Spis treści
--== JĄDRO KOMÓRKOWE ==--
1. Komórka
Bakterie
- translacja rozpoczyna się na końcu nici mRNA jeszcze podczas jej transkrypcji
- sekwencja sygnałowa (~20 aminokw.) zapewnia dostarczenie kompleksu translacyjnego (mRNA, rybosomy + sekwencja sygnałowa) do błony kom. w czasie krótszym niż 5s. by polipeptyd nie zdążył ulec zmianom konformacyjnym uniemożliwiającym mu opuszczenie komórki
Strategie utrzymywania równowagi w kom. eukariotycznej i bakterii:
- kompleksy enzymatyczne specyficzne tylko dla danych reakcji
- błony wew. kom. tworzą kompartymenty w kom. dzięki którym mogą zachodzić różne, przeciwstawne procesy katalizowane przez różne enzymy
Uwaga! W przypadku zniszczenia przedziałów kom. ulega samostrawieniu przez przeważające procesy degradacyjne
7 głównych przedziałów i reakcje za które są odpowiedzialne:
- jądro (synteza DNA i RNA)
- cytozol (metabolizm pośredni)
- siateczka śródplazmatyczna (synteza białek, lipidów i błon)
- aparat Golgiego (kontrola glikozylacji, obróbka białek na eksport)
- mitochondria + plastydy (synteza ATP)
- lizosomy (trawienie)
- peroksysomy (utlenianie, rozkład H2O2, detoksykacja)
Jądro
- kształt zbliżony do obrysu komórki
- średnica od kilku do kilkunastu mikrometrów
- trzy przedziały: macierz jądrowa (nukleoplazma), chromatyny jądrowej, jąderko
- otoczone w osłonką jądrową
Struktura chromatyny:
Euchromatyna (luźna)
- czynna transkrypcyjnie
- 7 -10% całej chromatyny jądrowej
- w komórkach w interfazie budują ją całkowicie rozwinięte chromosomy
- w jądrach plemników jest skondensowana ! ( nie mylić z heterochromatyną)
- może występować przejściowo w stanie skondensowany w ciałkach Barra u kobiet (w leukocytach jako pałeczka dobosza)
Zawiera:
- wysoce powtarzalne sekwencje niekodujące
- tandemowe
- rozproszone
- sekwencje umiarkowanie powtarzalne
- sekwencje unikatowe kodujące geny strukturalne
Heterochromatyna (skondensowana)
- nie posiada odcinków kodujących (odcinki chromosomów nierozwinięte w interfazie)
- zbudowana z krótkich sekwencji nukleotydów ułożonych tandemowo i powtórzonych milion razy w genomie
- czasami występują pomiędzy ruchome elementy kodujące -tzw. transpozony
- zawiera sekwencje satelitarne
- znajduje się przy centomerach, telomerach i w obszarach interstycjalnych
- można ją uwidocznić najłatwiej w skondensowanych chromosomach metafazowych.
Nukleosomowa budowa chromatyny
- chromatyna to kompleks niehistonowych, histonowych białek i kwasów dezoksyrybonukleinowych
- DNA jest owinięte na histonowym oktamerze
- nukleonom to podstawowa jednostka strukturalna chromatyny
- rdzeń histonowy tworzą: 2x (H2A,H2B,H3,H4) wokół nich nawinięte jest DNA, a w rejonie łącznikowym jest H1 lub H5 (razem jest to tzw. nić nukleosomowa)
Sposób upakowania chromatyny w jądrze
Dwuniciowa cząsteczka DNA → nić nukleosomowa → solenoid (właściwe włókno chromatynowe) → pętle domenowe → grona 1 rzędu i 2 rzędu → chromosom
Jąderko
- nie ma go w plemnikach i w erytrocytach ptaków
- wielkość zależna od syntezy białek
- jest integralną częścią chromosomów jąderkowych
- jego zadaniem jest synteza pre rRNA i formowanie jednostek rybosomów
- składa się z 3 rejonów: włóknistego, gęstego składnika włóknistego, ziarnistego zawieszonych w macierzy jąderkowej
- najważniejsze białka jąderkowe: polimeraza RNA 1, nukleolina, fibrylaryna, rybocharyna
Otoczka jądrowa
wyściółka podosłonkowa + 2 błony:
- zewnętrzna - podobna do siateczki śródplazmatycznej (jest z nią połączona, ma rybosomy)
- wewnętrzna - posiada białka receptorowe dla białek związanych z laminą B np. emeryna, murima i LAP1, 2, 2b (wiąże błonę osłonki z filamentami lamin) + wyściółka podosłonkowa
- pomiędzy błonami znajduje się przestrzeń wokółjądrowa (cysterna perynuklearna)
- wewnętrzna wyściółka białkowa osłonki jądrowej (z 3 typów białek: laminy A, B1 i B2) bierze udział w
utrzymaniu integralności jądra
Pory jądrowe
- są związane z kompleksem białkowym poru (zbudowanym z około 100 rodzajów białek tzw. nukleoporyn)
- znajdują się w miejscu połączenia błony zew. i wew.
