
Dr hab. Ewa Rembiałkowska
Dr hab. Ewa Rembiałkowska
Szkoła Główna Gospodarstwa
Szkoła Główna Gospodarstwa
Wiejskiego
Wiejskiego
Zakład Żywności Ekologicznej
Zakład Żywności Ekologicznej
e-mai
e-mai
l:
l:
rembiałk@alpha.sggw.wa
rembiałk@alpha.sggw.wa
w
w
.pl
.pl

EKOLOGIA PODSTAWOWA
EKOLOGIA PODSTAWOWA

Prekursorzy Haeckel’a
Arystoteles (384-322
p.n.e)
Hipokrates (ok. 460-377
p.n.e.)
OIKOΣ
Teofrast (ok. 370-287
p.n.e.)
Karol Linneusz (1707-
1778)
Ernst Haeckel, 1869
(żył 1834-1919)

CECHY ŻYCIA
CECHY ŻYCIA
• Swoisty skład chemiczny
• Nosicielami życia są osobniki
• Zachowanie właściwego poziomu organizacji
• Wymiana substancji z otoczeniem (pobieranie i przyswajanie,
wydalanie i
wydzielanie, oddychanie)
• Zdolność reagowania na bodźce zewnętrzne (wrażliwość,
pobudliwość)
• Wykonywanie ruchów
• Wzrost, rozwój, rozmnażanie
• Dziedziczenie
• Śmierć
• Zdolność przeciwstawiania się wzrostowi entropii

ROŚLINY A
ROŚLINY A
ZWIERZĘTA
ZWIERZĘTA
Podobieństwa
:
•budowa komórkowa
•ogólne prawidłowości procesów
życiowych
•właściwości struktur i zjawisk na molekularnym poziomie
organizacji
Różnice
(nie mają charakteru
powszechnego)
•samożywność
•nieruchome przytwierdzenie do
podłoża
•celulozowe ściany
komórkowe
•w zasadzie nieograniczony wzrost
•organy zawiązują się do końca życia osobnika
•procesy wzrostu
zlokalizowane
•brak tkanki nerwowej i organów zmysłów (?)

NAJSTARSZE ROŚLINY NA ZIEMI
Sosna ościasta
‘Matuzalem’ 4
800 lat rośnie w
Górach Białych,
Kalifornia USA
Mamutowiec
olbrzymi 3 300
lat
Fiztroya
cupresoides
3 622 lata
Jałowiec wschodni 2
675 lat

Cis pospolity 2000 lat
Drzewo oliwne w
Jerozolimie 2000 lat
Dąb Chrobry 750 lat
rośnie w gm.
Szprotawa
Dąb Bartek 700 lat
rośnie na terenie
Nadleśnictwa
Zagnańsk

Żółw słoniowy 200
lat
Wyspy Galapagos
Słoń afrykański
60-70 lat
Goryl (małpa
człekokształtna),
40 lat
Na świecie żyje około 40 osób, które maja
powyżej 110 lat, ale w tym tylko 4
mężczyzn:
Obecnie żyjący:
•Ruidavets Moll mieszkający na Majorce
ma 113 lat
Yukichi Chuganji w Japonii ma 114 lat
Rekordziści światowi:
•W Emiracie Al-Chajma zmarł w latach
osiemdziesiątych mężczyzna, który twierdził, że liczył
sobie 147 lat. Miał to być najstarszy człowiek w krajach
arabskich nad Zatoką Perską.
•Najstarsza mieszkanka Egiptu, licząca według jej słów
ponad 135 lat, oświadczyła, że podstawą osiągnięcia
sędziwego wieku jest spokój.
•Francuzka Jeanne Calmet, zmarła niedawno w wieku
122 lat.
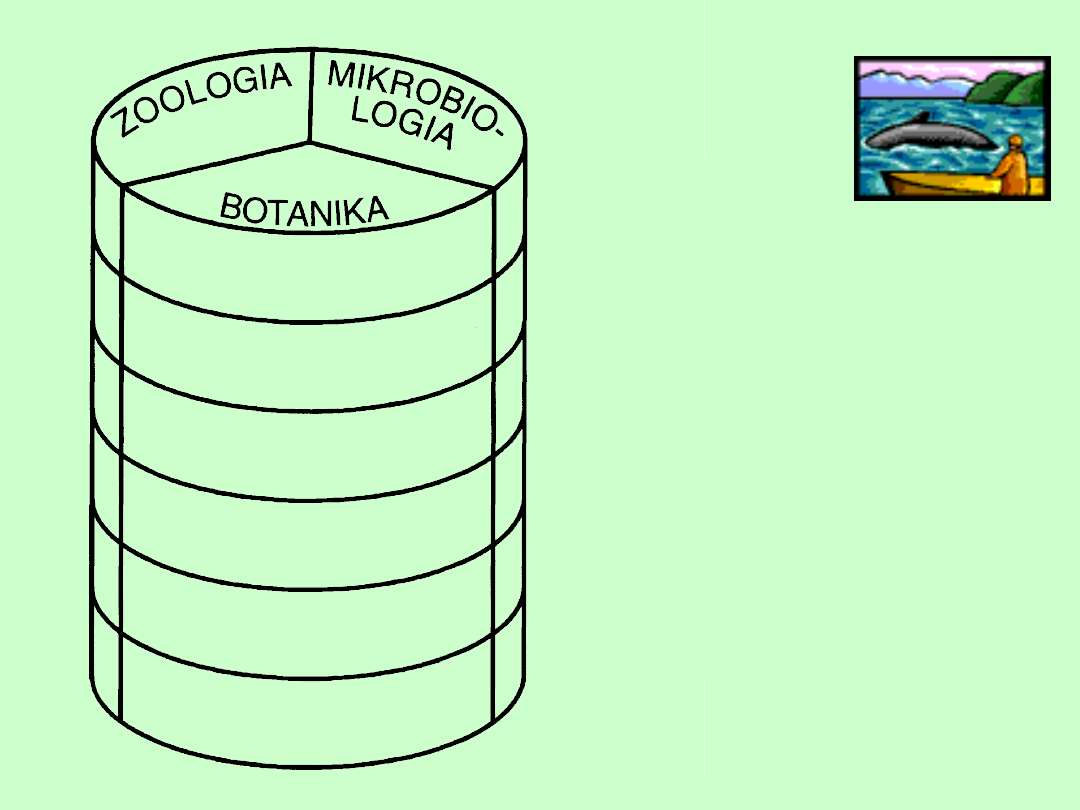
Biologia
molekularna
Morfologia
Fizjologia
Genetyka
Systematyka
Ekologia
POZIOMY ORGANIZACJI PRZYRODY
POZIOMY ORGANIZACJI PRZYRODY
(z punktu widzenia podejścia badawczego)
• molekularny
•komórkowy
•organizmalny (osobniczy)
• populacyjny
• gatunkowy
•biocenotyczny (ekosystemalny)
• fizjocenotyczny (krajobrazowy)
•biosfery
•kosmiczny

EKOLOGIA
EKOLOGIA
•
BIOLOGIA ŚRODOWISKA
•
EKONOMIA PRZYRODY
•
FILOZOFIA PRZYRODY

działania człowieka na rzecz naprawienia tego, co
człowiek zepsuł w przyrodzie
EKOLOGIA
OCHRONA
PRZYRODY
OCHRONA
ŚRODOWISKA
dziedzina nauk biologicznych, obok m.in. takich
jak biologia molekularna, fizjologia, genetyka,
taksonomia itd.
dziedzina wiedzy oraz działania praktyczne,
dotyczące zachowania istniejących zasobów
przyrody żywej i nieożywionej

AUTEKOLOGIA
osobnik
DEMEKOLOGIA = POPULACJOLOGIA
populacja ew.
gatunek
ekologia mikroorganizmów
ekologia roślin
ekologia zwierząt
ekologia człowieka
BIOCENOLOGIA
biocenozy lub
fizjocenozy
SYNEKOLOGIA
FIZJOCENOLOGIA

POJĘCIA ROZPATRYWANE NA POZIOMIE
POJĘCIA ROZPATRYWANE NA POZIOMIE
POPULACJI:
POPULACJI:
nisza ekologiczna (Charles Elton, 1927)
nisza siedliskowa
POJĘCIA ROZPATRYWANE NA POZIOMIE
POJĘCIA ROZPATRYWANE NA POZIOMIE
BIOCENOZY:
BIOCENOZY:
równowaga biocenotyczna (ekologiczna)
sukcesja
POJĘCIA ROZPATRYWANE NA POZIOMIE
POJĘCIA ROZPATRYWANE NA POZIOMIE
FIZJOCENOZY:
FIZJOCENOZY:
ekoton
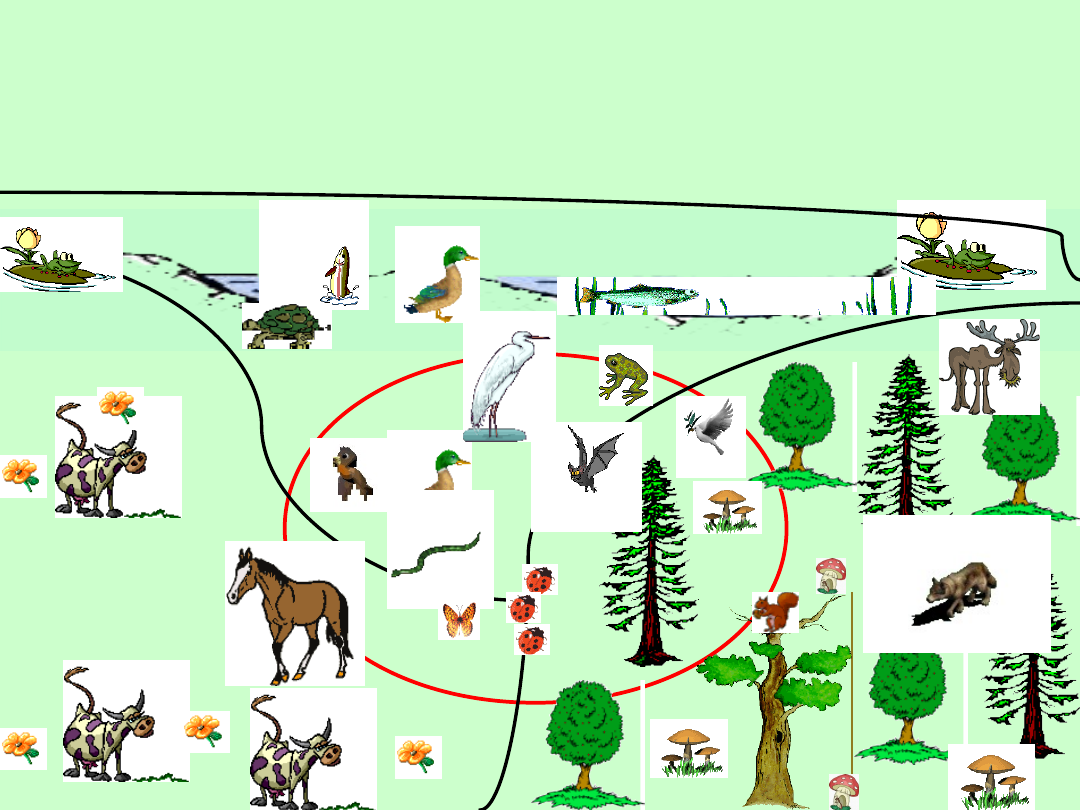
Pastwisko
Las
Rzeka
EKOTON

