7.Podaj fizjologiczny skład moczu ostatecznego wraz z fizjologicznie występującymi elementami upostaciowanymi.
Mocz ostateczny powstaje z moczu pierwotnego w wyniku zagęszczenia, zwrotnej resorpcji i sekrecji w kanalikach nerkowych. Ma on o wiele mniejszą objętość i większe stężenie substancji niż mocz pierwotny (około 200 razy większe stężenie od moczu pierwotnego) Gromadzi się on w miedniczkach nerkowych, skąd moczowodami spływa do pęcherza moczowego. Dobowa ilość moczu wydalanego przez zdrowego człowieka waha się od 750 do 2500 ml
W skład moczu ostatecznego wchodzą: woda (1000g na dobową ilość moczu) mocznik i NaCl (10g) kreatynina, kwas moczowy, amoniak, potas, nieorganiczne siarczany i fosforany (1g) Ca, Mg, fenole, estry siarczanowe (0,1g) białka (śladowe ilości ze złuszczonych nabłonka kanalikowego), ciała ketonowe, zasady purynowe, kwas szczawiowy, urobilinogen, urochromy, urobilina, indykan (0,001g) zbędne produkty przemiany materii, toksyny, metabolity leków oraz substancje obecne we krwi w stężeniu przewyższającym ich próg nerkowy (np. aminokwasy, Hb, glukoza) elementy upostaciowione: złuszczone nabłonki, do kilku erytrocytów i leukocytów w polu widzenia.
8. Opisz autoregulacyjny mechanizm przepływu krwi przez nerki
jest to najważniejsza cecha krążenia nerkowego, polega na tym że nerkowy przepływ krwi i przesączanie kłębuszkowe. (RBF) i osocza (RPF) jest względnie stały, mimo wahań średniego ciśnienia w tętnicach nerkowych w zakresie 80-200mmHg, zanika przy spadku ciśnienia poniżej 80mmHgi powyżej 200mmHg ( u osób z nadciśnieniem tętniczym dolna i górna granica jest przesunięta). Działa ona dwukierunkowo - zapobiega nadmiernemu wzrostowi i zbytniemu spadkowi przepływu krwi i ciśnienia w kapilarach kłębuszka Autoregulacja ma charakter autonomiczny, wewnątrzpochodny, zachodzi też w nerce odwodnionej, przeszczepionej, wyizolowanej. Autoregulacji nie podlega diureza nerkowa - ilość wyprodukowanego moczu, która wzrasta wraz ze wzrostem ciśnienia perfuzyjnego w nerkach.
Rolę w autoregulacji odgrywają:
-właściwości samej ściany naczyń - autoregulacja miogenna czyli skurcz mięśniówki gładkiej naczyń oporowych głównie tętniczek doprowadzających i zwieraczy prekapilarnych przy wzroście ciśnienia transmuralnego (różnicy ciśnienia między światłem naczynia a otaczającym płynem śródmiąższowym) mechanizm ten działa zgodnie z prawem Laplace'a, najsilniejsze właściwości regulacyjne mają naczynia kory nerek (więcej receptorów , są też lepiej unerwione) dlatego przepływ korowy i rdzeniowy mogą być regulowane niezależnie
- czynniki parakrynne - autoregulacja metaboliczna, są miejscowo produkowane:
a) angiotensyna II - powoduje lokalne skurcze tętniczek odprowadzających (dużo receptorów AT1)
b) NO i prostaglandyny PGE i PGI - działanie naczyniorozszerzające, zwiększony przepływ krwi przez nerki w wyniku wzrostu ciśnienia napędowego wypłukuje te substancje warunkując skurcz natomiast przy spadku ciśnienia, hipoksji i niedokrwieniu nerek następuje wzrost biosyntezy PGI ,PGE i NO
- inne czynniki np. alkohol etylowy, prokaina, cyjanki powodują ograniczenie autoregulacji nerkowej
9. Neurohormonalna regulacja krążenia nerkowego:
działanie naczyniorozszerzające:
-nerwy błędne - włókna cholinergiczne; Ach - działa na rec. muskarynowe (rozszerza naczynia, zwiększa przepływ krwi gł. w rdzeniu nerki)
- dopamina - produkowana w nerce
- prostacyklina i NO - wytwarzana przez śródbłonek naczyń kory nerki
- bradykinina (powstaje w nerkach z globulin osocza pod wpływem kalikreiny) uwalnia prostaglandyny które stymulują jej produkcję i uwalnianie
działanie naczyniozwężające:
- współczulne, adrenergiczne nerwy naczynioruchowe - ośrodki w rogach bocznych rdzenia Th12-L2, noradrenalina uwalniana na neuronach pozazwojowych; działa na naczynia poprzez receptory a1- i a2 - adrenergiczne; powoduje głównie skurcz tętnic i tętniczek kory; pobudzenie tego układu następuje w wyniku hipoksji, hiperkapni, hipowolemii, ciężkiej pracy fizycznej
- pobudzenie ośrodków naczynioruchowych opuszki rdzenia
-renina produkowana przez komórki ziarniste(przykłebuszkowe)w wyniku pobudzenia układu współczulnego powoduje przejście angiotensynogenu w angiotensynę I, angiotensyna I przechodzi pod wpływem enzymu konwertującego w angiotensynę II
- tromboksan A2
- prostaglandyny
Układ renina - angiotensyna - aldosteron (RAA)
( fajny schemat Konturek t. III strona 234)
Renina - kwaśna proteza wytwarzana w kom. ziarnistych (przykłębuszkowych) odszczepia od ANGIOTENSYNOGENU dekapeptyd, uwalniając ANGIOTENSYNĘ I , która pod wpływem enzymu konwertującego ACE (jak wybielacz :P) (obecny głównie w komórkach śródbłonkowych płuc, także w nerkach) zmienia się w oktapeptyd - ANGIOTENSYNĘ II jeżeli od angiotensyny II odszczepiona zostanie reszta aspartylowa to powstanie ANGIOTENSYNA III, stymuluje głównie uwalnianie aldosteronu, degradowana przez enzymy proteolityczne (angiotensynazy).
wydzielanie reniny i tworzenie angiotensyny pobudza:
- pobudzenie baroreceptorów w kom. ziarnistych ( mniejsze rozciąganie tętniczki doprowadzającej)
-pobudzenie chemoreceptorów plamki gęstej (przez spadek stężenia Na i Cl w moczu w części dalszej kanalików)
-pobudzenie beta1-receptorów adrenergicznych komórek ziarnistych
-prostaglandyny (prostacyklina) i NO
-stany fizjologiczne takie jak: spadek ciśnienia krwi, odwodnienie, krwotok, stan utraty sodu, marskość wątroby, niewydolność układu krążenia, nagła zmiana pozycji (Poziom-na Pion), czynniki stresorodne.
angiotensyna II ( działa przez receptory AT1 -w ścianie naczyń tętniczych i mózgu (aktywacja białka G, fosfolipazy C; w nadnerczach wzrost wydzielania aldosteronu) i AT2) powoduje:
1. skurcz tętniczek doprowadzających (najsilniejszy wpływ!!!) przez co wzrasta ciśnienie skurczowo-rozkurczowe, skurcz także ułatwia zwrotne wchłanianie Na i wody w kanalikach bliższych, angiotensyna II działa ona także bezpośrednio na ten efekt -> antagoniści: NO, prostacykliny
2. pobudzenie pozazwojowych neuronów współczulnych i uwalnianie noradrenaliny
3. pobudzenie neuronów podwzgórza - wzmożone uwalnianie wazopresyny, ACTH, wzrost pragnienia
4. pobudzenie kory nadnerczy - wzrost uwalniania aldosteronu
5. działa hamująco na uwalnianie reniny
rola fizjologiczna układu renina-angiotensyna-aldosteron:
1.utrzymanie odpowiedniego stosunku miedzy filtracją a zwrotnym wchłanianiem kanalikowym
(równowaga kłębuszkowo-kanalikowa)
2.utrzymanie prawidłowego ciśnienia tętniczego
3. wypełnienie łożyska naczyniowego (wydzielanie aldosteronu)
4. zapewnia zwrotne wchłanianie Na w kanalikach nerkowych
aldosteron - reguluje wchłanianie Na i wydalanie K, jego wydzielanie pobudza ACTH i beta-endorfiny, hamuje: NO i ANP(antagonista aldosteronu); pobudza receptory MR które są głownie w kanaliku dystalnym i cewce zbiorczej, na wskutek jego działania wzrasta synteza podjednostek 3Na/2K- ATPazy i jej aktywność , wzrasta synteza i aktywność kanałów Na, razem z Na wchłaniana jest woda dlatego aldosteron reguluje objętość płynów ustrojowych i wpływa równowagę kwasowo-zasadową (przeciwtransport Na-H)
zniszczenie kory nadnerczy -> brak aldosteronu -> choroba Addisona
nadprodukcja hormonu -> zespół Conna
11. Budowa aparatu przykłębuszkowego
1.nabłonkowate komórki ziarniste przykłębuszkowe - przekształcone komórki mięśniowki gładkiej tętniczki doprowadzającej; zawierają ziarnistości z reniną , funkcjonują jako baroreceptory.
