Biological Foundations of Human Behavior
Neuron
ion channels - Membranes, or phospholipids bilayers, build a hydrophobic, low dielectric barrier to hydrophilic and charged molecules. They are electrical insulators. Ion channels provide a high conducting, hydrophilic pathway across the hydrophobic interior of the membrane. The channel, or pore structure, is said to catalyze the 'reaction' of transporting charged molecules across a low dielectric medium. The 'catalytic site', the central channel, is either open or closed. The open channel conformation can be compared to the transition state of the enzyme-substrate complex, where ions are tightly associated to the catalytic site. The conformational change between closed and open state is called gating, as in opening and closing a gate. Channel gating is controlled by external factors like enzymes are controlled by modulators and effectors. Ion channels can be classified according to which chemical or physical modulator controls their gating activity
concentration gradient
The graduated difference in concentration of a solute per unit distance through a solution.
electrochemical gradient - The diffusion gradient of an ion, representing a type of potential energy that accounts for both the concentration difference of the ion across a membrane and its tendency to move relative to the membrane potential.
sodium-potassium pump (Na+/K+) - The Na+/K+ pump is found in the membranes of many types of cells. In particular, it plays a very important role in nerve cell membranes. Notice that 3 positive ions (Na+) are pumped out of the cell (towards ECF) for every 2 positive ions (K+) pumped into the cell (towards ICF). This means that there is more positive charges leaving the cell than entering it. As a result, positive charge builds up outside the cell compared to inside the cell. The difference in charge between the outside and inside of the cell allows nerve cells to generate electrical impulses which lead to nerve impulses. The Na+/K+ pump illustrates "active transport" since it moves Na+ and K+ against their concentration gradients. That is because there is already a high concentration of Na+ outside the cell and a high concentration of K+ inside the cell. In order to move the ions (Na+ and K+) against their gradients, energy is required. This energy is supplied by ATP (adenosine triphosphate). An ATP molecule floating inside the cell, binds to the pump transferring some energy to it. As the energy is used, the ATP falls off and having lost its energy it is converted into ADP (adenosine diphosphate).
Notice in the diagram that there are 3 binding sites for the 3 Na+ ions on the inner surface of the pump and 2 binding sites for the 2 K+ ions on the outer surface of the pump. The shape of these binding sites ensures that only Na+ and K+ can bind and be transported.
Depolarization - reduction, toward zero, of the membrane potential of a cell from its normal resting potential
Hyperpolarization - an increase in membrane potential of a cell, relative to the normal resting potential
dendrite - The treelike extensions of a neuron. Most neurons have multiple dendrites, which are short and typically highly branched. Dendrites are specialized for receiving information and form synaptic contacts with the terminals of other nerve cells to allow nerve impulses to be transmitted.
axon (collateral, axon hillock, myelin) - - axon - a single, elongated tubular extension of a neuron that conducts action potentials away from the cell body toward the neuron terminal.
axon hillock - the first portion of a neuronal axon plus the region of the cell body from which the axon leaves: the site of action potential initiation in most neurons
Myelin - a substance consists of multiple layers of a glial cell (Schwann cell or oligodendrocyte) membrane wrapped around the axon of a neighboring neuron. This myelin “insulation” greatly increases the conduction rate along the axon. That is, non-myelinated neurons carry impulses about 50 times slower than myelinated neurons. The portions of the axon of a myelinated neuron that are not wrapped in myelin are called the nodes of Ranvier. The axons of some neurons branch to form axon collaterals along which the bifurcated impulse travels simultaneously to signal more than one other cell.
synapse (presynaptic membrane, synaptic cleft, postsynaptic membrane) - Synapse is a junction at which the axon of the presynaptic neuron terminates at some location upon the postsynaptic neuron. The end of a presynaptic axon, where it is juxtaposed to the postsynaptic neuron, is enlarged and forms a structure known as the terminal button. An axon can make contact anywhere along the second neuron: on the dendrites (an axodendritic synapse), the cell body (an axosomatic synapse) or the axons (an axo-axonal synapse). Nerve impulses are transmitted at synapses by the release of chemicals called neurotransmitters. As a nerve impulse, or action potential, reaches the end of a presynaptic axon, molecules of neurotransmitter are released into the synaptic cleft
1. action potential invades presynaptic bouton, causing the whole bouton to be depolarized
2. depolarization triggers the opening of calcium voltage-gated channels
3. calcium ions flood in (they are in higher concentration in the extracellular solution)
4. calcium ions interact with the synaptic vesicles, allowing for their exocytosis
5. neurotransmitter spills out into the synaptic cleft (exocytosis released it) 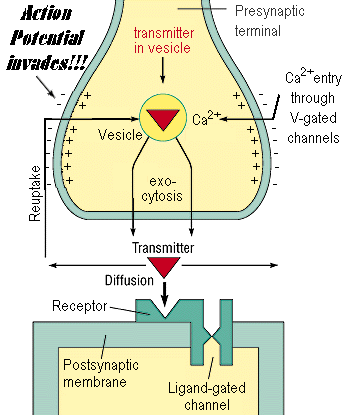
6. neurotransmitter diffuses across the synaptic cleft
7. neurotransmitter interacts with receptors at the postsynaptic membrane
8. receptors tend to be ligand-gated channels, so they open when neurotransmitter binds
9. ions flow through the ligand-gated channels, causing a change in the postsynaptic membrane potential
transmitters: excitatory (e.g. glutamate) and inhibitory (e.g. GABA). - Communication of information between neurons is accomplished by movement of chemicals across a small gap called the synapse. Chemicals, called neurotransmitters, are released from one neuron at the presynaptic nerve terminal. Neurotransmitters then cross the synapse where they may be accepted by the next neuron at a specialized site called a receptor. The action that follows activation of a receptor site may be either depolarization (an excitatory postsynaptic potential e.g. glutamate) or hyperpolarization (an inhibitory postsynaptic potential e.g. GABA). A depolarization makes it more likely that an action potential will fire; a hyperpolarization makes it less likely that an action potential will fire.
Glutamate: an amino acid; the most important excitatory neurotransmitter in the brain
GABA: an amino acid; the most important inhibitory neurotransmitter in the brain
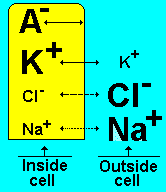
resting potential - When a neuron is not sending a signal, it is "at rest." When a neuron is at rest, the inside of the neuron is negative relative to the outside. Although the concentrations of the different ions attempt to balance out on both sides of the membrane, they cannot because the cell membrane allows only some ions to pass through channels (ion channels). At rest, potassium ions (K+) can cross through the membrane easily. Also at rest, chloride ions (Cl-) and sodium ions (Na+) have a more difficult time crossing. The negatively charged protein molecules (A-) inside the neuron cannot cross the membrane. In addition to these selective ion channels, there is a pump that uses energy to move three sodium ions out of the neuron for every two potassium ions it puts in. Finally, when all these forces balance out, and the difference in the voltage between the inside and outside of the neuron is measured, you have the resting potential. The resting membrane potential of a neuron is about -70 mV - this means that the inside of the neuron is 70 mV less than the outside. At rest, there are relatively more sodium ions outside the neuron and more potassium ions inside that neuron.
threshold potential - The threshold potential is the membrane potential to which a membrane must be depolarized to initiate an action potential
action potential (spike)
The resting potential tells about what happens when a neuron is at rest. An action potential occurs when a neuron sends information down an axon, away from the cell body. Neuroscientists use other words, such as a "spike" or an "impulse" for the action potential. The action potential is an explosion of electrical activity that is created by a depolarizing current. This means that some event (a stimulus) causes the resting potential to move toward 0 mV. When the depolarization reaches about -55 mV a neuron will fire an action potential. This is the threshold. If the neuron does not reach this critical threshold level, then no action potential will fire. Also, when the threshold level is reached, an action potential of a fixed sized will always fire...for any given neuron, the size of the action potential is always the same. There are no big or small action potentials in one nerve cell - all action potentials are the same size. Therefore, the neuron either does not reach the threshold or a full action potential is fired - this is the "ALL OR NONE" principle.