- zapewniają wybiórczy transport
- im aktywniejsza metabolicznie komórka tym więcej porów
- kompleks porowy jest promieniście symetryczny (3 pierścienie białkowe połączone 8 włóknami pionowo i poziomo, na środku obu pierścieni występuje kompleks transportujący, a w nim kanał transportujący z bezpostaciową substancją)
- białka łączące por z osłonką jądrową to gp62, gp190 i gp210
- w czasie mitozy kompleksy porowe ulegają rozpadowi i zostają odtworzone w telofazie
Rodzaje transportu z i do jądra komórkowego
- do jądra przedostają się makrocząsteczki, które wchodzą w interakcje z białkami poru bezpośrednio lub za pomocą innych białek
- cząsteczki <50kDa są importowane i eksportowane na zasadzie swobodnej dyfuzji
- cząsteczki >50kDa są transportowane za pomocą specyficznych białek i przy użyciu energii
- transport białek jądrowych z cytoplazmy do jądra odbywa się z udziałem sekwencji lokalizującej jądro
- transport cząsteczek z jądra (np. RNA) odbywa się z udziałem sekwencji eksportującej NES
(sekwencje te rozpoznawane są przez importyny β)
Etapy transportu
- importyny α i β rozpoznają sekwencje lokalizującą jądro NSL znajdującą się na białku importowane i tworzą razem kompleks importowy
- importyna β od strony cytoplazmy dokuje powstały kompleks
- kompleks Ran-GTP transportuje cząsteczkę przez kanał porowy
- po dostaniu się do nukleoplazmy jądra importyna β, a następnie α odłączają się i wracają do cytoplazmy
Skład macierzy jądrowej
- roztwór białek, cukrów, jonów, nukleotydów
Białkowy skład:
- Laminy
- laminy 60-70kDa
- białka nielaminowe w macierzy wewnętrznej jądra (wykryte w chromatynie interfazowej i chromosomach skondensowanych) = włóknisto - ziarniste matryny np. F, G, H i 3,4,15
- matryna F/G z DNA się wiąże za pomocą palców cynkowych odpowiedzialnych za kontrolę ekspresji genów (3 alfa - helisy połączone atomem cynku)
- Białka tworzące kompleksy z RNA
- Komórkowo specyficzne białka macierzy
- w konkretnych rodzajach komórek np. glejak ma polipeptydy specyficzne dla komórek nowotworowych
DNA macierzy jądrowej
- klasa główna 75%
- klasa stowarzyszona z macierzą białkową 25%
-DNA rozpuszczalny
- DNA resztkowy 1% (ze strukturami białkowymi tworzy sieć włókien)
RNA macierzy jądrowej
- podstawowa struktura macierzy jądrowej to rybonukleinowe filamenty rdzeniowe
- 65% całego RNA jądrowego
- 60-80% hnRNA
- 74% snRNA
- śladowe ilości rRNA
- większość prekursorowego mRNA rRNA
Uwaga! W oocytach i blastomerach ssaków mogą występować jądra dodatkowe z pseudojąderkami, które powstają przez odpączkowanie ja jadra komórkowego.
2. Cykl życiowy komórki
Interfaza:
G0
- stan spoczynkowy kom.
- kom. tracą zdolność do podziałów
- kom. ma bardziej skondensowaną chromatynę, inne rodzaje mRNA i swoiste białka
- kom. wchodzą w fazę G0 z G1
- czynniki patologiczne mogą spowodować przejście z tej fazy do G1 np. limfocyty B i T pod wpływem mutagenów
G1
-faza wzrostu
- dla komórek rzadko dzielących się jest to najdłuższa część cyklu
- wchodzi w nią komórka dwukrotnie mniejsza od macierzystej
- zachodzą intensywne procesy anaboliczne, wymiana chemiczna z otoczeniem i wzrasta ruchliwość i wrażliwość na bodźce zew.