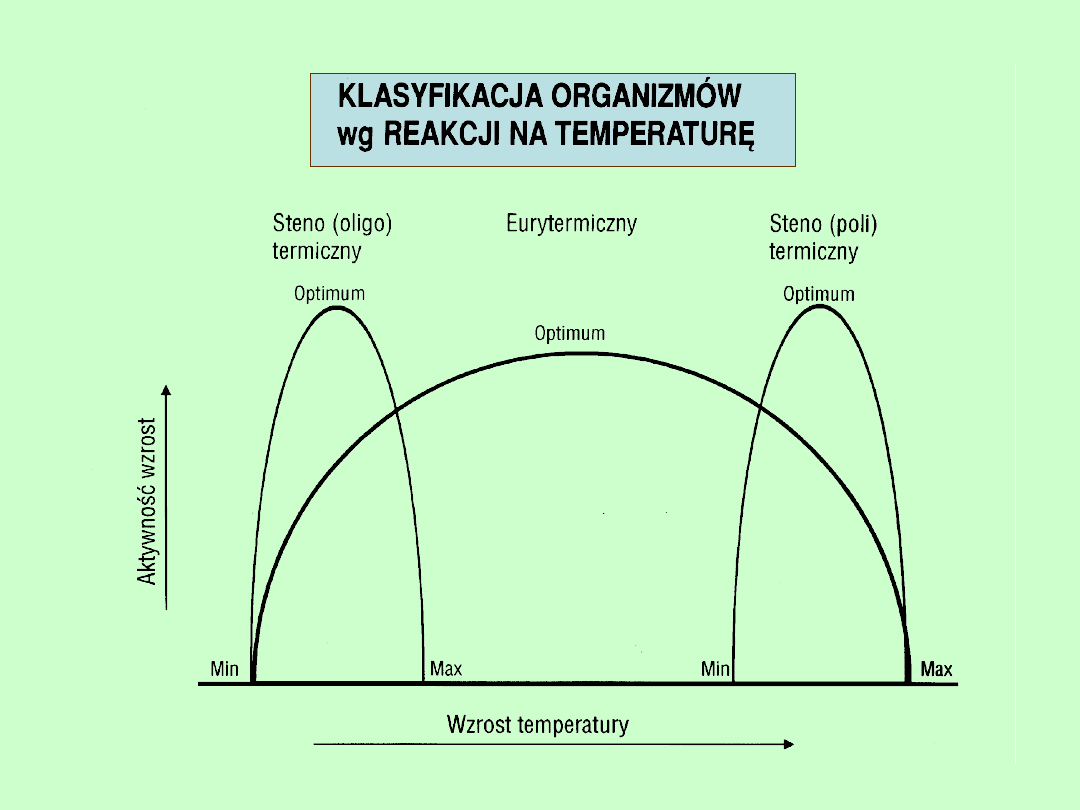
CZYNNIKI OGRANICZAJĄCE
CZYNNIKI OGRANICZAJĄCE
W ŚRODOWISKU FIZYCZNYM
W ŚRODOWISKU FIZYCZNYM
zakres (minimum, optimum maksimum)
TEMPERATURA
zmienność
kompensacja w stosunku do innych
czynników
ŚWIATŁO
WODA
TLEN
w wodach śródlądowych i w
glebie
ZASOLENIE i pH
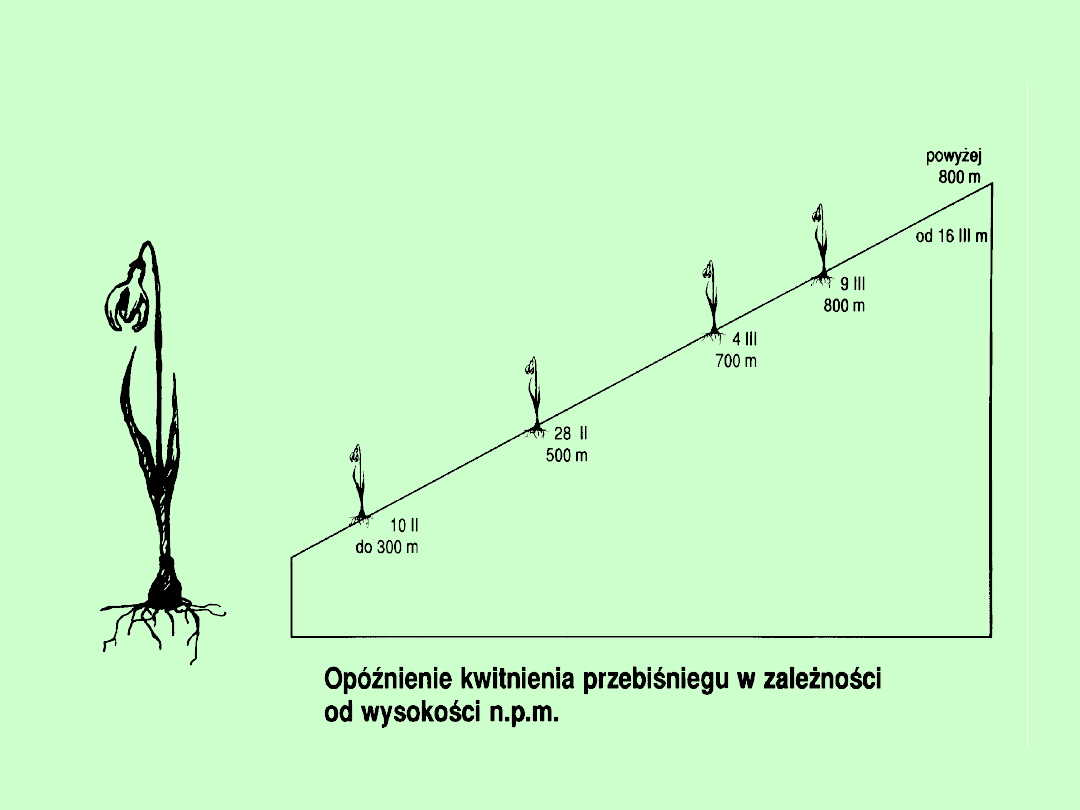
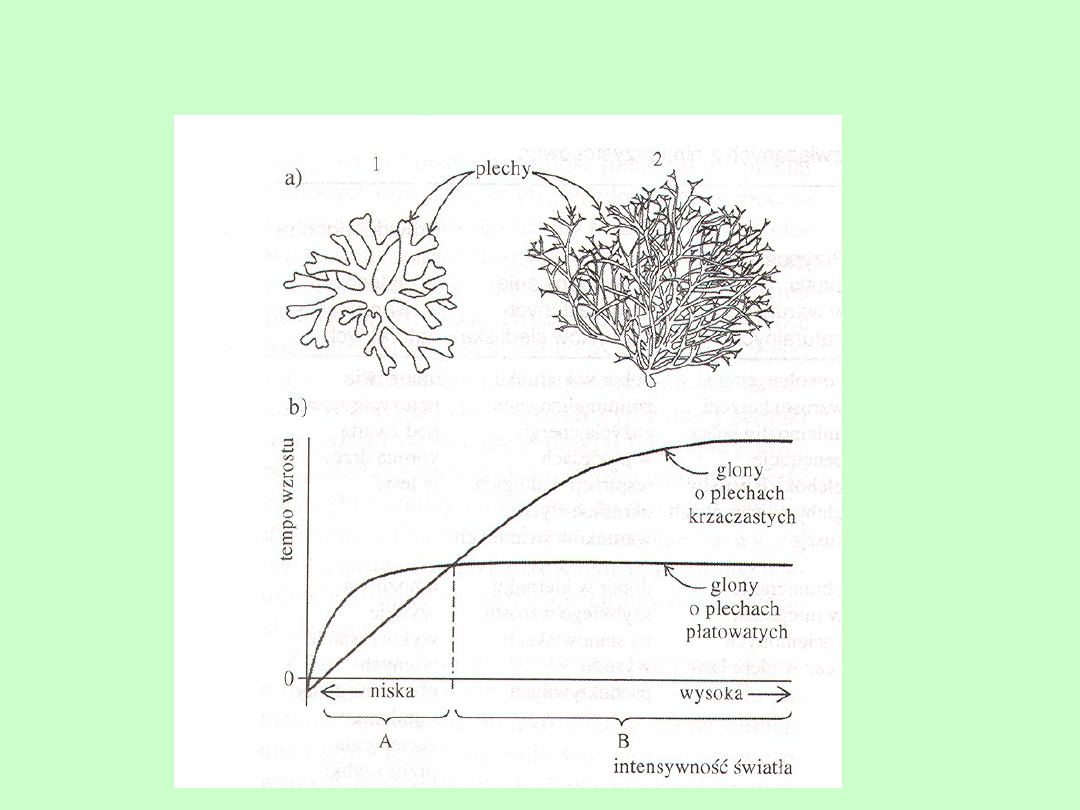
Wpływ światła na kształt plechy u glonów
morskich

JEŚLI ORGANIZM MA SZEROKI ZAKRES TOLERANCJI
W STOSUNKU DO WZGLĘDNIE STAŁEGO CZYNNIKA,
TO CZYNNIK TEN RACZEJ
NIE JEST CZYNNIKIEM OGRANICZAJĄCYM
(np. woda w zbiornikach wodnych).
JEŚLI ORGANIZM MA OKREŚLONE,
WĄSKIE GRANICE TOLERANCJI W STOSUNKU
DO ZMIENNEGO CZYNNIKA, TO TAKI CZYNNIK
MOŻE BYĆ OGRANICZAJĄCYM
(np. woda w glebie).

Gatunki
eurytopowe
Gatunki
kosmopolityczne
o dużym
zasięgu
=
Gatunek ten występuje prawie na
całej kuli ziemskiej z wyjątkiem
basenu Amazonki Występuje nad
brzegami wód stojących i
płynących, oraz na wszelkich
podmokłych terenach jak bagna,
przydrożne rowy, oraz wilgotne
łąki.
Trzcina
pospolita
Orlica
(paproć)
Roślina ta nie
występuje tylko w
rejonach arktycznych,
pustynnych oraz
stepowych oraz w
górach powyżej górnej
granicy lasu
traganek
Traganek duński, ale
we wszystkich
ekosystemach
półkuli północnej
znajdziemy jakieś
gatunki odpowiednio
przystosowanych
traganków
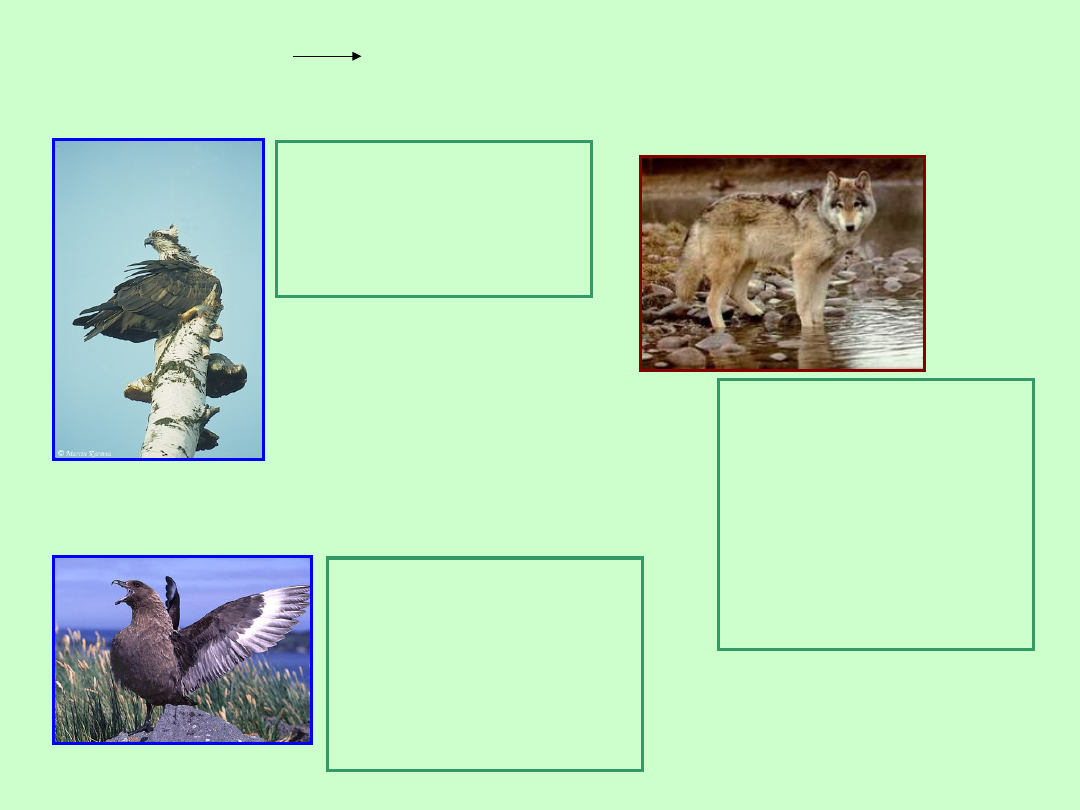
Gatunki
eurytopowe
Gatunki
kosmopolityczne
o dużym
zasięgu
=
rybołów
Rybołów występuje
nad wodami całego
świata np. w Polsce
nad Odrą oraz nad
Morzem Czerwonym
Wydrzyki występują
na wszystkich
brzegach mórz na
świecie. Są to ptaki
drapieżne np. w
okolicy Antarktydy
polują na pingwiny
Wydrzyk czyli
Skua
Wilk
Wilk zasiedla prawie
całą półkulę
północną, polując w
lasach Eurazji i
Ameryki Północnej na
zwierzynę drobną i
grubą (sarny, jelenie,
łosie). Preferuje życie
w tajdze

