
BYDŁO
WETERYNARIA W TERENIE
y
1/2009
y
17
A
dr n. wet. Grzegorz J. Dejneka, prof. dr hab. Jan Twardoń, lek. wet. Joanna Polańska
Katedra i Klinika Rozrodu, Chorób Przeżuwaczy oraz Ochrony Zdrowia Zwierząt Uniwersytetu Przyrodniczego we Wrocławiu
Aspekty fi zjologiczne
Aktywność rozrodcza
bydła mięsnego
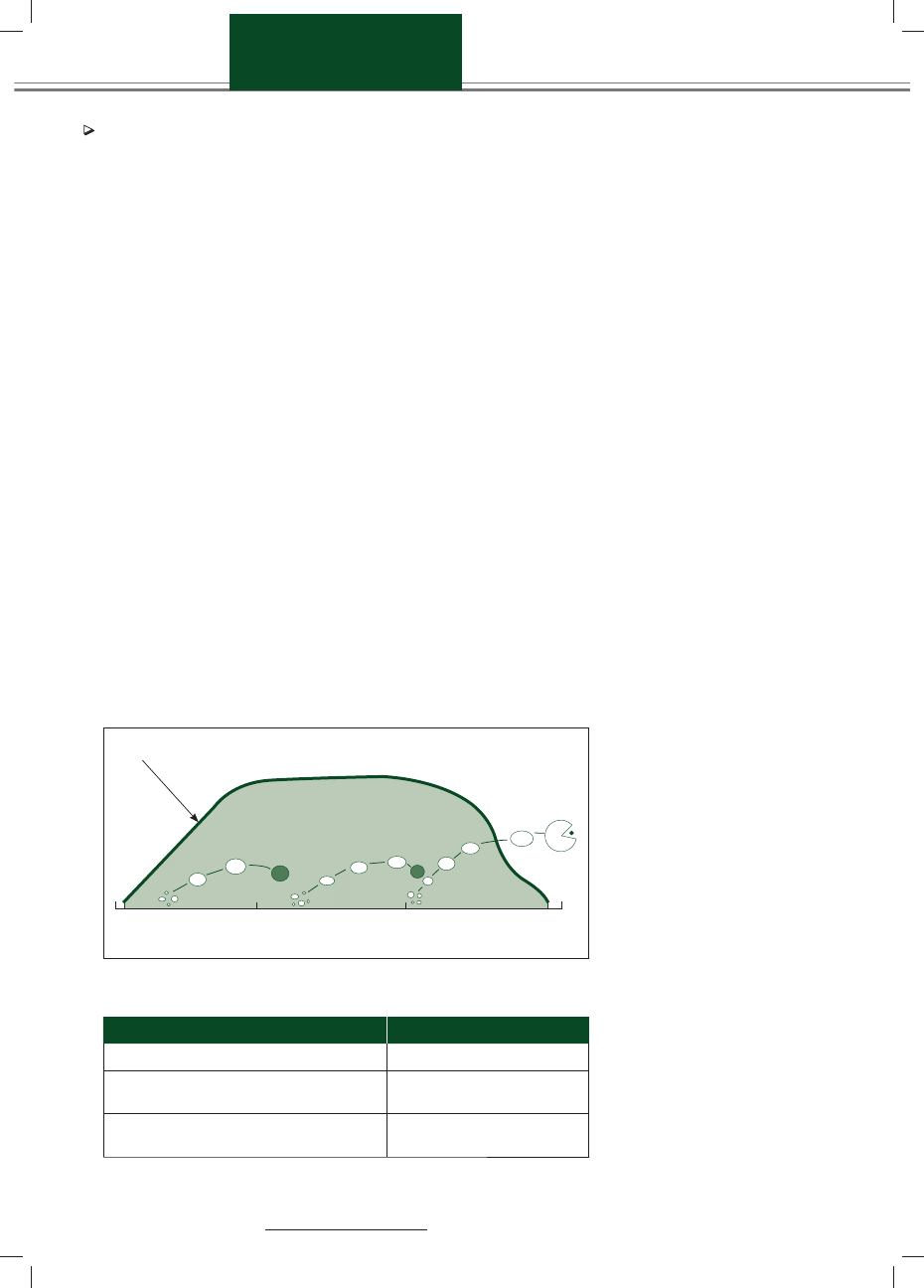
Istnieją 2 lub częściej 3 fale wzrostu pę-
cherzyków, przy czym ostatnia z nich
jest falą owulacyjną (ryc. 1).
