
Czynność komórek
nerwowych i
mięśniowych

Pobudliwość i
pobudzenie
• Pobudzenie (excitation) jest to
zmiana właściwości błony
komórkowej lub metabolizmu
komórkowego pod wpływem
czynników działających z zewnątrz
komórki, czyli pod wpływem
bodźców.

• W warunkach fizjologicznych
bodźcami działającymi na
przeważającą liczbę komórek w
organizmie są przekaźniki chemiczne.
Komórki natomiast, tworzące w
organizmie narządy odbiorcze, czyli
receptory, odbierają w warunkach
fizjologicznych również bodźce
fizyczne w różnej postaci, np. fal
świetlnych, fal akustycznych, energii
cieplnej, energii mechanicznej (jako
ucisk lub rozciąganie itp.).

• Bodźce fizjologiczne są to takie
bodźce, które nie uszkadzają komórki
wywolują całkowicie odwracalne
procesy.

• Pobudliwość (excitability) jest to zdolność
reagowania na bodziec.
• do pobudliwych zalicza się te tkanki, których
komórki szybko odpowiadają na bodźce.
• Są to tkanki zbudowane z komórek
nerwowych i ich wypustek oraz z komórek
mięśniowych: mięśni poprzecznie
prążkowanych, mięśni gładkich i mięśnia
sercowego.

• W komórkach mięśniowych i
nerwowych pod wpływem bodźca
dochodzi do wędrówki jonów do
wnętrza komórki, a następnie z
wnętrza komórki na zewnątrz. Tym
szybkim wędrówkom przez błonę
komórkową jonów obdarzonych
ładunkiem elektrycznym towarzyszą
wahania potencjału elektrycznego.

Potencjał spoczynkowy
• Pomiędzy wnętrzem komórek tkanek
pobudliwych a płynem
zewnątrzkomórkowym występuje
stale w spoczynku różnica potencjału
elektrycznego, czyli potencjał
spoczynkowy błony komórkowej
(resting membrane potential).

Potencjał spoczynkowy
• Jest on spowodowany właściwościami
błony komórkowej, przez ktorą jony o
dodatnim ładunku elektrycznym
przenikają z trudnością.

• Stężenie poszczególnych jonów w
płynie wevrnątrzkomórkowym nie
zmienia się, jeśli metabolizm nie
ulegnie zmianie i jeśli na błonę
komórkową nie działają bodźce z
zewnątrz. W tych warunkach
wytwarza się równowaga pomiędzy
stężeniem poszczególnych jonów
na zewnątrz i wewnątrz komórek.

Pompa sodowo-potasowa
• Utrzymanie wewnątrz komórek
dużego stężenia K+ i małego
stężenia Na+ wymaga aktywnego
transportu obu tych kationów przez
błonę komórkową przeciwko
gradientowi stężeń.

• Napęd pompy sodowo-potasowej
wiąże się z metabolizmem
wewnątrzkomórkowym. Okolo
30% całego metabolizmu
komórkowego tkanek pobudliwych
jest zużywane na napęd pompy
sodowo-potasowej.

Optymalna praca pompy i związana z tym
optymalna pobudliwość wymagają:
• 1) stałego dopływu do komórek tlenu i
substancji energetycznych (glukozy);
• 2) stałej resyntezy ATP z ADP i fosforanu w
procesie oddychania komórkowego;
• 3) stałego odprowadzania z komórek
ostatecznego produktu rozpadu substancji
energetycznych - dwutlenku węgla;
• 4) odpowiedniego stosunku kationów
[Na+] do [K+] w płynie
zewnątrzkomórkowym;
• 5) odpowiedniej temperatury dla procesów
enzymatycznych wewnątrzkomórkowych,
jaką jest 37°C.

• Wystarczy zmiana jednego z wymienionych
warunków, aby nastąpiło zwolnienie lub
zatrzymanie pompy sodowo-potasowej.
• Po zatrzymaniu pompy następuje
wyrównanie stężenia Na+ i K+ po obu
stronach błony komórkowej i zanika różnica
potencjałów elektrycznych pomiędzy
wnętrzem komórki a otoczeniem.