- następuje zwiększenie masy i objętości kom.
- zachodzi synteza cykliny A, C, D, E
- pod koniec znajduje się pierwszy punkt restrykcyjny (kontrola integralności mat. gen.) po którym komórka bez względu na warunki jest zmuszona do realizowania kolejnych etapów cyklu
S
- faza syntezy DNA
- ma względnie stałą długość ze względu na fakt, że replikacja DNA przebiega z jednakową prędkością w jednakowych warunkach
- ilość DNA podwaja się
- komórka zwiększa masę i objętość
- syntetyzowane są histony i rozpoczyna się produkcja cykliny B
G2
- bezpośrednie przygotowanie do podziału
- synteza białek wrzeciona podziałowego (tubulina) i cykliny B (składnik kinazy fazy M)
- produkcja składników koniecznych do odtworzenia błony w telofazie
- aktywacja kinazy białkowej CDK potrzebna do zaniku błony i kondensacji chromosomów
- drugi punkt restrykcyjny (kontrola prawidłowości kondensacji i segregacji mat. gen. Oraz tworzenie wrzeciona podziałowego)
Mitoza
- profaza
-chromosomy ulegają kondensacji
- uwidocznia się ich podział podłużny na dwie chromatydy siostrzane
- tworzą się kinetochory
- zanika błona jądrowa i jąderko
- metafaza
- silnie skręcone chromosomy grupują się w środku komórki tworząc tzw. płytkę metafazową
- anafaza
- chromatydy ulegają całkowitemu rozdzieleniu i są przemieszczane do przeciwległych biegunów przez nici wrzeciona podziałowego
- telofaza
chromosomy ulegają de kondensacji
- następuje defosforylacja białka blaszki jądrowej-laminy (wokół jąder potomnych powstaje błona jądrowa i odtwarzają się jąderka)
- cytokineza
- pierścień z włókien aktyno miozynowych w prostopadłej płaszczyźnie do długiej osi wrzeciona podziałowego zaciska się tworząc ciałko środkowe
- u roślin z soczewkowatą płytką podziałową łączą się pęcherzyki z prekursorami ściany komórkowej, która rośnie w kierunku przeciwległych ścian dzieląc cytoplazmę
Mejoza I (podział komórek rozrodczych)
- profaza I
• leptoten -chromosomy przybierają postać długich, cienkich nitek
• zygoten - chromosomy homologiczne układają się w pary (koniugują ze sobą) tworząc biwalenty, (tetrady)
• pachyten - zachodzi crossing-over, czyli wymiana odcinków chromatyd niesiostrzanych chromosomów homologicznych
• diploten - chromosomy homologiczne rozchodzą się, ale pozostają złączone w punktach zwanych chiazmami. Każdy z biwalentów połączony jest poprzez chiazmę. Następuje intensywna synteza RNA i dekondensacja chromosomów.
• diakineza - chromosomy ulegają kondensacji, dochodzi do spadku syntezy RNA, chromosomy oddzielają się od otoczki jądrowej, kinetochory tworzące biwalent zlewają się ze sobą.
- metafaza I
- chromosomy biwalentne układają się w płaszczyźnie równikowej komórki
- kończy się tworzenie wrzeciona podziałowego
- kinetochory są połączone z mikrotubulami wrzeciona
- anafaza I
- rozejście się biwalentow na 2 pary chromatyd do przeciwległych biegunów wrzeciona podziałowego
- telofaza I
- powstanie 2 komórek o liczbie chromosomów zredukowanej do połowy (haploidalna liczba chromosomów z diploidalną liczbą DNA) w stosunku do komórki macierzystej
Mejoza II
- przypomina podział mitotyczny
- ma kolejno miejsce druga profaza, metafaza, anafaza i telofaza
- w metafazie chromosomy układają się w płaszczyźnie równikowej
- w anafazie połączone dotąd centromerami chromatydy poszczególnych chromosomów rozdzielają się i wędrują ku biegunom komórki
- w rezultacie powstają 2 komórki o 23 chromosomach każda
Czynnik dojrzewania MPF
- składa się z białka p34 (produkt genu cdc2) mającego zdolność do przenoszenia grup fosforanowych z ATP na inne białka
- powoduje, że kom. jajowe zatrzymane w profazie I mejozy, wchodzą w mejozę II i dojrzewają do zapłodnienia