CECHY POPULACJI
CECHY POPULACJI
• liczebność (zagęszczenie)
• struktura płciowa
• struktura wiekowa
• liczba urodzeń
• śmiertelność (w tym w grupach wiekowych)
• organizacja wewnętrzna (rodziny, stada)
• odżywianie
• wykorzystywanie środowiska (rozmieszczenie)
• współwystępowanie z innymi populacjami

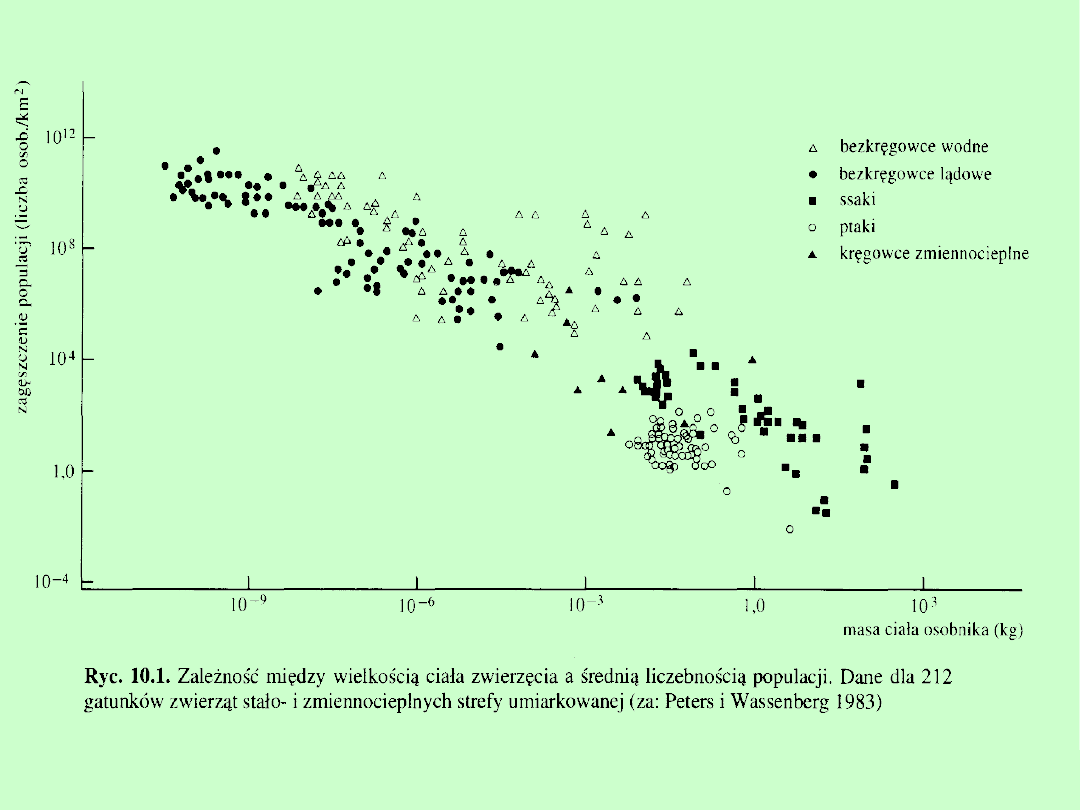
Zagęszczenie populacji
Zagęszczenie populacji
ludność Europy
6O osób/km²
ludność Australii 1,5 osoby / km²
jelenie w terenach leśno-polnych
1-15 osobników /
100 km²
sarny w terenach leśno-polnych
15-225 osobników /
100 km²
sadzonki sosny w uprawie leśnej
10-17 tysięcy
osobników/ ha
dorosłe sosny tworzące las
300-600 osobników / ha
dżdżownica kompostowa
3-5 tysięcy w m
3
kompostu
Właściwości populacji
Właściwości populacji
Populacja jest to grupa organizmów należących do tego samego gatunku,
współwystępujących na określonym obszarze i w określonym czasie.
Podstawowa cecha populacji to
zagęszczenie
.
Na zagęszczenie mają wpływ: rozrodczość, śmiertelność, imigracja i
emigracja.
Właściwości wtórne populacji: struktura wiekowa, struktura genetyczna)
struktura przestrzenna (sposób rozmieszczenia osobników).
imigracja
+
+ _
rozrodczość
zagęszczenie
śmiertelność
_
emigracja

Tab. 1. Liczba gatunków z głównych gromad roślin i typów zwierząt
Tab. 1. Liczba gatunków z głównych gromad roślin i typów zwierząt
Rośliny — gromada
Przybliżo
na liczba
gatunków
Zwierzęta - typy
Przybliżon
a liczba
gatunków
Zielenice (Chlorophyta)
Okrzemki (Bacillarophyta)
Brunatnice (Phaeophyta)
Krasnorosty (Rhodophyta)
Sinice (Cyanophyta)
Bakterie (Bacteriophyta)
Grzyby (Mycota)
Porosty (Lichenes))
Mszaki (Byophyta)
Widłaki (Lycododiophyta)
Paprocie (Polypodiophyta)
Nagonasienne(Gymnosperm
ae)
Okrytonasienne(Angiosperm
ae)
6 000
10 000
1 000
2 500
1 500
5 000
70 000
30 000
25 000
1 000
9 000
1 000
250 000
Pierwotniaki (Protozoa)
Gąbki. (Spongiae)
Jamochłony (Coelenterata)
Płazińce (Platyhelminthes)
Obleńce (Nemathelminthes)
PierścienIce (Annelida)
Mszywioły (Bryozoa)
Mięczaki (Mollusca)
Stawonogi (Arthropoda) bez
owadów
Owady (Insekta)
Szkarłupnie (Echinodermata)
Kręgowce (Vertebrata)
30 000
5 000
9 000
6 000
10 000
7 000
3 000
108 000
70 000
1 000
000
6 000
35 000
Rośliny - ogółem
412
000
1 289
000
Łączna liczba gatunków i zwierząt
1 701 000

Migracja osobników
Jest to proces o podstawowym znaczeniu, zapobiegający wsobnemu krzyżowaniu
w obrębie populacji, dzięki czemu odbywa się przepływ genów między
populacjami.
Migracja ogranicza geograficzne rozmieszczenie gatunku oraz wpływa na skład
zgrupowań wielogatunkowych.
Niektóre populacje istnieją tylko dzięki imigracji osobników, np. we wschodniej
części USA na małych płatach leśnych populacje drobnych ptaków śpiewających
utrzymują się jedynie dzięki imigracji.
Badanie migracji zwierząt jest trudne; główne metody to znakowanie i
radiotelemetria.

Jednym z głównych problemów ochrony przyrody jest
umożliwienie imigracji i emigracji w populacjach
zasiedlających izolowane fragmenty siedlisk.
Słabe zdolności migracyjne osobników mogą być przyczyną
rzadkości gatunku. Przykładem jest populacja
dzięcioła
skromnego Dendrocopos borealis,
niegdyś licznie
występującego na dużych obszarach USA, obecnie
uratowanego przed wyginięciem. Młode osobniki mogą albo
migrować w poszukiwaniu miejsc do osiedlania się, albo
pozostać jako osobniki towarzyszące parze lęgowej.
Kluczowym problemem jest wykucie nowej dziupli, które trwa
kilka lat. Dlatego ptaki wolą konkurować o istniejące już
miejsca lęgowe, niż osiedlać się w nowych miejscach, i
migrację rozpoczyna tylko 25% z prze samców.
Bardzo skuteczne okazało się wykucie dziupli w pniach sosen
w Karolinie Północnej - 18 na 20 stanowisk zostało
skolonizowanych przez dzięcioły skromne, które utworzyły tam
nowe grupy socjalne. Dzięki temu uratowano je przed
wyginięciem.

Rozrodczość
Rozrodczość
Płodność
- zdolność osobnika do wydawania potomstwa
Rozrodczość
- liczba potomstwa produkowanego w określonym czasie
Rozrodczość potencjalna
- w przypadku człowieka 1 noworodek /1 kobietę w wieku
rozrodczym / rok
Rozrodczość realizowana
- w przypadku człowieka 1 noworodek / kobietę w wieku
rozrodczym / 15 lat
Wskaźnik urodzeń
- liczba potomstwa produkowanego przez 1 samicę w jednostce
czasu,np.uostrygica.5Omln jaj/ rok, u ryb tysiące jaj/rok, u żab-setki jaj/rok, u ptaków 1-20
piskląt/rok,
u ssaków dużych 1- 2młode /rok, u ssaków małych np. u myszy 2O- 50/rok.
Śmiertelność
Śmiertelność
to liczba osobników umierających w jednostce czasu
Długość życia potencjalna
to maksymalny okres, który zdolny jest przeżyć osobnik danego
gatunku w optymalnych warunkach życia (określony przez granicę fizjologiczną)
Długość życia realizowana
jest to długość życia osobnika w populacji w określonych
warunkach środowiska, ograniczona przez choroby, drapieżców, inne zagrożenia. Np. rudzik
(Erithacus rubecula) w naturze żyje 1 rok, a w niewoli może żyć do 11 lat.
Średnia długość życia kobiety w Starożytnym Rzymie wynosiła 21 lat, w Anglii w 1780
roku -39 lat, a w USA w 1990 roku -79 lat.
Wskaźnik przeżywalności
- spadek liczebności osobników w kolejnych grupach wieku, np.
dla ryb:
Wskaźnik przeżywalności liczebność
*
ryb 3 letnich
= -----------------------------------
między 2 i 3 rokiem życia liczebność
*
ryb 2 letnich
* liczebność względna na podstawie ryb złowionych

Nie zawsze proporcja płci wynosi 1:1, np.
w populacjach lemingów leśnych w Finlandii udział
samców wynosi 25 %
w populacji wilgowronów meksykańskich (Teksas) w
grupie piskląt samce stanowią 50 %, ale wśród dorosłych
tylko 29 %
Proporcja płci wywiera wpływ na tempo rozrodu oraz na
interakcje socjalne.
Struktura płciowa

Są podobne do świnek morskich, co 3-6 lat
występują bardzo licznie w tundrze Arktyki. W
ciągu jednego lata samica może urodzić nawet do
78 młodych! Gdy brakuje pokarmu, zaczynają się
masowe wędrówki migracyjne w poszukiwaniu
lepszych miejsc bytowania. Większość ze zwierząt
ginie np. podczas próby pokonania jeziora czy
rzek. Nieprawdziwe są jednak opowieści o
samobójczych skokach lemingów do wody z
wysokich skał! Lemingi w okresach masowych
pojawów padają ofiara wielu drapieżców, dla
których
stanowią
podstawowy
pokarm,
np.
wydrzyków, sów śnieżnych, lisów polarnych i łasic
Sowa śnieżna
Lis polarny