• Po wyrównaniu różnicy
potencjałów pomiędzy wnętrzem
komórki a jej otoczeniem wskutek
zatrzymania pompy komórki tkanek
pobudliwych tracą swoje
właściwości. Przestają reagować na
bodźce i stają się niepobudliwe.

Komórka nerwowa
• W organizmie człowieka znajduje się około 30
miliardów komórek nerwowych, czyli neuronów.
• Znaczna większość z nich skupia się w
ośrodkowym układzie nerwowym.
• Stosunkowo niewielka ich liczba występuje poza
ośrodkowym układem nerwowym w zwojach
nerwowych. Są to zwoje nerwowe nerwów
czaszkowych i nerwów rdzeniowych oraz zwoje
nerwowe należące do układu autonomicznego.

• Zasadniczą funkcją
neuronu jest
przekazywanie
informacji
zakodowanych w postaci
impulsów nerwowych.

• W organizmie człowieka występują
zarówno neurony o krótkich
aksonach rozgałęziających się w
pobliżu ciała komórki, jak i o długich
aksonach, z których najdłuższe mają
około 1,2 m.

• Jedne z największych komórek
nerwowych w organizmie człowieka, o
długich aksonach, występują w
jądrach ruchowych pnia mózgu i
rdzenia kręgowego.
• Aksony tych komórek biegną w
nerwach czaszkowych lub nerwach
rdzeniowych do mięsni szkieletowych,
gdzie znajdują się ich zakończenia.

Odmienną budowę mają
neurony czuciowe w zwojach
rdzeniowych
• Jedna ich wypustka, długa, podobna do
aksonu, odbiera pobudzenie z
receptorów i przewodzi je w postaci
impulsów nerwowych do ciała
komórkowego.
• Za pośrednictwem zaś krótszej wypustki,
wstępującej do rdzenia kręgowego,
neurony przekazują do innych komórek
nerwowych impulsy nerwowe.

Potencjał czynnościowy
• Bodziec działając na błonę
komórkową neuronu zmienia jej
właściwości, co z kolei wywołuje
potencjał czynnościowy.

Depolaryzacja błony
komórkowej
• Zjawisko napływania jonów Na+ do
wnętrza neuronu, co powoduje wyrównanie
ładunków elektrycznych pomiędzy
wnętrzem a otoczeniem.
– Jony Na+ początkowo wnikają do wnętrza
neuronu tylko w miejscach zadziałania bodźca.
Z chwilą wyrównania ładunków elektrycznych w
tym jednym miejscu depolaryzacja zaczyna się
rozszerzać na sąsiednie odcinki błony
komórkowej, przesuwając się również wzdłuż
aksonów.

Impuls nerwowy
• Impulsem nerwowym jest
przesuwanie się fali
depolaryzacji od miejsca
zadziałania bodźca na błonę
komórkową aż do zakończenia
neuronu.

• Miejsce stykania się ze sobą błony
komórkowej zakończenia aksonu z
błoną komórkową drugiej komórki
nosi nazwę synapsy.
• Błonę komórkową neuronu
przekazującego impuls przyjęto
nazywać błoną presynaptyczną,
błona komórkowa neuronu
odbierającego impuls nosi zaś nazwę
błony postsynaptycznej.

Postsynaptyczny potencjał
pobudzający
• Z zakończeń aksonów w obrębie
synaps wydzielają się przekaźniki
chemiczne, czyli transmittery, które
zmieniają właściwości błony
postsynaptycznej.

Postsynaptyczny potencjał
pobudzający c.d.
• Pod wpływem cząsteczek transmittera, który
pośredniczy w przewodzeniu przez synapsę
impulsów pobudzających, jony Na+ wnikają do
wnętrza neuronu odbierającego impuls nerwowy.
• Zmniejszenie się ujemnego potencjału
wewnątrzkomórkowego przyjęto nazywać
postsynaptycznym potencjałem pobudzającym
- EPSP (excitatory postsynaptic potential),
• synapsy wywołujące depolaryzację błony
komórkowej noszą zaś nazwę synaps
pobudzających.

