
Transmisja synaptyczna w
zwojach współczulnych
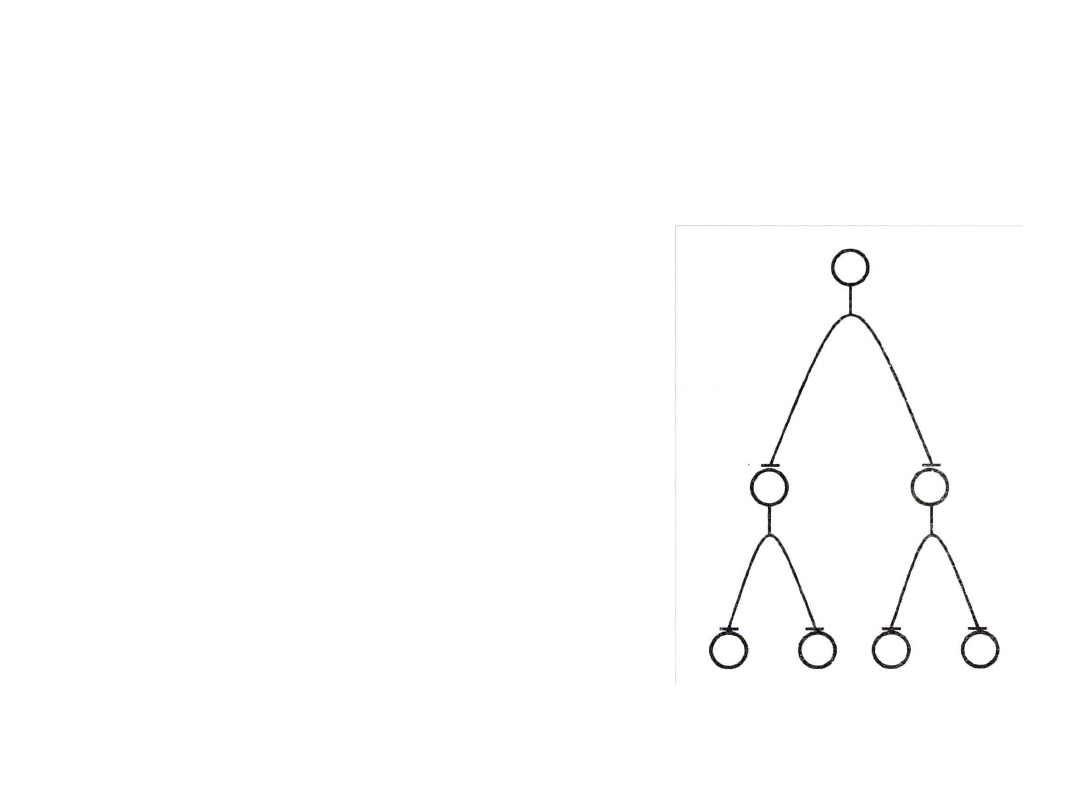
Dywergencja
• Każdy neuron
współczulny oddaje
synapsy na 10 – 100
neuronach zwojowych
• Nieliczne neurony
mogą
pobudzić
układ współczulny
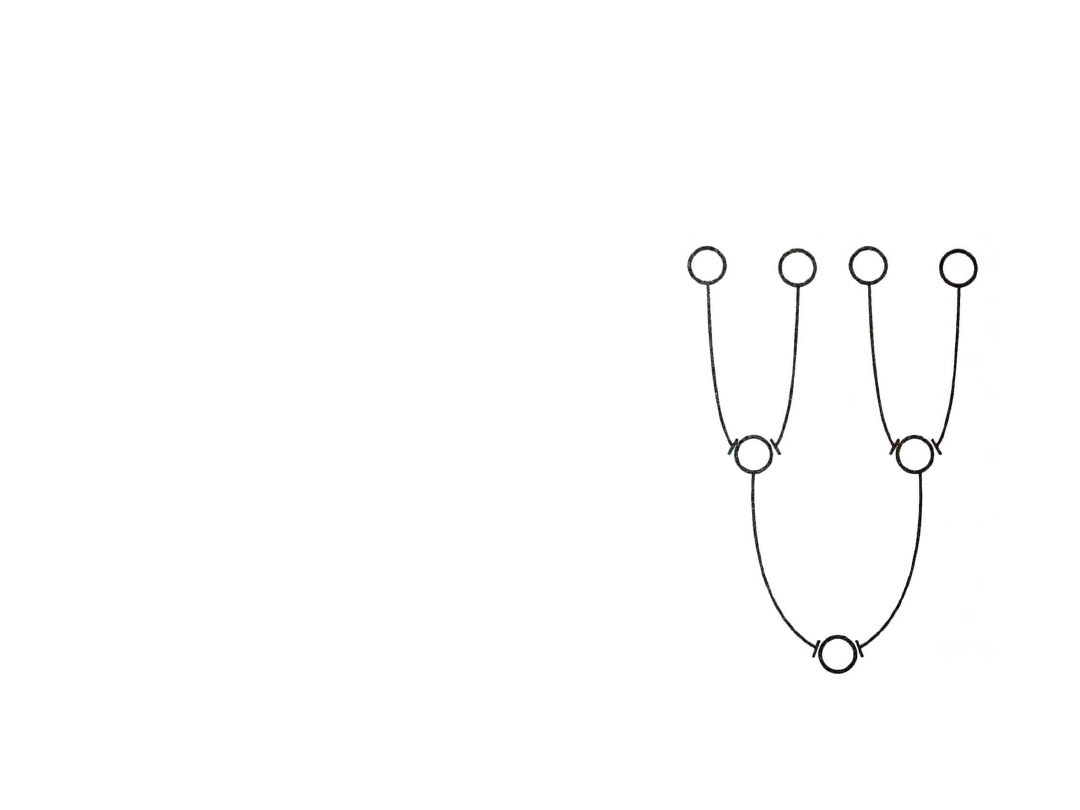
Konwergencja
• EPSP powstaje w
wyniku sumowania
postsynaptycznego
• Na pojedynczym
neuronie oddaje
4 - 15 neuronów
przedzwojowych