Są jeszcze inne cechy różniące osobniki w
obrębie populacji, np.
kolor
. Tak więc
populacja jest zbiorem różniących się
osobników, a jednocześnie może być
traktowana jako zbiór przeciętnych
osobników, gdy obliczamy np. jej
zagęszczenie.
Struktura wiekowa i
wielkościowa
Ma wpływ na przebieg wielu procesów. Osobniki starsze są
z reguły większe, a w efekcie składają więcej ikry (ryby) lub
wydają więcej nasion (rośliny). U ssaków osobniki młode są
podatne na choroby, na które nie zapadają osobniki
dorosłe.
Wiek i wielkość
są ważne u wszystkich
gatunków, u których wykształcona jest
organizacja socjalna, gdyż ma w to wpływ na
pozycję socjalną osobnika.
Wielkość
jest cechą szczególnie ważną u organizmów
występujących w formie struktur wieloosobniczych.
Należy tu wiele roślin rozmnażających się przez
odrosty, np. trawy, oraz wiele zwierząt z niższych grup
systematycznych, np. gąbki, koralowce, mszywioły.
Wielkość struktur określa ich rolę ekologiczną.
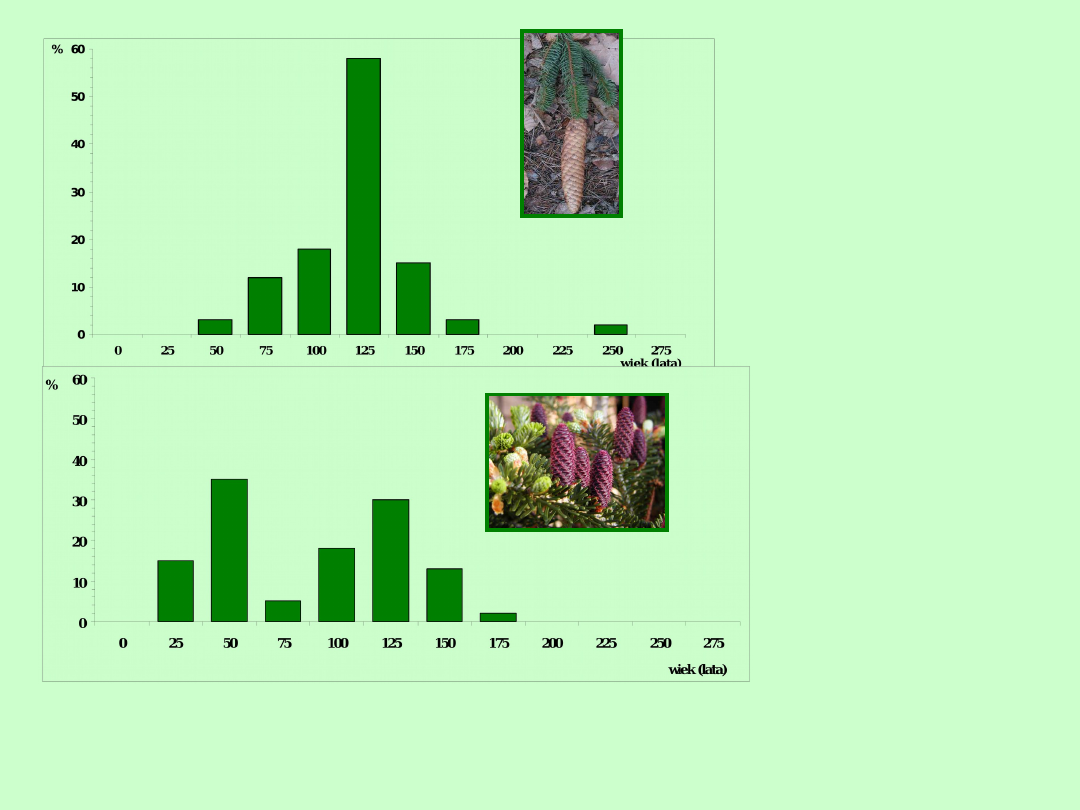
Świerk
Jodła
Struktura wiekowa świerka i jodły strefy subalpejskiej Gór Skalistych jest
nietypowa. Niektóre klasy wieku są znacznie liczniejsze niż inne; są to
struktury dominujące. Taka sytacja dotyczy gatunków o osobnikach
długowiecznych, np. ryb, drzew. W takich populacjach struktura wiekowa
może się zmieniać znacznie z roku na rok
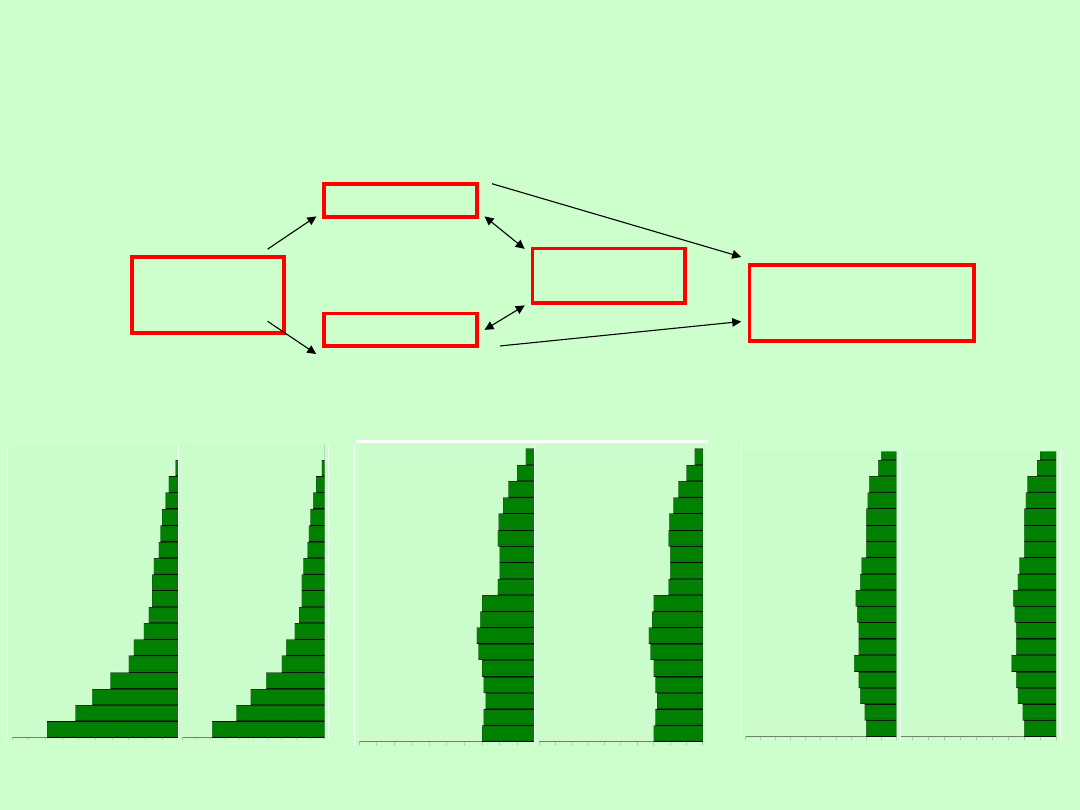
Informacja o strukturze wiekowej populacji może być użyta do określenia
jej stanu. Populacje będące w fazie wzrostu na ogół charakteryzują się
liczebną dominacją osobników młodych. Taka sytuacja była (i nadal jest) w
Kenii, gdzie tempo wzrostu wynosiło w 1991 r. 3,8% rocznie , a średnia
oczekiwana długość życia – 63 lata. W USA tempo wzrostu wynosiło 0,8%
rocznie, a średnia oczekiwana długość życia – 75 lat.
10 8 6 4 2 0 2 4
6 8 10
10 8 6 4 2 0 2 4
6 8 10
10 8 6 4 2 0 2 4 6
8 10
Populacja o
szybkim wzroście
(Kenia)
Populacja o
powolnym
wzroście (USA)
Brak wzrostu
populacji
(Dania)
% w populacji
85+
80-84
75-79
70-74
65-69
60-64
55-59
50-54
45-49
40-44
35-39
30-34
25-29
20-24
15-19
10-14
5-9
0-4
Przed 1905
1905-1910
1910-1915
1915-1920
1920-1925
1925-1930
1930-1935
1935-1940
1940-1945
1945-1950
1950-1955
1955-1960
1960-1965
1965-1970
1970-1975
1975-1980
1980-1985
1985-1990
Kobiety
Kobiety
Kobiety
Mężczyźni
Mężczyźni
Mężczyźni
Czynniki
środowiskow
e
śmiertelność
rozrodczość
Struktura
wiekowa
Tempo wzrostu lub
spadku liczebności
populacji

