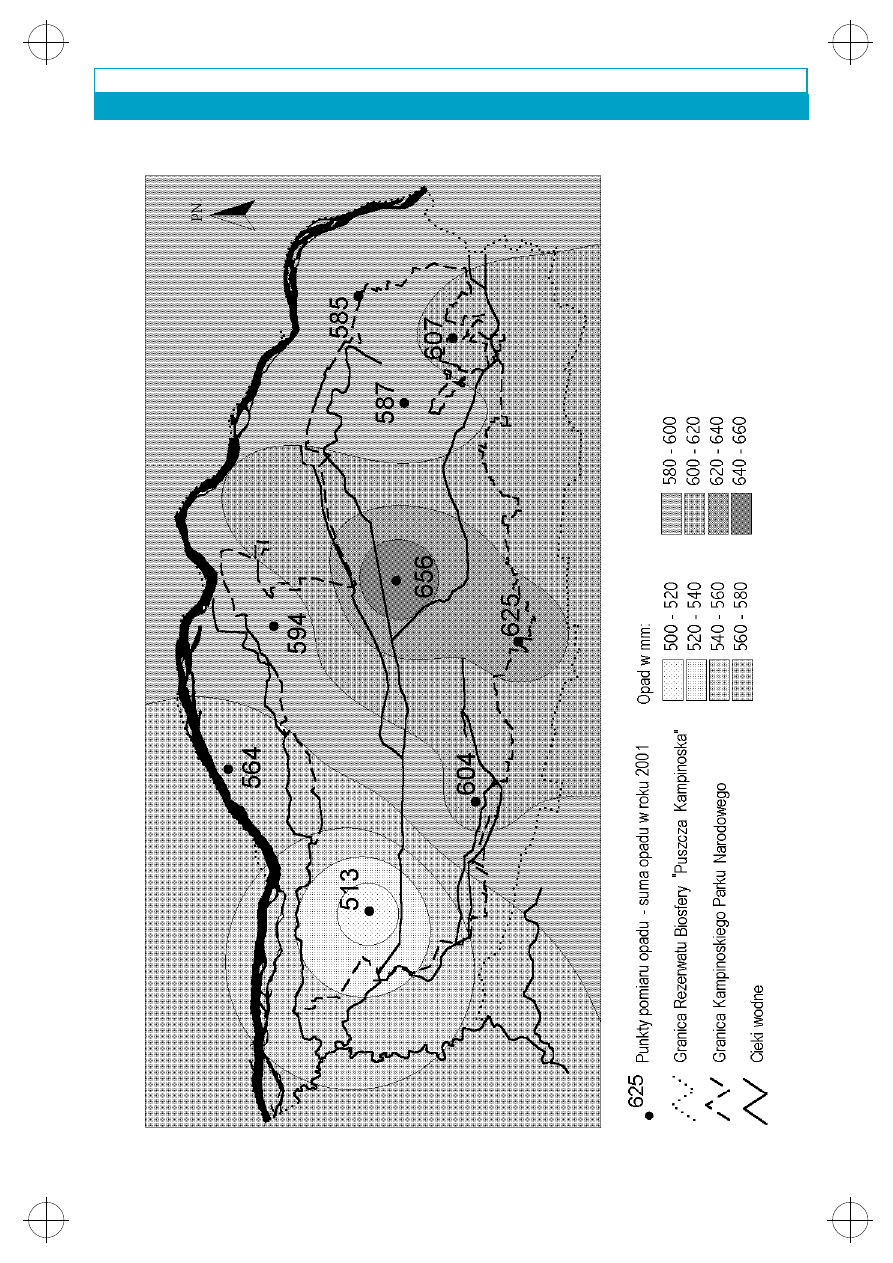
Rys. 2. Miesiêczne sumy opadu w latach hydrologicznych 1999–2000 zmierzone w Granicy
Rys. 1

71
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
a najni¿sze (3 mm) w kwietniu 2000 roku (rys.
2).
Widoczne jest zró¿nicowanie geograficzne
opadów na terenie Puszczy. Na podstawie danych
z posterunków opadowych Kampinoskiego Par-
ku Narodowego i Instytutu Meteorologii i Go-
spodarki Wodnej wykreœlono mapy izolinii o tych
samych wielkoœciach opadów. Rozpatrywano
pe³ny rok obserwacji 2001. Najbardziej zró¿ni-
cowane opady zanotowano w lipcu, kiedy ró¿ni-
ce pomiêdzy obszarami o najni¿szych i najwy¿-
szych opadach dochodzi³y do 90,9 mm. We
wszystkich porach roku zaznacza siê obszar o ni¿-
szych opadach w zachodniej lub pó³nocno-za-
chodniej czêœci Puszczy oraz wy¿sze opady
w centralnej jej czêœci. Najwy¿sze opady w roku
2001 (656 mm) wyst¹pi³y w centrum Parku we
wsi Kiœcinne, natomiast najni¿sze (513 mm), po-
dobnie jak w roku poprzednim w zachodniej czê-
œci Parku w Famu³kach Królewskich (rys. 3).
Trzyletnie obserwacje stanów wód gruntowych
s¹ niewystarczaj¹ce, by œwiadczyæ o d³ugotermi-
nowych zmianach poziomu wód. W celu okre-
œlenia zamian w poziomie wód gruntowych, dla
ca³ego obszaru Parku, obliczono œredni poziom
wody pod powierzchni¹ terenu (p.p.t) z wszyst-
kich piezometrów. Œrednia ta dla trzech lat wy-
nios³a 1,70
±
0,20 m p.p.t. Najwy¿sze stany wody
(1,30 m p.p.t.) zanotowano 28.06.1999, a najni¿-
sze 2,04 m p.p.t. – 30.10.2000. W okresie bada-
nych trzech lat obserwowano zmniejszaj¹cy siê
trend zmian poziomu lustra wody. WyraŸny
wp³yw na tê tendencje mia³ bardzo suchy rok
2000, kiedy roczna suma opadu wynios³a 406
mm. W roku 2001 zasoby wodne nie powróci³y
do stanów z roku 1999. Tak wiêc roczne staty-
styki: œredni poziom wody, mediana oraz poziom
maksymalny w badanym okresie opada³y. Jedy-
nie najni¿szy roczny poziom wody by³ wy¿szy
w roku 2001 ni¿ w roku 2000.