Charakterystyka EPSP
• Potencjał czynnościowy powstający w
wyniku
pobudzenia przez jeden
neuron
przedzwojowy
ma wyższą
amplitudę
niż powstający w wyniku
sumowania potencjałów
postsynaptycznych.
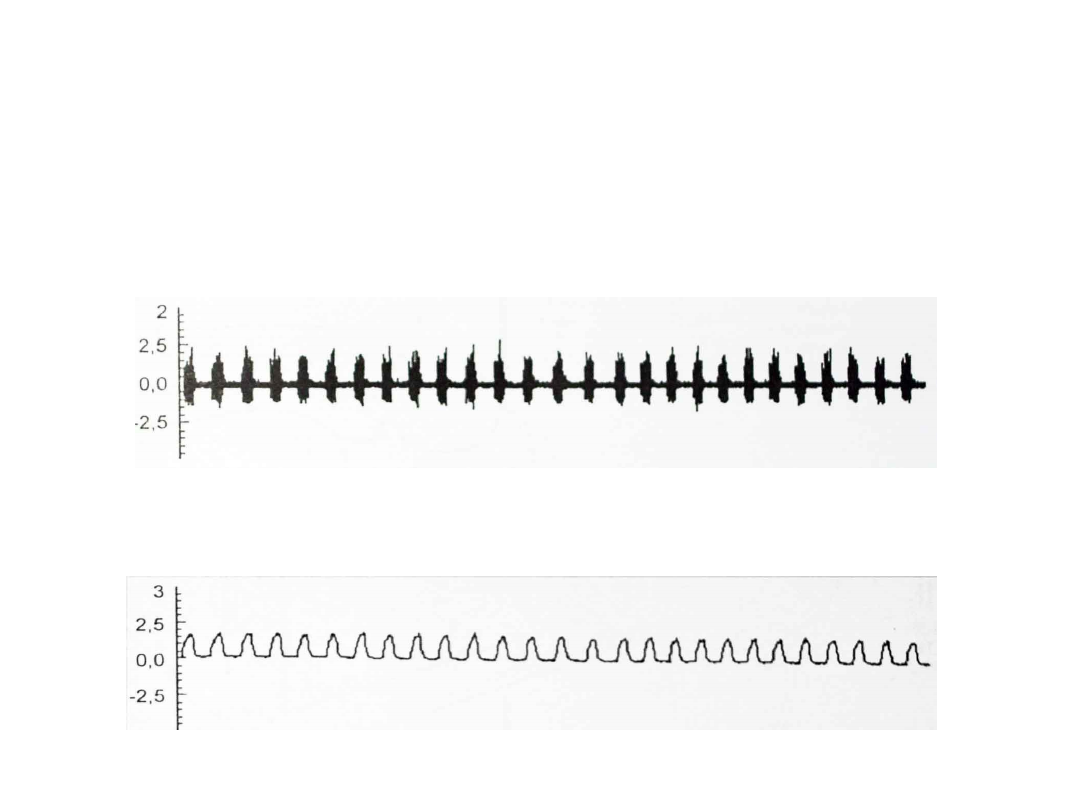
Typy potencjałów
postsynaptycznych
pobudzających
• Fazowe
– Neurony pobudzane przez pojedynczy
potencjał postsynaptyczny
– Długie serie potencjałów o dużej
częstotliwości
• Toniczne
– Wolny, stały rytm wyładowań
– Występuje w nich długa hiperpolaryzacja
następcza

Przyczyna hiperpolaryzacji
następczej
• Napływ jonów
Ca
i Na w fazie
depolaryzacji
aktywuje kanał potasowy
zależny od wapńa (hiperpolaryzacja)
• Neurony fazowe zawierają receptor
M
4
– Inaktywuje
kanały potasowe
• Skraca to odstępy między kolejnymi
pobudzeniami
• Zwiększa pobudliwość neuronu

Typy neuronów zwojowych
• B – większe
– Zmielinizowane
włókna przedzwojowe
typu B oddają na nich synapsę
• C – mniejsze
– Na nich zakończenia oddają cienkie ,
bezmielinowe
włókna typu C

• Acetylocholina uwalniana jest z
zakończeń przedzwojowych
– Działa na:
• Receptory cholinergiczne typu N
– Blokerem jest
heksametonium
(zarówno w
zwojach współczólnych i przywspółczulnych)
• Metabotropowe typu M
– Komórki wewnątrzzwojowe
dopaminergiczne
• mają tylko receptor typu M
Mają decydujące znaczenie dla
transmisji synaptycznej
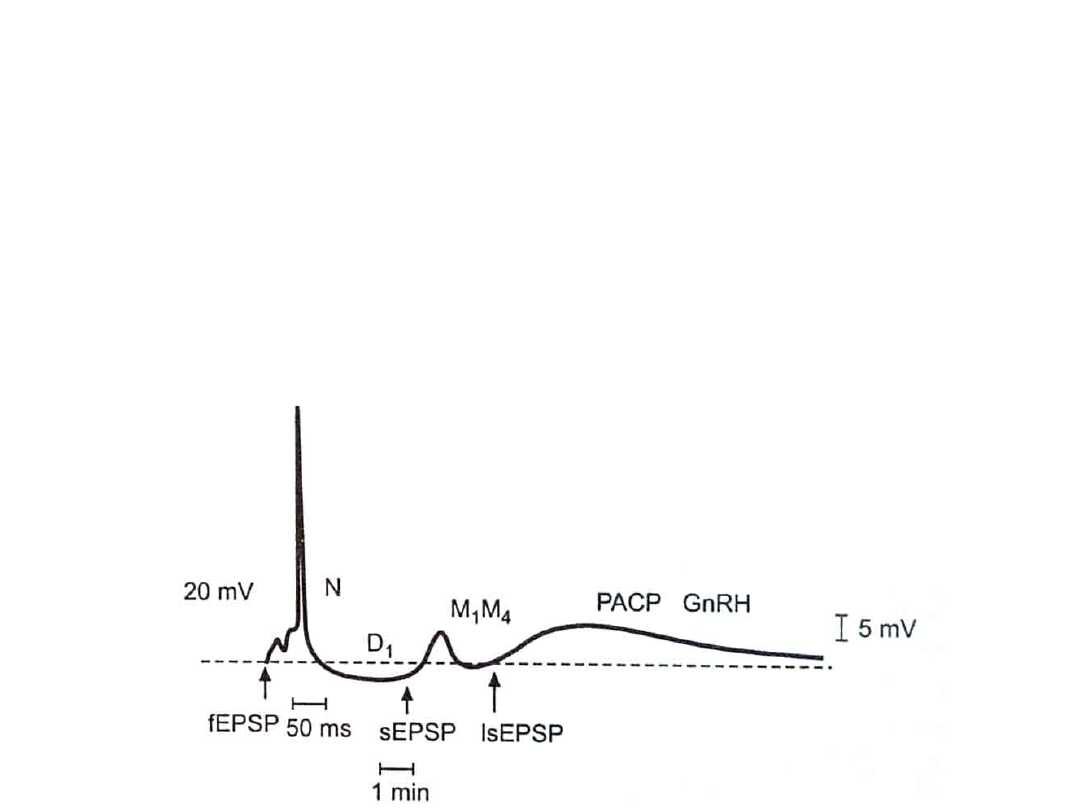
Transmisja synaptyczna -
fazy
• Wczesna
fEPSP
• Późna
s
EPSP
• Bardzo późna
isEPSP