2. komórki plamki gęstej - w ścianie kanalika dalszego, w miejscu gdzie przylega ona do bieguna naczyniowego ciałka nerkowego; wyższe od reszty komórek, są to chemoreceptory regulujące stężenie Na w pramoczu i szybkość jego przepływu przez dalszą część nefronu
3.Komórki mezangialne zewnętrzne(pozakłębuszkowe) - wypełniają przestrzeń między plamką gęstą a obydwoma tętniczkami bieguna naczyniowego; regulują lokalny przepływ krwi przez ciałko nerkowe.
Unerwienie: nerwy współczulne - zaopatrują kom ziarniste przez B1-rec adrenergiczne.
Chemoreceptory, baroreceptory i receptory adrenergiczne współdziałają w uwalnianiu reniny i aktywacji układu renina - angiotensyna.
12. czynniki regulujące wydzielanie reniny:
pobudzanie uwalniania:
- pobudzenie baroreceptorów w kom. ziarnistych przez mniejsze rozciąganie tętniczki doprowadzającej
-pobudzenie chemoreceptorów plamki gęstej (przez spadek stężenia Na i Cl w moczu w części dalszej kanalików)
-pobudzenie beta1-receptorów adrenergicznych komórek ziarnistych
-prostaglandyny,prostacyklina, NO
-stany fizjologiczne takie jak: spadek ciśnienia krwi, odwodnienie, krwotok, stan utraty sodu, marskość wątroby, niewydolność układu krążenia, nagła zmiana pozycji (Poziom-na Pion), czynniki stresorodne.
hamowanie uwalniania:
- angiotensyna II
- wazopresyna
- blokowanie biosyntezy prostaglandyn indometacyną
- blokada receptorów Beta-adrenergicznych np. propranololem
13. w kontekście wchłaniania glukozy wyjaśnij pojęcia: transport maksymalny i próg nerkowy
glukoza ulega przesączeniu w kłębuszkach i wchłania się całkowicie w kanalikach bliższych, wymaga to obecności jonów Na; glukoza i Na łączą się ze wspólnym nośnikiem SGLT 2 i SGLT 1 w błonie przyszczytowej komórek kanalikowych i dostają się do światła zgodnie z gradientem, potem Na zostają wypompowane do przestrzeni międzykomórkowej przez Na-K-ATPazę a glukoza przetransportowana tam za pomocą przenośnika GLUT 2, odbywa się ona na zasadzie wtórnego transportu czynnego w którym pompa Na-K-ATPaza w błonie bocznej i przypodstawnej stanowi główną siłę napędową (nie podlega on wpływom insuliny, można go zablokować za pomocą czynników hamujących fosforylację glukozy - cyjanków i florydzyny)
czynnikiem ograniczającym wchłanianie jest transport maksymalny (Tm) który dla glukozy wynosi 375mg/min (2,1mmol/min) glukoza zaczyna się pojawiać w moczu jeżeli jej stężenie w osoczu przekracza 160-180mg% (8,9-10 mmol/l) stanowi to jej próg nerkowy; wydalanie glukozy w moczu wzrasta stopniowo i proporcjonalnie do wzrostu jej stężenia we krwi
14. znaczenie wazopresyny w zagęszczaniu moczu
najważniejsza rola ADH - zwiększanie biernego transportu wody w kanalikach bliższych i dalszych zależnego od gradientu stężeń osmotycznych - ma to podstawowe znaczenie w procesie wydzielania hipertonicznego moczu ostatecznego ( gradient ten z kolei ściśle zależy od ilości związków osmotycznie czynnych nagromadzonych w rdzeniu nerki, ADH zwiększa współtransport Na-K-2Cl w grubej części ramienia wstępującego pętli Henlego, działa synergistycznie z aldosteronem zwiększając syntezę i aktywność kanału sodowego a także zwiększa transport mocznika w części dystalnej kanalików zbiorczych - te efekty wzmagają zdolność nerek do produkcji hiperosmotycznego moczu
wydzielanie ADH pobudza: wzrost ciśnienia osmotycznego, zmniejszenie objętości krwi krążącej, spadek ciśnienie tętniczego, pobudzenie baroreceptorów, alkohol, nikotyna, mannitol, stojąca pozycja ciała, wysiłek fizyczny, cukrzyca
niedobór ADH lub brak jego działania powoduje moczówkę prostą (wydalane jest dobowo12-15l moczu). Jeśli dotyczy ona zaburzenia wydzielania na poziomie podwzgórza lub przysadki jest to moczówka prosta pochodzenia ośrodkowego. Jeżeli występuje niewrażliwość cewek nerkowych na działanie hormonu ADH (defekt syntezy receptora) - jest to moczówka prosta nefrogenna (rzadziej występująca)
nadmiar ADH - zespół nieprawidłowego wydzielania wazopresyny (SIADH) powoduje przewodnienie lub nadmierne rozcieńczenie
15. mechanizm działania wazopresyny w nefronie
wazopresyna syntetyzowana przez neurony neurosekrecyjne w jądrach nadwzrokowych i przykomorowych podwzgórza, magazynowana w tylnym płacie przysadki;
działa przez receptory V2 w ścianie kanalików dalszych i cewek zbiorczych oraz V1 w ścianie naczyń; mechanizm stymulacji V2 polega na wbudowaniu akwaporyny 2 (APQ2) do ściany luminalnej (szczytowej) kanalika dalszego i cewki zbiorczej,
etapy:
1)niepobudzona APQ2 jest w pęcherzykach wewnątrz komórek
2)stymulacja wazporesyną receptora uaktywnia cyklozę andenylanową i wzrasta stężenie cAMP które pobudza kinazę białkową A
3)następuje fosforylacja APQ2,
4) ufosforylowana APQ2 wchodzi w interakcję z białkami cytoplazmy które umożliwiają mu transport i wbudowanie do błony luminalnej przez co wzrasta przepuszczalność dla wody co wywołuje jej re absorpcję.