Oprócz rocznych zmian poziomu wody wi-
doczne s¹ zmiany sezonowe. Najwy¿sze stany
notowane s¹ wiosn¹. W latach 2000 i 2001 wy-
st¹pi³y one w kwietniu, a w roku 1999 – w czerw-
cu. Od momentu wyst¹pienia wiosennego mak-
simum poziom wód zaczyna opadaæ i najni¿sze
poziomy notowane s¹ jesieni¹. W latach 1999
i 2000 wyst¹pi³y one pod koniec paŸdziernika i na
pocz¹tku listopada, natomiast w roku 2001 ju¿
od koñca sierpnia notowany by³ ponowny wzrost
poziomu wody.
W badanym okresie amplituda œredniego po-
ziomu wody dla ca³ego Parku wynios³a 74 cm,
ale roczne amplitudy s¹ ni¿sze i wynios³y od 30
cm w roku 2001 do 50 cm w roku 1999.
Badania warunków hydrogeologicznych, pro-
wadzone od wielu lat na terenie KPN przez Kazi-
mierskiego i Sikorsk¹-Maykowsk¹ (1979), pozwo-
li³y na wydzielenie stref o podobnej dynamice wód
podziemnych. Jako kryteria wydzieleñ, a jedno-
czeœnie czynniki powoduj¹ce odrêbnoœæ hydrody-
namiczn¹, oprócz procesów zasilania czy drena-
¿u, przyjêto: ró¿nice w budowie geologicznej
i geomorfologii, litologiê osadów przypowierzch-
niowych i zwi¹zane z tym pokrycie szat¹ roœlinn¹,
g³êbokoœæ zwierciad³a wód podziemnych, ampli-
tudê zmian stanów wód oraz gospodarcz¹ dzia³al-
noœæ cz³owieka. Wydzielono nastêpuj¹ce strefy:
A – pasy wydmowe, B – pasy dolinne (bagienne),
C – tarasy zalewowe i nadzalewowe, D – obszar
zasiêgu oddzia³ywania ujêcia wód podziemnych
w Wólce Smolanej, E - przedmieœcia Warszawy
oraz F – poziom b³oñski.
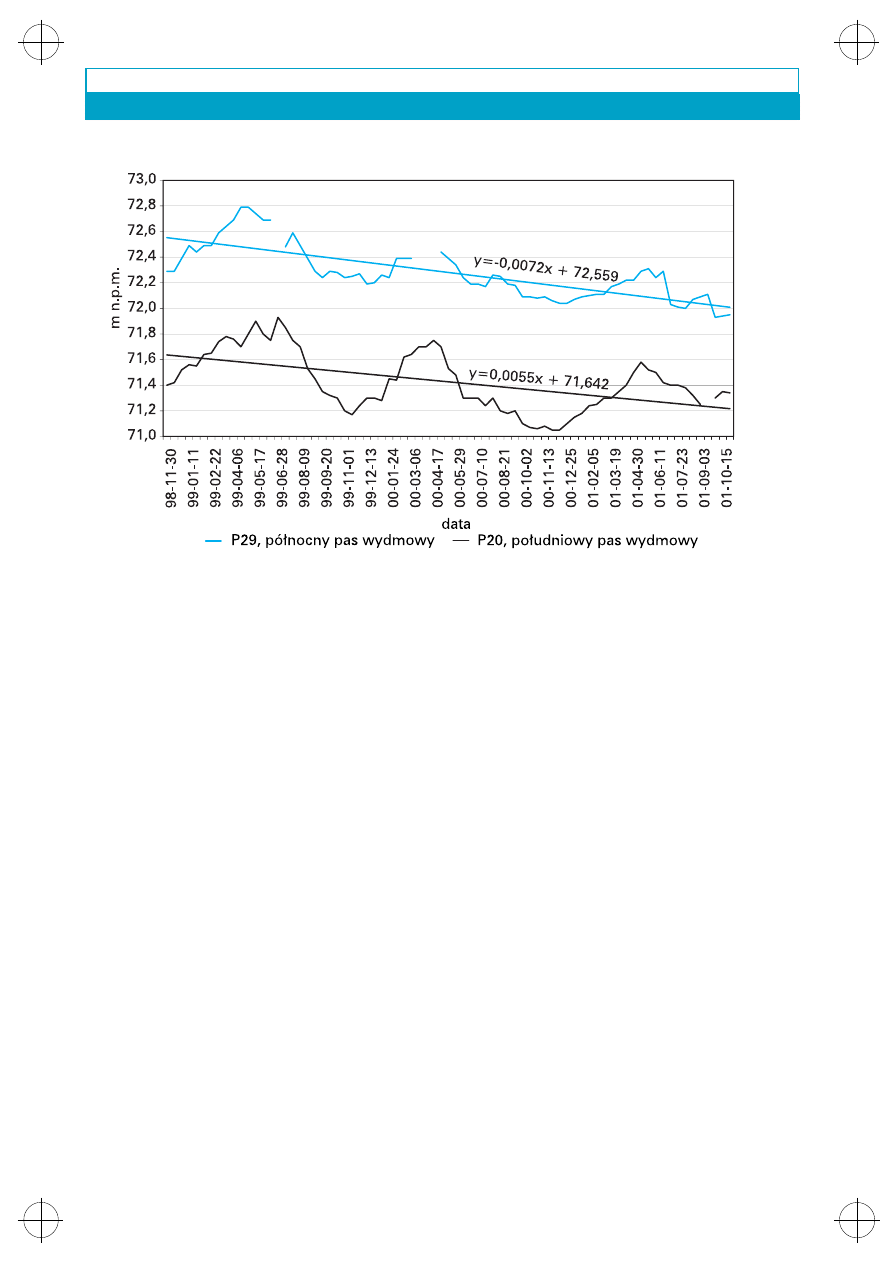
Pasy wydmowe s¹ obszarami intensywnej in-
filtracji opadów atmosferycznych. Zwierciad³o
wody podziemnej w okresie prowadzenia badañ
monitoringowych (listopad 1999 – grudzieñ
2001) stabilizowa³o siê na niewielkiej g³êboko-
œci, œrednio od 1,3 do 4,49 m (p.p.t.) w pó³noc-
nym pasie wydmowym i od 1,27 do 4,38 m p.p.t.
w po³udniowym pasie wydmowym; na rzêdnych
od 68,04 do 73,44 m n.p.m. w pó³nocnym pasie
wydmowym i od 68,60 do 77,88 m n.p.m. w po-
³udniowym pasie wydmowym
(rys. 4)
.
Zaznacza
siê tu wyraŸna tendencja wystêpowania wy¿szych
rzêdnych zwierciad³a wody podziemnej w po³u-
dniowym, wy¿szym pasie wydmowym ni¿ w pa-
sie pó³nocnym (Krogulec w druku).