Action potentials are caused by an exchange of ions across the neuron membrane. A stimulus first causes sodium channels to open. Because there are many more sodium ions on the outside, and the inside of the neuron is negative relative to the outside, sodium ions rush into the neuron. Remember, sodium has a positive charge, so the neuron becomes more positive and becomes depolarized. It takes longer for potassium channels to open. When they do open, potassium rushes out of the cell, reversing the depolarization. Also at about this time, sodium channels start to close. This causes the action potential to go back toward -70 mV (a repolarization). The action potential actually goes past -70 mV (a hyperpolarization) because the potassium channels stay open a bit too long. Gradually, the ion concentrations go back to resting levels and the cell returns to -70 mV.
postsynaptic potential - Postsynaptic conductance changes and the potential changes that accompany them alter the probability that an action potential will be produced in the postsynaptic cell. At the neuromuscular junction, synaptic action increases the probability that an action potential will occur in the postsynaptic muscle cell; indeed, the large amplitude of the EPP ensures that an action potential always is triggered. At many other synapses, PSPs actually decrease the probability that the postsynaptic cell will generate an action potential. PSPs are called excitatory (or EPSPs) if they increase the likelihood of a postsynaptic action potential occurring, and inhibitory (or IPSPs) if they decrease this likelihood. Given that most neurons receive inputs from both excitatory and inhibitory synapses, it is important to understand more precisely the mechanisms that determine whether a particular synapse excites or inhibits its postsynaptic partner.The principles of excitation just described for the neuromuscular junction are pertinent to all excitatory synapses. The principles of postsynaptic inhibition are much the same as for excitation, and are also general. In both cases, neurotransmitters binding to receptors open and close ion channels in the postsynaptic cell. Whether a postsynaptic response is an EPSP or an IPSP depends on the type of channel that is coupled to the receptor, and on the concentration of permeant ions inside and outside the cell. In fact, the only factor that distinguishes postsynaptic excitation from inhibition is the reversal potential of the PSP in relation to the threshold voltage for generating action potentials in the postsynaptic cell
- excitatory post synaptic potential (EPSP) - an excitatory depolarization of the postsynaptic membrane of a synapse caused by the liberation of the neurotransmitter by the terminal button
- inhibitory post synaptic potential (IPSP) - an inhibitory hyperpolarization of the postsynaptic membrane of a synapse caused by the liberation of neurotransmitter by the terminal button
coding stimulus strength by spike frequency - Stimulus strength is usually reflected in the firing frequency (spikes per second) of the fiber. The firing frequency of tonic afferents reflects absolute stimulus strength, whereas the firing frequency of phasic afferents reflects changes in stimulus strength.
spatial and temporal summation (slow potential theory) - Motor neuron receives over 1,000 presynaptic terminals, and each postsynaptic potential is below threshold. Thus postsynaptic potentials must summate to reach the depolarization threshold. Summation of postsynaptic potentials occurs when a presynaptic neuron fires repeatedly at a high rate ("temporal summation") or when several presynaptic terminals fire at the same time ("spatial summation") or from a combination of temporal and spatial summation. When the threshold for voltage-gated sodium channels activation is reached (at the hillock) an action potential occurs. This is an all-or-none process, like flushing a toilet: it either occurs or it does not. Finally, we compare a neuron with a circuit of the nervous system. In this analogy, dendrites and the soma are the afferent limb, and the axon is the efferent limb. The axon hillock is the integrative center, gathering information received by the dendrites and soma and deciding to fire an action potential or not
tonic and phasic cell activities (slow and fast adapting cells)
Neural Net
receptive field - There are many millions of receptors in the retina. There is only about one million optic nerve fibers sending visual signals up to the high brain centers. Consequently, individual receptors do not have private lines up to the visual cortex. Rather, multiple receptors converge on to subsequent neural units on their way to the higher visual centers. This convergence results in a physiological concept known as receptive fields.
Receptive field of a neuron in the visual system is the part of the visual field, that an individual neuron “sees”- that is, the part in which light must fall for neuron to be stimulated. The location of the receptive field of a particular neuron depends on the location of the photoreceptors that provide it with visual information. If a neuron receives inf. from photoreceptors located in the fovea, its receptive field will be at the fixation point-the point at which the eye is looking. If the neuron receives inf. from photoreceptors located in the periphery of the retina, its receptive field will be located off to one side.
concentric receptive field (center and surround), on-center and off-center) - A typical way to study the receptive fields is to stimulate the retina with patterns of light of every conceivable size, shape, color, or motion, while recording the activity of a single cell with microelectrodes. Working in this way, researchers found that the retina ganglion cells and the cells of the LGN(lateral geniculate nucleus) are cells with concentric, center-surround receptive fields. Kuffler discover that cat receptive field consists of a roughly circular center, surrounded by a ring. Stimulation of the center or surrounding fields had contarary effects: ON cells were excited by light falling in the central field (center) and were inhibited by light falling in the surrounding field (surround), whereas OFF cells responded in the opposite manner. On/off ganglion cells were briefly excited when light was turned on or off.
inhibitory interneuron - produce an inhibitors that bind to receptors on excited neurons. The binding of the inhibitor blocks the neuron from releasing the acetylcholine that is responsible for muscle contraction. As a result, the muscle relaxes.
retinal ganglion cell - A neuron located in the retina that receives visual information from bipolar cells; it's axons give rise to the optic nerve.
Retinal ganglion cells calculate color
The purpose of the ganglion cells is not fully known, but they are involved in color vision. Ganglion cells compare signals from any different cones
The ganglion cells add and subtract signals from many cones. For example, by comparing the response of the middle-wavelength and long-wavelength cones, a ganglion cell determines the amount of green-or-red. Moreover, they are excited in the middle of their field, and inhibited in the surround, which makes them particularly sensitive to edges.
The result of all these steps for color vision, is a signal which is sent to the brain. There are three signals, corresponding to the color attributes. These are:
the amount of green-or-red
the amount of blue-or-yellow
and the brightness.
One of the characteristic of the receptive fields of ganglion cells is their center-surround organization -enhances our ability to detect the outlines of objects even when contrast between the background is low. (The cells whose centers are located in the brighter region but whose surrounds are located at least partially in the darker region will have the highest rate of firing)
spatial summation in the receptive field (optimal stimulus for cell activation)
spatial summation: then integration of signals that occur at different sites on the neuron
lateral inhibition
Some photoreceptors are activated when they detect light, while others are activated in the absence of light. These two types usually encircle each other and are spread throughout the retina creating receptive fields. The image to the right is a depiction of how photoreceptors and receptive fields are arranged.
These fields can be called either ON-Center or OFF-Center depending on whether the photoreceptors in the center region become activated in the presence of light
Often, light can fall onto both light and dark photoreceptors causing the two regions to compete with one another. One part of the receptive field wants to become active while the other part does not. This competitive interaction is called Lateral Inhibition
Mach bands - Most sensory tissue (retina of the eye, cochlea of the ear, pressure sensitive nerves of the skin) and even portions of the brain, is organized so that stimulation of any given location produces inhibition of the surrounding nerve fibers, the effect, called lateral inhibition, is to (mathematically) differentiate the signal being processed (or high pass filter). In the case of visual information, such (spatial) differentiation causes gradual changes in the contrast between an object and its background to be enhanced, i.e., to become more visible. In human perception, this contrast enhancement produces what is known as Mach Bands: between two regions of different intensity a thin bright band appears at the lighter side and a thin dark band appears on the darker size. These bands are not physically present are "overshoot" and "undershoot" caused by our neural circuits in processing a step discontinuity in illumination
feed-forward and recurrent inhibition - Nearly all sensory information arrives in the cerebral cortex via the thalamus, thus making the thalamocortical synapse our gateway to consciousness. Interestingly, inhibitory interneurons, which are a minority (~20%) of all cortical neurons, are much more reliably excited by incoming sensory inputs, compared to excitatory neurons. These inhibitory interneurons, which use GABA as a neurotransmitter, then relay “feedforward” inhibition onto neighboring excitatory neurons. Feedforward inhibition is thought to be critical for focusing our attention on the event of interest, at the expense of various background distractions. Inhibitory interneurons are highly variable, and can be classified into anywhere from 5 to 50 subtypes, depending on investigator
excitatory and inhibitory feedback loops
synapse types: axosomatic, axodendritic and axoaxonal - a junction between the terminal buton o fan axon and the terminal membrane of another neuron( small gap separating neurons). Axon can make contact anywhere along the second neuron
1. Axodendritic Synapse (axodendritic synapse, one between the axon of one neuron and dendrites of another.) Axon to dendrite, Excitatory
2. Axosomatic Synapse (axosomatic synapse, one between the axon of one neuron and the body (soma) of another). Axon to cell body , Inhibitory
3. Axoaxonic Synapse ( axoaxonic synapse, one between the axon of one neuron and the axon of another neuron.). Axon to terminal endings , Presynaptic inhibition , Reduced polarity.
presynaptic inhibition - The action of a presynaptic terminal buton in an axoaxonic synapse; reduces the amount of neurotransmitter released by the postsynaptic terminal button so if the activity of axoaxonic synapse decreases the release of the neurotransmitter, the effect is called PRESYNAPTIC INHIBITION.