Nondysjunkcja
- nierozdzielenie się chromosomów podczas podziału
- przyczyną
3. Występowanie chromatyny płciowej X i Y.
Ciałko X = Ciałko Barra
- ciemniejsza grudka zasadochłonnej chromatyny dyskowato przylegająca do błony jądrowej i wybarwiająca się w jądrach interfazowych komórek osobników żeńskich
- w granulocytach obojętnochłonnych-w postaci pałeczek dobosza (chromatyna wyrzucona poza obręb jądra)
- obecna w komórkach niemal wszystkich tkanek
- u człowieka najczęściej bada się w rozmazach nabłonka jamy ustnej i w komórkach płynu owodniowego (20-60% i to uznano za normę, próg jest 15%)
- w kom. nerwowych [nie dzielą się] - obecność wykazuje 70-90% komórek
- niewielki odsetek w nie dzielących się komórkach
- występowanie chromatyny X nie jest związane z płcią lecz liczbą akt. chromosomów X w kariotypie (n=X-1)
Inaktywacji na ch. X nie ulegają:
- geny na końcu ramion krótkich
- geny w regionie koniugacyjnym (PAR) z Y
- część proksymalna ramienia długiego i krótkiego
- jeśli 1 X jest błędny, to on ulega inaktywacji
Uwaga! Niski odsetek ciałek Barra w mat. z biopsji z tk. nowotworowych świadczy o złośliwości guza
Ciałko Y
- silnie fluoryzująca dystalna heterochromatynowa cz. długiego ramienia (q12)
- uwidacznia się w jądrze komórkowym komórek interfazowych męskich (30-50%), przy delecji q12 lub małym Y może być brak ciałka Y i można się pomylić
- może uwidocznić się w postaci pałeczki Dobosza, która u mężczyzn barwi się techniką fluorescencyjną, co odróżnia ją od pałeczki żeńskiej
- ich liczba odpowiada liczbie chromosomów Y w kariotypie (n=Y)
4. Hipoteza powstawania chromatyny X (teoria Lyon).
Założenia Teorii Lyon:
- Występowanie chromatyny płciowej nie jest bezwzględnie powiązane z płcią
- Obecność ciałka Barra jest związane z liczbą chromosomów X w kariotypie
- Chromatyna płciowa nie powstaje w kariotypie zawierającym jeden chromosom X
- Liczba chromosomów Y nie wpływa w żadnym stopniu na liczbę ciałek Barra
- Jeżeli liczba chromosomów X jest większą niż jeden, wtedy jeden chromosom zawiera czynny materiał genetyczny, a pozostałe ulegają unieczynnieniu- lionizacji (kondensacji)
Lionizacja (heterochromatyzacja, kondensacja)- jest to losowa inaktywacja jednego z heterochromosomów (chromosomów X), będąca mechanizmem mającym na celu wyrównanie ilości informacji genetycznej, która jest zawarta w na ogół dwóch chromosomach X u kobiety w stosunku do jednego chromosomu X u mężczyzny
- we wczesnym okresie rozwoju embrionalnego, przed gastrulacją (16 dzień rozwoju zarodka człowieka)
- inaktywacja losowa chromosomów - tworzy się mozaika (organizm ma komórki Xpat/Xmat)
- obejmuje większość genów, poza mniej lub bardziej homologicznymi do tych w chromosomie Y (tzw. geny pseudoautosomalne)
- inaktywacja przez metylację (nieaktywny chromosom X jest w znacznym stopniu zmetylowany, czego skutkiem jest zahamowanie transkrypcji)
- proces inaktywacji jest inicjowany w centrum inaktywacji XIC (X Inactivation Center - q13), przez gen XIST
- XIST ulega transkrypcji w nieaktywnym chromosomie X, w chromosomie aktywnym nie podlega temu procesowi; nie koduje białka, ale RNA, które rozprzestrzeniając się powoduje inaktywację
- utrata genu XIST w wyniku np. delecji uniemożliwia inaktywację tego chromosomu
- nieaktywny chromosom X jest bardzo odporny na większość mutagenów, ale niektóre związki mutagenne mogą wywołać derepresję genów np. 5- azacytydyna
- nieczynne chromosomy X charakteryzują się opóźnioną replikacją DNA w późnej fazie S i dlatego różnią się wzorem prążków
Mechanizmy inaktywacji chromosomu X:
Przypadkowa inaktywacja prawidłowa → mozaicyzm tkankowy
Inaktywacja prawidłowego chromosomu → drugi chromosom z mutacją wpływa na przeżywalność kom.