Transmisja synaptyczna -
fazy
• Wczesna
– Odpowiada jej krótkotrwały duży
potencjał postsynaptyczny pobudzający
(
fEPSP
)
• Wywołany pobudzeniem receptora N
• Trwa 0,5 – 1 ms

Transmisja synaptyczna -
fazy
• Późna (sEPSP)
– Pojawia się po 100 – 200 ms
– Wywołana działaniem
acetylocholiny
na
• receptor metabotropowy M1
– Zmniejsza przewodność błony komórkowej dla
potasu.
• Receptor metabotropowy M4
– Hamuje zależny od wapńa kanał potasowy

Wpływ serotoniny
• serotonina
(obecna w zwojach
współczulnych)
– Działa pobudzająco poprzez receptor
metabotropowy
5-HT
3
– Wywołuje późną i bardzo późną
depolaryzację

Hamowanie oboczne
• Sprzyja ograniczeniu pobudzenia
– Noradrenalina działa na receptor
2B
hamuje syntazę cAMP
• Wzrost przewodności dla K i hiperpolaryzacja
– Adenozyna działa na receptor A1
redukując aktywność cyklazy
adenylanowej.

Wpływ ATP
• Kolaterale współczólne uwalniają ATP
– Działa pobudzająco na jonotropowy
receptor P2x
• Zwiększa napływ Ca

Postsynaptyczny potencjał
hamujący IPSP
• Zależy od małych komórek
zwojowych zawierających dopaminę
(SIF)
– Oddaja synapsę hamującą
• Dopamina działa na
• receptory typu D
1
– Zwiększa przewodność dla jonów K
• Hamuje uwalnianie acetylocholiny z
zakończeń przedzwojowych

Transmisja synaptyczna -
fazy
• Bardzo późna (IsEPSP)
– Wywołują go neuropeptydy uwalniane
przez cholinergiczne włókna C
• PACAP
– przysadkowy polipeptyd aktywujący
cyklazę adenylową
• GnRH – hormon podwzgórzowy uwalniający
gonadotropiny.
– Powolne zamykanie kanału potasowego i
zmniejszenie prądu odkomórkowego K

Transmisja synaptyczna -
fazy
• Bardzo późna (IsEPSP)
• PACAP
– przysadkowy polipeptyd aktywujący
cyklazę adenylową
– PACAP 27 i PACAP 38
– Pobudza neurony zwojów współczólnych
– Działa poprzez receptor PACAP
1
– Receptory dla PACAP obecne są w błonie
neuronów B oraz komórek zwojowych typu C
– Działają troficznie ułatwiając przeżycie
niedojrzałych neuroblastów w okresie
rozwojowym.

Depolaryzację powodują
także
• Substancja P
– Działa na metabotropowy receptor NKA
1
• CGRP (peptyd pochodny genu
kalcytoniny)
– Aktywuje receptor CGRP
1
• CKK-8 cholecystokinina
– Receptor CKK
A

Modulacja presynaptyczna
• Receptory opioidowe na włóknach
przedzwojowych.
– Synapsy aksonalno-aksonalne
• Uwalniana enkefalina leucynowa (ENK-L)
- hamuje uwalninie acetylocholiny
- Wpływa na strukturę molekularną
receptorów M i N (zmniejsza powinowactwo
do acetylocholiny)
- zmniejszenie liczby receptorów N

Enkefaliny
• Komórki chromochłonne rdzenia
nadnerczy są źródłem enkefalin
• Uwalniane są do krwi w stanach
stresu razem z adrenaliną i
noradrenaliną.
• Ograniczają długotrwałe pobudzenie
układu współczulnego
( ujemne sprzężenie zwrotne)

Plastyczność transmisji
synaptycznej
• Jest to zdolność do utrwalania śladów
pobudzenia
• Seria wysoko czestotliwych bodźców
-> krótkowrwałe wzmocnienie ->
zwiększenie amplitudy fEPSP
– Wzrost dokomórkowego Ca (kanły P i N)
– Zwiększenie wytważania acetylocholiny
(spowodowane wzrostem Ca)

Bloery kanałów wapniowych
• ω Agatotoksyna – Kanały typu P
• ω Konotoksyna – Kanały typu N

LTP – długotrwałe
wzmocnienie
postsynaptyczne
• Wzmożona ekspresja mRNA białka
receptora N
• NO w zwojach współczólnych

ANP
• Hamuje transmisję synaptyczną w
zwojach współczólnych u człowieka
– Aktywacja cyklazy guanylanowej