Inhibitory synapse onto presynaptic terminal of excitatory synapse
Post-synaptic inhibition momentarily reduces ability of cell to respond to excitatory input,
Presynaptic inhibition specific to particular input, without affecting response to others. Example: stimulation of hairs on tail of insect normally triggers escape behavior, but must be suppressed during mating, without suppressing response to other dangerous stimuli
Presynaptic inhibition reduces amount of neurotransmitter released from an excitatory terminal
Works by reducing Ca2+ influx into presynaptic terminal, therefore reducing amount of neurotransmitter released
convergence and divergence - Komórki nerwowe zorganizowane są w obwodach!! Ośrodkowy układ nerwowy (CNS) zbudowany jest z milionów komórek , lecz nie jest bynajmniej chaotycznie splątaną ich masą. Komórki nerwowe są zorganizowane w sieciach neuronów, a w obrębie sieci-w specyficznych szlakach, czyli obwodach nerwowych. Chociaż każda z sieci ma pewne szczególne właściwości, struktura i organizacja wszystkich obwodów nerwowych jest podobna. I tak na przykład, powszechną ich właściwością jest zdolność do konwergencji i dywergencji ( convergence , divergence ):
KONWERGENCJA- polega na tym, że pojedyncza komórka nerwowa otrzymuje pobudzenie z dwóch lub z wielu włókien presynaptycznych. Na przykład w neuronach pośredniczących (interneurons) w rdzeniu kręgowym mogą zbiegać się inform. z neuronów czuciowych (sensory neurons), z neuronów dostarczających inform. z różnych części mózgu i z neuronów pochodzących z różnych części rdzenia. Informacje napływające ze wszystkich tych źródeł są integrowane, zanim rozprzestrzeni się potencjał czynnościowy i zanim nastąpi podrażnienie odpowiedniego neuronu ruchowego (motor neuron). Konwergencja jest więc ważnym mechanizmem, za pomocą którego ośrodkowy układ nerwowy (CNS) koordynuje informacje z rozmaitych źródeł.
DYWERGENCJA z kolei polega na tym, że pojedyncze włókno presynaptyczne pobudza przewodzenie impulsu w wielu neuronach postsynaptycznych. Każdy neuron presynaptyczny może rozgałęziać się i stykać synaptycznie aż z 25 000 ( i większą liczbą) neuronów postsynaptycznych. Np. pojedynczy przewodzący impuls z ośrodków ruchowych w mózgu może synaptycznie stykać się z setkami neuronów pośredniczących ( interneurons)w rdzeniu kręgowym, a każdy z nich może rozgałęziać się dalej i przekazywać pobudzenie do setek włókien mięśniowych
simple receptive fields of primary visual cortex (V1) neuron - V1 has a topographic/retinotopic map of the visual world (see above). This means that there is a "neural image" that retains the spatial layout of the pattern of light that falls on the retina. This map has several interesting characteristics:
Remember that there are 2 V1s in each person (left and right hemispheres). Each V1 has a representation of the opposite half of the visual field (e.g., left V1 has a map of the right visual field, and vice versa). Note that each V1 does not simply receive input from the opposite eye. The outputs of each retina are split (left half/right half) and then run through the LGN to the appropriate V1.This diagram of the visual fields is helpful
Just as othe image of the world is inverted when projected onto the retina, the retinotopic V1 map is upside down. As discussed earlier, the right hemisphere's V1 has a topographic map of the left visual field, and vice versa
Cortical magnification: more cortical space is dedicated to the fovea than the periphery (remember the higher density of photoreceptors in the fovea, hence clearer vision).
Hebb's ensemble
post-stimulus-time-histogram (PSTH) - As with the amplitude Histogram, the Post-Stimulus Time Histogram (PSTH) is not really a waveform average. Instead, it shows the temporal distribution of threshold events. The X-axis shows the same time span as a conventional waveform display. At each sample time point, the Y-height shows the percent of sweeps that contained a threshold event at that point. Threshold events are those that have the specified slope polarity when they cross the specified threshold, as set by the controls beneath the PSTH selector button. With a repetitive waveform that has Tone Sync active, the default positive slope and zero threshold level will result in a threshold event at each positive zero crossing. If the wave repeats exactly on each sweep, the PSTH will show a 100% spike at each of these times. If there is a little jitter in the timing, the peaks may be broader and of lower amplitude (since 100% of the sweeps don't have an event that lands on the same time point.) With a lot of jitter or with no sync, the peaks may appear at random positions and at very low levels. The PSTH will allow you to quickly pick out timing problems. If the input is a pulse train, this will easily show skipped or extra pulses. Note that the threshold detector needs a "running start" to determine the slope. So if the threshold is set to zero and the waveform phase is such that each sweep starts exactly at zero, the detector will not recognize the first sample as a threshold event. If you want to count it, you should set the threshold level a little above zero, say 2% or so. Or you can set the main tone Phase to start a little before the zero crossing, say -10 degrees or so. PSTH mode behaves like Linear or Histogram in that it accumulates a discrete number of sweeps as set by the Sweeps Request. One typical use for a PSTH is in electrophysiological studies. Neurons typically have some baseline "spontaneous" firing rate; with no stimulus applied, the PSTH would show a low-level random pattern, because there would be no correlation between the neural discharge spikes and any stimulus signal. Now if a stimulus is applied, such as a tone burst that repeats once per sweep, the neuron will be much more likely to discharge at the start of the stimulus. It won't necessarily fire at the start of each and every stimulus, since it may have fired spontaneously just before that, and not yet recovered. But there should be a big peak in the PSTH at the stimulus time and shortly thereafter. During the time the stimulus remains on, the neuron may show an elevated firing rate. The PSTH would show a lower-level region after the onset peak, decaying after the stimulus offset. For some acoustic neurons, the discharge spikes from an ongoing tone are more likely to occur at a certain phase of the tone. The particular phase won't be known in advance, since there will be acoustical and neural delays between the stimulus generator and the neural spike mechanism. But this phase-locking behavior will show up as a series of peaks or clumps in the ongoing portion of the PSTH.
corellogram
Brain
Spinal Cord
The spinal cord is the main pathway for information connecting the brain and peripheral nervous system. The human spinal cord is protected by the bony spinal column. The spinal column is made up of bones called vertebrae.
The spinal cord is located in the vertebral foramen and is made up of 31 segments: 8 cervical, 12 thoracic, 5 lumbar, 5 sacral and 1 coccygeal. A pair of spinal nerves exits from each segment of the spinal cord.
The spinal cord is about 45 cm long in men and 43 cm long in women. The length of the spinal cord is much shorter than the length of the bony spinal column. In fact, the spinal cord extends down to only the last of the thoracic vertebrae. Therefore, nerves that branch from the spinal cord from the lumbar and sacral levels must run in the vertebral canal for a distance before they exit the vertebral column. This collection of nerves in the vertebral canal is called the cauda equina (which means "horse tail").
Receptors in the skin send information to the spinal cord through the spinal nerves. The cell bodies for these nerve fibers are located in the dorsal root ganglion. The nerve fibers enter the spinal cord through the dorsal root. Some fibers make synapses with other neurons in the dorsal horn, while others continue up to the brain. Many cell bodies in the ventral horn of the spinal cord send axons through the ventral root to muscles to control movement.
brain stem (arousal)
The portion of the brain closest to the spinal cord. It consists of the medulla, pons, and midbrain and controls many of the involuntary functions that keep us alive.
thalamus
A collection of nerve cells in the brain. Although it performs many functions, the primary role of the thalamus is to relay sensory information from other parts of the brain to the cerebral cortex.
lateral geniculate body - The “visual middleman,” the lateral geniculate body receives information from the eyes via the optic tract, does a preliminary analysis of the information, and then sends it onto the visual cortex.
nucleus reticularis thalami* (*nRt*) - covers the lateral surfaces of the thalamus. Through it pass nearly all the pathways coursing between the thalamus and cerebral hemispheres. Via these reciprocal nRt circuits, the cerebral cortex and brainstem reticular formation _selectively modulate their own information processing activities_ in the service of conscious perceptions, intentions and/or plans.
Striatum (sensory-motor coordination, automatic movements)
Part of the basal ganglia of the brain. The basal ganglia are interconnected masses of gray matter located in the interior regions of the cerebral hemispheres and in the upper part of the brainstem. The striatum is also called the striate body. It includes the caudate nucleus and the lentiform nucleus. The term striatum (striate) refers to the layered appearance of bands of gray matter that course from one of these nuclei to the other and represent the connections between them.
Cerebellum (correcting movement trajectories)
Major region of the brain concerned with coordinating movements. The cerebellum is where learned movements are stored. The “little brain” attached to the brain stem. The cerebellum is responsible for coordinated movements. Attached to the brain at the rear of the skull, the twin lobes of the cerebellum coordinate movement, control posture, and maintain equilibrium.
cerebral cortex - the part of the brain that is visible from the outside. The cerebral cortex is the outermost layer and consists of a collection of nerve cell bodies. The thin gray matter covering of the cerebrum, consisting of a ¼-inch layer dense with cell bodies of neurons. The cerebral cortex carries on the major portion of our “higher” mental processing, including thinking & perceiving.
association cortex - cortical areas that are neither motor or sensory but are thought to be involved in higher processing of information
techniques measuring brain activity
CT(computerized tomography) - a method of examining body organs by scanning them with X rays and using a computer to construct a series of cross-sectional scans along a single axis. Tomography detects the soft-tissue structures of the brain that normal X rays do not reveal.