16. Opisz losy jonów sodu w nefronie
W kłębuszkach nerkowych przesączone zostaje około 25 000 mmol na dobę. Około 70% wchłania się w kanalikach bliższych, 15% w ramieniu wstępującym pętli nefronu, 9% w kanalikach dalszych, a 5% w kanalikach zbiorczych. 1% jest wydalany z moczem. Reabsorbcja sodu w kanalikach bliższych odbywa się na zasadzie transportu wtórnie czynnego i jest procesem dwustopniowym: (1) Pierwszy etap polega na transporcie ułatwionym przez błonę przyszczytową do komórek kanalikowych dzięki: gradientowi elektrochemicznemu i obecności nośników dla transportu ułatwionego z glukozą i aminokwasami (2) Drugi etap polega na aktywnym transporcie przez błonę przypodstawną i boczną za pośrednictwem Na+-K+ pompy ATP-azowej. Wchłanianie zwrotne w kanalikach dalszych i zbiorczych odbywa się w pierwszej kolejności(1) poprzez specjalne kanały sodowe zależne od: aldosteronu (powodującego otwieranie się kanałów sodowych oraz wzrost aktywności Na+-K+ pompy ATP-azowej), ANP (indukującego zamykanie się kanałów poprzez wzrost stężenia cytoplazmatycznego c-GMP), PGE2 (indukującego zamykanie się kanałów poprzez wzrost stężenia cytoplazmatycznego jonów Ca2+ oraz niezależnie od tego stężenia zmniejszającego aktywność Na+-K+ pompy ATP-azowej) (2) następnie przy pomocy Na+-K+ pompy ATP-azowej do płynu śródmiąszowego.
17. Opisz losy jonów potasu w nefronie
W kłębuszkach nerkowych przesączone zostaje około 750 mmol na dobę. Około 65% wchłania się w kanalikach bliższych, 25% w ramieniu wstępującym pętli nefronu. Do kanalików dalszych dopływa mocz zawierający 10% przesączonego potasu, zachodzi tu wydzielanie potasu do światła kanalika w ilości 10-150% przesączonego potasu. W kanalikach zbiorczych zostaje wydalone ok. 5-30%.
Wchłanianie potasu z kanalików bliższych jest czynne. Wydzielanie K+ w kanalikach dalszych i zbiorczych jest bierne i zależy od (1) gradientu elektrycznego, (2)ilości moczu kanalikowego i (3)stężenia K+ w osoczu. Gradient elektryczny wytworzony jest przez zwrotne wchłanianie Na+ , które to jony wymieniane są na H+ ,co potęguje zmianę gradientu. Wzrost wchłaniania Na+ > wzrost gradientu elektrycznego > wzrost dyfuzji K+
18. Wskaż znane Ci mechanizmy działania czynników regulujących wydalanie wody wraz z moczem
Czynniki regulujące wydalanie wody z moczem to: (1) klirens osmotyczny: wzrost lub spadek wchłaniania zwrotnego substancji osmotycznie czynnych powoduje odpowiednio zmniejszenie lub zwiększenie diurezy osmotycznej, której wzrost jest spowodowany zatrzymaniem substancji osmotycznie czynnych w kanalikach, co zmniejsza wchłanianie zwrotne wody;
(2) ciśnienie onkotyczne białek osocza: jego wzrost hamuje, a spadek wzmaga filtracje kłębuszkową. Dodatkowo wszelkie zmiany ciśnienia onkotycznego wpływają na wchłanianie płynu śródmiąższowego nerki i składników w nim rozpuszczonych
(3) stopień pobudzenia układu sympatycznego: jego wzrost powoduje skurcz tętniczek doprowadzających > zmniejszenie przepływu krwi przez kłębuszki > spadek filtracji i vice versa
(4) ciśnienie tetnicze krwi: jego wzrost nieznacznie zwiększa filtrację kłębuszkową (przy wzroście ciśnienia do 200mm Hg wzrost filtracji o zaledwie 20%, spadek dopiero poniżej 60mm Hg ją zatrzymuje), lecz wzmaga ciśnienie hydrostatyczne w kapilarach okołokanalikowych co znacznie zmniejsza resorbcję zwrotna wody.