72
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
Rys. 3
73
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
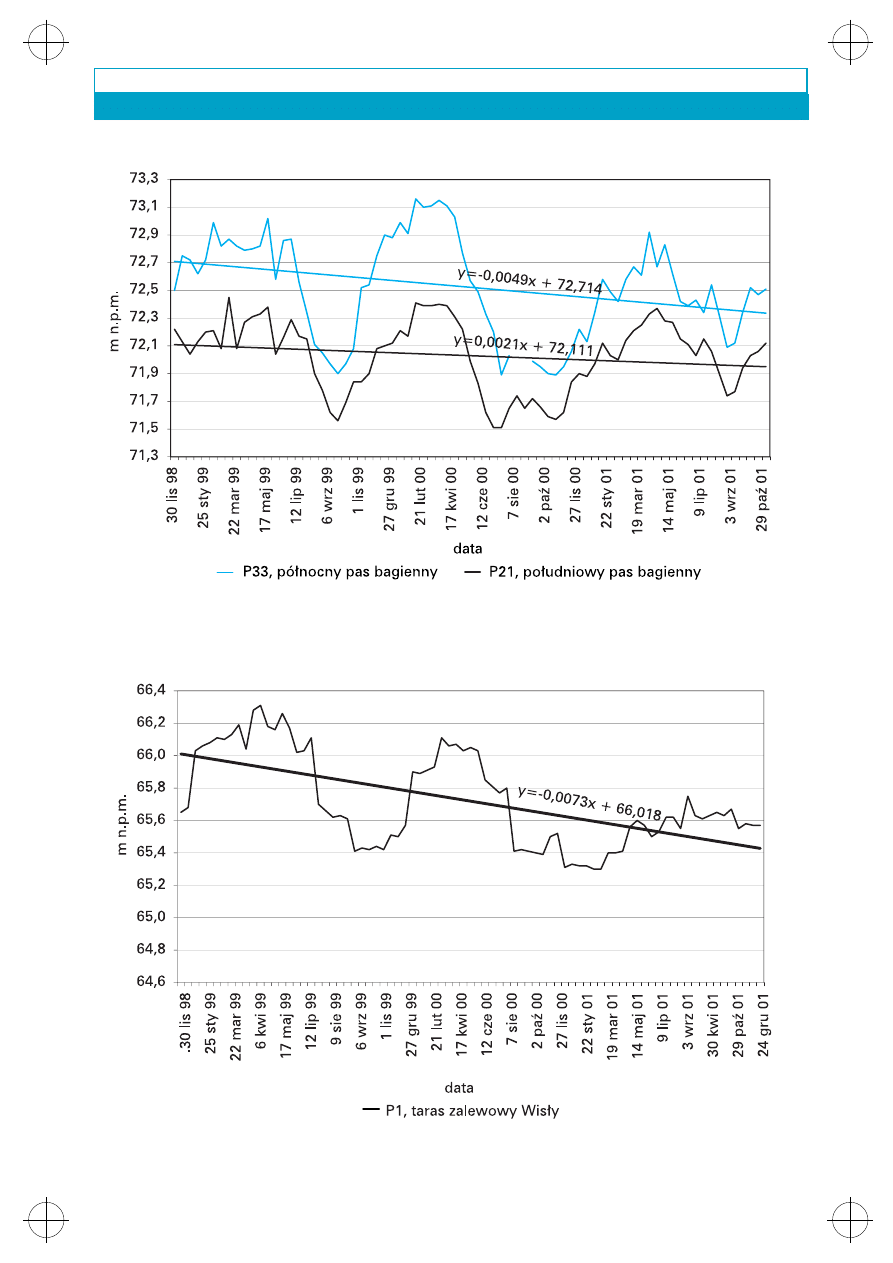
W rejonie pasów dolinnych (bagiennych) od-
bywa siê intensywny drena¿ wód podziemnych.
Zwierciad³o wody podziemnej wystêpuje w tej
strefie p³ytko, œrednio na rzêdnych od 68,45 do
73,93 m n.p.m. w pó³nocnym pasie bagiennym
i od 69,05 do 81,81 m n.p.m. w po³udniowym
pasie bagiennym
(rys. 5).
Stany wody i zakres
zmian zwi¹zane s¹ tutaj przede wszystkim z trze-
ma czynnikami: wielkoœci¹ opadu, procesami
ewapotranspiracji oraz drenuj¹cym charakterem
cieków (Krogulec w druku).
Na obszarze tarasu zalewowego s¹ notowane
znaczne amplitudy wahañ stanów wód podziem-
nych, warunkowane stanami wód w Wiœle (rys.
6).
W okresie prowadzenia obserwacji w punk-
tach monitoringowych KPN (od listopada 1998
do grudnia 2001) notowane by³y najczêœciej kil-
kudziesiêciocentymetrowe zmiany po³o¿enia
zwierciad³a wody podziemnej w ci¹gu roku (Kro-
gulec w druku).
Rejon oddzia³ywania ujêcia wód podziemnych
– tarasy Bzury (rejon wsi Wólka Smolana) wraz
z ocen¹ skutków wp³ywu eksploatacji na tereny
s¹siednie by³ przedmiotem licznych opracowañ.
Kompleksow¹ dokumentacjê zmian stanów wód
podziemnych, jak równie¿ wielkoœci i zasiêgu
zdepresjonowania wskutek eksploatacji ujêcia
przeprowadzono na modelu numerycznym (B.
Kazimierski, E. Pilichowska-Kazimierska 1993)
oraz w trakcie wieloletnich badañ hydrogeolo-
gicznych w tym rejonie. Obliczenia numeryczne
filtracji w warunkach niezaburzonych eksplo-
atacj¹ wskazuj¹ na drenuj¹cy charakter cieków:
Bzury, £asicy, Kana³u Olszowieckiego oraz nie-
wielki dop³yw od strony poziomu b³oñskiego.
Z porównania dwóch wariantów symulacji: wa-
runków hydrogeologicznych z rzeczywist¹ eks-
ploatacj¹ ujêcia, wynosz¹c¹ 912 m
3
/d, oraz wa-
runków hydrogeologicznych bez eksploatacji
(tzw. warunki naturalne) wynika, ¿e wskutek eks-
ploatacji wody na ujêciu w Wólce Smolanej,
z Bzury i Kana³u Olszowieckiego zaczê³y dop³y-
waæ znaczne iloœci wody do warstwy wodono-
œnej przy jednoczesnym zmniejszonym drena¿u
tych cieków. Pod wp³ywem eksploatacji i zdre-
nowania terenu wielkoϾ zasilania tarasu kampi-
Rys. 4. Zmiany poziomu wody w wybranych piezometrach pó³nocnego i po³udniowego pasa
wydmowego
74
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
Rys. 5. Zmiany poziomu wody w wybranym piezometrze pó³nocnego i po³udniowego pasa
bagiennego
Rys. 6. Zmiany poziomu lustra wody w wybranym piezometrze tarasu zalewowego Wis³y
75
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
noskiego od strony poziomu b³oñskiego w rejo-
nie zdrenowanym eksploatacj¹, obejmuj¹cym
strefê krawêdziow¹ o d³ugoœci oko³o 7000 m,
wynosi ponad 0,58 m
3
/d na 1 m d³ugoœci skarpy
(Krogulec w druku).