PET(positron emission tomography) - using a computerized radiographic technique to examine the metabolic activity in various tissues (especially in the brain)
Microelectrodes
LFP
fMRI - a form of magnetic resonance imaging of the brain that registers blood flow to functioning areas of the brain
EEG - a graphical record of electrical activity of the brain waves, picked up by electrodes posted on the scalp(without opening the skull); produced by an electroencephalograph
Visual system
retina - A layer of light-sensitive receptors at the back of the eye that forwards information to the optic tract and eventually to the brain. Cells:
photoreceptor - A cell or group of cells that can sense and receive light
Rods - especially sensitive to dim light but not to colors; 125 million thin rods
Cones - especially sensitive to colors but not to dim light; 7 million fat cones
Bipolar cells - other types of cells in retina that do not respond directly to light. Instead they gather the responses from groups of nearby receptors; they collect impulses from many photoreceptors and send them to the ganglion cells.
Horizontal cells Also called cells of Cajal, horizontal cells are found in the outermost layer of the cerebrum. They extend their receptive dendrite fibers laterally along the surface of the gray matter. Their primary function is correlation, or providing linkages for cells distant from each other.
ganglion - A group of nerve cell bodies in the Central Nervous System (CNS) or Peripheral Nervous System (PNS). (Plural form: ganglia.)
horizontal cell - inhibitory interneuron
horizontal cell: a neuron in the retina that interconnects adjacent photoreceptorsand the outer processes of the bipolar cells
interneuron: a neuron located entirely within the central nervous system.
On - and Off-center receptive fields
These cells are primarily concerned not with assessing levels of illumination but rather with making a comparison between light level in one small area of the visual scene and the average illumination of the immediate surroundings. Some retina ganglion cells have an ON and an OFF response, signalling the overall illumination of the visual scene and are important for pupillary reflexes. In the visual cortex, however, the situation is far more complex. There is a wide variety of cell types: some are orientation selective cells, named simple cells; some have both orientation and direction selective receptive fields and are named complex cells; some even respond to a particular length or width and they are named hypercomplex cells or end-stopped cells. The properties of these feature sensitive cells play a fundamental role in theories of visual perception.
visual acuity - sharpness of vision; the visual ability to resolve fine detail (usually measured by a Snellen chart
parallel processing of stimuli attributes (channels: parvo- (P) and magno-cellular (M)
parvocellural pathway: one of the four outer layers of neurons in the dorsal lateral geniculate nucleus; transmits information necessary for perception of color and fine details to the primary visual cortex.
magnocellural pathway: one of the inner two layers of neurons in the dorsal lateral geniculate nucleus; transmits information necessary for the perception of form, movement, depth, and small differences in brightness to the primary visual cortex.
Retinotopy: the notion that receptor cells in the retina are mapped to points on the surface of the visual cortex of the brain
Primary visual cortical area-V1 Visual processing area of cortex in the occipital lobes. Processes sensations from the eyes. Input from the center part of the retina at the back of the eye, the area that transmits the most detailed visual information, chains the greatest proportion of the visual cortex. Also within this region are highly specialized areas that process color, movement, shape, shading, and other aspects of visual stimulation, including a patch of cortex responsible for the recognition of human faces.
Cortical columns:
V1 Cortical Columns The term "cortical column" refers to the notion that cells arranged vertically from the surface of the cortex to the white matter might comprise functional or anatomical units. Thus, a cortical column can be defined on the basis of anatomical features (e.g. stereotyped patterns of pyramidal cell apical dendrite bundles), functional features (e.g. columns of cortical cells all responding to the same stimulus orientation) or both. Many types of columns have been proposed including ocular dominance, orientation, spatial frequency, and color columns. The details of these columnar arrangements will be described in later sections dealing with the physiology of V1. However, I will here briefly describe the ocular dominance columns in order to provide an example of columnar arrangement in the primary visual cortex.
Ocular Dominance Columns. Visual signals from the two eyes remain segregated in the LGNd (Figure 11) and in the geniculorecipient layers of area V1 (Figure 14). One can observe this segregation by measuring the electrophysiological responses of the units in layer 4C. As the recording electrode is moved within layer 4C, there is an abrupt shift as to which eye drives the unit. In layer 4C, the shift from one eye to the other takes place over a distance of less than 50 microns. Signals from these bands converge on individual neurons in the superficial layers of the cortex, thereby forming columns dominated by one eye or the other in an alternating fashion.
blobs (color processing, V1) peglike, cytochrome oxidase-rich, dual-opponent color columns
hypercolumn
higher level areas: V2, V3, V4, V5
Visual area V2 is the second major area in the occipital lobe of the cerebral cortex. It receives strong feedforward connections from Primary visual cortex (V1) and sends strong connections to Visual area V4 and Visual area MT. It also sends strong feedback connections to the Primary visual cortex. Anatomically, V2 is split into four quadrants, a dorsal and ventral representation in the left and the right hemispheres. Together these four regions provide a complete map of the visual world. Functionally, V2 has many properties in common with Primary visual cortex. Cells are tuned to simple properties such as orientation, spatial frequency, and color. Recent research has shown that V2 cells are tuned to the orientation of illusory contours, shows a small amount of attentional modulation (more than V1, less than V4), are tuned for moderately complex patterns, and may be driven by multiple orientations at different subregions within a single receptive field.
Area V2 has a long common border with V1. It receives a somewhat patchy input from V1 and has a rather disorderly topographic organisation. Interestingly the mapping of the visual field onto V2 is the mirror-image of the V1 map.
Like V1 the distribution of cytochrome oxidase in V2 is of great interest. Staining for this metabolic enzyme reveals a pattern of alternating thick and thin cytochrome oxidase stripes, each separated by a thin interstripe region.
The thick stripes (drawn in yellow)are clearly a part of the magnocellular pathway. They receive input from layer 4B of V1 and they project to V3 and MT.
The thin stripes (red), and the interstripes (grey), receive input from the blobs and interblob regions of layers 2 & 3 of V1. They then project to V4, thus are a part of the parvocellular pathway.
Although it may be fashionable to talk of the M and P pathways as if they were completely separate this is clearly not the case. For example, there are connections between the thin stripes and the thick stripes of V2 and there is also a projection from V4 back to the thick stripes. There are also direct connections between MT and V4 and V3.
Visual area V3 This area (colored blue in the picture) receives inputs from the thick stripes in V2, and from layer 4B in V1. Only the lower part of the visual field is represented in V3 (a corresponding area representing the upper part is no longer considered part of V3, but has been given a different name, ventral posterior, or VP). Properties of cells in V3 offer few clues as to its function. Most are selective for orientation, and many are also tuned to motion and to depth. Relatively few are color sensitive.
Visual area V4 This area was originally discovered by Semir Zeki. It receives input mainly from the thin and interstripe regions of V2, but also has connections from V1 and V3.
Although the area contains many cells that are colour selective, indicating a role in colour analysis, cells are also found with complex spatial and orientation tuning, suggesting that the area is also important for spatial vision.
Visual area V5 The middle temporal area is sometimes called V5. It receives connections from layers 4B and 6 in V1, and from thick stripes in V2. These connections are ultimately linked with the M pathway from the LGN.
Most cells in MT are tuned to motion, and the area can be divided into direction and 'axis of motion' columns. The connections from V2 are puzzling, because few cells there are motion selective. Nevertheless, many agree that MT is intimately linked with the analysis of image motion.
inferotemporal cortex
neuropsychology
IT lesions cause agnosia (“psychic blindness”)
monkeys and humans
neurophysiology
neurons are selective to complex features
high degree of spatial invariance
Neurons in inferotemporal cortex respond only to visual stimuli and a majority have receptive fields that extend well into both visual half-fields. After bilateral removal of striate cortex, no inferotemporal neurons responded to visual stimuli. After unilateral removal of striate cortex, inferotemporal neurons in both hemispheres responded only to stimuli in the hemifield contralateral to the intact striate cortex. After section of the corpus callosum and anterior commissure, inferotemporal neurons in both hemispheres responded only to stimuli in the hemifield contralateral to the recording site. These results indicate that inferotemporal cortex visual information from striate cortex and that the pathway from striate cortex to the contralateral inferotemporal cortex includes the forebrain commissures. This same striate-temporal pathway is also necessary for normal discrimination learning. We suggest that the converging input onto single inferotemporal neurons from widely separated retinal areas may provide a mechanism for stimulus equivalence over different parts of the visual field, and it may be the absence of such a mechanism that contributes to the visual discrimination deficit that follows inferotemporal lesions.
gnostic unit gnostic modules (Konorski 1969), established through learning, to play a fundamental role as higher-order primitives/building blocks in the perceptual representation of visual forms
Konorski (1967): concept of gnostic units, i.e. cells which respond to one specific object. Later known as `grandmother cells,' as it was thought there may be a cell representing one's grandma
gnostic theory (J. Konorski) Konorski (1967) had referred to `gnostic units', which makes these cells sound as though they are semantic and not just visual entities; however, Konorski divided up the cortex purely into sensory, motor, and emotion `analyzers' with no regions of multimodal or amodal responsiveness. Grandmother's face cells would therefore be found in the visual analyzer and grandmother's voice cells in the auditory analyzer, with associative links running directly between them
area V4, „color constancy”
area V5 (MT), responses for moving stimuli
depth perception
The eye uses three methods to determine distance:
The size a known object has on your retina - If you have knowledge of the size of an object from previous experience, then your brain can gauge the distance based on the size of the object on the retina.