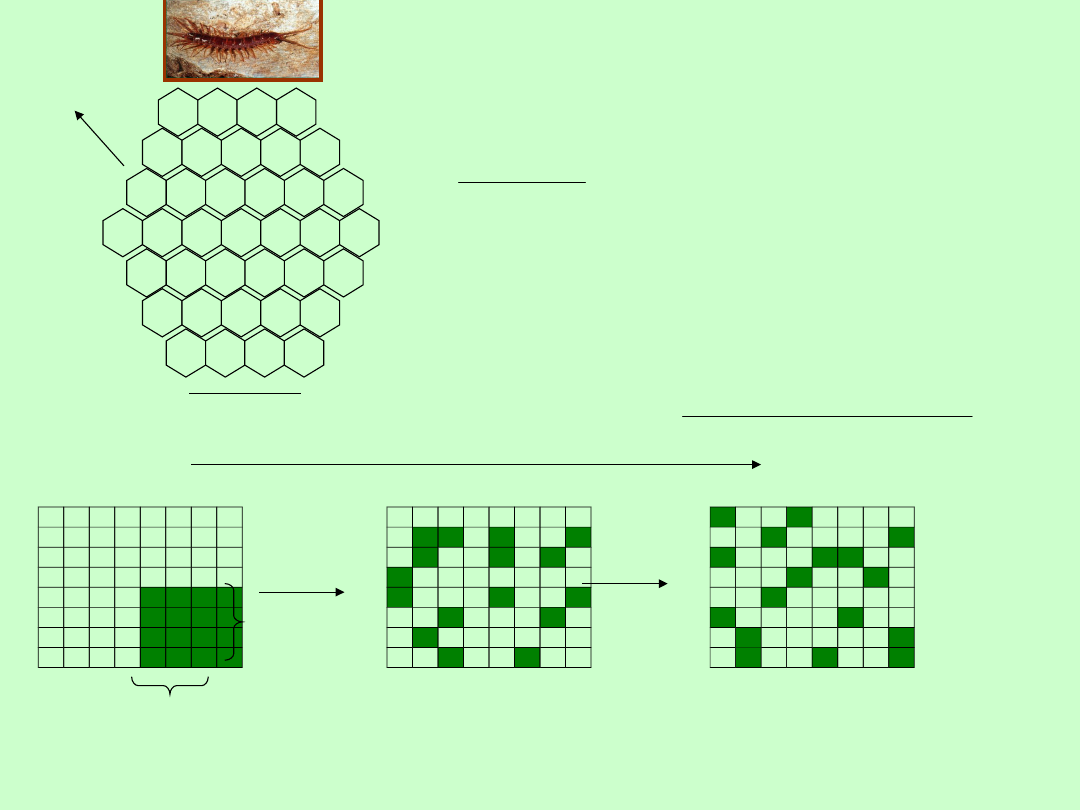
Metody oceny zagęszczenia bezwzględnego
Metody oceny zagęszczenia bezwzględnego
1. Liczenia bezpośrednie, np. powszechny spis ludności, liczenie drzew w lesie, fok podczas
rozrodu, osiadłych bezkręgowców
2. Pobieranie prób z populacji
a)
Metoda kwadratów
- policzenie wszystkich osobników na określonej powierzchni;
konieczna dokładność oceny i losowy dobór kwadratów do badań; metoda szeroko
stosowana przy ocenie zagęszczenia bezkręgowców glebowych i roślin
b)
Metoda znakowania i ponownych złowień, tzw. metoda Petersena
- polega na łowieniu
osobników, indywidualnym ich znakowaniu, wypuszczaniu i ponownym łowieniu.
Stosowana do oceny zagęszczenia zwierząt ruchliwych. Metoda jest znacznie prostsza
do zastosowania w przypadku populacji zamkniętej (jej wielkość nie zmienia się między
pierwszym a drugim połowem) niż otwartej (liczebność zmienia się w czasie trwania
badań).
W tej metodzie powinny być spełnione 3 podstawowe założenia:
•
zwierzęta znakowane i nie znakowane łowią się przypadkowo, to znaczy z takim samym
prawdopodobieństwem
•
śmiertelność osobników znakowanych jest taka sama jak nie znakowanych [ istocie
metoda Petersena zakłada brak śmiertelności osobników podczas badań]
•
oznakowanie osobników jest trwale i wyraźne
[ problemów: niektóre myszy polne przyzwyczajają się do pułapek, inne - unikają ich, ryby
mogą być
zranione podczas połowów siecią, długo żyjące ptaki mogą gubić obrączki itp.]
Metoda Petersena
powtarzana wielokrotnie często
służy
nie tylko
do oceny zagęszczenia
, ale
także
dynamiki liczebności w czasie
,
wskaźnika śmiertelności (ubywania) i wskaźnika rozrodczości
(przybywania).
0
2
0
1
1
2
1
0
0
0
0
2
0
1
0
2
2
0
0
2
0
0
1
1
0
3
1
0
0
3
1
0
2
1
0
0
1
1 m
N
Lloyd oceniał zagęszczenie wijów w
37 „kwadratach” w lesie bukowym w
środkowej Anglii. Wyniki jakie
uzyskał, są przedstawione na
rysunku. Średnie zagęszczenie
wynosi więc:
kwadracie
w
ika
0,811osobn
kwadratów
37
osobników
30
lub, ponieważ powierzchnia każdego
„kwadratu wynosi 0,08 m
2
zagęszczenie wijów można ocenić
jako10,1 osobnika na m
2
Osobniki
złowione i
wyznakowane m
= 16
Mieszanie się
osobników
znakowanych i
nieznakowanych
liczba złowionych
osobników = 20
Każdy kwadracik reprezentuje jednego
osobnika
Połów pierwszy
Połów drugi
populacji
h w
znakowanyc
Udział
h
znakowanyc
osobników
Liczba
populacji
liczebnosc
Oceniana
Metoda znakowania i ponownych złowień, znana
jako metoda Petersena lub wśród ekologów
terenowych metoda Lincolna
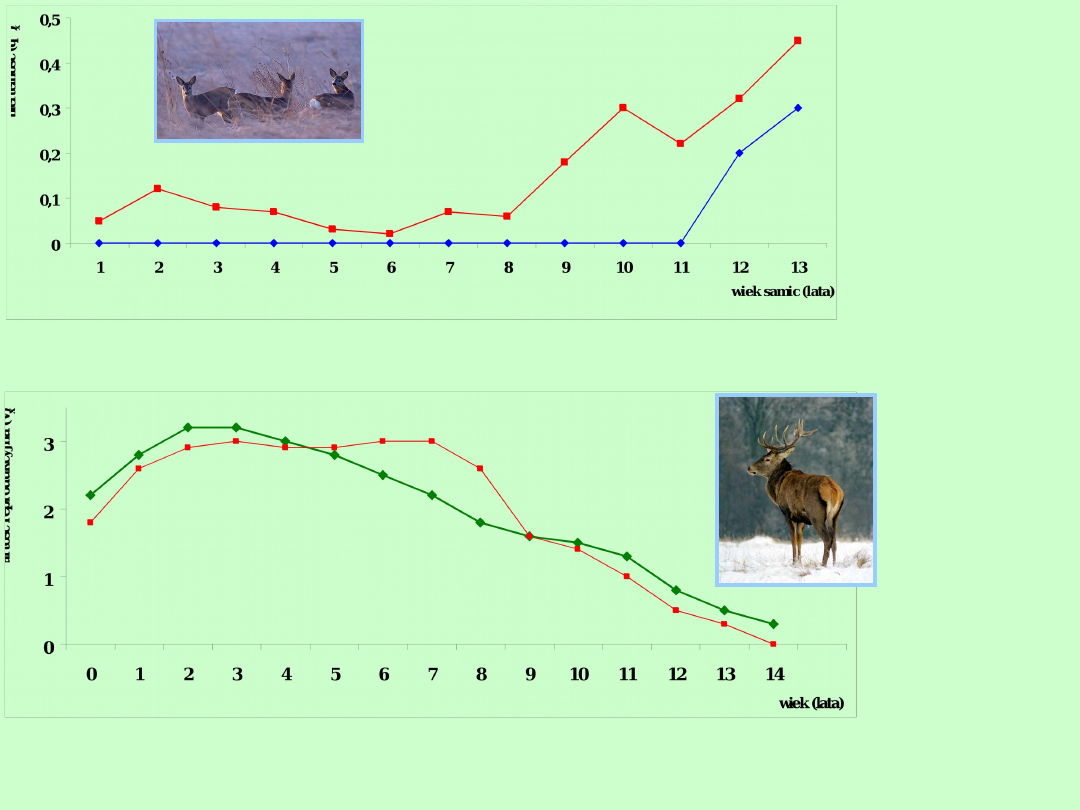
Rozród związany jest z określonymi korzyściami biologicznymi, ale także z
kosztami. Polegają one na obniżeniu tempa wzrostu lub spadku
przeżywalności, co jest konsekwencją przeznaczenia większej ilości energii na
reprodukcję. Szczególnie laktacja u samic jest kosztowna energetycznie
Samice
biorące udział
w rozrodzie
Samice nie
uczestniczące
w rozrodzie
Samce jeleni żyjących w Szkocji bronią swoich haremów przy pomocy poroża i
najefektowniejszy okres przypada między 6 a 11 rokiem życia. Samice jeleni
(łanie) rodzą młode od 3 do 15 roku życia i żyją dłużej niż samce. Dlatego
krzywe wartości reprodukcyjnej samic i samców są różne
byki
łanie

Metody oceny zagęszczenia
Metody oceny zagęszczenia
względnego
względnego
Metody te służą do obliczania mniej lub bardziej
dokładnych wskaźników liczebności rzeczywistej i do
porównywania zagęszczenia populacji na dwóch lub
więcej terenach. Nie można jednak na ich podstawie
twierdzić, że zagęszczenie na terenie x jest np.
dwukrotnie większe niż na terenie y.
Metoda pułapkowa, stosowana głównie do oceny liczebności gryzoni i owadów
Metoda liczenia odchodów stosowna do oceny liczebności zajęcy, jeleni, myszy
Metoda liczenia głosów (w odniesieniu do ptaków), np. liczba krzyków bażantów
słyszanych w ciągu 15 minut wczesnym rankiem)
Liczenie zwierząt upolowanych przez myśliwych, np. liczba skór skupowanych w
Ameryce Płn.
Ocena wielkości połowów, np. liczba ryb złowionych w ciągu 100 godz. trałowania.
Metoda liczenia różnych śladów pozostawianych przez zwierzęta, np. liczenie
wypluwek ptaków drapieżnych, gniazd wiewiórek na drzewach czy też nor
wygrzebanych w mule przez raki