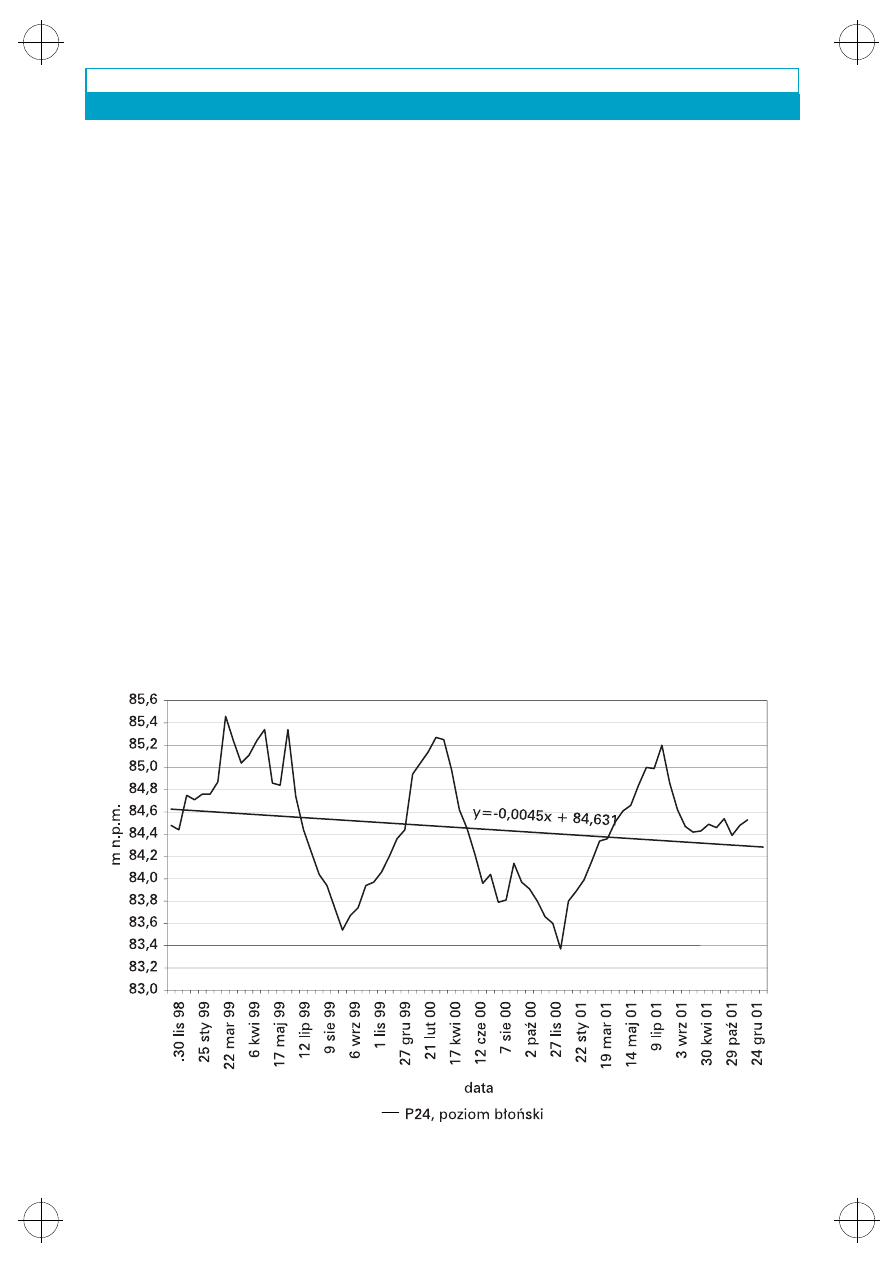
Na poziomie b³oñskim wystêpuj¹ dwie war-
stwy wodonoœne: przypowierzchniowa i g³êbiej
po³o¿ona – u¿ytkowa warstwa wodonoœna. Ba-
dania wykaza³y (Krogulec 1997), ¿e zasilanie
warstwy wodonoœnej tarasu kampinoskiego
w strefie przykrawêdziowej poziomu b³oñskiego
zwi¹zane jest z dop³ywem wód podziemnych tyl-
ko z g³êbszej warstwy wodonoœnej poziomu b³oñ-
skiego. Zwierciad³o wody podziemnej o charak-
terze swobodnym w rejonie kontaktu z tarasem
kampinoskim (rys. 7) kszta³tuje siê na rzêdnych
od 82,97 do 91,83 m n.p.m. (Krogulec w druku).
Po³udniowo-wschodnia czêœæ KPN i jego otu-
liny (przedmieœcia Warszawy) to rejon tzw. Wiel-
kiej Warszawy, g³ównie obszar zwartego budow-
nictwa jedno- i wielorodzinnego oraz doϾ
licznych zak³adów pracy. Warunki hydrogeolo-
giczne przedmieœæ Warszawy w rejonie KPN s¹,
pomimo doϾ dobrego rozpoznania, trudne do
scharakteryzowania. Strefa ta zosta³a wyznaczo-
na ze wzglêdu na znaczny stopieñ zagospodaro-
wania terenu. Pod wzglêdem geomorfologicznym
i geologicznym wystêpuje tu du¿a zmiennoœæ po-
woduj¹ca tak¿e znaczne zró¿nicowanie warun-
ków hydrogeologicznych (Krogulec w druku).
Podsumowanie
Przedstawiony monitoring wód powierzchnio-
wych i podziemnych na terenie Rezerwatu Bios-
fery „Puszcza Kampinoska” jest uzupe³niany
automatycznymi pomiarami w wybranych 4 pie-
zometrach, w których odczyty nastêpuj¹ w inter-
wa³ach godzinowych. Pomiary te umo¿liwiaj¹
analizê zmian stanu wód w okresach pomiêdzy
pomiarami manualnymi (Andrzejewska, Hardej
w druku).