W każdej fali rośnie grupa kilku, kil-
kunastu pęcherzyków, przy czym jeden
z nich, osiągający największe rozmiary,
nazywa się pęcherzykiem dominują-
cym, podczas gdy pozostałe określane
są mianem pęcherzyków podrzędnych,
a ich wzrost jest ograniczany właśnie
przez pęcherzyk dominujący (Steven-
son, 2007; Noakes, 1997). Wzrastające
pęcherzyki przechodzą przez fazę re-
krutacji, selekcji i różnicowania (Mihm
i Bleach, 2003). Dominujący pęcherzyk
fali owulacyjnej jest tym, który podle-
ga jajeczkowaniu (owulacji), przy czym
jego średnica jest większa niż średnica
osiągana przez pęcherzyk dominujący
fali nieowulacyjnej. Brak owulacji pę-
cherzyka dominującego pierwszej fali
(w przypadku wzrostu dwufalowego)
lub pierwszej i drugiej fali (w przypadku
wzrostu trójfalowego) wynika z hamują-
cego wpływu progesteronu produkowa-
nego przez ciałko żółte okresowe (corpus
luteum periodicum), przy czym wpływ
ten kończy się w związku z luteolizą.
Rozwój pęcherzyków jajnikowych uwa-
runkowany jest działaniem hormonów
gonadotropowych, tj. FSH i LH (ich wy-
dzielanie przez przysadkę stymulują go-
nadoliberyny – GnRH), które realizują
swój wpływ na jajnik poprzez wykształ-
cone dla nich receptory (Hartigan, 2004;
Noakes, 1997; Stevenson, 2007).
Brak rui
Brak rui (anoestrus) może mieć cha-
rakter fi zjologiczny lub patologiczny.
Stan ten charakteryzuje się brakiem
cyklicznych zmian czynnościowych
i morfologicznych w obrębie narzą-
du płciowego. Fizjologiczna acyklia
ma miejsce w trzech przypadkach. I tak,
w przypadku zajścia w ciążę cykliczne
zmiany w jajnikach są wstrzymane
(incydentalnie u niektórych ciężarnych
Ruja (oestrus) trwa stosunkowo krótko
– średnio kilkanaście godzin, a sam
okres (interwał) międzyrujowy (a zatem
i cykl jajnikowy, czyli rujowy) wynosi
ok. 21 dni (zakres od 18. dni do 24. dni).
Tak więc zdrowa krowa (lub dojrzała
płciowo jałówka), jeżeli nie jest sku-
tecznie zainseminowana lub pokryta,
„powtarza” ruję mniej więcej co 3 tygo-
dnie (ryc. 1). Owulacja ma miejsce kil-
ka/kilkanaście godzin po zakończeniu
objawów rujowych (Hartigan, 2004; Ste-
venson, 2007).
Falowy wzrost pęcherzyków
Podczas cyklu u krów ma miejsce falo-
wy wzrost pęcherzyków, które osiągają
różne stadia rozwojowe (Noakes, 1997).
Bydło domowe (Bos taurus)
należy do zwierząt poliestral-
nych, czyli takich, które ak-
tywność rozrodczą wykazują
przez cały rok.
fot. Shutters
tock
BYDŁO
WETERYNARIA W TERENIE
y
1/2009
y
18
samic może się pojawić ruja), a obecne
na jednej z gonad ciałko żółte ciążowe
(corpus luteum graviditatis), poprzez
produkcję progesteronu, blokuje aktyw-
ność podwzgórza i przysadki, na zasa-
dzie sprzężenia zwrotnego ujemnego.