Potencjał iglicowy
• Pod wpływem transmittera
wydzielającego się na synapsach
pobudzających depolaryzacja błony
komórkowej neuronu osiąga pewien
próg, przy którym dochodzi do
wyzwolenia potencjału iglicowego.
• Potencjał progowy czyli krytyczny
potencjał błony komórkowej ciała
neuronu.

• Po szybko
narastającej
depolaryzacji
zakończonej
nadstrzałem
następuje
repolaryzacja
błony
komórkowej,

Potencjał następczy
• Zjawisko zwalniania repolaryzacji
błony komórkowej neuronu po
powrocie ujemnego potencjału
wewnątrzkomórkowego i po
przekroczeniu potencjału progowego.

Postsynaptyczny potencjał
hamujący
• Transmitter hamujący zmienia właściwości
błony komórkowej w ten sposób, że jony K+
uciekają z wnętrza neuronu na zewnątrz i
jednocześnie jony Cl- wnikają do wnętrza.
• Ubywanie jonów o dodatnim ładunku
elektrycznym i przybywanie jonów o
ładunku ujemnym powoduje zwiększenie
ujemnego potencjału elektrycznego we
wnętrzu komórki.

Transmittery pobudzające
• Do transmitterów chemicznych
pobudzających zalicza się
acetylocholinę, aminy (dopamina,
adrenalina, noradrenalina, serotonina
i histamina) oraz aminokwasy (sole
kwasu asparaginowego i
glutaminowego).

Transmittery hamujące
• Zasadniczym transmitterem
hamującym jest kwas gamma-
aminomasłowy - GABA powstający
w wyniku dekarboksylacji
aminokwasu - kwasu glutaminowego.

Modulatory synaptyczne
• poza transmitterami z pęcherzyków
synaptycznych uwalniają się również
związki o większej cząsteczce,
którym przypisuje się rolę
modulatorów synaptycznych.

Modulatory synaptyczne
• Do związków tych zalicza się biologicznie
aktywne peptydy. (wazopresyna
argininowa, neurotensyna, cholecystokinina)
• Modulatory synaptyczne uwolnione z
zakonczeń nerwowych oddziałują na błonę
postsynaptyczną i presynaptyczną.
• Aktywują lub inaktywują enzymy
występujące w tych błonach, tym samym
zmieniają właściwości błon komórkowych,
wzmacniając lub tłumiąc działanie
transmitterów.

Hamowanie
presynaptyczne
W zależności od lokalizacji synaps na błonie
postsynaptycznej neuronu odbierającego dzielą
się one na trzy zasadnicze rodzaje:
1)
dendrytów, zwane synapsą aksono-
dendrytyczną,
2)
ciała neuronu, czyli synapsa aksono-
somatyczna,
3)
aksonów, zwane synapsą aksono-aksonalną.

• Neurony kończące się synapsami
pobudzającymi mogą również
hamować przekazywanie impulsów
przez inne neurony pobudzające w
wyniku hamowania
presynaptycznego.
• Ten typ hamowania wywołują synapsy
aksono-aksonalne.

Błona komórkowa ciała neuronu i
odcinka początkowego aksonu:
I w spoczynku,
II - w czasie depolaryzacji ciała
neuronu,
III - w czasie przewodzenia
depolaryzacji wzdłuż aksonu bez
osłonki mielinowej.
Przewodzenie impulsów we
włóknach bezrdzennych.

Błona komórkowa ciała neuronu i
odcinka początkowego aksonu:
I w spoczynku,
II - w czasie depolaryzacji ciała
neuronu,
III - w czasie przewodzenia
skokowego depolaryzacji wzdłuż
aksonu z osłonką mielinową.
Przewodzenie impulsów
we włóknach rdzennych.