Metoda ankietowa - ankiety wysyłane są do myśliwych oceniających
subiektywnie zmiany liczebności; przydatna w ocenie głębokich zmian
Iiczebności dużych zwierząt.
Metoda oceny stopnia pokrycia podłoża - % powierzchni gleby pokryty
przez rośliny lub % pokrycia skalistego dna morskiego przez osiadłe
bezkręgowce, np. małże.
Metoda oparta na ocenie ilości zjedzonej przynęty, np. do oceny liczebności
myszy i szczurów.
Liczenie wzdłuż określonej trasy, np. jadąc samochodem można liczyć
wszystkie widziane po drodze ptaki drapieżne.
Metody oceny zagęszczenia względnego są - użyteczne
jako uzupełnienie metod bezpośrednich oraz przy ocenie
głębokich zmian liczebności populacji.
Dokładne policzenie wszystkich osobników populacji jest
możliwe bardzo rzadko
, najczęściej określa się tyko rząd
wielkości liczebności populacji.
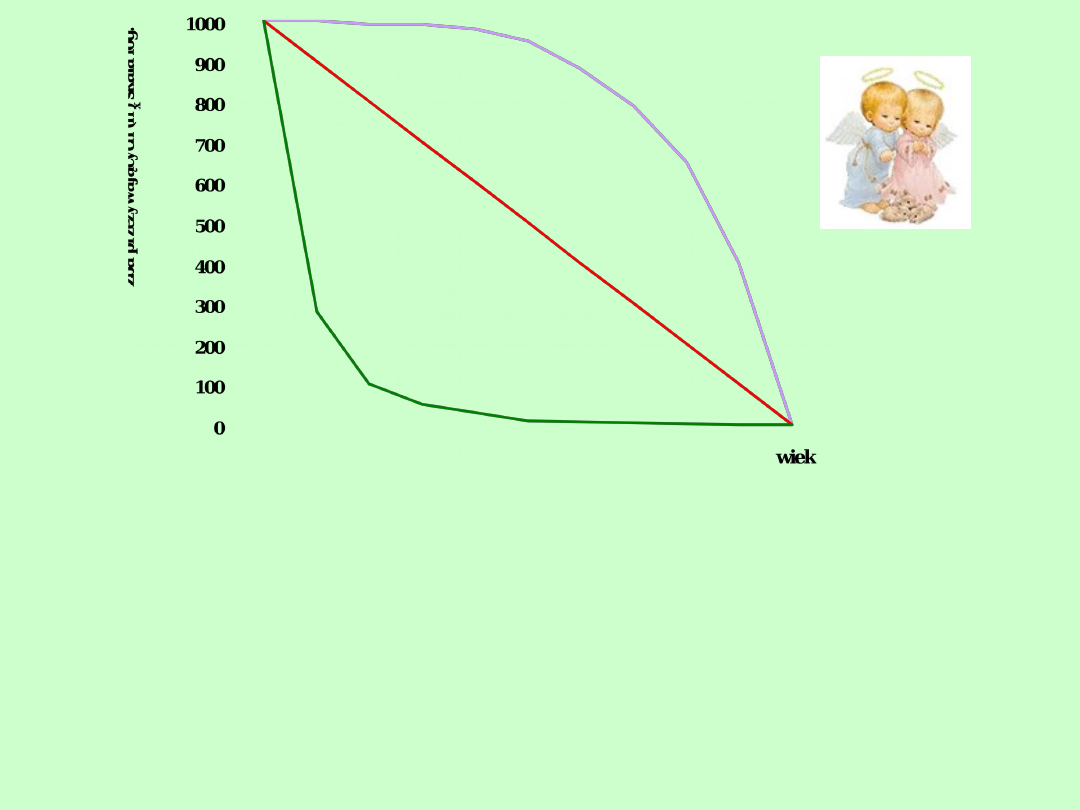
Typ I
Typ II
Typ III
Podstawowe typy krzywych przeżywania wg Pearla (1928)
1.Krzywa przezywania Typu I
Jest charakterystyczna dla populacji w których śmiertelność jest niewielka, przez
większą część życia, a wzrasta znacznie w klasach osobników najstarszych
2.
Krzywa przeżywania Typu II
Wykazuje na stałe tempo ubywania (śmiertelność) osobników, niezależnie od wieku.
3. Krzywa przezywania typu III
Dotyczy populacji w których śmiertelność osobników młodych w pierwszych okresach
życia Jest największa, zaś w starszych klasach wiekowych śmiertelność jest niewielka i
względnie stała (niezależna od wieku)
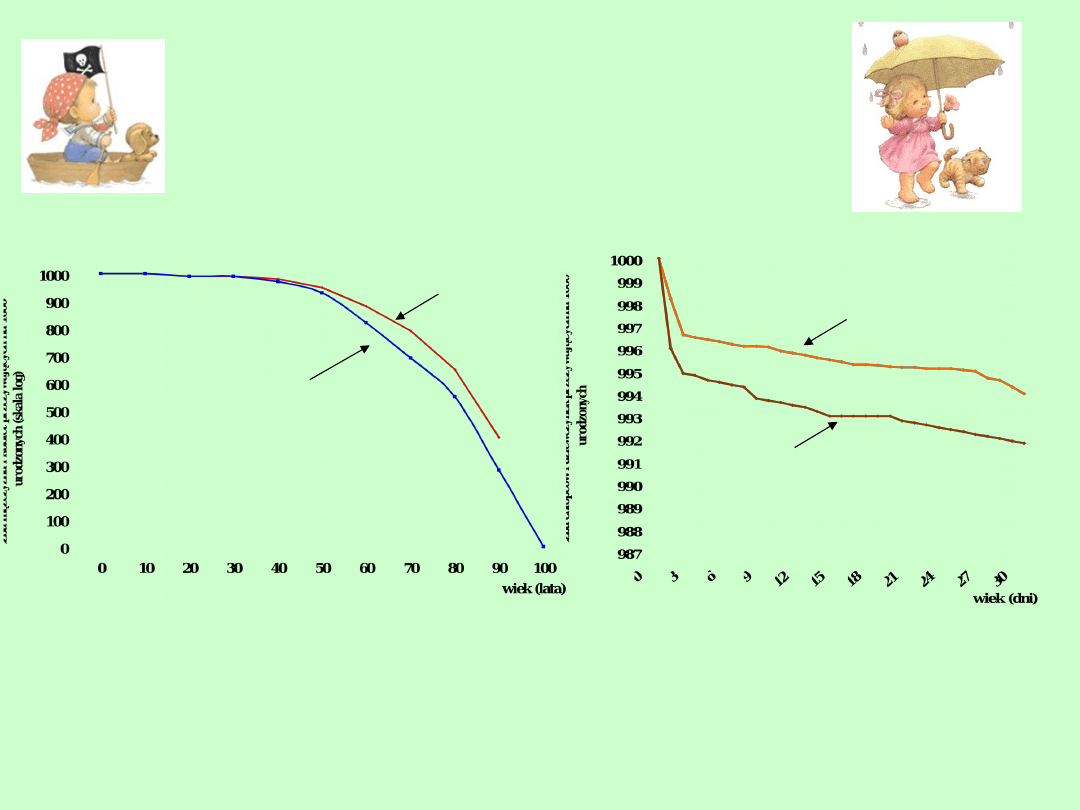
a.
Krzywa
przeżywalność
ludności
USA
z
uwzględnieniem płci.
b. Krzywa przeżywalności noworodków w pierwszym
miesiącu życia
a
kobiety
mężczyźni
b
dziewczynki
chłopcy
Rzeczywiste krzywe przeżywania zawsze nieznacznie różnią
się od teoretycznych. Na przykład krzywe przeżywania
populacji ludzkich w krajach wysoko rozwiniętych są zbliżone
do krzywej typu I. Krzywe typu II są charakterystyczne dla
wielu ptaków, natomiast typ III dotyczy wielu gatunków ryb,
bezkręgowców morskich a także pasożytów
Ważna zasada demograficzna
W populacji o stałej rozrodczości i śmiertelności osobników (w
określonym wieku):
1. liczebność wzrasta zgodnie z krzywą wykładniczą, stosownie do wartości wrodzonego
tempa wzrostu,
2. ustala się określona stała struktura wiekowa,
3. struktura wiekowa jest stałą i może pozostawać bez zmian przez dowolnie długi
okres.
Struktura wiekowa populacji nie zmienia się tak długo, jak długo pozostają nie
zmienione jej tabele pożywienia i płodności
Populacje o wzroście wykładniczym osiągają stałą strukturę wiekową, zaś te, które
okresowo utrzymują stałą liczebność, cechują się niezmienną strukturą wiekową.
W populacjach naturalnych struktura wiekowa populacji jest zwykle zmienna w czasie.
Stała struktura
wiekowa
populacji
gwałtowny wzrost
liczebności populacji
Niezmienna struktura wiekowa
liczebność populacji stała
Jest to struktura
wiekowa populacji o
wykładniczym wzroście
liczebności
Jest to struktura
hipotetyczna i pokazuje,
jaka byłaby struktura
wiekowa populacji o
określonym wskaźniku
rozrodczości, gdyby ten
wskaźnik śmiertelności by
idealnie taki sam
Nornik
zwyczajny
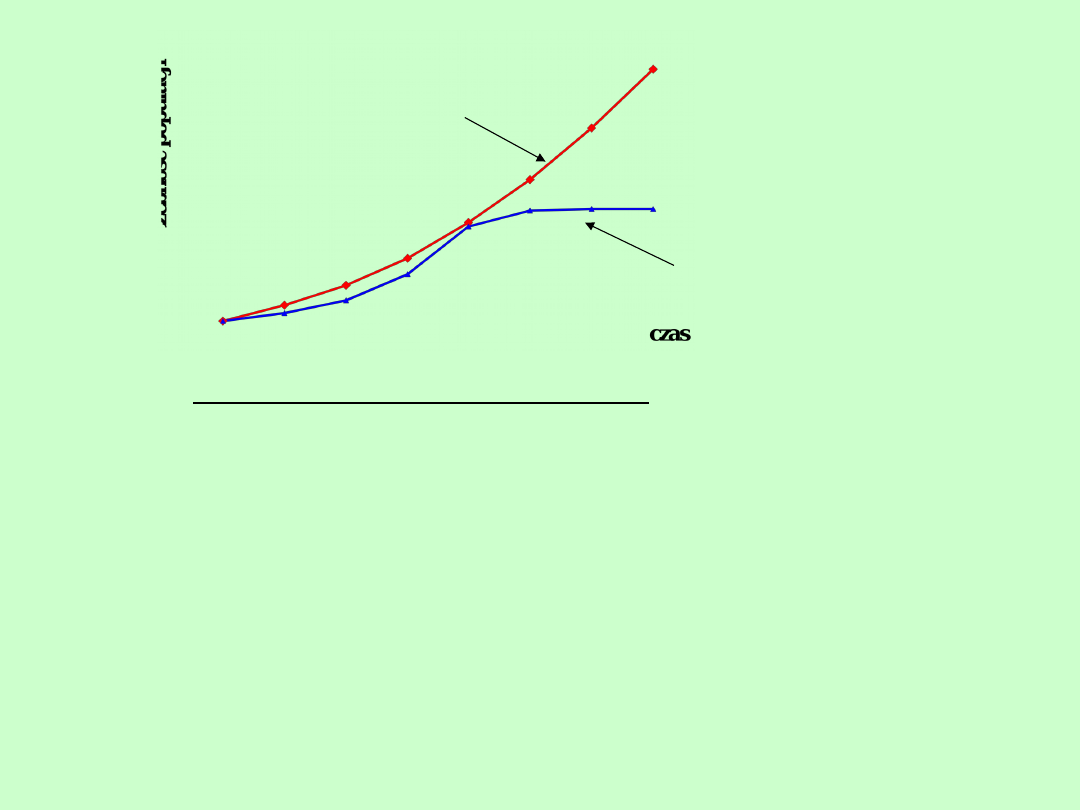
Wzrost
logistyczny
Wzrost
wykładniczy
Populacje o pokoleniach ciągłych:
1.Tempo wzrostu jest stałe
Powstaje krzywa wykładniczego wzrostu
nieograniczonego
2. Tempo wzrostu jest zależne od zagęszczenia
Powstaje krzywa logistycznego wzrostu
ograniczonego przez pojemność środowiska
Krzywa logistyczna = krzywa esowata. Ma ona
górną asymptotę, do której zmierza liczebność
populacji
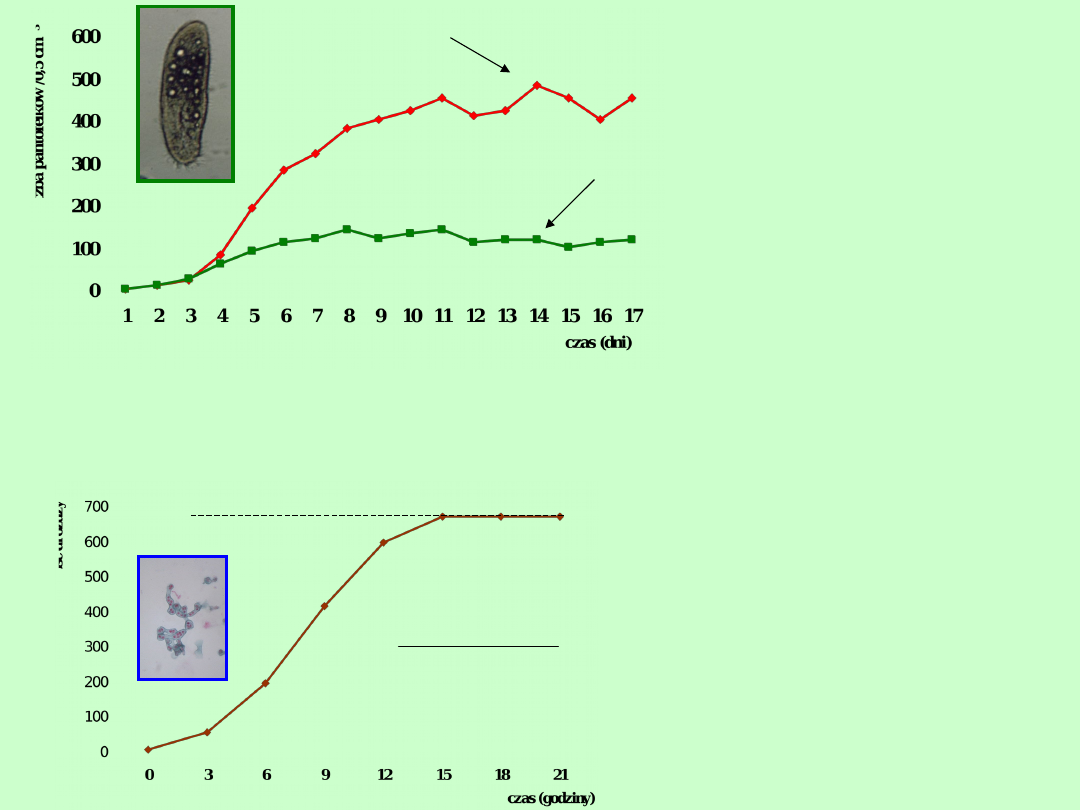
Paramecium
aurelia
Paramecium
caudatum
Wzrost liczebności populacji pantofelków Paramecium aurelia i
Paramecium caudatum w temp. 26
o
C w zbuforowanym roztworze
Osterhouta przy pH = 8,0 i stałym dopływie bakterii jako
pożywienia (dane z: Gause 1934)
Wzrost populacji drożdży
(za: Pearl 1927; dane z:
Carlson 1913
r
0,54
4,19
1
665
e
N
K = 665
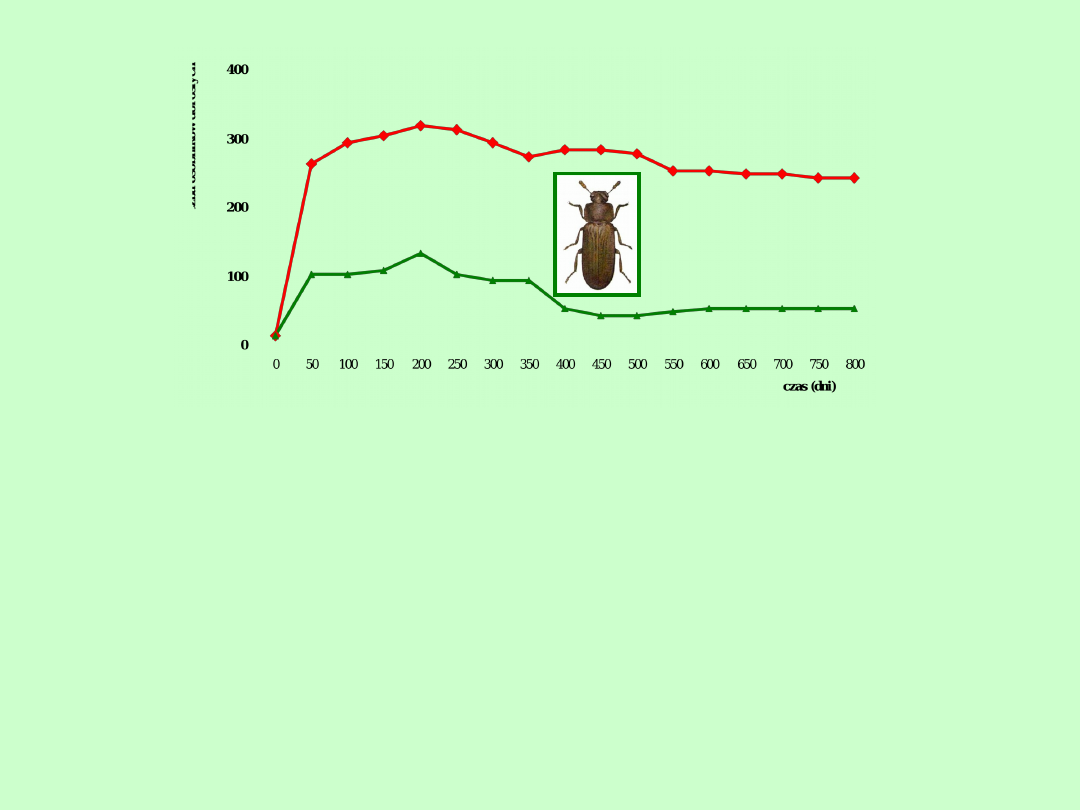
Wyniki doświadczalne często wskazują dużą zgodność
z krzywą logistyczną. Jednak górna asymptota
populacji może się różnić w zależności od wrodzonego
tempa reprodukcji (np. pantofelki w eksperymencie
Gausego). Ponadto liczebność populacji po osiągnięciu
poziomu pojemności środowiska może wykazywać
spadek (np. trojszyki w doświadczeniu Parka
linia c
I
linia c IV
Wzrost liczebności populacji trojszyka Tribolium castaneum w
hodowli w 8 g mąki w temp. 29
o
C I wilgotności względnej 70%.
Widoczne są znaczne różnice w przebiegu wzrostu populacji
różnych linii genetycznych u tego gatunku
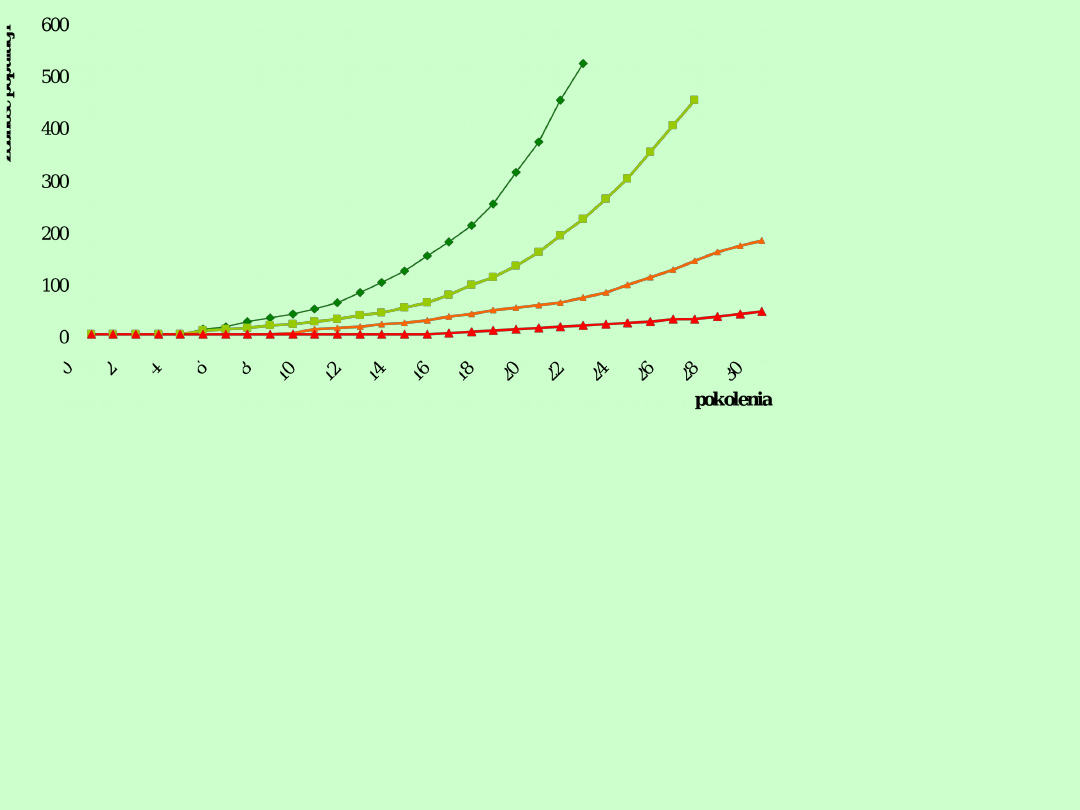
Populacje o pokoleniach nieciągłych
1.Tempo wzrostu jest stałe
Jeżeli gatunek rozmnaża się tylko raz w
roku, a tempo wzrostu jest stałe, to wzrost
populacji
będzie
wykładniczy.
Tempo
wzrostu będzie w tym przypadku zależne
od tempa reprodukcji netto, czyli od liczby
potomstwa płci żeńskiej dożywającego do
sezonu rozrodczego w roku następnym.
R
o
=1,2
0
R
o
=1,1
5
R
o
=1,1
0
R
o
=1,0
5
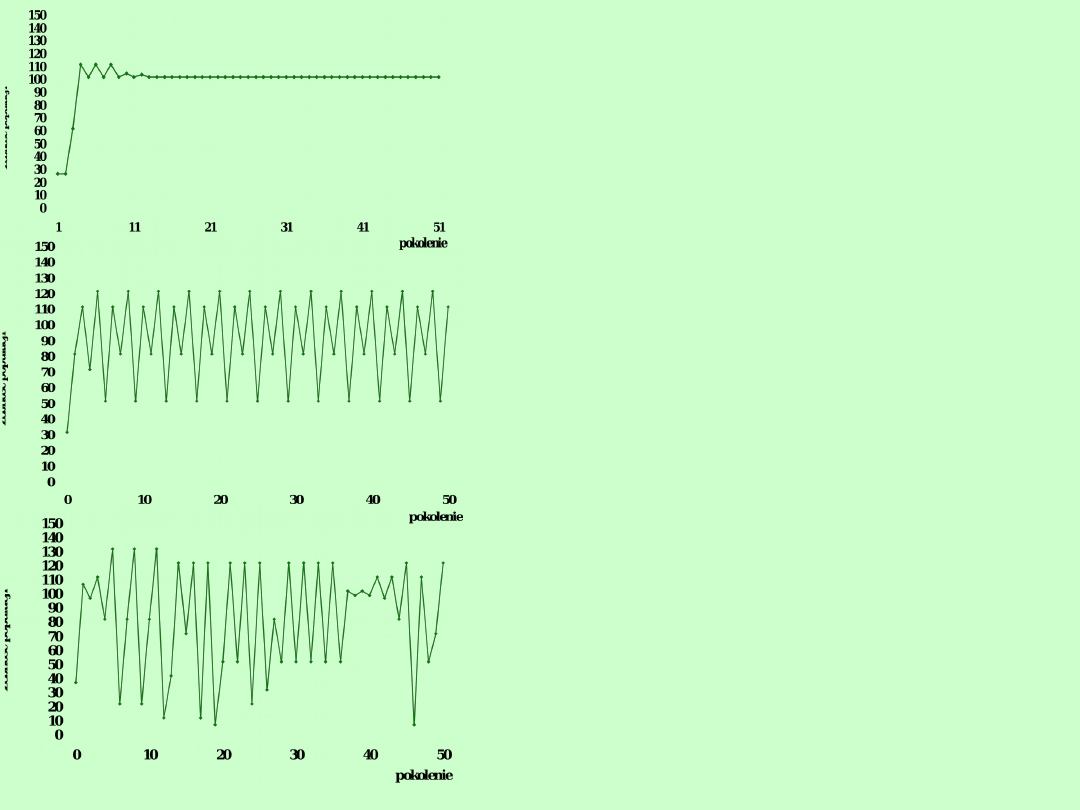
2. Tempo wzrostu jest zależne od
zagęszczenia
Przy
dużych
zagęszczeniach
rozrodczość spada, a wzrasta
śmiertelność (epidemie, brak
pokarmu). Fluktuacje liczebności
zależne są od korelacji między
zagęszczeniem
populacji
i
tempem jej wzrostu.
Zagęszczenie
zrównoważone
oznacza
punkt
w
którym
śmiertelność
równoważy
rozrodczość.
Im
bardziej
faktyczne
zagęszczenie populacji odbiega
od
zagęszczenia
zrównoważonego, tym większe
będą oscylacje liczebności przed
osiągnięciem zrównoważenia.
Przy dużej różnicy mamy do
czynienia
z
chaotycznymi
fluktuacjami
o
charakterze
przypadkowym
R=0,0
18
R=0,0
25
R=0,0
29
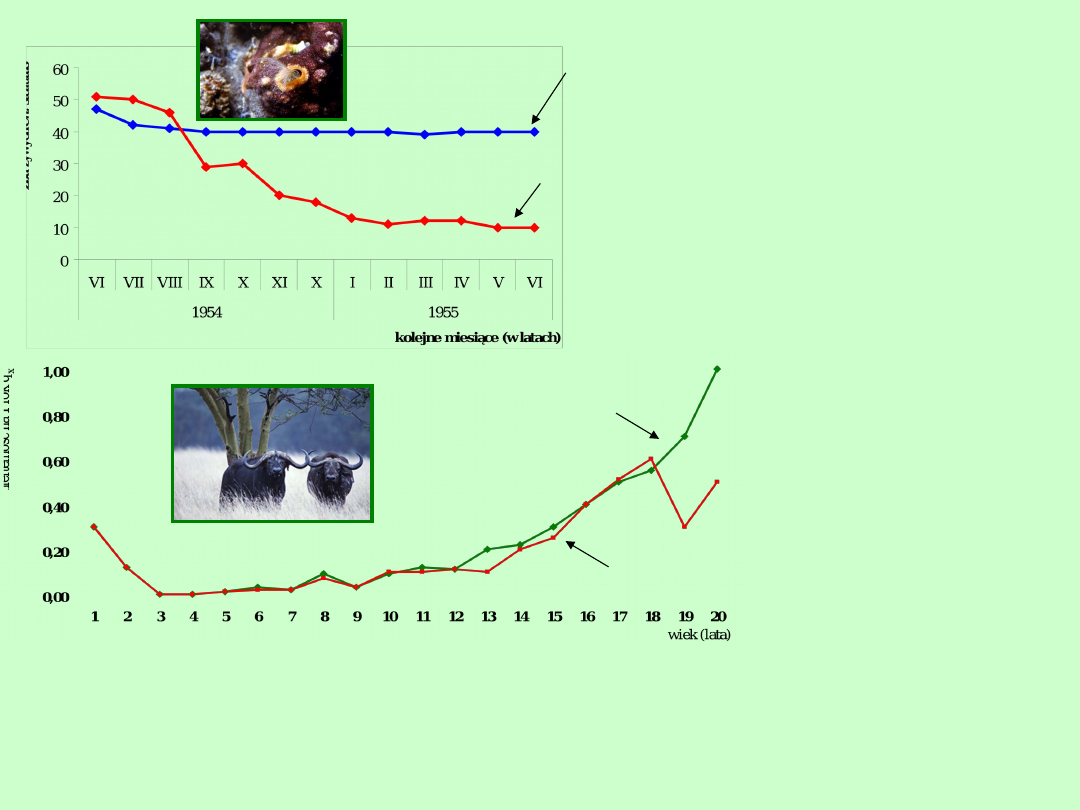
Przykład bezpośredniej obserwacji
przeżywalności
Pąkle Chtalamus stellatus i Balanus
balanoides przytwierdzone do skał
podwodnych w Szkocji konkurują ze
sobą.
Osobniki
Chtalamus
stellatus
przeżywały
znacznie
lepiej,
gdy
doświadczalnie
usunięto
Balanus
balanoides
(gatunek
w
naturze
dominujący)
Stanowiska z których
usunięto Balanus
balanoides
Stanowiska z których nie
usunięto Balanus
balanoides
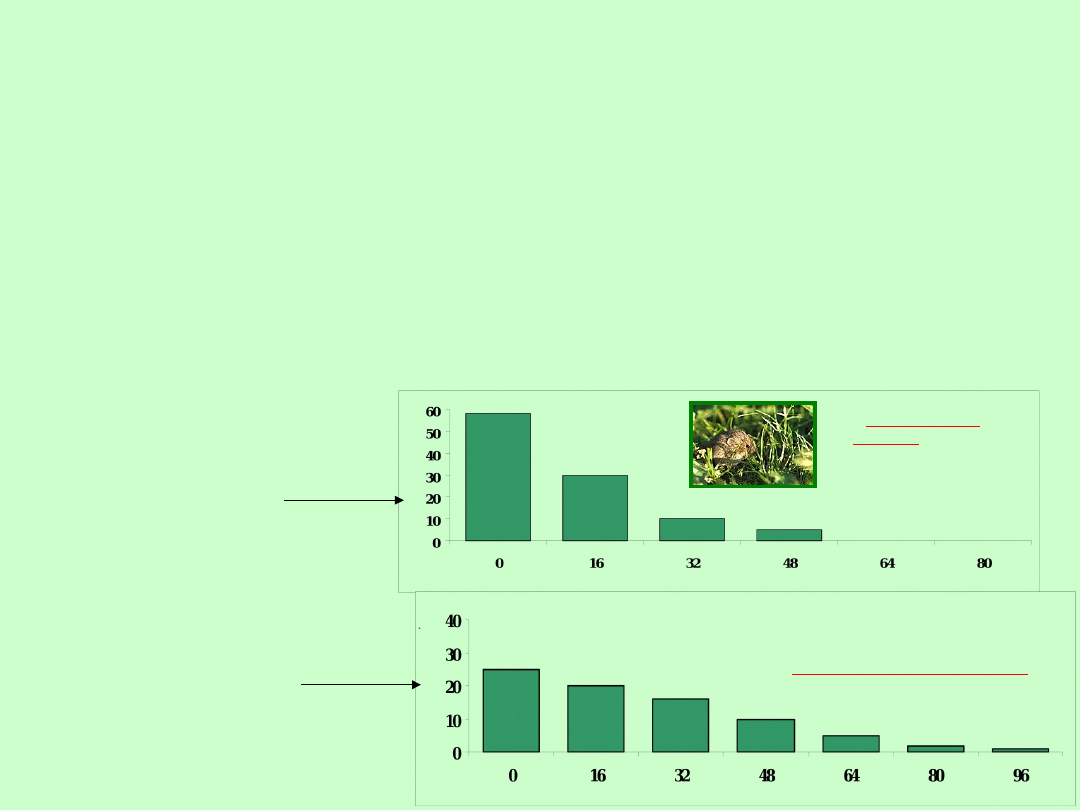
Na obszarze Parku Narodowego Serengeti w Afryce Wschodniej zbierano
wszystkie czaszki bawołu afrykańskiego (Syncerus caffer), które umierały w
sposób naturalny. Wiek określano na podstawie rocznych pierścieni przyrostów
rogu. Ubywanie osobników młodych (do lat 2) badano bezpośrednio w składzie
Przykład
przeżywalności na
podstawie określania