Cykliczna aktywność jajników nie wy-
stępuje także u jałowic przed osiągnię-
ciem dojrzałości płciowej oraz u krów
przez pewien czas po wycieleniu, tzw.
poporodowe anoestrus (Hopkins, 1986;
Jaśkowski i Zbylut, 1998).
Dojrzałość płciowa
Dojrzałość płciowa u bydła (deter-
minowana wystąpieniem pierwszej
rui) występuje generalnie pomiędzy
7. a 18. miesiącem życia, kiedy jałów-
ki mleczne osiągają 35% wagi dorosłej
samicy w danej rasie, a jałówki mię-
sne – 40%. U ras mięsnych dojrzałość
hodowlana ma miejsce, gdy jałówka
osiągnie ok. 2/3 masy ciała dorosłej
krowy danej rasy (Wierzbowski i Żu-
kowski, 2007). Należy przy tym pa-
miętać, że istnieją różnice międzyraso-
we – np. hereford czy aberdeen angus
są rasami wcześniej dojrzewającymi
niż charolaise.
Sezon rozrodczy
Opisane wyżej w zarysie aspekty re-
produkcji dotyczą bydła jako gatunku.
W zakresie hodowli, chowu i rozrodu
istnieją jednak pewne znaczące różni-
ce pomiędzy rasami mlecznymi a mię-
snymi. Jedną z nich jest fakt, że chów
bydła mięsnego jest ściśle związany
z wypasem na użytkach zielonych,
które w naszej strefie klimatycznej
są dostępne od maja do listopada.
Inna rozbieżność dotyczy systemu
odchowu cieląt. U bydła mlecznego
cielęta są zwykle zabierane od kro-
wy tuż po urodzeniu – w przeciwień-
stwie do ras mięsnych, u których cie-
lak przebywa dłuższy czas przy matce
i jest przez nią karmiony. Mleko krów
mięsnych jest w całości wykorzysty-
wane przez potomstwo i nie stanowi
surowca skupowego. W stadach by-
dła mięsnego – znacznie rzadziej niż
w przypadku mlecznego – znajduje
zastosowanie sztuczne unasiennianie.
Z reguły ma miejsce krycie haremowe
lub z ręki dokonywane przez buhaje
podczas sezonu rozrodczego.
W stadach bydła mlecznego dąży się
do mniej więcej równomiernego roz-
łożenia wycieleń w ciągu całego roku
– w celu optymalnego wykorzystania
potencjału produkcyjnego krów. Zgo-
ła inna sytuacja jest u bydła mięsnego,
gdzie ze względów ekonomicznych i or-
ganizacyjnych pożądana jest sezono-
wość okresu krycia, a co za tym idzie,
także okresu wycieleń. Powinno się
przy tym dążyć do tego, aby pierwiastki
(jałówki remontowe) cieliły się w wieku
ok. 2 lat, a więc w sezonie wycieleń dla
całego stada. Biorąc pod uwagę prze-
bieg pór roku na półkuli północnej,
korzystne są porody zimowo-wiosen-
ne, dzięki czemu cielęta są odchowy-
wane w ciągu sezonu pastwiskowego,
a obfitość zielonki zapewnia odpo-
wiednią mleczność krów karmiących
potomstwo.
Racjonalne podejście do powyższych
zagadnień wymaga surowego prze-
strzegania pewnych norm dotyczą-
cych rozrodu bydła mięsnego. W tabe-
li 1. są przedstawione zalecane wartości,
które dotyczą stada zarodowego bydła
mięsnego utrzymywanego w klimacie
umiarkowanym.
Utrzymanie pełnej sezonowości wy-
cieleń nie jest łatwe, zwłaszcza bez re-
strykcyjnego brakowania niepłodnych
samic. Krótki sezon rozrodczy powo-
duje, że kluczowej roli nabiera kwestia
płodności krów mięsnych w okresie
poporodowym.