Mutacja genu XIST → nieprzypadkowa inaktywacja
5. Hodowla limfocytów krwi obwodowej - uzyskiwanie chromosomów metafazowych.
Fazeolina = Fitohemaglutynina = wyciąg z nasion fasoli
- posiada właściwości mitogenne
- stosowana do sedymentacji krwinek czerwonych
- pod jej wpływem krwinki białe ulegają odróżnicowaniu (transformacji blastycznej) odzyskując zdolność podziału
Etapy procesu:
- Umieszczenie niewielkiej ilości jałowej, heparynizowanej krwi pełnej lub osocza w naczyniach zawierających odpowiednie podłoże: płyn hodowlany (aminokwasy, cukry, witaminy, sole mineralne), surowica ludzka lub cielęca + fazeolina.
- 2-3-dniowa inkubacja w temperaturze 37 stopni C
- Dodanie kolchicyny lub jej pochodnych na 2-3 godziny
- Wirowanie- oddzielenie komórek od podłoża hodowlanego
- Inkubacja w płynie hipotonicznym- rozluźnienie i rozproszenie garnituru chromosomowego
- Utrwalenie w płynie Carnoy
- Końcowe odwirowanie
- Naniesienie kropli zawiesiny na szkiełko podstawowe. W zawiesinie znajdują się limfocyty nieodróżnicowane, limfocyty w rożnych stadiach oraz limfocyty ulegające podziałom i hamowane przez kolchicynę w stadium metafazy
- Barwienie np. czynnikiem Giemsy, orceiną
- Obserwacja kariotypu w największych powiększeniach
- Mikrofotografia odpowiednich metafaz
- Sporządzanie kariogramu, wycinanie chromosomów z odbitek fotograficznych
6. Budowa chromosomy metafazowego.
Chromosom metafazowy to najbardziej skondensowana forma chromatyny. Każdy chromosom zawiera jedną cząsteczkę DNA upakowaną w kilku poziomach uorganizowania. Jest to niezbędne, aby podczas podziału komórkowego materiał genetyczny został idealnie i równo podzielony. Chromosom składa się z dwóch, równolegle ułożonych chromatyd połączonych w obszarze przewężenia pierwotnego tj. centromeru. Jest on miejscem przyczepu włókien wrzeciona kariokinetycznego. Ze względu na jego umiejscowienie wyróżniamy cztery podstawowe rodzaje chromosomów metafazowych: submetacentryczny, metacentryczny, teocentryczny i akrocentryczny.
Chromatyna posiada kilka stopni upakowania. Podwójna helisa DNA wraz z białkami tworzy nukleosomy. Nukleosom obejmuje łańcuch DNA nawinięty na rdzeń zbudowany z 4 rodzajów białek histonowych (H2A, H2B, H3, H4), nazywamy go oktamerem histonowym. Pomiędzy nukleosomami znajduje się DNA łącznikowy.Nukleosomy i DNA łącznikowe układają się w specyficzny, zygzakowaty sposób, tworząc solenoid. Ten układa się w pętle, z których składają się chromatydy.
7. Metody uzyskiwania prążków G, Q, C, R.
Chromosomy badamy w celu określenia ich liczby i struktury. Pobiera się wtedy limfocyty krwi obwodowej lub komórki szpiku kostnego. W szczególnych wypadkach chromosomy otrzymujemy dzięki hodowli fibroblastów skóry lub tkanek pochodzących z guzów nowotworowych. Kariotyp płodu oceniamy na podstawie badania komórek pobranych z trofoblastu lub płynu owodniowego.
Najczęściej hoduje się limfocyty krwi obwodowej. Pobrane znajduja się w fazie G1 lub G0. Muszą zatem zostać pobudzone do podziałów. Do tego celu stosuje się mitogenny wyciąg z nasion fasoli fitohemaglutyninę (PHA).
Charakterystyczny dla każdego z chromosomów układ prążków ujawniony podczas barwienia umożliwia identyfikację poszczególnych par homologicznych oraz analizę ich nieprawidłowości (aberracje strukturalne).