• W zwojach współczulnych zamykają się
łuki odruchowe.
• Neurony zwojowe tworzą rodzaj ośrodków
dla miejscowych odruchów w części
jelitowej AUN.
• Zwoje wsółczulne odbierają pobudzenie z
różnych receptorów przez zawracające
odgałęzienia trzewno-czuciowych
bezmielinowych włókien SP-ergicznych i
CGRP-ergicznych
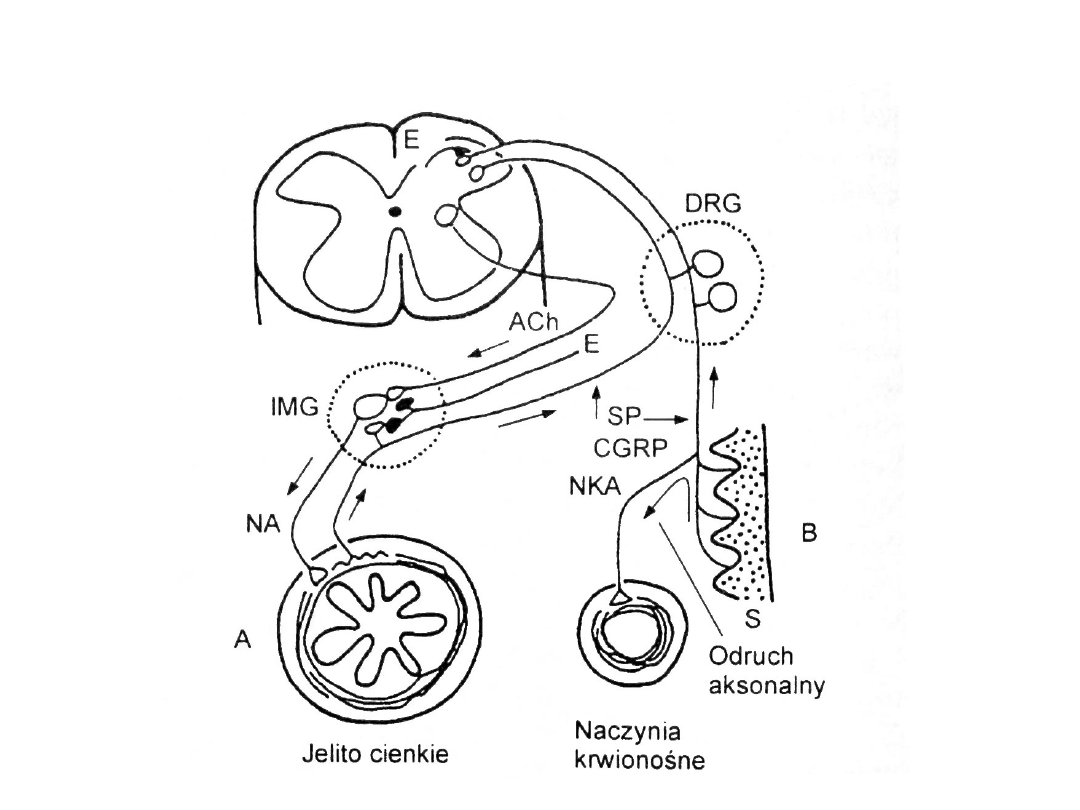

Bodźce nocyceptywne
• Pobudzają trzewnoczuciowe
bezmielinowe włókna bólowe, które
oddają zawracajace odgałęzienia do
zwojów współczólnych.

Rola substancji P oraz
cCGRP
• Torują transmisję synaptyczną
• Wzmagają bólową reakcję
współczulną i presyjną.
• Działają poprzez receptory:
– NKA
1
– substancja P
– CGRP
1
- cCGRP

Sprzężenie zwrotne
• Występuje pomiędzy
zwojami
współczulnymi
a
cholinergicznymi
neuronami zwojów części jelitowej
układu autonomicznego.

CZĘŚĆ
TRZEWNOCZUCIOWA
AUTONOMICZNEGO
UKŁADU
NERWOWEGO:
odruchy współczulne i
aksonalne

Część trzewnoczuciową tworzą
włókna aferentne
bezmielinowe
typu Cdr
lub
cienkie mielinowe
typu Aδ
oraz ich neurony czuciowe w
zwojach grzbietowych rdzenia
kręgowego.Przewodza
pobudzenia z narzadów
wewnętrznych do
ośrodkowego układu
nerwowego w trzewnych
odruchach autonomicznych.

Dzielimy je na:
1)
trzewno – trzewne
2)
trzewno – somatyczne
3)
somatyczno - trzewne

1)
Bodziec działa na
wisceroreceptory w narządach
wewnętrznych. Efektorem jest
układ krążenia, przewód
pokarmowy (np.protekcyjne
rdzeniowe odruchy żołądkowe i
jelitowe) i inne. Najważniejsze
to:
a.
Odruch nerkowo – nerkowy
b.
Odruch wątrobowo – nerkowy
c.
Odruch wątrobowo - jelitowy

2)
Odruchy trzewno – somatyczne,
od wisceroreceptorów do
neuronów ośrodkowych i następnie
do somatycznych neuronów
eferentnych. Przykładem
patologicznego odruchu jest objaw
tzw.
obrony mięśniowej
, oraz
zjawisko tzw.
bólu odniesionego
-
np. ból wieńcowy przy zawale
obejmujący lewe ramię, obojczyk
czy podbrzusze, imitując ból
żołądka.

3)
Odruchy somatyczno – trzewne.
Odpowiedż ze strony układu
autonomicznego na bodżce działające
na somatyczne eksteroreceptory czy
narządy zmysłów. Przykładem są:
a.
Presyjne odruchy z ergoreceptorów
lub
metaboreceptorów
mięśni
szkieletowych pobudzanych podczas
wysiłku fizycznego, zwłaszcza
statycznego.
b.
Odruchy
somatyczno – współczulne
(odpowiedź
wczesna
,
późna
,
bardzo
późna
) np..przyspieszenie rytmu
serca i presyjna reakcja bólowa na
podrażnienie nocyceptorów skórnych.

Charakterystycznym dla układu
autonomicznego rodzajem regulacji są
odruchy zwojowe zamykające się na
poziomie neuronów zwojowychbez
udziału ośrodkowego układu
nerwowego.W zwojach pnia
współczulnego znajdują się komórki
czuciowe włókien trzewnoczuciowych
pochodzących z receptorów serca, aorty,
przełyku, oskrzeli.