Grupy włókien nerwowych
• Włókna nerwowe dzieli się zarówno
pod względem morfologicznym, jak i
czynnościowym.

Morfologiczne kryteria
podziału
odnoszą się do:
• obecności lub braku osłonki
mielinowej,
• średnicy aksonów,
• miejsca występowania:
– zarówno w ośrodkowym układzie
nerwowym, gdzie tworzą drogi nerwowe,
jak i obwodowym układzie nerwowym,
gdzie są skupione w postaci nerwów.

Czynnościowe kryteria
podziału
pod względem czynnościowym
włókna nerwowe dzielą się na
przewodzące impulsy:
• z obwodu do ośrodków - są to włókna
dośrodkowe, czyli aferentne,
• z ośrodków na obwód - włókna
odśrodkowe, czyli eferentne,

Mięśnie poprzecznie
prążkowane
Komórka mięśniowa:
Miesień poprzecznie
prążkowany, czyli
szkieletowy, jest
zbudowany z wielu
tysięcy komórek
mięśniowych
tworzących pęczki.

• Włókienko mięśniowe, czyli
miofibryla, ma odcinki o większym i
mniejszym współczynniku załamania
światła występujące naprzemiennie.
• Odcinki silniej załamujące światło
tworzą ciemniejsze prążki zwane
prążkami anizotropowymi (A),
odcinki słabiej załamujące światło
tworzą zaś jasne prążki izotropowe
(I),
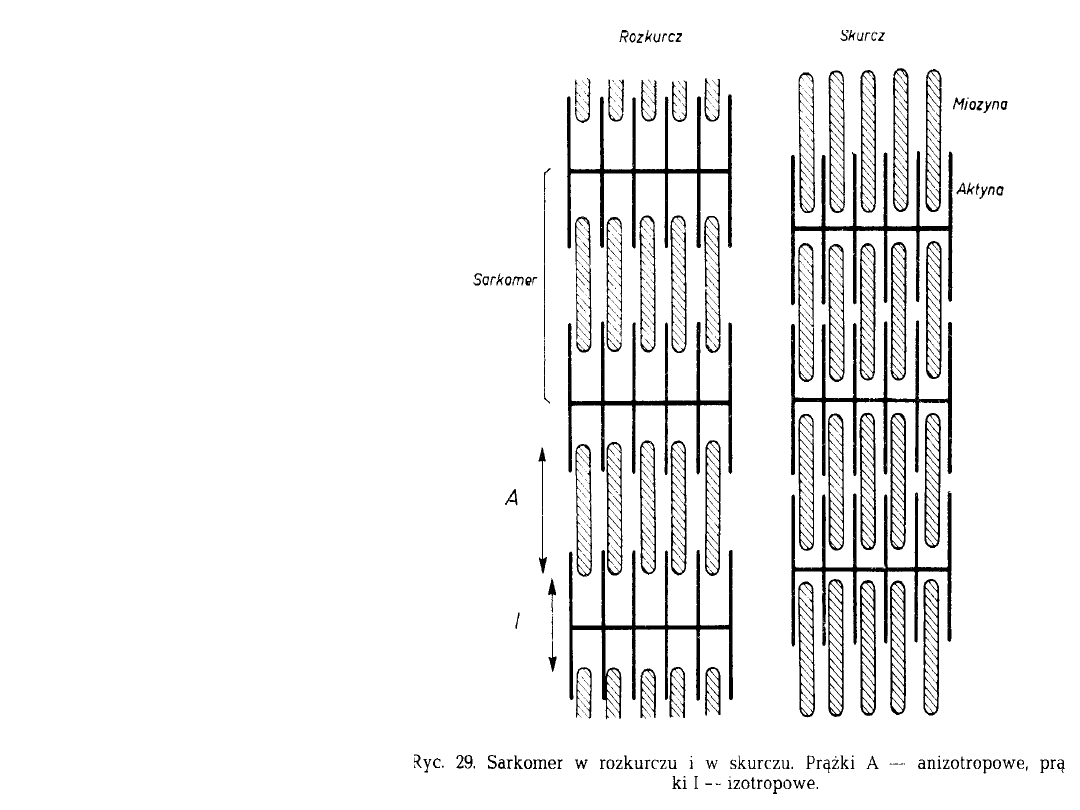
Sarkomer
Sarkomer obejmuje
jeden cały prążek
anizotropowy i
sąsiadujące z nim dwie
połówki prążka
izotropowego.