Ponadto szczegó³owy monitoring podstawo-
wych elementów œrodowiska przyrodniczego pro-
wadzony jest w Stacji Bazowej Zintegrowanego
Rys. 7. Zmiany lustra wody w wybranym piezometrze poziomu b³oñskiego

76
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
Monitoringu Œrodowiska Przyrodniczego, gdzie
na obszarze zlewni Po¿ary dokonuje siê codzien-
nych pomiarów stanów wód w 21 piezometrach,
z których 3 to tzw. stanowiska „multilevel”, uj-
muj¹ce wody gruntowe z ró¿nych poziomów
(Wierzbicki 1997).
Monitoring wód s³u¿y nie tylko do badañ hy-
drologicznych i hydrogeologicznych, ale jest tak-
¿e podstaw¹ do badañ interdyscyplinarnych. Ba-
danie te s¹ u³atwione dziêki stosunkowo dobremu
wyodrêbnieniu zlewni £asicy, w znacznym stop-
niu pokrywaj¹cej siê z powierzchni¹ Kampinoskie-
go Parku Narodowego. Na podstawie sieci moni-
toringu wód zosta³a za³o¿ona sieæ sta³ych
powierzchni monitoringu zmian roœlinnoœci. Na-
wi¹zuje te¿ do niej monitoring wybranych grup
zwierz¹t wskaŸnikowych.
Na podstawie dotychczasowych badañ stwier-
dzono, ¿e poziom wód gruntowych, a zw³asz-
cza stan najd³u¿ej utrzymuj¹cy siê w roku
(moda), stale siê obni¿a (Krogulec 2002). Mo¿-
na wyró¿niæ kilka naturalnych i antropogenicz-
nych przyczyn tego zjawiska:
– czynniki atmosferyczne, a przede wszystkim
niskie opady atmosferyczne,
– melioracje wodne o charakterze drena¿owym,
– obni¿aj¹cy siê poziom wód Wis³y i Bzury,
stanowi¹cych lokaln¹ bazê drena¿u,
– systemy obwa³owañ Wis³y i Bzury, które nie
pozwalaj¹ na dop³yw wód powodziowych na
obszar obni¿eñ bagiennych i uzupe³nianie w ten
sposób zasobów wodnych,
– pobór wód podziemnych z ujêæ zlokalizowa-
nych na terenie Warszawy oraz w Wólce Smola-
nej (okolice Sochaczewa),
– brak wodoci¹gów i pobór wód podziemnych
ze studni gospodarskich,
– zmiana u¿ytkowania obszarów KPN w kie-
runku zbiorowisk o wiêkszej ewapotranspiracji:
zalesianie pól, zarastanie ³¹k, przebudowa drze-
wostanów z sosnowych na mieszane i liœciaste.
Ca³oœæ prac zwi¹zanych z rozwojem monito-
ringu przyrody jest ukierunkowana na stworze-
nie modelu jej funkcjonowania i takim doborze
elementów monitoringu, które umo¿liwi¹ stwo-
rzenie tego modelu (Andrzejewski 2001).
W obliczu zmian globalnych oraz zwiêkszaj¹-
cej siê urbanizacji, monitoring ten ma znaczenie
nie tylko dla Kampinoskiego Parku Narodowe-
go. Mo¿e dostarczyæ danych, bêd¹cych ogólno-
krajowymi i miêdzynarodowymi informacjami
o przemianach przyrody pod wp³ywem zmian lo-
kalnych i urbanizacyjnych.
Literatura
Andrzejewska A., Hardej M. w druku. Elec-
tronic measurements of groundwater level in the
Kampinoski National Park. International Journal
of Ecohydrology & Hydrobiology.
Andrzejewski R. 2001. Park narodowy jako
obszar monitoringu przyrody. Biuletyn Monito-
ringu Przyrody 1(2) Biblioteka Monitoringu Œro-
dowiska. Warszawa pp 66 –72.
Fic M., Wierzbicki A. 1994. Organizacja mo-
nitoringu wód podziemnych na terenie rezerwa-
tu Po¿ary w Kampinoskim Parku Narodowym.
Prz. Geol., vol. 42, nr 12, 1994.
Kondracki J. 1981. Geografia Fizyczna Polski
PWN. Warszawa
Kazimierski B. 1980. Ocena parametrów cha-
rakteryzuj¹cych zwi¹zki hydrauliczne wód pod-
ziemnych z wodami w rzece w œwietle analizy
modelowej. Archiwum. IHiGI, Warszawa.
Kazimierski B. i In. 1990. Warunki hydroge-
ologiczne Puszczy Kampinoskiej – w œwietle ob-
serwacji z lat 1985 –90, niepubl., Archiwum KPN.
Kazimierski B., Sikorska-Maykowska M., Pi-
lichowska-Kazimierska E. 1996. Operat ochro-
ny zasobów i ekosystemów wodnych. [w:] Plan
Ochrony KPN. NFOΠniepubl. Archiwum KPN
Krogulec E. 1997. Numeryczna analiza struk-
tury strumienia filtracji w strefie krawêdziowej
poziomu b³oñskiego (Kotlina Warszawska).
Wydawnictwo Uniwersytetu Warszawskiego,
Warszawa pp. 79.
Krogulec E. w druku. Hydrogeological condi-
tions of the Kampinos National Park (KNP) re-
gion.

77
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
Krogulec E., Sikorska-Maykowska M. 1996
Projekt lokalnej sieci monitoringu wód po-
wierzchniowych i podziemnych na obszarze KPN
wraz z otulin¹. niepubl. Archiwum KPN.
Sikorska-Maykowska M. 1981. Modelowanie
matematyczne zmiennoœci zasilania efektywne-
go warstwy wodonoœnej na podstawie badañ
w wid³ach Wis³y i Bzury, niepubl. Archiwum
Wydzia³u Geol. UW.
Sikorska-Maykowska M., Kazimierski B.
1989. Warunki hydrogeologiczne tarasu kampi-
noskiego a potrzeby wodne siedlisk roœlinnych
Puszczy Kampinoskiej [w:] Wspó³czesne proble-
my geologii Polski centralnej, Wyd. Zak³ad Ma-
³ej Poligrafi UW, Warszawa
Sikorska Maykowska M., Krogulec E., Fic M.
1998. Dokumentacja hydrologiczna z wykona-
nych piezometrów wraz z programem monitorin-
gu na obszarze Kampinoskiego Parku Narodo-
wego. niepubl. Archiwum KPN.
Wierzbicki A. 1999. System pomiarowy Zin-
tegrowanego Monitoringu Œrodowiska Przyrod-
niczego na terenie Stacji Bazowej „Po¿ary”
w Kampinoskim Parku Narodowym [w:] Zinte-
growany Monitoring Œrodowiska Przyrodnicze-
go Stacja Bazowa Po¿ary w Kampinoskim Par-
ku Narodowym, Inspekcja Ochrony Œrodowiska
Biblioteka Monitoringu, Warszawa, 85–12.