Okres poporodowy
Powszechnie przyjęto, że okres po-
porodowy (puerperium) u bydła trwa
od zakończenia porodu do momentu
powrotu narządu płciowego do stanu
przedciążowego – z czym związana
jest możliwość ponownego zapłodnie-
nia samicy i rozwoju następnej ciąży.
Procesy przebiegające podczas puer-
perium prowadzą do inwolucji (zwijania
się) macicy oraz wznowienia cyklicznej
aktywności jajników.
W warunkach fizjologicznych, za-
równo u krów mlecznych, jak i mię-
snych, pierwsze pęcherzyki (średnich
rozmiarów) pojawiają się na jajnikach
stosunkowo wcześnie, bo już pod ko-
niec pierwszego tygodnia post partum
(Hartigan, 2004; Stevenson, 2007).
Wyczerpane zasoby LH sprawiają
jednak, że we wczesnym puerperium
(do 10.-15. dnia) nie dochodzi do pełne-
go rozwoju pęcherzyków i jajeczkowa-
nia ze względu na brak odpowiedniego
uwalniania hormonu luteotropowego
(Karg i Schallenberger, 1983; Ptaszyń-
ska, 2008; Stevenson i wsp., 1997). Po-
wyższa sytuacja dotyczy bydła niezależ-
nie od rasy. Od 3. tygodnia po porodzie
zachodzą jednak pewne różnice – w za-
kresie aktywności gonad – pomiędzy
rasami mlecznymi a mięsnymi.
Blokada progesteronowa – ciałko żółte
Dni cyklu rujowego
0
7
14
21
Ryc. 1. Schematyczne przedstawienie falowego wzrostu pęcherzyków jajnikowych (3 fale wzrostu) u bydła
podczas cyklu rujowego
Parametr
Wartość zalecana
Długość sezonu rozrodczego
Poniżej 63 dni
Odsetek zacieleń
(35 dni po zakończeniu sezonu rozrodczego)
Powyżej 95%
Odsetek cieląt żywo urodzonych
(od krów, u których zdiagnozowano ciążę)
Powyżej 93%
Tabela 1. Wskaźniki wydajności rozrodczej stada zarodowego bydła mięsnego i wartości zalecane (wg Kom-
pendium Rozrodu Zwierząt Intervet, red. M. Ptaszyńska)

BYDŁO
WETERYNARIA W TERENIE
y
1/2009
y
19
I tak u zdrowych krów mlecznych
do pierwszej poporodowej owulacji
przebiegającej z reguły bez zewnętrz-
nych objawów rui dochodzi zwykle
w 3-4 tygodniu po wycieleniu (Harti-
gan, 2004; Noakes, 1997; Stevenson,
2007). Jajeczkowanie w tym czasie jest
możliwe dzięki odbudowie zasobów
LH, a co za tym idzie – pojawieniu się
pierwszego po porodzie przedowula-
cyjnego wylewu tego hormonu (Pta-
szyńska, 2008; Sheldon i wsp., 2004).
Druga poporodowa owulacja poprze-
dzona pełnowartościową rują pojawia
się przeważnie do 50. dnia po porodzie
(Stevenson, 2007; Żebracki, 1993).
W porównaniu z bydłem mlecznym
– u ras mięsnych pierwsze poporodo-
we jajeczkowanie ma miejsce znacznie
później, ze względu na obecność czyn-
nika hamującego, jakim jest akt ssania
przez cielę (Borsberry, 2004; Hopkins,
1986; Jaśkowski i Zbylut, 1998; Ste-
venson i wsp., 1997; Yavas i Walton,
2000). Na początku lat 80. ubiegłego
wieku autorzy brytyjscy (Peters i Ri-
ley, 1982) odnotowali, że u karmiących
krów ras mięsnych średni okres od wy-
cielenia do wznowienia cyklu jajniko-
wego wyniósł 59 dni ± 2,5 dnia, przy
czym istniały spore różnice zarówno
w obrębie danego stada, jak i pomię-
dzy poszczególnymi stadami. Powyż-
sze dane liczbowe należy traktować
zarówno ze zrozumieniem, jak i pew-
ną rezerwą, gdyż u bydła mięsnego
na wznowienie cyklicznej aktywności
jajników wpływają – oprócz ssania –
inne czynniki, jak genotyp, warunki
klimatyczne, warunki utrzymania czy
status żywieniowy (Borsberry, 2004).