Moving parallax - When you move your head from side to side, objects that are close to you move rapidly across your retina. However, objects that are far away move very little. In this way, your brain can tell roughly how far something is from you.
Stereo vision - Each eye receives a different image of an object on its retina because each eye is about 2 inches apart. This is especially true when an object is close to your eyes. This is less useful when objects are far away because the images on the retina become more identical the farther they are from your eyes.
A classic problem in visual perception is how we come to perceive a three-dimensional world. Our retinal images are flat, after all. There are a wide variety of depth cues or pieces of information within flat retinal images that help us infer the correct three-dimensional structure of the visual world
Pictorial cues to depth
Interposition or occlusion is said to occur when one object obscures a more distant one. The object that is blocking the view of the other object is closer
Linear perspective was first used systematically in fine art during the Renaissance. Painters wanted to represent lights from the three-dimensional world on a flat canvas. Lights are projected onto the two-dimensional canvas according to the laws of linear perspective, illustrated below in this work of Albrecht Durer
Relative size refers the comparison of retinal image sizes of identical or nearly-identical objects. The sizes of the retinal images from two adults of about the same height, for example, readily inform the viewer which adult is closer (the adult with the larger retinal image is closer, of course!).
Texture gradients can be used to determine the way that a surface with a uniform surface is receding away from the viewer.
In general, many of these cues to depth operate simultaneously
Depth from motion As mentioned above, information about objects' depths is also available from the way their retinal images move
Motion parallax. Imagine you are driving along in a car and looking out the side window. Suppose further that there are trees at various distances. Now the trees that are close to the road appear to zip by, but the trees that are off in the distance move much more slowly. This motion in the retinal image induced by the viewer's motion is called motion parallax, and it is clear that the amount of motion parallax is an excellent cue to depth: lots of motion parallax means close, little motion parallax means far away
Optic flow. As the observer moves through the environment, things move.
- When the observer looks directly at a point and then starts moving towards it, the images of things close to the point appear to diverge from the point: the retinal image seems to expand.
- When the observer looks directly at some point and then moves away it, the images of things appear to move towards the point or converge.
- When an object (often fast-moving!) travels towards one's eyes, its retinal image expands in an often alarmingly rapid way! This expansion is called looming.
binocular disparity
Binocular cues Binocular cues that are available through comparison of the (different) images seen by the two eyes.
Disparity is a binocular cue to depth. Disparity refers to disparity in the retinal image locations of a single point in the three-dimensional world. This disparity is illustrated in the figure below.
Suppose that the two eyes are converged on the central point. Then the images of the central point fall on the retina of the left eye and the retina of the right eye and there is no disparity in retinal image location. A point that is farther away will produce images that are both displaced towards the nose (nasally)--this is called far disparity. A point that is nearer to the eyes will produce images that are both displaced away towards the ears (temporally)--this is called near disparity. Evidently, disparity is a strong cue to depth. In monkeys at least, there are neurons in visual area V2 (which neighbors V1) that are sensitivity to disparity.
Now a person who can use disparity information to create a fused, three-dimensional percept is said to possess stereo vision. Such a person can have fun with the variety of stereograms that people have made over the past several years. Indeed, a host of stereograms can be retrieved off the Net by the adventurous. Click here to try!
Monocular cues There are stereoblind people who cannot fuse the two eyes' images from stereograms to perceive variations in depth. Yet in everyday viewing they certainly do see depth. There are many cues to depth that are available from a single retinal image. People with stereo vision can easily convince themselves of this simply by covering one eye and looking around. Depth perception is barely altered. Static cues are ones that exist within a single image to a single eye; these are sometimes called pictorial cues. Motion cues are ones that can be taken advantage of by comparing images from a sequence of "frames" presented to a single eye; depth is inferred from the types of motion evident in the image.
Notice that the tree and the policeman are about the same size on the retina. One would undoubtedly perceive the tree to be further away than the policeman. There are multiple reasons for this. One, of course, is that most trees, this well developed, are much larger than people. Consequently knowing that the size of the tree is likely to be much larger than the policeman helps to make it appear further away. In the disparity diagram the eyes are fixated on the tree. The red lines between the tree and the retinas describe the angle of convergence the eyes make when the tree is imaged on both foveae. It is said that the tree is imaged on corresponding points of the retina and thus there is no disparity. By corresponding points I mean those points that would be identical if one retina was moved over to superimpose the other retina. Because the eye are fixated on the tree, the policeman, who is closer to the observer, has its image falling on non-corresponding points of the retinas. That is to say its images are binocular disparate. This binocular disparity provides an additional cue to the distance of the policeman relative to the distance of the tree. As an aside, if images are too disparate on the retina one will perceive two images instead of one (double vision). You can create this for yourself by holding your two pointing fingers in front of your face. Hold one so that it is behind the other. Now fixate on the finger in the front. You will notice that if the second finger is no too far behind the first it will appear as a single finger. But as you move the second finger further way from the first it will appear double. Now fixate the second finger while it is a this distant position. Note, that now it appears singular but the finger closer to your face now appears double. The single appear finger is on corresponding points of your retinas the one that appears double is not. The visual system can tolerate a certain amount of disparity without causing double vision. As long as two images lie within what is called Panum's area disparate images will not appear double.
ventral pathway (occipito-temporal [what?]) carries information about static object properties such as shape and colour. A hierarchical Pathway runs from V1 to V2, V4 and the inferior temporal cortex. There are subsequent links in occipitotemporal pathway to limbic structures. This permits cognitive association of objects with other events, e.g. motor, emotions.
dorsal pathway (occipito-parietal [where? how?]) carries information about dynamic object properties such as motion and spatial relationships. A hierarchical pathway runs from V1 to V2, MT and the Parietal lobe. There are subsequent links in occipitoparietal pathway to dorsal limbic and dorsal frontal cortical structures. Latter stages become polysensory.
binding theory, synchronization of oscillatory activities
Other Sensory Systems
Skin receptors
Nociceptors - pain receptors- Networks of free nerve endings in the skin ( 3 types):
high- threshold mechanoreceptors - free nerve endings that respond to intense pressure caused by striking, pinching, or stretching skin
respond to extreme heat, acids, capsaicin receptors (chile peppers' ingredient)- VR1 receptor- ionotropic receptor that contain a cation channel
receptors sensitive to ATP- sodium and calcium ionchannels; cause angina, migraine, damage to muscles, cancer
Skin mechanoreceptors
Sensitivity to pressure and vibration is caused by movement of the skin
Pacinis corpsule- when it is bent relatively to the axon, the membrane becomes depolarized- action potential. The axon responds briefly when the organ is bent and again when it is released. The organ is sensitive to vibration but not to steady pressure. The bending of the tip of the nerve ending opens ion channels in the membrane
Skin receptors rarely act alone; contact with skin usually activates multiple classes of receptors, total pattern of responses yields sensations
Fibers carrying info to CNS vary in conduction velocity, A alpha fibers fastest, A beta, A gamma, and A delta are successively slower, all A fibers myelinated, convey information about skin contact, cold, and sharp pain, C fibers slow, non-myelinated, mediate touch and tickle, hot, and dull, burning pain
Thermoreceptors - thermal receptors which respond to warmth and coolness
Sodium - potassium pump is responsible for sensory transduction in coolness receptors. A drop in temperature slows the action of the pump, which allows sodium to accumulate in the free nerve ending and depolarize its membrane. Warmth- deep and cold- close to surface -receptors lay in different layers of skin
Proprioceptors - On the basis of what environmental stimulus they detect, DRG (Dorsal Root Ganglia) neurons (the neurons that allow us to sense balance and coordination, pressure and vibration, pain and temperature) are functionally classified as PROPRIOCEPTORS, low-threshold mechano receptors, and cells that sense pain and/or temperature. Proprioception is an automatic sensitivity mechanism in the body that sends messages through the central nervous system (CNS). The CNS then relays information to the rest of the body about how to react and with what amount of tension
Muscle spindles are receptors that actually contain muscle fibers enveloped in a connective tissue sheath. Nerve fibers are wrapped around the fibers to detect the degree of stretch on the muscle. Muscle spindles are unique as receptors because they are end organs which sense distortion but they can also distort themselves. Also called intrafusal muscle fibers - specialized sensory organs that are served by two axons: sensory and motor
Tendon receptor (tissue) - Connect muscle and bone, transmit contractile forces to bone. Tendons in areas that bend sharply, such as the tendons which cause the fingers to bend (flexor digitorum) pass through a tendon sheath that directs the path of the tendon.
Afferent pathway
Afferent fibers originate at nociceptors: transmit pain sensations to the dorsal horn of the spinal cord - substantia gelatinosa laminae. Pain sensation is transmitted across to the other side of the spinal cord then through spinothalamic tracts to thalamus
Afferent fibers- A-delta fibers - myelinated, large A-delta fibers; transfer pain sensations more rapidly, transmit well localized, sharp pain sensations
Small A-delta or C fibers : C fibers are small and unmyelinated and transmit pain sensations slowly, C fibers transmit diffuse burning or aching sensations.