Informacja z tych narządów jest
przekazywana wewnątrz zwoju do
neuronówodśrodkowych, które
wysyłają swoje włókna pozazwojowe
do tych samych lub sąsiednich
narządów.Takie współczulne odruchy
zwojowe pobudzają np. czynność
serca pod wpływem rozciągnięcia
mechanicznego ścian komór lub
pobudzenia nocyceptorów, np. w
niedokrwieniu lub zawale serca.

Włókna trzewnoczuciowe pełnią też
funkcję eferentną, oddając kolaterale
do wielu narządów.
W neuronach czuciowych ekspresji
ulega gen preprotachykininy (PPT):
α- PPT -A
Β- PPT -A
γ- PPT -A
Neurokinina K

Pobudzenie wisceroreceptorów lub
receptorów powierzchni ciała
powoduje uwolnienie całej rodziny
neurokinin orazCGRP z obwodowych
kolaterali włókien czuciowych
neurokininy i CGRP powodują
miejscowe rozszerzenie naczyń
krwionośnych, obrzęk, objawy stanów
zapalnych, pobudzają migrację
leukocytów i innych komórek
immunoreaktywnych do miejsca
uszkodzenia i uwalnianie przez nie
cytokin.

Neurony części jelitowej wbrew
swojej nazwie rozmieszczone są w
całej ścianie przewodu
pokarmowego od przełyku aż po
odbyt.

Skupiają się one w dwóch splotach
śródściennych:
1) Podśluzówkowym, regulującym czynność
wydzielniczą komórek bony śluzowej,
resorpcję i śluzówkowy przepływ krwi.
2) Śródmięśniowy, kierujący motoryką mięśni
gładkich
Neurony splotów jelitowych, podobnie jak
neurony mózgu, otoczone są komórkami
glejowymi. Działają one jako ośrodki
lokalnych odruchów, integrując wzorce
ruchowe przewodu pokarmowego,
wydzielanie gruczołów trawiennych, lokalne
zmiany przepływu krwi, wchłanianie, a także
wydzielanie śluzu i soku jelitowego.

Neurony splotów jelitowych odkrył
w XIX wieku wybitny
neurohistolog polskiego
pochodzenia Aleksander Dogiel

W splotach śródściennych znajdują się szczególne
neurony-komórki Calaja.Działają one jak komórki
rozrusznikowe-wykazują powolną spoczynkową
depolaryzację, niezależną od pobudzeń
synaptycznych.
Rytmiczna aktywność neuronów splotu
śródmięśniowego powoduje depolaryzacje mięśni
gładkich żołądka i jelit, generując podstawowy rytm
elektryczny(BER).
Falom BER towarzyszą zsynchronizowane fale
skurczów mięśniowych, tworząc łącznie wędrujące
kompleksy mioelektryczne(MMC).Tworzą one fale
perystaltyczne, przebiegające od dna żołądka, gdzie
znajduje się strefa rozrusznikowa aż do
odźwiernika.Spoczynkowa częstotliwość
podstawowego rytmu elektrycznego BER w żołądku
wykosi 2-4/min.

W dwunastnicy znajduje się druga
strefa rozrusznikowa BER.Jest on
szybszy niż w żołądku, wynosi około
12/min i rozchodzi się z dekrementem
w stronę jelita grubego.

Synapsy splotu mięśniowego uwalniają liczne
transmitery i modulatory. Główną rolę
pobudzającą odgrywa acetylocholina.
Towarzysza jej liczne neuropeptydy
pobudzające:motylina, VIP, peptyd uwalniający
gastrynę(GRP), substancja P oraz
hamujące:somatostatyna, enkefaliny, galanina,
NPY.Z amin biogennych są to:serotonina i
dopamina.

Włókna współczulne za pośrednictwem
receptora adrenergicznego typu alfa hamują
także bezpośrednio napięcie i ruch samych
komórek mięśniowych, z wyjątkiem mięsni
zwieraczy pobudzanych do skurczu.
Nerw błędny przez swe cholinergiczne włókna
przedzwojowe i receptor typu N wzmaga
aktywność splotów jelitowych i przyspiesza
częstość rytmu komórek Calaja, za ich
pośrednictwem, częstość rytmu
podstawowego BER i wędrujących
kompleksów mioeletrycznych MMC.

Wydzielanie soku jelitowego pobudzają także
inne neuromodulatory. Jako aminy biogenne
działają sokopędnie w jelitach histamina i
serotonina, a także prostaglandyny typu E.
Z grupy peptydów są to:neurotensyna, GRP,
wazopresyna, bradykinina i kalcytonina.
Prawdopodobnymi transmiterami
wzmagającymi absorpcję jelitową
są:somatostatyna, NPY, CCK-8 oraz enkefaliny.
Spośród amin biogennych :noradrenalina,
adrenalina i dopamina.

Odruchy nocyceptywne z błony śluzowej
jelita działają jak sygnalizacja alarmowa
chroniąca błonę śluzowa żołądka,
dwunastnicy i jelita przed uszkodzeniem.
Głównym transmiterem ochronnym
uwalnianym na drodze krótkich,
zwojowych i długich, rdzeniowych i
ponadrdzeniowych odruchów z
chemoreceptorów błony śluzowej
żołądka i jelit jest peptyd pochodny genu
kalcytyninowego(CGRP).

Pewien udział ma także tachykinina
A(NKA).
Działanie ochronne CGRP i NKA polega
na uwalnianiu tlenku azotu.Te dwie
substancje zmniejszają także kwasotę
soku żołądkowego.

Eferentne neurony splotów trzewnych
unerwiają immunoreaktywne komórki tuczne
ściany przewodu pokarmowego, powodując
ich degranulację i uwolnienie histaminy.
Alergeny i antygeny działające od strony
przewodu pokarmowego często powodują
biegunki. Włókna eferentne splotów
trzewnych unerwiają także płytki Peyerta
ściany jelita, gdzie gromadzą się limfocyty,
leukocyty kwasochłonne i makrofagi. W ten
sposób część jelitowa układu autonomicznego
współdziała w komórkowej odpowiedzi
immunologicznej.

Transmisję synaptyczną w
autonomicznych splotach jelitowych
określamy jako wielochemiczną.