(5) Działanie ADH na kanaliki: Jego uwalnianie do krwioobiegu przez układ podwzgórzowo-przysadkowy pod wpływem wielu różnych czynników świadczących o utracie wody z organizmu (a także nikotyny, alkoholu, barbituranów, morfiny), a następnie wiązanie się z kompleksem receptorowym V2, powoduje wzrost produkcji akwaporyny-2 tworzącej kanały wodne w kanalikach zbiorczych, co ułatwia wchłanianie zwrotne wody i zagęszczanie moczu.
19. Opisz mechanizm mikcji i jego kontrolę nerwową
Mikcja: czyli oddawanie moczu. W miarę wypełniania się pęcherza, jego mięśnie gładkie rozluźniają się utrzymując ciśnienie w jego wnętrzu na stałym poziomie, aż do momentu, kiedy jego objętość przekroczy 400 mL. Podrażnienie mechanoreceptorów przez gromadzący się mocz prowadzi do odruchowego skurczu pęcherza w wyniku impulsacji aferentnej w trzewnych włóknach czuciowych do rdzenia i z powrotem eferentnie we włóknach parasympatycznych (nerw miedniczy) pobudzenie włókien eferentnych przez aferentne w segmentach krzyżowych rdzenia zależy od torowania z ośrodka w moście i hamowania z ośrodka w śródmózgowiu. Część włókien po dotarciu do ośrodka torującego w moście biegnie dalej do kory, dzięki czemu występuje uczucie parcia na mocz. Wzrost impulsacji w nerwie miedniczym (przywspółczulnym) powoduje skurcz mięśnia wypieracza