• Prążek anizotropowy tworzą nitki
grube miozyny, prążek izotropowy
zaś nitki cienkie aktyny, które są
doczepione do błony granicznej Z.
• Błona Z dzieli każdy prążek
izotropowy na dwie połówki, należące
do dwóch sąsiednich sarkomerów.

• W czasie skracania się komórki
mięśniowej nitki aktyny tworzące
prążki izotropowe wsuwają się
pomiędzy nitki miozyny i dzięki
temu wszystkie prążki I nikną.
• W rozkurczu nitki aktyny wysuwają
się spomiędzy nitek miozyny i prążki
I ponownie stają się widoczne.

Skurcze mięśni
• Pod wpływem działającego na komórkę
mięśniową pojedynczego bodźca o sile
progowej lub wyższej od progowej jego
błona komórkowa ulega depolaryzacji, po
której następuje skurcz całej komórki.
• Bodziec podprogowy me wywołuje
depolaryzacji błony komórkowej i komórka
mięśniowa nie kurczy się.
• Komórka mięśniowa odpowiada na bodziec
zgodnie z prawem „wszystko albo nic”,

Wyróżnia się dwa rodzaje
pojedynczych skurczów mięśni
szkieletowych
• izotoniczne
• (W czasie skurczu izotonicznego komórki
mięśniowe skracają się i cały miesień ulega
skróceniu, jego napięcie zaś nie zmienia się.
Przyczepy mięśnia w układzie szkieletowym
zbliżają się do siebie).
• izometryczne
• (Skurcz izometryczny charakteryzuje się
wzrostem napięcia mięśnia bez zmian jego
długości. Przyczepy mięśnia w układzie
szkieletowym nie zmieniają swojej
odległości).

• Powtarzające się z pewną
częstotliwością bodźce nadprogowe
wywołują kolejne skurcze mięśnia.
• Przy pewnej częstotliwości bodźca
zaczyna występować zjawisko
sumowania się skurczów
pojedynczych.
• Dochodzi do skurczu tężcowego
zupełnego lub do skurczu
tężcowego niezupełnego.

• Skurcz tężcowy zupełny występuje
wtedy, kiedy bodźce pobudzają
miesień w odstępach czasu krótszych,
niż trwa skurcz pojedynczy.
• Pobudzanie mięśnia w odstępach
czasu dłuższych niż czas trwania
skurczu pojedynczego pozwala na
częściowy rozkurcz mięśnia pomiędzy
bodźcami.

• Pobudzenie wszystkich komórek
mięśniowych wywołuje skurcz
maksymalny.

Jedna komórka nerwowa, jej
wypustka biegnąca do mięśnia i
wszystkie komórki mięśniowe
przez nią unerwione stanowią
jednostkę motoryczną.

Siła skurczu mięśnia w
organizmie zależy od:
1. liczby jednostek motorycznych
biorących udział w skurczu,
2. częstotliwości, z jaką poszczególne
jednostki motoryczne są pobudzane,
3. stopnia rozciągnięcia mięśnia przed
jego skurczem.

Napięcie mięśniowe jest
regulowane:
• przez nadrzędne ośrodki ruchowe w
ośrodkowym układzie nerwowym.
2. dzięki samoregulacji (muscle tonus
regulation).

Samoregulacja napięcia
mięśniowego
• Rozciągnięcie mięśnia wyzwala
odruchowy wzrost jego napięcia.
• Na przykład skurcz izotoniczny zginaczy
powoduje jednoczesne rozciągnięcie mięśni
antagonistycznych, czyli prostowników, i
wzrost ich napięcia mięsniowego na drodze
odruchowej.