Czucie bólu
BODZIEC
NOCYCEPTYWNY
NIEAKTYWNE
KALIKREINY
TKANKOWE
AKTYWNE
KALIKREINY
TKANKOWE
KININOGENY
TKANKOWE
KININY (tez
histamina)
SPOLARYZOWANE
NAGIE
ZAKOŃCZENIA
NERWOWE
ZDEPOLARYZO
WANE NAGIE
ZAKOŃCZENIA
NERWOWE
SALWY
IMPULSÓW
BÓLOWYCH

Impulsy bólowe są
przewodzone przez włókna
nerwowe:
•Z osłonką mielinową – należą one do grupy A ,o
średnicy 2-5 μm i prękości przewodzenia 12-30m/s.
•Bez osłonki mielinowej- należą do grupy Cdr, o
śr. 0,4-1,2 μm i prędkości przewodzenia 0,5-
2m/s
Podwójna odpowiedź bólowa:
•Wywołane jest to różnicą prędkości przewodzenia
impulsów bólowych we włóknach grupy A i Cdr
•Bodźce bólowe są podwójnie odczuwane.
•Pierwsza odpowiedź bólowa odznacza się silnym
ostrym bólem, który jest ściśle umiejscowiony.
•Druga natomiast daje nieprzyjemne uczucie bez
ścisłego umiejscowienia.

„Czuciowe neuropeptydy”:
•Są uwalniane na synapsach w rogach tylnych rdzenia
oraz z zakończeń włókien czuciowych w unerwianych
tkankach (np. skóra, mięśnie, stawy, narządy
wewnętrzne)
•Do tej grupy należą: substancja P (+), peptyd
pochodny kalcytoninowego genu(+), wazoaktywny
peptyd jelitowy(+), somatostatyna(+, -), galanina(-).
•Powodują rozkurcz naczyń krwionośnych, zwiekszenie
przepuszczalności naczyń włosowatych i powstawanie
obrzęku zapalnego.
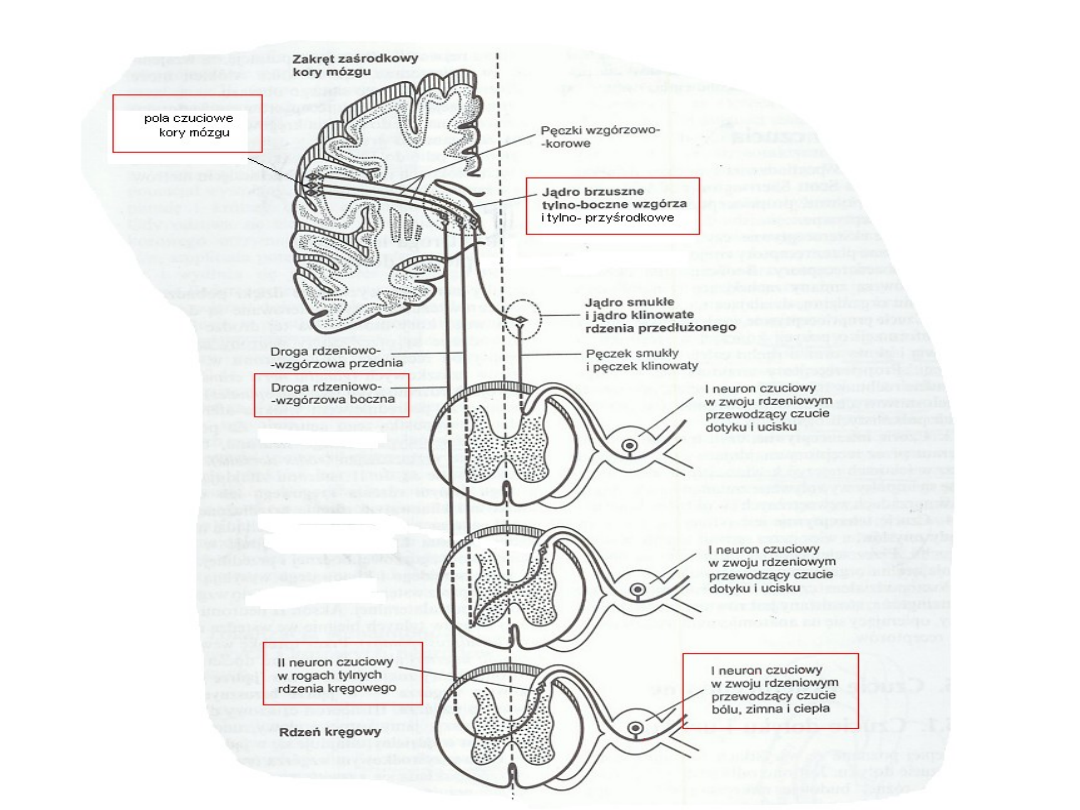
Droga impulsacji bólowej:

Hamowanie czucia bólu:
•Hamowanie impulsacji bólowej zachodzi w obrębie
rogów tylnych rdzenia kręgowego oraz w jądrach
wzgórza.
•Ten mechanizm objaśnia hipoteza „bramki kontrolnej”
dla bólu. Według niej w rogach tylnych rdzenia znajduje
się mechanizm działający jak bramka przepuszczająca
impulsy biegnące od pierwszego neuronu czuciowego (w
zwojach rdzeniowych) do drugiego neuronu czuciowego
(w rogach tylnych).
•Dzięki istnieniu bramki może zachodzić zwiększanie lub
zmniejszanie dopływu impulsów nerwowych.
•Przewodzenie impulsów czuciowych może być także
regulowane w jądrach części tylnej wzgórza między II a
III neuronem czuciowym.

Ośrodkowa modulacja czucia
bólu:
•W błonie komórkowej takich struktur mózgowia jak
śródmózgowie, podwzgórze i układ limbiczny występują
receptory opioidowe, które wiążą się z peptydami
opioidowymi (np.. Enkefalina, β-endorfina) działającymi
jak modulatory synaptyczne.
•U ludzi skarżących się na przewlekłe bóle zawartośc
peptydów opioidowych w płynie mózgowo-rdzeniowym
jest niewielka.