moczu, którego włókna pociągają ku górze włókna zwieracza wewnętrznego, co powoduje otwarcie cewki moczowej. Jednocześnie zahamowaniu ulegają neurony współczulne, co powoduje rozluźnienie pęcherza poprzez działanie noradrenaliny na receptory adrenergiczne typu β. Po połączeniu skurczu wypieracza moczu z rozluźnieniem zwieracza wewnętrznego cewki moczowej i wzrostem ciśnienia wewnętrznego cewki moczowej, może dojść do wyparcia moczu. Na drodze stoi jednak zwieracz zewnętrzny cewki moczowej unerwiony przez nerw sromowy i podległy naszej woli, co umożliwia nam zahamowanie oddawania moczu. W miarę rozwoju rola ośrodków odruchowych rdzenia jest częściowo przejmowana przez kolejno: ośrodek mostowy > śródmózgowiowy > w tylnym podwzgórzu i w obrębie gornego zakrętu czołowego.
20. Wskaż udział nerek w czynnościach pozawydalniczych.
Poza wydalaniem zbędnych lub szkodliwych produktów końcowych metabolizmu, jak mocznik, kreatynina, kwas moczowy i inne. Nerki biorą udział w:
regulacji objętości i składu substancji nieorganicznych i organicznych oraz osmolarnosci płynu zewnątrzkomórkowego
regulacji równowagi kwasowo-zasadowej, głównie przez wydalanie nadmiaru kwasów i oszczędzanie zasad
wydzielaniu do krwi substancji o działaniu hormonalnym: (1)erytropoetyna, (2)renina, (3)bradykinina, (4)prostaglandyny, (5)aktywna postać witaminy D3