Roman Andrzejewski
Dzia³ Nauki i Monitoringu Przyrody
Kampinoski Park Narodowy
MONITOROWANIE DROBNYCH
SSAKÓW
zanym z drobnymi ssakami jest ocena ich zagêsz-
czenia, zwykle trudna do wykonania.
By prze¿yæ w przyrodzie, drobne ssaki musz¹
intensywnie siê rozmna¿aæ. Samica rodzi czêsto
i du¿o potomstwa, które szybko dojrzewa p³cio-
wo i rodzi kolejne pokolenie m³odych. U niektó-
rych gatunków drobnych ssaków, w naszym kli-
macie i sprzyjaj¹cych warunkach pokarmowych,
od jednej samicy, ¿yj¹cej na wiosnê, mo¿e przy-
byæ do jesieni razem dwieœcie jej dzieci, wnu-
ków i prawnuków, a nawet praprawnuków, mimo
¿e wiele z nich w miêdzyczasie zostanie zjedzo-
nych przez drapie¿niki lub padnie z powodu in-
fekcji. Mówi¹c jêzykiem bardziej precyzyjnym:
niektóre gatunki drobnych ssaków (np. nasz nor-
nik zwyczajny) mog¹ zwiêkszyæ zagêszczenie po-
pulacji od wiosny do jesieni 100 razy (po³owa
w tym jest samców, st¹d ka¿da samica musi uro-
dziæ dwa razy wiêcej potomstwa). Równoczeœnie
cech¹ drobnych ssaków jest krótkie ¿ycie, œred-
nio mniej ni¿ trzy miesi¹ce, a w naszym klimacie
u wiêkszoœci gatunków drobnych ssaków obserwuje
siê maksymaln¹ d³ugoœæ ¿ycia, trwaj¹c¹ od wiosny
jednego roku do jesieni nastêpnego. Jest to spowo-
dowane wielkim nara¿eniem na œmieræ ze strony
drapie¿ników, chorób paso¿ytniczych, z³ych warun-
ków atmosferycznych i pokarmowych.
W efekcie takiej biologii zagêszczenie drob-
nych ssaków mo¿e wykazywaæ du¿e wahania.
Je¿eli w danym roku nagromadzi siê dzia³anie
czynników sprzyjaj¹cych rozrodowi i prze¿yciu
m³odych, a bêdzie niewielkie dzia³anie czynni-
ków powoduj¹cych œmiertelnoœæ, zagêszczenie
danego gatunku lub gatunków o podobnych wy-
maganiach œrodowiskowych (ekologowie mówi¹:
„o podobnej niszy ekologicznej”) mo¿e byæ bar-
dzo du¿e. Je¿eli dzia³anie czynników u³o¿y siê
odwrotnie, to zagêszczenie ró¿nych gatunków
mo¿e byæ bardzo niskie, a w niektórych œrodo-
wiskach drobne ssaki ca³kowicie zanikn¹.
Badacze drobnych ssaków zaobserwowali ty-
powy obraz zmian zagêszczenia tych zwierz¹t.
Po roku o bardzo wysokim zagêszczeniu danego
gatunku nastêpuje rok o bardzo niskim zagêsz-
„Drobne ssaki” to okreœlenie przyjête dla naj-
mniejszych ssaków, wystêpuj¹cych w naszej fau-
nie: ryjówkowatych i kretów, myszowatych i nor-
nikowatych, pilchowatych. W ró¿nych krajach
i kontynentach t¹ nazw¹ obejmuje siê tak¿e inne
grupy systematyczne ssaków, ale zawsze o nie-
wielkich rozmiarach cia³a i masie. Podstawowym
zagadnieniem badawczym i praktycznym zwi¹-

78
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
czeniu, a nastêpnie, w kolejnych latach, zagêsz-
czenie roœnie a¿ do roku o zagêszczeniu znowu
bardzo wysokim. Taka sytuacja zwykle powta-
rza siê cyklicznie – ekologowie mówi¹ o cyklach
zmian liczebnoœci wielu gatunków drobnych ssa-
ków, ze szczytami liczebnoœci powtarzaj¹cymi siê
co oko³o 4 lata.
Od prawie 100 lat badacze obserwuj¹ na œwie-
cie cykle liczebnoœci drobnych ssaków i trwa
dyskusja, czy s¹ to naprawdê regularne cykle, a je-
¿eli tak, to co jest ich przyczyn¹. Powsta³o kilka
teorii na temat tych przyczyn i ¿adna, jak dot¹d,
dobrze nie t³umaczy obserwowanego zjawiska.
Istotn¹ jest jednak obserwacja, ¿e du¿e zmiany
liczebnoœci wystêpuj¹ u gatunków ¿yj¹cych
w œrodowiskach ubogich (np. lemingi w tundrze)
lub silnie zmienionych przez cz³owieka (np. nor-
nik zwyczajny na obszarach rolniczych), nato-
miast zmiany znacznie mniejsze s¹ w œrodowi-
skach o du¿ej ró¿norodnoœci biologicznej, gdzie
wystêpuje silniejsza regulacja ekologiczna za-
gêszczenia gatunków (np. w lasach).
Wysokie zagêszczenia drobnych ssaków po-
woduj¹ niszczenie upraw polowych w równych
czêœciach œwiata (tak¿e czasem i w Polsce). W ta-
kich przypadkach powstaj¹ znaczne straty eko-
nomiczne – wiele gatunków drobnych gryzoni
zaliczanych jest do powa¿nych szkodników
w rolnictwie. W warunkach œrodowisk bogatych,
np. leœnych, rzadko dochodzi do tak du¿ych za-
gêszczeñ, by powsta³o naruszenie równowagi eko-
logicznej ekosystemów, a drobnym ssakom przy-
pisuje siê rol¹ wa¿nego czynnika uczestnicz¹cego
w równowadze przyrody. Zarówno zbyt du¿e za-
gêszczenia, jak i odwrotnie, brak drobnych ssa-
ków wskazuj¹, ¿e ekosystemy o takiej sytuacji maj¹
zubo¿on¹ ró¿norodnoœæ biologiczn¹, zwykle z po-
wodu antropogennych przekszta³ceñ. St¹d pocho-
dzi zainteresowanie nauki i gospodarki rolnej lub
leœnej zagêszczeniami drobnych ssaków w ró¿nych
ekosystemach, jak czêsto wystêpuj¹ zagêszczenia
skrajne, i jakie s¹ ich przyczyny.