Wzór prążkowy - liczba, wymiary i umiejscowienie prążków charakterystyczne dla każdego gatunku.
Metody
G - preparat wybarwiamy enzymami proteolitycznymi, trypsyną i barwnikiem Giemsy, otrzymujemy prążki ciemne - prążki G o dużej zawartości par zasad A-T z późno replikującym DNA, poprzedzielane międzyprążkami (jasne) dużo par G -C - jest to wcześnie replikująca chromatyna aktywna transkrypcyjnie
Q - do wybarwiania używmy dwóch rodzajów fluorochromów
- oranż akrydynowy (łączy się jonowo z DNA lub interkaluje tj. wsuwa się pomiędzy płaszczyznę jego zasad)
-atebryna (Q) oraz iperyt akrychinowy (QM) (wykazują zdolność wsuwania się pomiędzy płaszczyzny DNA oraz powinowactwo do pewnych par zasad)
Silnie fluoryzujące prążki regiony DNA bogate w pary A-T
Wygaszanie fluorescencji regiony bogate w pary G-C
C - po uprzedniej inkubacji w Ba(OH)2 do wybarwiania używamy odczynnika Giemsy, wykrywamy w ten sposób chromatynę konstytutywną- regiony centromerowe i przycentromerowe
R - jest to odwrotność prążków G; prązki ciemne - pary G-C, prążki jasne - pary A-T. Metoda ta jest potrzebna przy wykrywaniu drobnych aberracji nieujawnionych przy barwieniu prążków G lub Q.
8. Kryteria klasyfikacji chromosomów człowieka.
Prawidłowy kariotyp człowieka zawiera 46 chromosomów: 44 autosomy i 2 chromosomy płciowe.
- Autosomy (chromosomy somatyczne)- wszystkie chromosomy znajdujące się w komórce poza chromosomami płciowymi
- Chromosomy płci (heterochromosomy, allosomy)- chromosomy determinujące płeć
Kryteria klasyfikacji:
- Wielkość
- Położenie centromeru
- Rozmieszczenie prążków w chromosomach
Ze względu na położenie centromeru chromosomy człowieka dzielimy na 4 grupy:
- Chromosomy metacentryczne: centromer znajduje się w środku chromosomu, ramiona górne (p), są równe dolnym (q)
- Chromosomy submetacentryczne: centromer jest przesunięty bliżej jednego końca, ramiona górne są krótsze od dolnych
- Chromosomy akrocentryczne: centromer znajduje się blisko jednego końca, ramiona górne są bardzo krótkie, a dolne bardzo długie
- Chromosomy telocentryczne: centromer znajduje się na samym końcu chromosomu, ramiona górne nie występują, obecne są tylko długie (u człowieka chromosomy telocentryczne nie występują)
Ze względu na wielkość i położenie centromeru, chromosomy człowieka zostały podzielone na 7 grup:
A- Chromosomy 1-3, duże chromosomy metacentryczne
B- Chromosomy 4-5, duże chromosomy submetacentryczne
C- Chromosomy 6-12 i chromosom X, średnie chromosomy submetacentryczne
D- Chromosomy 13-15, duże chromosomy akrocentryczne, mogące zawierać nitki satelitonośne i satelity
E- Chromosomy 16-18, 16- mały chromosom prawie metacentryczny, 17 i 18- małe chromosomy submetacentryczne
F- Chromosomy 19-20, najmniejsze chromosomy metacentryczne
G- Chromosomy 21-22 i chromosom Y, chromosomy 21 i 22 to małe chromosomy akrocentryczne, mogące posiadać nitki satelitonośne i satelity, chromosomy 21 pary są mniejsze niż pary 22, chromosom Y nie posiada satelit