czynnościach metabolicznych jak detoksykacja np. Kwasu benzoesowego przez połaczenie go z glicyną do kwasu hipuronowego, glukoneogeneza w warunkach głodu, utlenianie aminokwasów
21. Narysuj schemat przedstawiający nerkowe mechanizmy utrzymywania izohydrii
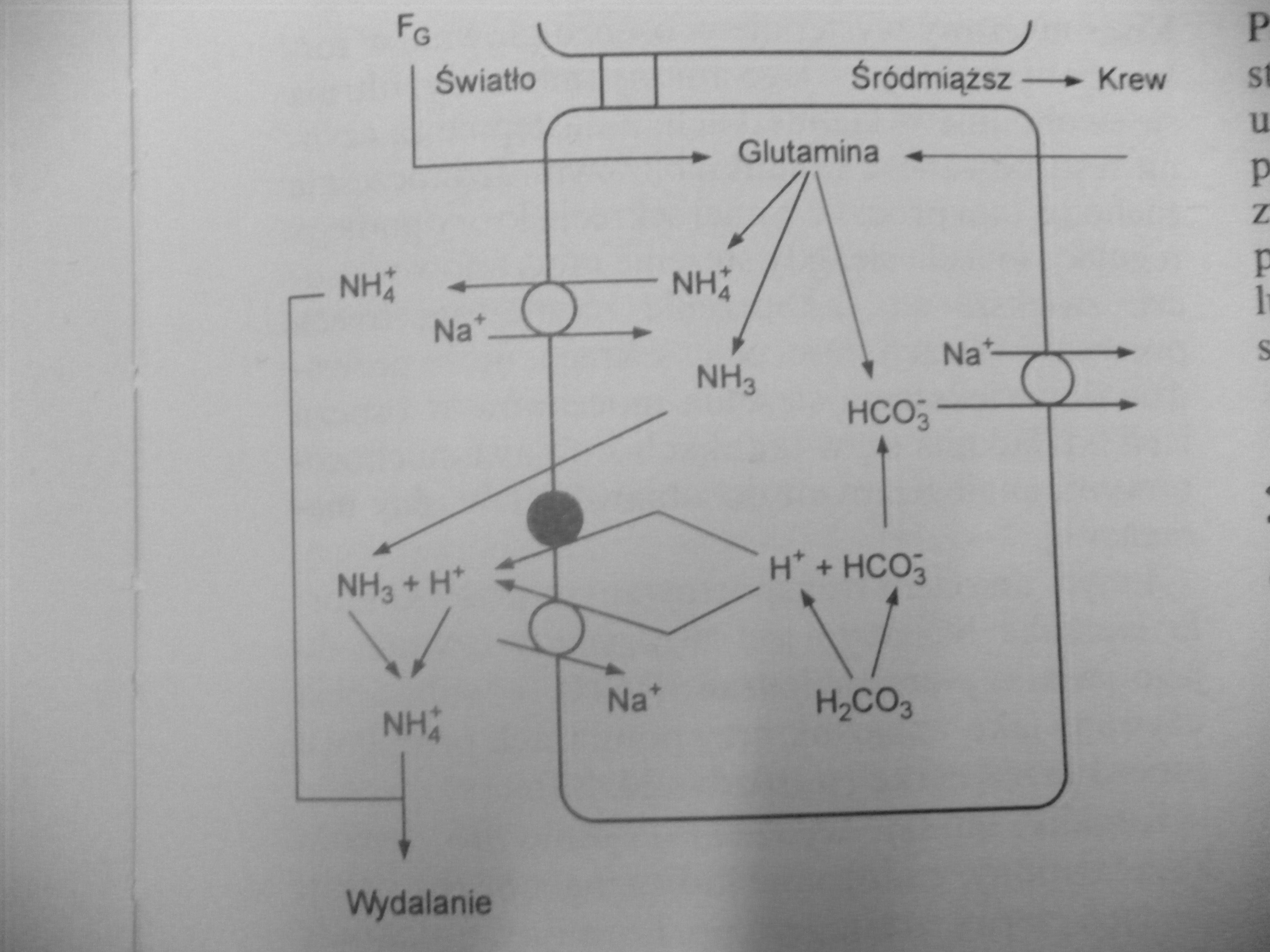
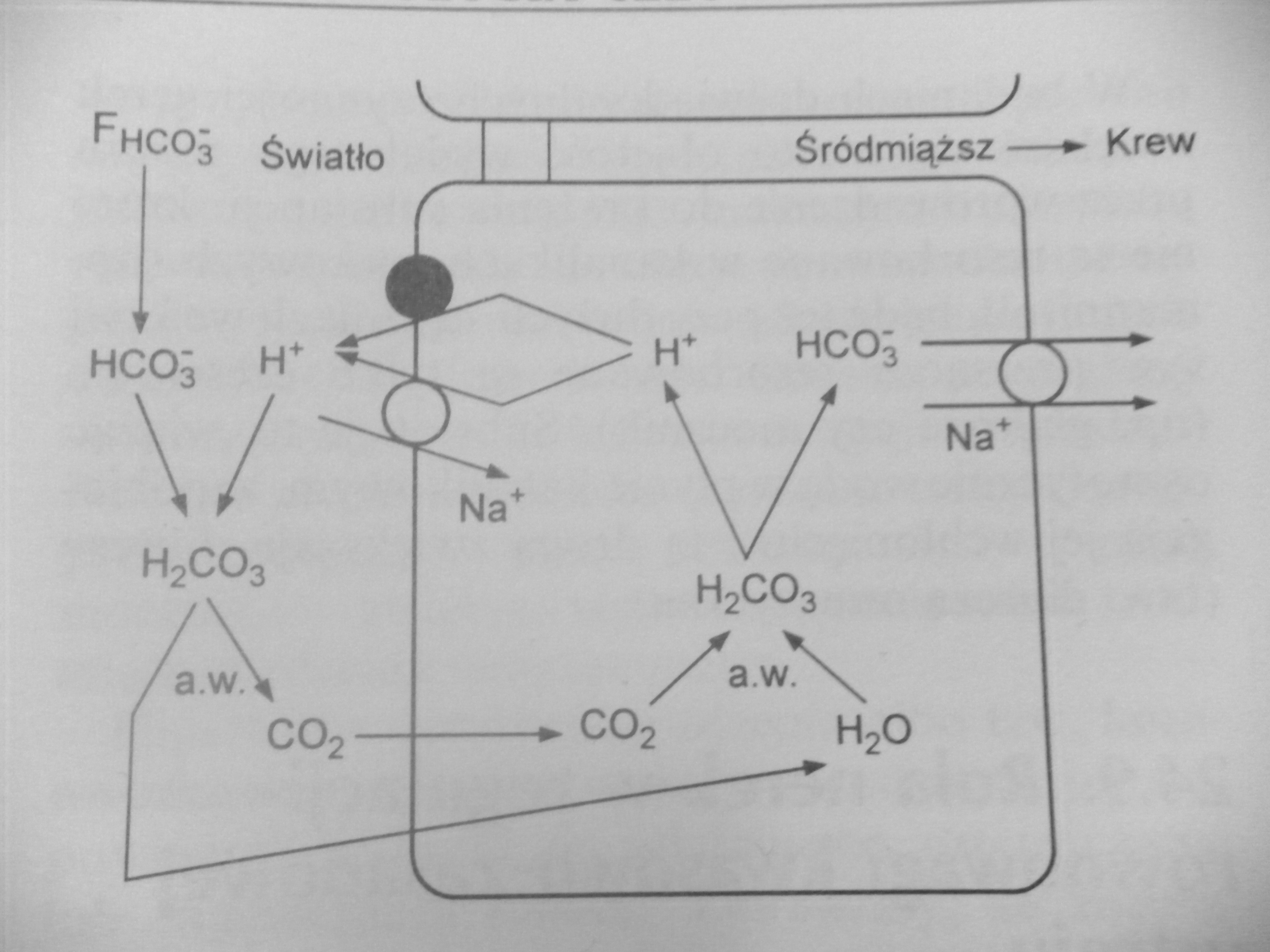
Izohydria polega na utrzymaniu stałego stężenia jonów H+ w płynie zewnątrzkomórkowym. Nadmiar tego jonu wytwarzany jest w procesach metabolicznych, zwłaszcza przy diecie zawierającej duże ilości białka, a jego dzienne wytwarzanie w przypadku tej diety może sięgać nawet 60mmol. Powstający w komórkach organizmu nadmiar jonów H+ jest z nich usuwany do płynu zewnątrzkomórkowego, gdzie łączy się z buforem HCO3 - lub buforem fosforanowym, lub trafia w niezmienionej postaci do nerek. Nerki są jedynym narządem, który może usuwać jony H+ i odtwarzać bufor HCO3 -. W komórkach kanalikowych zawarta jest anhydraza węglanowa umożliwiająca odtwarzanie kwasu węglowego z H2O i CO2
Schemat pierwszy obrazuje odtwarzanie jonu wodorowęglanowego w kanalikach bliższych. Proces ten jest niezbędny dla przeżycia organizmu, gdyż HCO3- jest przefiltrowany w kłębuszku nerkowym, a jego utrata prowadziłaby do zakwaszenia płynu zewnątrzkomórkowego.