Drobne ssaki maj¹ jeszcze jedn¹ ciekaw¹
i wa¿n¹ cechê. Jest ni¹ ³atwoœæ przemieszczania
siê na stosunkowo du¿e odleg³oœci; opuszczania
siedlisk, w których warunki dla ich ¿ycia pogor-
szy³y siê, i zasiedlania siedlisk, w których ich nie
by³o, a w których warunki sta³y siê dla nich przy-
chylne.
Zdolnoœæ do szybkiego rozmna¿ania siê, wy-
mierania, opuszczania jednych siedlisk i ekosys-
temów, a zasiedlania innych powoduje, ¿e drob-
ne ssaki traktuje siê jako dobre bioindykatory
(wskaŸniki) zmian zachodz¹cych w œrodowisku.
Sk³ad gatunkowy i zagêszczenie poszczególnych
gatunków pozwalaj¹ oceniæ wskaŸnik ró¿norod-
noœci biologicznej tej grupy zwierz¹t i odnieœæ
go do ca³ego œrodowiska.
Cechy biologii, ekologii (zmiennoœæ zagêsz-
czeñ) i bioindykacyjnoœæ drobnych ssaków spra-
wiaj¹, ¿e warto je obj¹æ monitoringiem przyrod-
niczym.
S poœród polskich parków narodowych dobre
dane, dotycz¹ce wielu lat monitorowania zmian
zagêszczenia drobnych ssaków, ma Bia³owieski
Park Narodowy. Dane te zbiera i opracowuje
Zak³ad Badania Ssaków PAN
1)
.
W Kampinoskim Parku Narodowym od 50 lat
by³y prowadzone badania nad drobnymi ssaka-
mi, g³ównie przez Instytut Ekologii PAN w Dzie-
kanowie Leœnym. Choæ niektóre serie badawcze
obejmowa³y paroletnie odcinki czasu, to jednak
na podstawie tych obserwacji nie da siê odtwo-
rzyæ zmian liczebnoœci drobnych ssaków w tym
ca³ym okresie. Inne parki narodowe nie maj¹ in-
formacji o drobnych ssakach, które mo¿na by
uznaæ za d³ugookresowy monitoring zmian ich
liczebnoœci.
W roku 1992 rozpoczêliœmy oceny zmian za-
gêszczenia drobnych ssaków w Kampinoskim
Parku Narodowym, poszukuj¹c sta³ej metody,
przystosowanej do „parkowych warunków” przy-
1)
Pucek Z., Jêdrzejewski W., Jêdrzejewska B., Pucek
M. 1993. Rodent population dynamics in primeval deci-
duous forest (Bia³owie¿a National Park) in relation to
weather, seed crop and predation. Acta Therilogica 38 (2):
199–232.

79
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
rodniczych i organizacyjnych. Wiedz¹c, ¿e nie
ma metod oceny zagêszczenia drobnych ssaków,
daj¹cych rzeczywisty obraz, przyjêliœmy nastê-
puj¹ce za³o¿enia wstêpne.
Metoda powinna dawaæ ocenê wzglêdn¹, po-
równywaln¹ w czasie i przestrzeni, nie sil¹c siê
na podanie rzeczywistego zagêszczenia drobnych
ssaków, np. liczonego w liczbie osobników wy-
stêpuj¹cych na hektarze danego œrodowiska.
Metoda powinna byæ przy¿yciowa (nie uœmier-
caj¹ca zwierz¹t) chocia¿ musi dopuœciæ okreœlon¹
œmiertelnoœæ i umo¿liwiaæ wyjêcie z parku ogra-
niczonej liczby osobników, je¿eli te bêd¹ po-
trzebne do monitorowania innych zjawisk (np.
wystêpowania w populacjach ssaków mikroor-
ganizmów zagra¿aj¹cych zdrowiu cz³owieka lub
zwierz¹t hodowlanych).
Metoda powinna byæ ³atwa, ma³o czasoch³on-
na i niekosztowna, a co wa¿ne mo¿liwa do wy-
konania ewentualnie przez terenowe s³u¿by par-
ku.
Jeden z pracowników parku powinien byæ od-
powiedzialny za coroczne przeprowadzenie ocen
terenowych, analizê i podsumowanie wyników
oraz przedstawienie dyrektorowi parku odpo-
wiedniego sprawozdania.
W roku 2002 Dyrektor Kampinoskiego Parku
Narodowego podpisa³ ostateczn¹ decyzjê o sta-
³ym monitorowaniu drobnych ssaków. W decy-
zji tej przewidziano nastêpuj¹c¹ metodê i zasady
organizacyjne.
Drobne ssaki od³awia siê w 25 pu³apkach
¿ywo³ownych typu „dziekanowskiego”, usta-
wionych w pêtlê (pocz¹tek i koniec pêtli znaj-
duj¹ siê niedaleko od siebie ze wzglêdu na
oszczêdnoœæ czasu obs³ugi) na obszarze bada-
nego œrodowiska
2)
.
Pu³apki w linii tworz¹cej pêtlê stoj¹ w odle-
g³oœci oko³o 10 m od siebie (linia ma wiêc 250
m) przez 4 doby (100 pu³apkodób). Pu³apki usta-
wia siê w poniedzia³ek rano, kontroluje, opisuje,
znakuje, wypuszcza z³owione zwierzêta raz
dziennie (w godzinach rannych) do pi¹tku, gdy
wraz z kontrol¹ pu³apek nastêpuje ich zebranie.
Przed uwolnieniem zwierz¹t znakuje siê je dla
wyró¿nienia z³owieñ powtórnych, obcinaj¹c koñ-
ce w³osów w postaci plamki na sierœci. Termi-
nem przeprowadzenia corocznej kontroli jest
okres miêdzy 1 i 15 paŸdziernika (po zakoñcze-
niu sezonu rozrodczego, w szczycie zagêszcze-
nia drobnych ssaków w cyklu rocznym).
Odnotowuje siê nastêpuj¹ce dane z kontroli
pu³apek: (1) ogólne – data, numer pêtli pu³apek,
aktualne warunki œrodowiskowe, specyficzne
w stosunku do danych najbli¿szej stacji meteoro-
logicznej, których trzy posiada KPN, (2) indywi-
dualne dla ka¿dego z³owionego osobnika – nu-
mer stanowiska pu³apki, gatunek z³owionego
zwierzêcia, p³eæ, masê (okreœlon¹ na wadze sprê-
¿ynowej firmy Pesola), cechy kondycji p³ciowej.
Notuje siê tak¿e fakty: zamkniêcie pu³apki bez
zwierzêcia, widoczne zablokowanie pu³apki unie-
mo¿liwiaj¹ce z³owienie, inne z³owione zwierzê-
ta (poza ssakami), np. p³azy, gady, ptaki.