Efferent pathway responsible for modulation (inhibition) of pain impulses, stimulation of the periaqueductal gray (PAG) site in the midbrain results in activation of the efferent pathway
Efferent nerves: nerves that carry motor signals from the central nervous system to the skeletal muscles or internal organs.
Hierarchical processing of sensory information- sensory processing of incoming information essentially is hierarchical, i.e., consisting of a chain of abstracting steps by which the multitude of primary sensory cues is organized into a subjective representation of external objects. It appears that hierarchical processing principally operates on metathetic sensory attributes - as opposed to the prothetic ones. (Metathetic attributes are related to "what, and where"; prothetic attributes to "how much".) Primary methatetic attributes are the contours in vision and the spectral pitches in audition. Understanding the perception of pitch and several related auditory phenomena essentially was enabled by, and is crucially dependent on, the hierarchical approach
Functional columns in somatosensory cortex- 1) Dorsal Column-Medial Lemniscus Pathway- primarily associated with low-threshold mechanosensation (fine touch) as well as proprioception. This phylogenetically recent pathway can transmit information to the cortex with a minimum of synapses involving three neurons. To sending several axon terminals to arborize within the dorsal horn, many primary sensory afferents (largely, A ; A fibers), have a central ascending axon within the dorsal column, which terminate within one of the dorsal column nuclei, located in the dorsal medulla. The dorsal column also contains axons of second-order neurons. The cell bodies are located medial aspects of the dorsal horn and their axons terminate in either the cuneate or gracile nucleus, depending on segment of origin. In cats, at least, the functions of dorsal columns have been suggested to play a role in nociception. 2) Anterior Lateral System- The central pathways associated with pain (nociceptive) and temperature are much more complex than the dorsal column medial lemniscus system. Route: ascends anterior lateral quadrant; largely crossed with some fibers ascending ipsilaterally. Some fibers terminate only in the reticular formation of the brainstem; others send collaterals to the reticular formation and the thalamus. Route: Fibers terminate in the midbrain reticular formation, lateral part of the periaqueductal grey (reciprocally interconnected with the limbic system via the hypothalamus) and other midbrain sites. Route: Axons ascend the ipsilateral dorsal lateral funiculus terminating in the lateral cervical nucleus (LCN). LCN neurons cross the midline and join the medial lemniscal fibers. Collaterals to midbrain nuclei. Route: ipsilateral dorsal columns terminating in the DCN.
Projectional maps - The parameter that varies within the map is a directly perceivable sensory parameter, such as the position of a tactile stimulus on your skin. Such maps can be set up within a CNS by the orderly projection of the axons of the "output" cells in the sensory epithelium into the "map" region, with local topographical relationships (i.e., nearest neighbor relationships) between the axons maintained
Primary Somatosensory Cortex (S1) - Parts of the cerebral cortex in the parietal lobe are involved with processing information related to touch. One such area is the primary somatosensory cortex which is located behind the central sulcus. Neurons in the primary somatosensory are activated when the skin is touched. However, the body is NOT represented in the cortex in proportion to the amount of skin. The face and hands take up a good portion of the primary somatosensory cortex. This is because the amount of primary somatosensory cortex is directly related to the sensitivity of a body area and the density of receptors found in different parts of the body. The areas of skin with the higher density of receptors (like the face, hands and fingers) have more cortical tissue devoted to them. When you touch the screen, an electrical impulse is sent down the length of your finger, down your hand and arm, up your spinal cord to your brain. Within the somatosensory cortex is a representation of the human body called the homunculus or „little man". Neurons in this location can identify the area of the body being stimulated by the information they receive from the somatic receptors in the skin
Homunculus - Within the somatosensory cortex is a representation of the human body called the homunculus or „little man". Neurons in this location can identify the area of the body being stimulated by the information they receive from the somatic receptors in the skin
S1
Motor System
Motoneurons:
Alpha motor neurons- neurons whose axons form synapses with extrafusal muscle fibers (responsible for the force exerted by contraction of a skeletal muscle) of a skeletal muscle; activation contracts the muscle fibers. Contraction of these fibers provides the muscle's motive force
Gamma motor neuron ( has efferent axos) causes the intrafusal muscle fibers (sensory organs that are served by 2 axons: sensory and motor) to contract; however, this contraction contributes an insubstantial amount of force. The function of this contraction is to modify the sensitivity of the fiber's afferent ending to stretch
Monosynaptic stretch reflex - A reflex in which a muscle contracts in response to its being quickly stretched; involves a sensory neuron and a motor neuron, with one synapse between them. Starting at the muscle spindle, afferent impulses are conducted to terminal buttons in the gray matter of the spinal cord. These terminal buttons synapse on an alpha motor neuron that innervates (unerwiac) the extrafusal muscle fibers of the same muscle. Only one synapse is encountered along the route from the receptor to effector- hence the term monosynaptic. When a person is asked to move his/ her leg as quickly as possible after being touched on the knee, the response is not reflexive but it involves sensory and motor mechanisms of the brain
Muscle:
Antagonist - a muscle whose contraction resists or reverses a particular movement. The agonist moves the limb in the being studied , and because muscles cannot push back, the antagonist muscle must move the limb back in the opposite direction. When a stretch reflex is elicited in the agonist, it contracts quickly, thus causing the antagonist to lengthen. The antagonist is presented with a stimulus that should elicit its stretch reflex. When the afferent axons of intrafusal muscle fibers fire, they excite inhibitory interneurons that slow the rate of firing of the alpha motor neurons that serve the antagonistic muscles, causing antagonist to relax and the agonist to contract
Synergistic muscle - …
Inhibition of antagonistic muscles by interneurons
The role of cerebellum
The cerebellum (“little brain”), with its two hemispheres, resembles a miniature version of the cerebrum. It is covered by the cerebellar cortex and has a set of deep cerebellar nuclei. These nuclei receive projections from the cerebellar cortex and themselves send projections out of the cerebellum to other parts of the brain. Each hemisphere of the cerebellum is attached to the dorsal surface of the pons by bundles of axons: the superior, middle, and inferior cerebellar.
Damage to the cerebellum impairs standing, walking or performance of coordinated movements. The cerebellum receives visual, auditory, vestibular, and somatosensory information, and it also receives information about individual muscle movements being directed by the brain. The cerebellum integrates this information and modifies the motor outflow, exerting a coordinating and smoothing effect on the movements. Cerebellar damage resuls in jerky, poorly coordinated, exaggerated movements; extensive cerebellar damage makes it impossible even to stand.
Cerebellum - a major part of the brain located dorsal to the pons, containing the two cerebellar hemispheres, covered with the cerebellar cortex; an important component of the motor system.
The role of basal ganglia
The basal ganglia are a collection of subcortical nuclei in the forebrain, which lie beneath the anterior portion of the lateral ventricles. The major parts of the basal ganglia are the caudate nucleus, the putamen, and the globus pallidus ( the “nucleus with a tail”, the “shell”, and the “pale globe”). The basal ganglia are involved in the control of movement. For example, Parkinson's disease is caused degeneration of certain neurons located in the midbrain the send axons to the caudate nucleus and the putamen. The symptoms of this disease are of weakness, tremors, rigidity of the limbs, poor balance, and difficulty in initiating movements.
Basal ganglia - a group of subcortical nuclei in the telencephalon, the caudate nucleus, the globus pallidus, and the putamen, important parts of the motor system.
Postural reflexes
During the development of the foetus a group of reflexes emerge which are called primitive reflexes. These primitive reflexes should be present at birth and they provide and indication of the status of the CNS. These primitive reflexes should slowly be inhibited during the first year of life. As these primitive reflexes are inhibited, so another group of reflexes, called the postural reflexes, should emerge to help the infant or child to cope with demands of a gravity-based environment, and provide the basis for the control of automatic balance, posture and voluntary movement.
The postural reflexes are regulated by higher centres in the brain involved in the execution of voluntary movement.
It is only as postural reflexes replace primitive reflexes that the infant begins to gain control of the body and body movements. A child who has cerebral palsy never makes the transition from primitive to postural reflexes, and so movements remain random and uncontrolled. Most children in the early months of life rapidly gain control of the primitive reflexes, and thus lay the foundations for later voluntary movement.
Cortical control of movements (M1)
The primary motor cortex, or M1, is one of the principal brain areas involved in motor function. M1 is located in the frontal lobe of the brain, along a bump called the precentral gyrus . The role of the primary motor cortex is to generate neural impulses that control the execution of movement. Signals from M1 cross the bodys midline to activate skeletal muscles on the opposite side of the body, meaning that the left hemisphere of the brain controls the right side of the body, and the right hemisphere controls the left side of the body. Every part of the body is represented in the primary motor cortex, and these representations are arranged somatotopically -- the foot is next to the leg which is next to the trunk which is next to the arm and the hand. The amount of brain matter devoted to any particular body part represents the amount of control that the primary motor cortex has over that body part. For example, a lot of cortical space is required to control the complex movements of the hand and fingers, and these body parts have larger representations in M1 than the trunk or legs, whose muscle patterns are relatively simple. This disproportionate map of the body in the motor cortex is called the motor homunculus.