Niemniej jednak podstawowa istota po-
porodowego anoestrus u ras mięsnych
sprowadza się do faktu, że impulsy wi-
zualne i dotykowe związane z karmie-
niem potomstwa hamują na wyższych
poziomach regulacyjnych wydzielanie
GnRH i LH, a w konsekwencji dojrze-
wanie i owulację pęcherzyków jajniko-
wych (Ptaszyńska, 2008). Należy w tym
miejscu zaznaczyć, że do analogicznej
sytuacji (tj. przedłużenia okresu popo-
rodowej nieczynności jajników) może
dochodzić również u bydła mlecznego
– w przypadku, gdy cielęta przebywają
przy matkach i są przez nie karmione
przez kilka tygodni po urodzeniu (Ja-
nowski i wsp., 1986).
U ras mięsnych w 3-4 tygodniu po wy-
cieleniu dochodzi wprawdzie do odbu-
dowy zasobów LH, ale brak jest pulsa-
cyjnego wydzielania tego hormonu,
ponieważ ssanie wymienia przez cielę
hamuje sekrecję GnRH – a więc tak-
że i LH (Ptaszyńska, 2008, Stevenson,
2007). Dodatkowym czynnikiem ha-
mującym jest zwiększenie wrażliwo-
ści podwzgórza na wpływ ujemnego
sprzężenia zwrotnego wywieranego
przez estrogeny jajnikowe, co związa-
ne jest z faktem, że ssanie stymuluje
uwalnianie endogennych opioidów
(Yaves i Walton, 2000). Na przełomie
pierwszego i drugiego miesiąca docho-
dzi jednak do obniżenia w hamowaniu
uwalnianego GnRH i LH, w wyniku
spontanicznego zmniejszenia często-
tliwości ssania lub ewentualnie przez
odstawienie cielaka (Ptaszyńska, 2008;
Stevenson, 2007). W związku z tym
może już dochodzić do przedowula-
cyjnego wylewu LH i do jajeczkowania.
Tak więc karmiące krowy ras mię-
snych zaczynają zwykle wchodzić
w cykl rujowy w 2. miesiącu po po-
rodzie (Crowe i wsp., 1993; Day i wsp.,
1993; Humphrey i wsp., 1984; Hurnik
i King, 1987; Radford i wsp., 1978; Stagg
i wsp., 1998; Stevenson i wsp., 1997).
Bardzo ważną rolę w podejmowaniu
przez jajniki krów mięsnych cyklicznej
aktywności w okresie puerperium pełni
odpowiednie żywienie (Hopkins, 1986;
Jaśkowski i Zbylut, 1998; Stevenson,
2007). Słabe zaopatrzenie w energię
przed i po porodzie owocuje przedłu-
żaniem się poporodowego anoestrus
(Hill i wsp., 1970). Natomiast odwrotna
sytuacja – tj. wzrastająca podaż energii
w okresie okołoporodowym – przyśpie-
sza powrót cyklicznej aktywności jaj-
ników po wycieleniu (Borsberry, 2004;
Yaves i Walton, 2000). Należy przy
tym pamiętać, że niski poziom gluko-
zy wpływa negatywnie na metabolizm
tkanki nerwowej (a więc i podwzgó-
rza). Dobra kondycja podczas porodu
wpływa pozytywnie na wydzielanie
hormonów płciowych i rozwój pęche-
rzyków jajnikowych w okresie puerpe-
rium, dzięki czemu większość takich
samic zachodzi w ciążę przed zakoń-
czeniem sezonu rozrodczego (Yaves
i Walton, 2000).