Na schemacie drugim mamy bufor fosforanowy. 25% przesączonego HPO42- nie ulega resorpcji zwrotnej i buforuje dostające się tutaj z krwią poprzez komórki kanalika jony H+ z jednoczesnym odtwarzaniem buforu HCO3 -. Bufor ten umożliwia wydalenie z moczem dziennie 30mmol H+
Reszta jonów H+ jest wydalana w formie NH4+. Jon ten powstaje w wyniku rozkładu glutaminy. Następnie dyfunduje do światła kanalika, gdzie łączy się jonem H+. Amoniogeneza może znacznie wzrosnąć w wyniku zakwaszenia organizmu dzięki czemu organizm radzi sobie z procesami metabolicznymi znacznie obniżającymi pH
22. Erytropoetyna (EPO) - wytwarzanie, działanie, fizjologiczne znaczenie
Erytropoetyna powstaje w wyniku odszczepienia od α2 globuliny osocza - proerytropoetynogenu, produkowanego w wątrobie. Odszczepienie to przeprowadza erytrogenina, wytwarzana w nerkach. Erytropoetyna wzmaga przekształcanie się niezróżnicowanych komórek macierzystych szpiku w bardziej zróżnicowane komórki układu erytroblastycznego. Oprócz tego pobudza syntezę hemoglobiny. Przypuszcza się, że erytropoetyna działa za pośrednictwem swoistego receptora. Gwałtowny wzrost wytwarzania erytropoetyne obserwuje się w stanach upośledzenia transportu tlenu do tkanek (hipoksja, spadek liczby erytrocytów, spadek objętości krwi, choroby serca i płuc, redukcja zawartości hemoglobiny). Szczyt erytropoezy w czasie wzmożonej produkcji erytropoetyny ma miejsce po upływie 5-7 dni. Z chwilą ustąpienia hipoksji, gdy komórki otrzymują wystarczającą ilość tlenu, ustaje zupełnie wytwarzanie erytropoetyny i erytropoeza spada niemal do zera. Mechanizm kontroli erytropoezy przez erytropoetynę działa więc na zasadzie zwrotnego sprzężenia ujemnego.
23. Opisz regulację mechanizmu wytwarzania aktywnej witaminy D3 oraz znaczenie fizjologiczne tego procesu.
7-hydroksycholesterol
↓skóra pod wpływem światła
cholekalcyferol (wit.D)
↓transport we krwi w połączeniu z DBP do wątroby, w hepatocytach
25-hydroksycholekalcyferol
Transportowany we krwi do nerek
1,25-dihydroksycholekalcyferol 24,25-dihydroksycholekalcyferol
DBP - globulina wiążąca witaminę D
regulacja
Reakcja prowadząca do powstania 25(OH)D3 jest hamowana na zasadzie sprzężenia zwrotnego przez samą 25(OH)D3, uwalnianą przez hepatocyty do krwi.
1,25-dihydroksycholekalcyferol jest bardziej aktywny w odniesieniu do mobilizacji wapnia i może być uważany za właściwy hormon.
24,25-dihydroksycholekalcyferol jest produkowany w ilościach około 1000 razy większych, ale wykazuje słabsze działanie na komórki jelitowe i jest mniej aktywny.
Zamiana 25(OH)D3 na 1,25(OH)2D3 przebiega tylko w obecności parathormonu, który pobudza syntezę i aktywność odpowiedniej hydroksylazy w komórkach kanalików proksymalnych nerek.
Znaczenie
Witamina D3 ma dwa główne narządy docelowe, tj. jelito i kości, oraz jeden narząd pomocniczy, tzn. nerki.
podnosi ona stężenie wapnia i fosforanów w płynie zewnątrzkomórkowym, zapewniając prawidłową mineralizacje i odnowę tkanki kostnej
zapewnia utrzymanie stałego stężenia Ca2+ w osoczu
wywiera silny wpływ na nabłonek jelitowy, pobudzając proces resorpcji wapnia z jelit
jej wpływ na kości polega na ułatwieniu działania parathormonu
24. Opisz wpływ angiotensyny II na czynność nerek.
Angiotensyna II powoduje zmniejszenie filtracji kłębuszkowej oraz zwiększenie resorpcji sodu. Efekty te są wynikiem dokuczającego działania angiotensyny II, stymulacji wydzielania aldosteronu, stymulacji pozajelitowych neuronów współczulnych do wydzielania NA oraz stymulacji neuronów podwzgórza do wzmożonego uwalniania wazopresyny. Ponadto angiotensyna II działa na komórki przykłębuszkowe, hamując uwalnianie reniny.
Wyszukiwarka
Podobne podstrony:
wyklad III- uklad wydalniczy, Biologia, zoologia
language tests 8b
language test 8b
historyjka obrazkowa 8b
exam skills test 7&8b
Ćwiczenie 13a, Patofizjologia, Ćwiczenia 13-15 (wydalniczy, nerwowy, nowotwory, toksykologia, rytmy
Metabolizm tłuszczowców, Patofizjologia, Ćwiczenia 13-15 (wydalniczy, nerwowy, nowotwory, toksykolog
VI klucz uklad wydalniczy, sprawdziany, biologia
układ wydalniczy i rozrodczy
układ wydalniczy i odpornosciowy
Language Test 8B
Ściąga Podsumowanie działu 6 Układ Wydalniczy
Fizjologia Zwierząt Trawienie, Wydalniczy Ściąga
Pytania z kolosów i egz Wydalniczy pytania Puzio
więcej podobnych podstron