W mięśniach szkieletowych
występują dwa rodzaje
komórek mięśniowych:
• komórki ekstrafuzalne
• komórki intrafuzalne

Komórki mięśniowe
intrafuzalne
• w swej części środkowej nie mają poprzecznego
prążkowania i część ta nie kurczy się.
• są skupione w pęczki, czyli wrzecionka
nerwowo-mięśniowe, które otacza torebka
łącznotkankowa.
• wrzecionka nerwowo-mięśniowe przyczepiają
się swoimi końcami do komórek
ekstrafuzalnych. We wrzecionkach tych
znajdują się receptory wrażliwe na rozciąganie
mięśnia.

Komórki mięśniowe
ekstrafuzalne
• stanowią podstawową masę każdego mięśnia
szkieletowego,
• unerwione są przez duże neurony ruchowe,
zwane neuronami alfa.
• Komórki intrafuzalne unerwiają mniejsze
neurony ruchowe, zwane neuronami gamma.
• Pobudliwość receptorów wrażliwych na
rozciąganie we wrzecionkach nerwowo-
mięśniowych jest regulowana przez neurony
gamma.

Wrzecionko nerwowo-mięśniowe.
W nerwie mięśniowym biegną
włókna aferentne:
pierwotne od zakończeń
pierścieniowato-spiralnych (IA),
wtórne - od zakończeń w postaci
„wiązanki kwiatów",
oraz wlokna eferentne kończące
się zakończeniami synaptycznymi
nerwowo-mięśniowymi w
odcinkach poprzecznie
prążkowanych intrafuzalnych
komórek mięśniowych (wg
Barkera).

• W każdej jednostce motorycznej
występuje jedna komórka nerwowa,
która łączy się z komórkami
mięśniowymi za pomocą zakończeń
aksonu.
• Miejsce stykania się wypustki komórki
nerwowej, czyli aksonu, z komórką
mięśniową nosi nazwę zakończenia
synaptycznego nerwowo-
mięśniowego (terminatio synapsis
neuromuscularis).

Mięśnie gładkie
• Komórka mięśniowa gładka
(myocytus glaber) w odróżnieniu od
komórki mięśniowej poprzecznie
prążkowanej nie ma jednostek
kurczliwych w postaci sarkomerów.

Komórki mięśniowe gładkie występują
w organizmie w dwóch różnych pod
względem czynnościowym
skupieniach. Tworzą one:
1. Wielojednostkowe mięśnie
gładkie, w których poszczególne
komórki kurczą się niezależnie i
pobudzenie nie przenosi się z jednej
komórki na drugą. Występują w
ścianach naczyń krwionośnych i w
tęczówce.

2. Trzewne mięśnie gładkie
stanowiące warstwy lub pierścienie
równolegle ułożonych komórek, w
których pobudzenie przenosi się z
jednej komórki na drugą. Trzewne
mięśnie gładkie występują w
ścianach przewodu pokarmowego, w
moczowodach, pęcherzu moczowym
i macicy.

Mięśnie gładkie kurczą się pod
wpływem:
• Samoistnego pobudzenia
które występuje rytmicznie w
niektórych komórkach trzewnych
mięśni gładkich. Pobudzenie szerzy
się na komórki sąsiednie
doprowadzając cały miesień do
skurczu. Komórki te spełniają funkcję
rozrusznika dla pozostałych komórek..

Mięśnie gładkie kurczą się pod
wpływem:
• Czynnika miejscowego
mechanicznego lub chemicznego
działającego bezpośrednio na
komórki, np. rozciąganie mięśnia,
zmiany pH, wzrost prężności
dwutlenku węgla.

Mięśnie gładkie kurczą się pod
wpływem:
• Związków chemicznych
wytworzonych w odległych tkankach i
przenoszonych drogą humoralną -
kontrola humoralna, np. hormony
rdzenia nadnerczy.