Pory roku
a aktywność rozrodcza
Innym aspektem, który warto poru-
szyć, jest kwestia wpływu pory roku
na aktywność rozrodczą bydła. Nale-
ży bowiem pamiętać, że u wielu gatun-
ków ssaków (szczególnie dzikich) dużą
rolę w podejmowaniu funkcji gonad
pełni długość dnia, co związane jest
z działaniem melatoniny wydzielanej
przez szyszynkę. Większość dzikich
gatunków bydłowatych (Bovinae) wy-
kazuje sezonowość rozrodczą (samice
są sezonowo poliestralne), dzięki któ-
rej okres porodów przypada na taką
porę roku, w której istnieje optymal-
na dostępność karmy. Chociaż bydło
domowe było przez setki lat – w trak-
cie procesu udomowiania – selekcjo-
nowane w kierunku poliestralności,
to można zaryzykować twierdzenie,
że w zakresie aktywności rozrodczej
u współczesnych krów mięsnych ist-
nieje szczątkowa wrażliwość na dłu-
gość dnia. Przykładem może tu być
fakt, że przy wycieleniach wiosennych
okres od porodu do pierwszej popo-
rodowej owulacji jest dłuższy niż przy
wycieleniach jesiennych (Borsberry,
2004; King i Macleod; 1985; Yaves
i Walton, 2000).
Warto również odnotować, że u pier-
wiastek acyklia poporodowa jest z reguły
1-4 tygodnie dłuższa niż u wieloródek,
co jest m.in. związane z kontynuacją
wzrostu u młodych matek (Dunn i Kal-
tenbach, 1980; Hopkins, 1986; Noakes,
1997).
Przebieg ocielenia wywiera również
wpływ na aktywność gonad podczas
puerperium – komplikacje porodowe,
porody bliźniacze oraz zatrzymania
łożyska wydłużają bowiem okres po-
porodowego anoestrus (Yaves i Walto,
2000).
Utrzymanie odpowiedniej sezonowo-
ści w stadach bydła mięsnego nie jest
łatwe, ale jest możliwe. Różnorodność
czynników wpływających na rozpo-
częcie cyklicznej aktywności jajników
u krów mięsnych po porodzie sprawia,
że ciężko jest dokładnie sprecyzować
termin pierwszej poporodowej owula-
cji. Aby jednak zapewnić opłacalność
hodowli poprzez utrzymanie pożąda-
nych parametrów, należy z jednej stro-
ny dozorować w odpowiedni sposób
okres okołoporodowy, z drugiej zaś
rozważać ewentualność racjonalnego
wdrażania technik wspomaganego roz-
rodu, takich jak np. farmakologiczna
kontrola cyklu.
Piśmiennictwo
dostępne u autorów
Wyszukiwarka
Podobne podstrony:
wetwterenie artykul 2009 01 24389 cukrzyca bydła
wetwterenie artykul 2009 01 24387(2)
wakcji artykul 2009 01 27633
weterynaria artykul 2009 01 23855
wetwterenie artykul 2009 02 25181
choroby trzustki i watroby 2008 2009 (01 12 2008)
laboratorium artykul 2010 01 28 Nieznany
2009 01 Analiza powłamaniowa
2009 01 Firewall w Linuksie [Poczatkujacy]
2009 01 Vim – konkurent prawie doskonały! [Poczatkujacy]
2009 01 testy odpowiedzi
[2009-01-20] MIKROEKONOMIA - Kolokwium 2 v2, mikro, Mikroekonomia, Ćwiczenia, Ściągi na kolokwia, [2
2009 01 rozszODP (2)
.SIMR-ALG1-EGZ-2009-01-30b-rozw
[2009-01-20] MIKROEKONOMIA - Kolokwium 2, mikro, Mikroekonomia, Ćwiczenia, Ściągi na kolokwia, [2009
2009 01 07
więcej podobnych podstron