wieku osobników
umierających
samice
samce
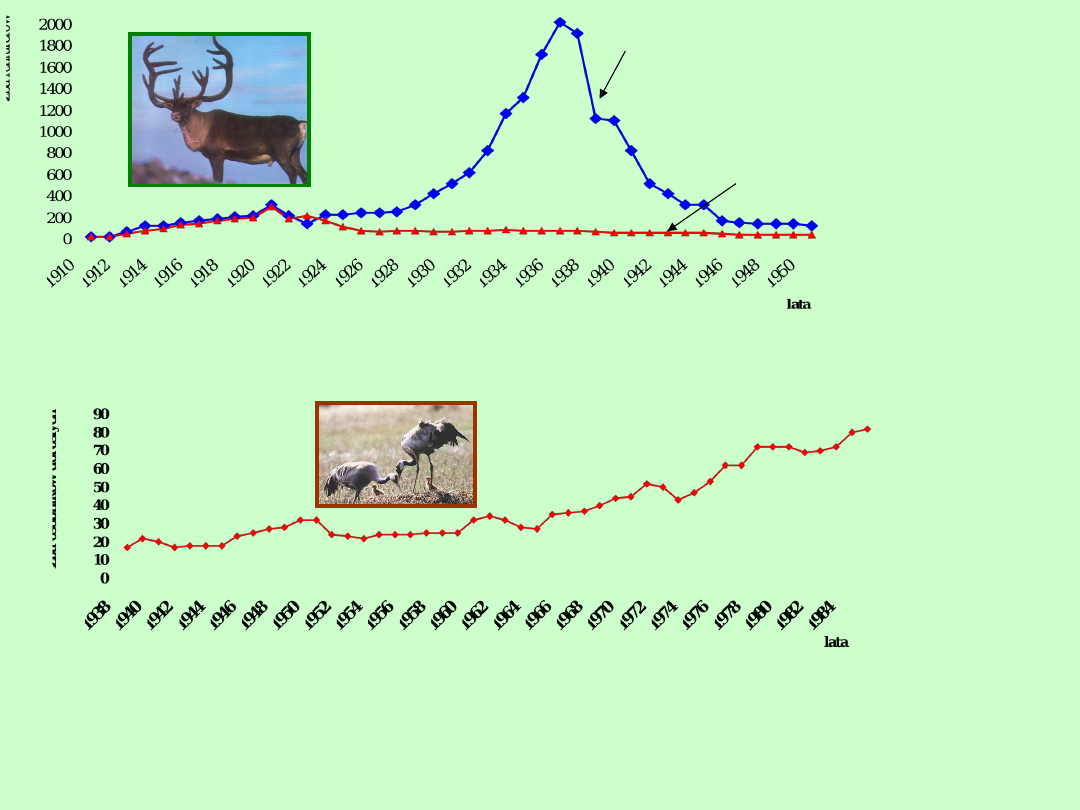
Wyspy St. Paul
Wyspy St.
George
W roku 1911 wprowadzono renifery na dwie wyspy na Morzu Berlinga, blisko
wybrzeży Alaski, jednak dalszy wzrost populacji przebiegał zupełnie inaczej na każdej
z wysp, pomimo, że warunki ekologiczne były bardzo podobne na obu wyspach
Żuraw krzykliwy (Grus americana) jest dobrym przykładem odnawiania się
gatunku niegdyś zagrożonego wyginięciem . Zaczęto go chronić w roku
1916, jednak dopiero po roku 1956 liczebność populacji powiększa się
szybciej. Obserwuję się cykliczność wzrostu spowodowaną być może
cyklicznie zmienną presją drapieżników na terenach lęgowych żurawi w
kanadyjskim Parku Narodowym Wood Bufflo. Na zimę ptaki migrują do
Teksasu
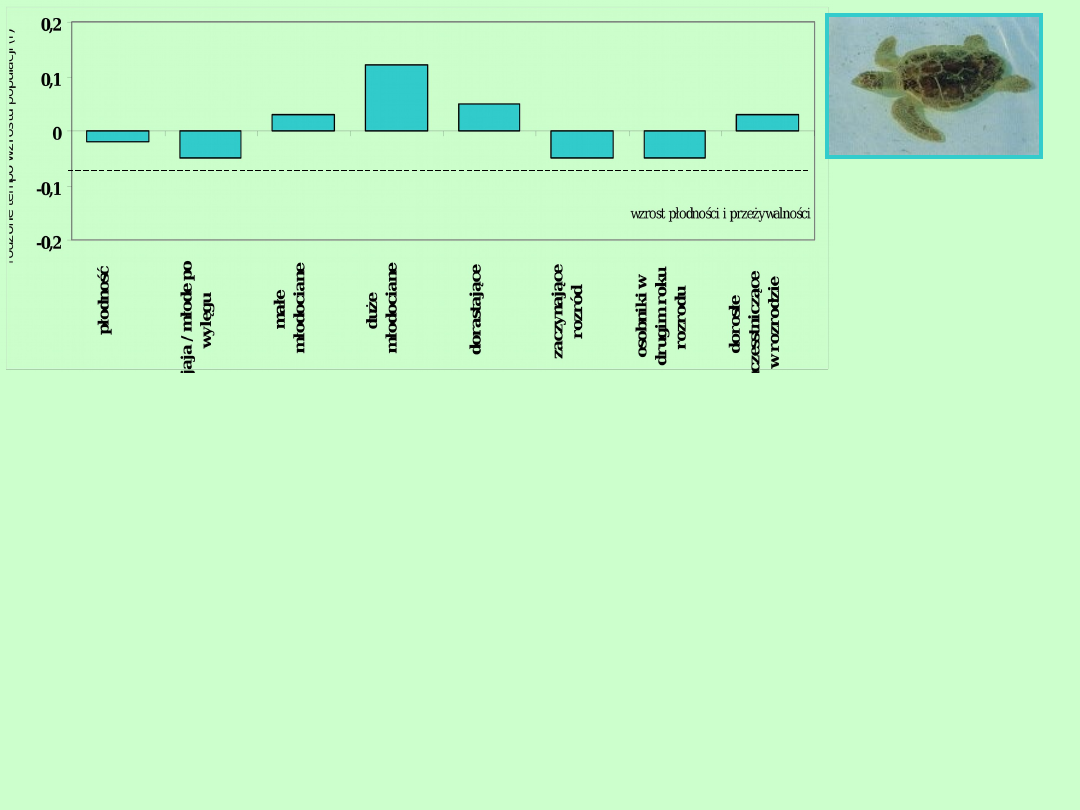
Żółw Karetta
Stan normalny
Zastosowanie teorii populacji do celów ochrony
gatunkowej
Żółw Karetta zagrożony wyginięciem żółw morski występujący w Oceanie
Atlantyckim wzdłuż południowo- wschodnich wybrzeży USA. Na
podstawie modelu macierzowego dla cyklu życiowego żółwia przebadano
różne sposoby zatrzymania spadku liczebności tego gatunku. Rycina
pokazuje skutki podwyższenie płodności o 50% lub podniesienia
przeżywalności na każdym etapie cyklu życiowego. Najlepszy efekt
uzyskuje się podnosząc przeżywalność osobników młodocianych.
Obecnie największą wagę przykłada się do ochrony jaj składanych przez
żółwie na plaży, jednak nawet po 30 latach ochrony gniazd żółwi nie
obserwuje się żadnego wzrostu populacji. Należy zadbać, by zwiększała
się przeżywalność żółwi młodocianych w morzu. Duża liczba młodych
żółwi topi się złowiona w sieci zastawione do połowów krewetek. Obecnie
więc tak konstruuje się te sieci, by nie wpadały w nie żółwie.
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
- Slide 41
- Slide 42
- Slide 43
- Slide 44
- Slide 45
- Slide 46
- Slide 47
- Slide 48
- Slide 49
- Slide 50
- Slide 51
Wyszukiwarka
Podobne podstrony:
Ekologiczne podstawy systemu ws Nieznany
Ekologiczne podstawy hodoeli lasu, AR Poznań - Leśnictwo, ekologiczne podstawy hodowli lasu
ephl egzamin cwiczenia, Leśnictwo UWM Olsztyn, Semestr IV, Ekologiczne Podstawy Hodowli Lasu, Egzami
Ekologiczne podstawy systemu wspierania rozwoju energetyki odnawialnej, Studia, ekologia
Zestawy na egzamin z EPHL, Leśnictwo UWM Olsztyn, Semestr IV, Ekologiczne Podstawy Hodowli Lasu, Egz
BRZOZA BRODAWKOWATA, AR Poznań - Leśnictwo, ekologiczne podstawy hodowli lasu, Prezentacja Brzoza br
ekologiczne podstawy gospodarki żywymi zasobami EPGZZ cw
poprawka, LEŚNICTWO, III ROK, EKOLOGICZNE PODSTAWY HODOWLI LASU
ekologiczne podstawy gospodarki Nieznany
10 Ekologiczne podstawy ksztaltowania doborów roślinnych
jadczyk,ekologia, Podstawy ekologii krajobrazu
ekologiczne podstawy gospodarki żywymi zasobami - tszydel, Materiały dla studentów, ochrona srodowis
ekologiczne podstawy gospodarki żywymi zasobami epgzz
ekologiczne podstawy gospodarki żywymi zasobami AAA
ekologiczne podstawy gospodarki żywymi zasobami wykladOS
ekologiczne podstawy gospodarki żywymi zasobami zagadnienia opracowane
poprawka, Leśnictwo UWM Olsztyn, Semestr IV, Ekologiczne Podstawy Hodowli Lasu, Egzamin i kolokwia
więcej podobnych podstron