Development, Recovery From Damage
Developmental plasticity
the ability of the cells and tissue of some species to develop in response to their current environment. Examples of this are the human immune system, ahich may change throughout life, and water snails, who are known to develop according to lake or pond conditions.
Critical periods in development
As a child's brain develops, it goes through several “critical periods,” a developmental phase in which the brain requires certain environmental input or it will not develop normally. “This is a term some people outside of biology are not comfortable with. Some people prefer `sensitive periods,' which makes the issue more open-ended,” Eliot said. “At a circuitry level, we know there truly are critical periods - stages of development - in which the brain needs certain types of experience, or the circuits don't get put together properly.” Eliot cited the landmark work of David Hubel and Torsten Wiesel, who earned the 1981 Nobel Prize for Physiology for their research on the effects of early visual experience on the brain. Hubel and Wiesel studied the visual cortex of the brain to better understand how the brain processes vision. They conducted experiments on kittens, which involved suturing shut one eye and comparing the kittens with others of the same litter left alone. The sutured eye, when opened after two months, would remain functionally blind for life. The blindness was not due to a defective eye, but because the other eye had established most of the pathways in the brain to compensate. “We know there a critical period: If child or animal, from birth, does not have a normal visual experience with both eyes, he or she is at risk of having permanent visual deficits,” Eliot said. The experience of sight changes the fundamental architecture of the brain because of the brain's plasticity, Eliot said. The effect is similar in all mammals. Not only sight, but hearing, touch and the other senses and functions go through different critical periods.“What we are learning through neuroscience is that each different part of the brain carries on a different cognitive function, whether it's perception, movement, emotion, language or memory. Each has a different developmental time table, and along with that different critical periods,” she said.
Sensory deprivation (e.g. monocular - visual)
The experimental or natural reduction of environmental stimuli, as by physical isolation or loss of eyesight, often leading to cognitive, perceptual, or behavioral changes, as disorientation, delusions, or panic.
Sensory deprivation, partial to total deprivation of visual, auditory, and tactile stimuli, such as may be produced experimentally or by solitary confinement, loss of sight or hearing, paralysis, or even hospital bed rest; it may result in some combination of anxiety, irritability, boredom, loss of ability to concentrate and organize thoughts, increased suggestibility, delusions, panic, and unpleasant vivid hallucinations.
Cortical blindness - blindness caused by damage to the optic radiations or primary visual cortex
Visual agnosia - deficits in visual perception in the absence of blindness; caused by brain damage. Agnosia - “failure to know”. Inability to perceive or identify a stimulus by means of a particular sensory modality, even though its deatails can be detected by means of this modality and the person retains relatively normal intellectual capacity. Apperceptive visual agnosias are failures in high level perception (failure to perceive objects, even though visual acuity is relatively normal), whereas associative visual agnosias are disconnections between these perceptions and verbal systems (inability to identify objects that are perceived visually, even though the form of the perceived object can be drawn or matched with similar objects).
Visual ataxia - difficulty in reaching for objects under visual guidance
Learning and Memory
Brain plasticity - jedyne, co znalazłam w tym stylu to plasticity of nervous system, ale myślę, że to mniej więcej o to samo chodzi ;P plasticity - the ability of the nervous system to adapt or change as a result of experience. Plasticity sometimes can also help the nervous system adapt to physical damage.
Declarative memory - memory that can be verbally expressed, such as memory for events in a person's past, it doesn't require repetitions for consolidation. Divided into: semantic (memory of facts and ideas, knowledge of languages and conception) and episodic memory (memory of places, emotions and events from one's own life). hippocampus
Procedural memory (non-declarative memory) - memory whose formation doesn't depend on the hippocampal formation; a collective term for perceptual, stimulus-response, and motor memory. Requires repetition for consolidation. Divided into: skills (motor, cognitive, associative, perceptual) priming (of recall, reactions, associations), conditioned reflexes (classical, instrumental), nonassociative learning (habituation, sensitization)
Nonassociative learning (habituation) - habituation (learning not to respond to the repeated presentation of a stimulus)
Associative learning (classical and instrumental conditioning)
Classical conditioning - a learning procedure, when a stimulus that initially produces no particular response is followed several times by an unconditional stimulus that produces a defensive or appetitive response (the unconditional response), the first stimulus (now called a conditional stimulus) itself evokes the response (now call the conditional response)
Instrumental conditioning - the learning procedure whereby the effects of a particular behavior in a particular situation increase (reinforce) or decrease (punish) the probability of the behavior, also called operant conditioning.
Hebb's rule (for increase of synaptic weight) - the hypothesis proposed by Donald Hebb that the cellular basis of learning involves strengthening of a synapse that is repeatedly active when the postsynaptic neuron fires
LTP (long term potentiation, its specificity and associativeness) - a long-term increase in the excitability of a neuron to a particular synaptic input caused by repeated high-frequency activity of that input. Associative long-term potentiation - a LTP in which concurrent stimulation of weak and strong synapses to a given neuron strengthens the weak ones.
The NMDA receptor (NMDAR) - is an ionotropic receptor for glutamate (NMDA is a name of its selective specific agonist). NMDARs play a critical role in synaptic plasticity mechanisms and thus are necessary for several types of learning and memory.
Structure: Each NMDA receptor contains four or five subunits, the exact number has not yet been established unambiguously. The receptor oligomers are formed from members of three gene families. The subunits are called NR1 (sigma, not related to the sigma receptors), NR2 (epsilon) A-D and NR3 A-C. Each subunit has an extracellular ligand binding domain, which links to the transmembrane ion pore. The most common arrangement contains two NR1 subunits that bind glycine and two NR2 that bind glutamate. Five NMDA receptor subunits have now been characterized in both rat and mouse brains
Agonists: A unique feature of NMDA receptor is the requirement for both glutamate and the co-agonist glycine for the efficient opening of the ion channel which is a part of this receptor. A third requirement is membrane depolarization. A change in electrical potential toward equilibrium between the two sides of the membrane in which the receptor sits will make it more likely that the ion channel in the NMDA receptor will open by expelling the Mg2+ ion that blocks the channel from the outside. This property is fundamental to the role of the NMDA receptor in memory and learning, and it has been suggested that this channel is a biochemical substrate of Hebbian learning, which states that "Neurons that fire together, wire together"
Modulators: NMDA receptor is modulated by a number of endogenous and exogenous compounds. Mg2+ not only blocks the NMDA channel in a voltage-dependent manner but also potentiates NMDA-induced responses at positive membrane potentials. Na+, K+ and Ca2+ not only pass through the NMDA receptor channel but also modulate the activity of NMDA receptors. Zn2+ blocks the NMDA current in a noncompetitive and a voltage-independent manner. It has been demonstrated that polyamines do not directly activate NMDA receptors, but instead act to potentiate or inhibit glutamate-mediated responses. The activity of NMDA receptors is also strikingly sensitive to the changes in H+ concentration, and partially inhibited by the ambient concentration of H+ under physiological conditions. regulated by ion channels that are permeable to Ca2+, Na+, K+ and are sensitive to voltage-dependent Mg2+ block
Role: This channel complex contributes to excitatory synaptic transmission at sites throughout the brain and the spinal cord, and is modulated by a number of endogenous and exogenous compounds. NMDA receptors play a key role in wide range of physiologic and pathologic processes
Arousing systems in learning - Learning and memory are essential requirements for every living organism in order to cope with environmental demands, which enables it to adapt to changes in the conditions of life. Research on the effects of hormones on memory has focused on hormones such as adrenocorticotropic hormone (ACTH), glucocorticoids, vasopressin, oxytocin, epinephrine, corticotropin-releasing factor (CRF) that are released into the blood and brain following arousing or stressful experiences.Most of the information have been derived from studies on conditioned behavior, in particular, avoidance behavior in rats. In these tasks, an aversive situation was used as a stimulus for learning. Aversive stimuli are associated with the release of stress hormones and neuropeptides. Many factors play a role in different aspects of learning and memory processes. Neuropeptides not only affect attention, motivation, concentration and arousal or vigilance, but also anxiety and fear. In this way, they participate in learning and memory processes. Furthermore, neuropeptides such as CRF and vasopressin modulate the release of stress hormones such as epinephrine. In turn, systemic catecholamines enhance memory consolidation. CRF and vasopressin are colocalized in neurons from the nucleus paraventricularis, which project to nuclei in the brainstem involved in autonomic regulation. The objective of this paper is to discuss the role of CRF, vasopressin, and the autonomic nervous system (ANS) in learning and memory processes. Both CRF and vasopressin have effects in the same direction on behavior, learning and memory processes and stress responses (release of catecholamines and ACTH). These neuropeptides may act synergistically or in a concerted action aimed to learn to adapt to environmental demand