Mięśnie gładkie kurczą się pod
wpływem:
• Związków chemicznych
wydzielonych z aksonów neuronów
należących do układu
autonomicznego,
czyli na zasadzie kontroli nerwowej
za pośrednictwem uwalnianych
transmitterów.

Na komórki mięśniowe gładkie
działają dwa transmittery:
• noradrenalina (NA) wydzielana na
zakończeniach neuronów zazwojowych
należących do układu współczulnego
(pars sympathica systematis
autonomici)
• acetylocholina (ACh) wydzielana na
zakończeniach neuronów układu
przywspółczulnego (pars
parasympathica systematis autonomici).

Mięsień sercowy
Mięsień sercowy składa się w istocie
z dwóch mięśni. Są to:
• miesień przedsionków i
• miesień komór,
odgrodzone od siebie pierścieniami
włóknistymi (anuli fibrosi)
otaczającymi ujścia przedsionkowo-
komorowe (ostia atrioventricularia).

• Siła skurczu mięśnia sercowego
zależy od początkowej długości jego
komórek, podobnie jak to występuje
w mięśniach poprzecznie
prążkowanych szkieletowych.

• Energię potrzebną do skurczów
miesień sercowy czerpie ze
składników odżywczych.
• Są nimi w 60% tłuszcze, których
połowę stanowią wolne kwasy
tłuszczowe – FFA, w 35%
węglowodany oraz aminokwasy, ciała
ketonowe, kwas mlekowy i kwas
pirogronowy.

Komórki układu
przewodzącego serca
• Poza komórkami mięśnia
przedsionków i mięśnia komór
występują w sercu komórki układu
przewodzącego.

Komórki układu
przewodzącego
W komórkach tych brak jest wyraźnego
poprzecznego prążkowania, są bogate w glikogen i
zawierają więcej sarkoplazmy.
Lezą bezpośrednio pod wsierdziem tworząc
skupienia:
• węzeł zatokowo-przedsionkowy, czyli węzeł
Keitha-Flacka,
• węzeł przedsionkowo-komorowy, czyli węzeł
Aschoffa-Tawary,
• pęczek przedsionkowo-komorowy, czyli pęczek
Palladino-Hisa, który dzieli się na dwie odnogi
kończące się komórkami mięśniowymi sercowymi
przewodzącymi (włóknami Purkinjego).

• komórki układu przewodzącego
samoistnie i rytmicznie pobudzają
się. Tworzą tym samym rozrusznik
(pacemaker) dla całego mięśnia
sercowego

• komórki węzła zatokowo-
przedsionkowego
narzucają swój rytm wszystkim
pozostałym komórkom układu
przewodzącego i komórkom całego
mięśnia sercowego.

KONIEC
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
- Slide 41
- Slide 42
- Slide 43
- Slide 44
- Slide 45
- Slide 46
- Slide 47
- Slide 48
- Slide 49
- Slide 50
- Slide 51
- Slide 52
- Slide 53
- Slide 54
- Slide 55
- Slide 56
- Slide 57
- Slide 58
- Slide 59
- Slide 60
- Slide 61
- Slide 62
- Slide 63
- Slide 64
- Slide 65
- Slide 66
- Slide 67
- Slide 68
- Slide 69
- Slide 70
- Slide 71
- Slide 72
- Slide 73
- Slide 74
- Slide 75
Wyszukiwarka
Podobne podstrony:
ok Fizjologia Czynność komórek nerwowych i mięśniowych
Czynność komórek nerwowych i mięśniowych
Czynność komórek nerwowych
Czynność komórek nerwowych
ok Fizjologia czynności mięśni
Zagadnienia z fizjologii nerwów i mięśni na kolokwium OPRACOWANIE(1) (1)
Budowa komorek tkanki miesniowej i nerwowe j
ok Fizjologia czynności mięśni
czynność komórek mięśniowych 1
Złącze nerwowo mięśniowe
Fizjologia czynności tkanek krwiotwórczych
CZYNNOŚĆ STATYCZNA I DYNAMICZNA MIĘŚNIA, Fizjoterapia, Biomechanika
więcej podobnych podstron