Podstawowym wskaŸnikiem wzglêdnego za-
gêszczenia drobnych ssaków jest liczba z³owio-
nych osobników (nie licz¹c z³owieñ powtórnych)
danego gatunku na 100 pu³apkodób, a dla oceny
ró¿norodnoœci biologicznej mo¿na zastosowaæ
wskaŸnik Shannona-Wienera. Istnieje mo¿liwoœæ
wykonania bardziej skomplikowanych analiz ze-
branego materia³u.
W przypadku gdy potrzebne s¹ ¿ywe zwierzê-
ta do innych badañ, mog¹ w tym celu byæ one
zabrane tylko w pi¹tek, w czasie ostatniej kon-
troli pu³apek. Zwierzêta pad³e w pu³apkach mog¹
byæ wykorzystane do innych badañ lub preparo-
wane i gromadzone jako zbiory dokumentacyjne
(tak¿e dydaktyczne) parku. Te czynnoœci nie s¹
czêœci¹ monitoringu drobnych ssaków.
Na terenie Kampinoskiego PN wyznaczono 7
pêtli pomiarowych (miejsca pêtli s¹ trwale ozna-
2)
Kszta³t linii w postaci pêtli podyktowany jest tylko
oszczêdnoœci¹ czasu, traconego na powrót do miejsca po-
cz¹tkowego. Je¿eli kszta³t œrodowiska uniemo¿liwia za³o-
¿enie pêtli, pu³apki mog¹ tworzyæ liniê prost¹ lub o ³agod-
nych zakrêtach.

80
Biuletyn Monitoringu Przyrody – Numer 1/2003 (4)
ARTYKU£Y I KOMUNIKATY
kowane w terenie) w trzech typach œrodowisk:
w ponadstuletnim borze i w olsie oraz na turzy-
cowisku (Macrocaricetum). Trzy takie pêtle (po-
³o¿one w zasiêgu kilometra miêdzy sob¹) znaj-
duj¹ siê w zachodniej i trzy we wschodniej czêœci
Parku. Siódma pêtla znajduje siê w turzycowi-
sku na terenie Stacji Zintegrowanego Monitorin-
gu Przyrody w zlewni Po¿ary. Tak rozmieszczo-
ne miejsca monitorowania drobnych ssaków daj¹
mo¿liwoœæ porównania miêdzy sob¹ zagêszcze-
nia zwierz¹t w trzech podstawowych dla Kampi-
noskiego PN œrodowiskach oraz miêdzy zachod-
ni¹ i wschodni¹ czêœci¹ parku (odleg³oœæ oko³o 40
km). Równoczeœnie istnieje mo¿liwoœæ zanalizo-
wania wyników po³owów na tle danych metero-
logicznych, zbieranych na stacjach mieszcz¹cych
siê w odleg³oœci do 3 km od pêtli, i w³¹czenia
danych z po³owów do wyników Stacji Monito-
ringu Zintegrowanego. W pobli¿u pêtli znajduj¹
siê tak¿e miejsca pomiaru zmiany g³êbokoœci
zwierciad³a wód gruntowych i powierzchnie mo-
nitorowania szaty roœlinnej.
Przeprowadzona wed³ug tej metody ocena li-
czebnoœci drobnych ssaków w roku 2002 wyka-
za³a we wszystkich œrodowiskach wyj¹tkowo
wysokie liczebnoœci gryzoni i umiarkowane li-
czebnoœci ryjówek. Metod¹ zbli¿on¹ do powy¿-
3)
Adamczewska-Andrzejewska K. 1998. Dynamika li-
czebnoœci i struktura zgrupowania drobnych ssaków w
zdegradowanych i niezdegradowanych œrodowiskach Kar-
konoszy. w: Geoekologiczne problemy Karkonoszy. Sa-
rosiek J., Stursa J. (red.). Wyd. Acarus, Poznañ, 2: 45–52.
4)
Adamczewska-Andrzejewska K. 2002. Operat „zoo-
log” Drobne ssaki. Manuscrypt materia³ów do Planu
Ochrony Parku Narodowego Bory Tucholskie.
szej stosowano dla oceny zagêszczenia drobnych
ssaków w ró¿nych œrodowiskach Karkonoszy,
w tym Karkonoskiego Parku Narodowego
(Adamczewska-Andrzejewska 1998)
3)
, i ostatnio
w Parku Narodowym Bory Tucholskie w zwi¹z-
ku z dokumentowaniem stanu fauny dla Planu
Ochrony tego Parku (Adamczewska-Andrzejew-
ska 2002)
4)
.
Na zakoñczenie nale¿y zrobiæ nastêpuj¹ce
uwagi. Metoda nie obejmuje drobnych ssaków
z rodziny pilchowatych, mo¿liwe tak¿e, ¿e nie bê-
dzie przydatna dla oceny karczownika ziemno-
wodnego (który w Kampinoskim PN jest gatun-
kiem bardzo rzadkim). Metoda pod wzglêdem
otrzymywanych danych jest zapewne prymityw-
na, jednak nie ma innej, która dawa³aby wiêksz¹
dok³adnoœæ oceny zagêszczenia. Walorem tej
metody jest niska praco- i kosztoch³onnoœæ.
Proponujemy sta³e stosowanie jej w innych
parkach narodowych. Wyniki bêd¹ porównywal-
ne w danym parku narodowym i miêdzy parka-
mi, co najmniej w odniesieniu do synchronicz-
noœci cykli populacyjnych ró¿nych gatunków
drobnych ssaków, na który to temat nie mamy
w Polsce odpowiednich danych. Chêtnie s³u¿y-
my pomoc¹ w organizacji monitoringu drobnych
ssaków.
Wyszukiwarka
Podobne podstrony:
PRZEKRA J TEOWY 2012 03 23 id 3 Nieznany
programowanie st7 2011 03 14 id Nieznany
Cennik INTERIA PL 2013 03 21 id Nieznany
cwicz e3 Kapanowski 8 03 13 id Nieznany
Cw inz Pn 2015 part 03 cw 4 id Nieznany
ei 2005 03 s024 id 154147 Nieznany
Edukacja spol przyr 5 kl 1 id Nieznany
Notatki 03 PRODUKT id 322319 Nieznany
ei 2005 03 s006 id 154146 Nieznany
cw 03 formularz id 121361 Nieznany
atmwp recenzja re 03 2006 id 71 Nieznany (2)
kolos automatyka cw PAiTS 03 id Nieznany
monitor 03 2011 id 307221 Nieznany
24 03 2011 id 30495 Nieznany (2)
23 03 2011 id 29948 Nieznany
3 03 2011 2 id 32823 Nieznany (2)
więcej podobnych podstron