The role of emotional system in learning - Much has been made about visual learning's increasing relevance towards understanding complex concepts in science and engineering. In this media-saturated age, the visualization of subject matter is quickly supplanting text and lecture-based presentation in the classroom and in our day-to-day lives. However, vision is only one part of our sensory complement. Though comprising the “highest bandwidth” of information received, isolated visual learning risks suffering the inherent limitations inherent in any single-sensory modality. Our work at USC's Institute for Creative Technologies explores effective combinations of visual, auditory and other sensory functions to provide an integrated learning experience that exploits the natural connectivity of the brain. As each sense holds a proprietary “memory location” within the brain, the effective orchestration of multiple sensory inputs ensures a wider degree of neural stimulation.[8] This elicitation creates stronger memories by virtue of its collaborative effect, thereby enhancing the learning experience. The strength of these “experience memories” is also dependant in large part on their associated emotional attachment. As memories are stored in fragments across the cerebral cortex, they must be retrieved and reassembled. [1] The hippocampus is responsible for both storage and retrieval of these loosely linked memories and is a key component of the limbic system. The limbic system itself contributes emotional meaning to these fragmented elements. Also controlled by this part of the brain is motivation, and it is conjectured that these two aspects of our mental processes are closely related.[4] An increased emotional connection should have a positive effect on motivation and the retrieval of memories, and indeed, some research indicates that we tend to remember emotional salient events better than those perceived as emotionally neutral. [7] To capitalize on these relationships, we have taken an integrated sensory design approach towards creating training environments with an emphasis on the emotional connectivity the senses can elicit. While our current work focuses on critical thinking skills as applied to ethical and doctrinal command decisions within military situations, the research is extensible to all educational applications. Our prototype environment, “DarkCon” [3] employs emotional cues within the participant's experience to increase retention of specific elements in the training situation. By skillfully blending sights, sounds, and visceral frequencies we not only present the information to be learned, but create emotional catalysts as well. These persistent, timed, and triggered events are all designed to ensure longer-lasting memories of the experience. This multi-sensory design approach may prove to be instrumental in providing enriched learning experiences in science and engineering. In addition, the application of this approach may serve to attract students who might otherwise be dissuaded by the abstract nature of these fields. While this amalgam of ideas is as of yet untested, planned evaluation studies for Fall 2003 will provide insights to the validity of our hypothesis: that emotional connectivity and holistic involvement through multiple-sensory stimulation significantly enhances the learning experience. Recently, researchers in neurosciences and psychology have found that emotions are widely related to cognition, they exert influences in various behavioral and cognitive processes, such as attention, long-term memorizing, decision-making. Moreover, positive affects are fundamental in cognitive organization and thought processes; they also play an important role to improve creativity and flexibility in problem solving. Reciprocally, negative affects can block thought processes; people who are anxious have a reduced memory capacity and a deficit in inductive reasoning. E-learning environments require to consider all factors that can improve and facilitate learning
Higher Nervous Activity, Consciousness
*automatism in perception
Searchlight hypothesis - It is suggested that in the brain the internal attentional searchlight, proposed by Treisman and others, is controlled by the reticular complex of the thalamus (including the closely related perigeniculate nucleus) and that the expression of the searchlight is the production of rapid bursts of firing in a subset of thalamic neurons. It is also suggested that the conjunctions produced by the attentional searchlight are mediated by rapidly modifiable synapses--here called Malsburg synapses--and especially by rapid bursts acting on them. The activation of Malsburg synapses is envisaged as producing transient cell assemblies, including ``vertical'' ones that temporarily unite neurons at different levels in the neural hierarchy.
Synchronization of neuronal assembly (percept)- Synchronous network excitation is believed to play an outstanding role in neuronal information processing. Due to the stochastic nature of the contributing neurons, however, those synchronized states are difficult to detect in electrode recordings. We present a framework and a model for the identification of such network states and of their dynamics in a specific experimental situation. Our approach operationalizes the notion of neuronal groups forming assemblies via synchronization based on experimentally obtained spike trains. The dynamics of such groups is reflected in the sequence of synchronized states, which we describe as a renewal dynamics. We furthermore introduce a rate function which is dependent on the internal network phase that quantifies the activity of neurons contributing to the observed spike train. This constitutes a hidden state model which is formally equivalent to a hidden Markov model, and all its parameters can be accurately determined from the experimental time series using the Baum-Welch algorithm. We apply our method to recordings from the cat visual cortex which exhibit oscillations and synchronizations. The parameters obtained for the hidden state model uncover characteristic properties of the system including synchronization, oscillation, switching, background activity and correlations. In applications involving multielectrode recordings, the extracted models quantify the extent of assembly formation and can be used for a temporally precise localization of system states underlying a specific spike train. Over the past several decades, more than twenty different cortical areas have been recognized as processing loci of specific properties of visual scenes, such as forms, colours, motion, etc. At the same time, those investigations led to the conclusion that there are few, if any, places in the nervous system where the parcellated information of visual scenes is resynthesized to obtain a unified and coherent percept. Along this line, it has been suggested that synchronization of action potentials within a time-window of 1 to 5 ms between two or more neurons belonging to distant pools of cells may be an encoding process, which enables binding of various features of a single visual object. Binding, in general terms, is a process permitting the functional linkage of the distributed neuronal activity. Numerous previous studies have been carried out with multiunit recordings where several cells are recorded simultaneously. This method fails to disclose which cells participate in the synchronization process. Our investigations are aimed at further clarifying the processes leading to the formation of an encoding assembly by synchronization. Toward this end we investigated the modulation of synchronization between pairs of neurons sorted out from multiunit recordings. Our results show that synchronization may be observed in spontaneous activity. We also demonstrate that the amplitude of synchronization is modulated in relationship to the visual stimulus structure. Moreover, we found that, when considering one pair of cells, the spontaneous synchronization is high enough to mask stimulus-related-synchronization, whereas when other pairs of cells are added in the neuronal assembly, random synchronization is progressively outweighed, and the pattern of stimulus induced synchronization is progressively revealed
*layer V cells as cognitive output from cortical cells (Crick hypothesis)
*amplitude maps as a trace of meaning (Freeman hypothesis)
Readiness potential (Libet) - The readiness potential (BP) consists of movement-related cortical potentials (MRCPs) peaking in the motor potential (MP). Libet's readiness potential is recorded over what's called "supplementary motor cortex", an area of the brain associated with motor planning. Motor planning is generally studied only with respect to simple actions, like reaching your arm out to a target or (say) flicking your wrist. Complex activities, like picking up a gun and shooting somebody, get murky real fast, but presumably the supplementary motor area is still involved in planning the necessary movements. Higher-order planning - like "I'm gonna go kill that guy" - is typically associated with more frontal parts of the brain. You wouldn't expect to see the same sort of readiness potential until you actually started the sequence of motions needed to shoot him. In the situation you describe, where you spontaneously pick up a gun and shoot somebody (without, for the sake of argument, planning to kill him beforehand), I suspect that Libet would claim this was not a "conscious" action on your part. The readiness potential shows up, the nerve impulses necessary to start the action start on their way, and only then do you become conscious of your decision. But it's worth noting that most of the time, we don't just spontaneously pick up guns and shoot each other without on some level planning to do so beforehand. People who do are technically described as "bat-shit crazy", and you could make a pretty decent argument that you really were not quite in control of your mind at the time - whether you could convince a jury is another story. However, if you were already planning to off the guy, it seems to me that free will is still alive and well. The exact timing of your motions may not be under conscious control to the same extent that we'd like to think, but you are still capable, even according to Libet (as I understand) of deciding not to shoot at any time up until the nerve signal to pull the trigger starts heading for your finger. You could think of this abstract planning as "priming" motor planning, telling supplementary motor cortex that it's okay to chart out the movements to grab a gun and shoot. Once you give the motor area the all-clear, though, the exact timing and control of your actions is to a certain extent unconscious.
*sensory-motor laws as bases for consciousness
*examples of "external memory" (change blindness, dichotomic figures, illusion of seeing everything)
Wyszukiwarka
Podobne podstrony:
Basic English Key Words List 5
letters words find the right key
Prezentacja konsument ostateczna
WYKŁAD PL wersja ostateczna
05 DFC 4 1 Sequence and Interation of Key QMS Processes Rev 3 1 03
13 G06 H04 ostateczna wersjaid 14452 ppt
Prezentacja harcerstwo ostateczne 3
rodowiskowe uwarunkwania zdrowia i choroby ostateczny 1
rehabilitacja chorego nieprzytomnego ostateczna wersja[1]
high key
OSTATECZNA Łukowska, M (2011) Mechanizmy kontroli głębokości zanurzenia przestrzennego w środowisko
Ostateczny porządek Świata wg żydów
first certificate practice tests and key 2
CARPROG BMW Key programmer manual
plan wypowiedzi ostateczny, szkoła
CD-KEY The Godfather (PC GAME) All, CD KEY'E
więcej podobnych podstron