Metody usuwania czucia bólu:
FARMAKOLOGICZNE:
•Za pomocą znieczulenia miejscowego- hamuje
przewodzenie impulsów.
CHIRURGICZNE:
•Zniszczenie lub uszkodzenie dróg przewodzących
impulsy bólowe w rdzeniu kręgowym (jedno lub
obustronne)- CHORDOTOMIA (polega na przecięciu
drogi rdzeniowo- wzgórzowej bocznej . Znosi czucie
bólu i temperatury poniżej miejsca przecięcia.
Zachowane jest czucie dotyku.
Trwale zniesione zostają także wszystkie rodzaje
czucia z narządów płciowych.

Badanie czucia bólu:
•Metoda do oceny siły bodźca (J. D. Hardy) wykorzystująca
bodziec termiczny w postaci 1000-watowej lampy (zapalonej
na 3s i ustawionej w stałej odległości od skóry)
•Dzięki tej metodzie można oznaczyć bodziec progowy i
maksymalny czucia bólu. Energia do wywołania
maksymalnego czucia bólu jest tylko dwukrotnie większa od
en. Wywołującej minimalne czucie bólu.
•Ta mała różnica ma ogromne znaczenie, ponieważ bodźce
wywołujące ból są jednocześnie bodźcami uszkadzającymi tkankę.
•Różnorodne czynniki mogą wybiórczo znieść przewodzenie
impulsów np. brak tlenu znosi przewodzenie bólowe we
włóknach A, a kokaina we włóknach Cdr.
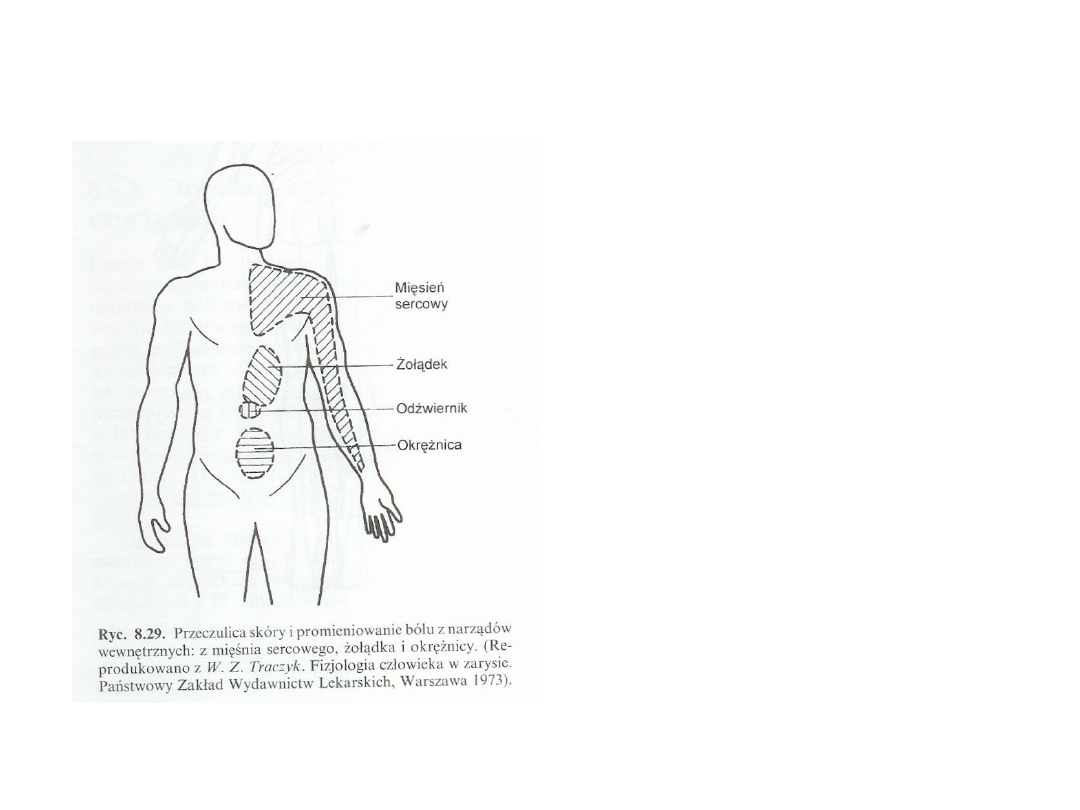
Unerwienie metameryczne:
Budowa metameryczna ciała
zachowała się w rdzeniu
kręgowym oraz w unerwieniu
skóry i mięśni. Narządy
wewnętrzne w rozwoju
ontogenetycznym utraciły w
znacznym stopniu budowę
metameryczną. Mimo to
stwierdza się przeczulicę skóry
unerwionej przez ten sam
odcinek rdzenia, który odbiera
impulsację aferentną z chorego
narządu wewnętrznego. Ma to
duże znaczenie praktyczne
ponieważ na podstawie
określenia miejsca przeczulicy
skóry można rozpoznać proces
chorobowy toczący się w
narządzie wewnętrznym (odruch
trzewno-skórny).

Czucie interoceptywne:
•Czucie trzewne odbierane jest przez interoreceptory
znajdujące się w narządach wewnętrznych i ścianach
naczyń krwionośnych.
•Są one pobudzane przez bodźce chemiczne,
mechaniczne i osmotyczne.
•Impulsacja wysyłana z tych receptorów biegnie w
nerwach czaszkowych, rdzeniowych i w nerwach
należacych do układu autonomicznego.
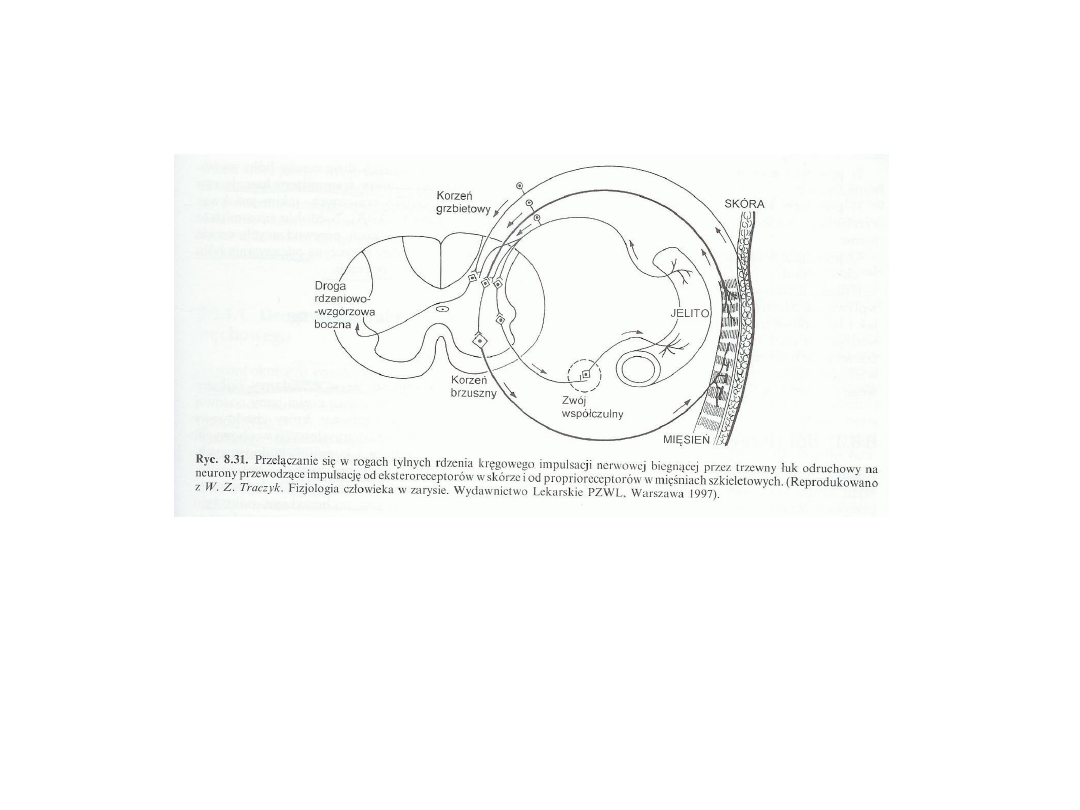
Czucie bólu trzewnego:
Proces chorobowy w narządach wewnętrznych wywołuje
podrażnienie interoreceptorów. Przez trzewny łuk odruchowy jest
przenoszona większa liczba impulsów, co powoduje wzmocnienie w
rdzeniu kręgowym impulsacji biegnącej od proprioreceptorów w
mięśniu poprzecznie prążkowanym. Prowadzi to do wzmożonego
napięcia mięśnia (odruch trzewno mięśniowy)
Również zostaje wzmocniona impulsacja z eksteroreceptorów
skórnych (przeczulica, odruch trzewno skórny)

Często jest tak, że ból trzewny odczuwa się w innym
miejscu niż leży chory narząd. Na przykład ból w napadzie
kolki żółciowej odczuwany jest pod łopatką, a ból w chorobie
wieńcowej serca promieniuje do ręki.
Wynika to z pobudzania leżących obok siebie szlaków
nerwowych i ośrodków w centralnym układzie nerwowym.
Impuls bólowy "rozlewa się" po sąsiednich komórkach
nerwowych.
Ból który budzi dziecko ze snu lub pojawia się po spożyciu
Ból który budzi dziecko ze snu lub pojawia się po spożyciu
płynów lub pokarmów stałych, ma często przyczynę w
płynów lub pokarmów stałych, ma często przyczynę w
przełyku.
przełyku.
Typowy ból trzewny trwa godzinami stopniowo zwiększając i
Typowy ból trzewny trwa godzinami stopniowo zwiększając i
zmniejszając natężenie.
zmniejszając natężenie.

Odnerwienia znoszące czucie
bólu:
1. Obłuszczenie przydanki tętnic doprowadzających krew do
narządu – usuwa się włókna aferentne biegnące wzdłuż
tętnic
2. Przecięcie nerwów należących do układu autonomicznego
3. Przecięcie korzeni grzbietowych przez które przebiegają
włókna aferentne od interoreceptorów.
4. Przecięcie drogi rdzeniowo-wzgórzowej bocznej
(chordotomia)

Ból przewlekły:
Przecięcie aferentnych dróg czucia bólu wywołuje
zmniejszenie syntezy transmittera hamującego w neuronach
GABA-ergicznych jakim jest kwas γ-aminomasłowy.
Niedobór transmittera hamującego w ośrodkach
przewodzących czucie bólu może być również przyczyną
odczuwania bólu pomimo wykonanej operacji.

KONIEC
Dziękujemy za uwagę
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
- Slide 41
- Slide 42
- Slide 43
- Slide 44
- Slide 45
- Slide 46
- Slide 47
- Slide 48
- Slide 49
- Slide 50
- Slide 51
- Slide 52
- Slide 53
- Slide 54
- Slide 55
- Slide 56
- Slide 57
- Slide 58
- Slide 59
- Slide 60
- Slide 61
- Slide 62
- Slide 63
- Slide 64
- Slide 65
- Slide 66
- Slide 67
Wyszukiwarka
Podobne podstrony:
Wpływ greliny na przewód pokarmowy 2007
Wpływ temp na przewo-dnictwo elektryczne, f2, Temat: Pomiar czujnikiem indykcyjnym.
Wpływ stresu na układ pokarmowy(1)
Wpływ stresu na układ pokarmowy
Wp3yw niesterydowych leków przeciwzapalnych na przewód pokarmowy, INTERNA, Gastroentero
Wpływ stresu na układ pokarmowy1
3 wpływ stresu na układ pokarmowy (agnieszka jednacz krzysztof korczynski)
Wpływ temperatury na przewodzenie złącza PN
Wpływ stresu na motorykę przewodu pokarmowego ready
Wpływ wielonienasyconych kwasów tłuszczowych na perystaltykę przewodu pokarmowego
Wpływ stresu na motorykę przewodu pokarmowego ready
(5) Wpływ leków na wchłanianie składników pokarmowyc h
25 WPŁYW STĘŻENIA MOCMNEGO ELEKTROLITU NA PRZEWODNICTWO
LEKI WPŁYWAJĄCE NA FUNKCJĘ PRZEWODU POKARMOWEGO
Dolny odcinek przewodu pokarmowego a alergia na pokarm
Naturalny lek na zakażenia?kteryjne i grzybicze przewodu pokarmowego
więcej podobnych podstron