9. Zasady układania prawidłowego kariogramu człowieka.
W celu ułożenia prawidłowego kariogramu człowieka, niezbędne było uzyskanie z badanych komórek(najczęściej są to białe krwinki)chromosomów w stanie podziałów metafazowych. Do tego celu używa się fazeolinę(inaczej fitohemaglutynina PHA). Pod jej wpływem dojrzałe komórki(limfocyty) ulegają transformacji blastycznej, tzn. że z formy dojrzałej powracają do postaci młodocianych, dzięki czemu mogą się dzielić. Metoda ta jest o tyle ważna, ze umożliwiła uzyskanie z nawet niewielkiej liczby limfocytów dużej grupy dzielących się komórek. Limfocyty trzeba umieścić na podłożu złożonym z cukrów, witamin, soli mineralnych, surowicy ludzkiej oraz fazeoliny. Po 2,3-dniowej inkubacji w temperaturze ok37˚C dodaje się kolchicynę, która hamuje mitozę w stadium metafazy oddziaływując na wrzeciono kinetyczne. Następnie wiruję się hodowlę w celu rozluźnienia garnituru chromosomowego, po czym utrwala się ją w płynie Carnoy. W celu ułatwienia rozróżnienia chromosomów wybarwia się je barwnikiem Giemsy. W tym stanie zawiesinę nanosi się na szkiełka podstawowe i dokonuje analizy pod mikroskopem. Pod największym powiększeniem wykonuje się zdjęcia metafaz,i z wywołanych odbitek wycina się chromosomy, dopasowuje się je w odpowiednie pary. W celu ułatwienia(chromosomy często trudno dobrać w pary)oraz by uwidocznić abberacje chromosomowe korzysta się z metody autoradiografii. Polega ona na wbudowywaniu w materiał genetyczny nukleotydów, które posiadają radioaktywne izotopy wodoru i węgla.
--== GENETYKA ==--
10. Podstawowe pojęcia genetyczne.
Gen- podstawowa jednostka dziedziczności, to fragment DNA odpowiedzialny za wykształcenie określonej cechy. Koduje jeden łańcuch polipeptydowy lub jeden rodzaj RNA.
Allel- jedna z form występowania genów warunkującego dana cechę. Allele to geny położone w identycznych miejscach w homologicznych chromosomach.
Gen letalny- gen, który powoduje, że organizm jest niezdolny do życia. Geny letalne są zazwyczaj genami recesywnymi, ponieważ gdyby były dominujące to nie mogłyby być przekazywane dalej(bo wszystkie homozygoty dominujące i heterozygoty by umarły).
Fenotyp- wygląd organizmu, jest związany z współdziałaniem genotypu z środowiskiem życia organizmu.
Genotyp- to zespół wszystkich genów danego organizmu(zawarta w każdej komórce).
Homozygota- organizm powstały z połączenia gamet o jednakowym genotypie.Termin odnosi się najczęściej do jednej pary alleli np. AA lub aa. Możemy wyróżnić homozygoty recesywne(które mają oba allele danego genu recesywne: aa)oraz homozygoty dominujące, które posiadają oba allele danego genu dominujące: AA).
Heterozygota- organizm powstały z połączenia gamet o różnym składzie genetycznym, heterozygota posiada w allelu 2 różne geny(gen dominujący i recesywny): Aa
Cecha dominująca- jest to cecha, która ujawnia się zarówno u homozygot dominujących, jak i u heterozygot.
Cecha recesywna- to cecha, która ujawnia się tylko u homozygot recesywnych.
Dominacja zupełna- występuje wtedy, gdy fenotyp heterozygoty nie różni się od fenotypu homozygoty dominującej.
Dominacja niezupełna- występuje wówczas gdy fenotyp heterozygotyczny różni się od fenotypu homozygoty dominującej i recesywnej.
Krzyżówka testowa- krzyżówka pomiędzy nieznanym genotypem o fenotypie dominującym, z homozygotą recesywną.
Krzyżówka wsteczna- polega na krzyżowaniu heterozygotycznego potomka z jedną z homozygotycznych form rodzicielskich (P). Efektem krzyżowania wstecznego jest krzyżówka genetyczna zwana pokoleniem wstecznym.
11. I i II prawo Mendla, allele wielokrotne, geny kumulatywne i dziedziczenie cech ilościowych, współdziałanie genów w wykształceniu cech(epistaza, penetracja, ekspresja, poligeny, plejotropia, komplementacja), chromosomowa teoria dziedziczenia.
Pierwsze Prawo Mendla- prawo czystości gamet- każda gameta wytwarzana przez organizm posiada tylko jeden allel z danej pary alleli genu. Podczas mejotycznego podziału kom. następuje rozdział odpowiadającej sobie pary genów (alleli).
Drugie Prawo Mendla- cecha uwarunkowana jedną parą genów (alleli) dziedziczy się niezależnie od cechy uwarunkowanej drugą parą genów (alleli) w związku z czym w pokoleniu F2 obserwuje się rozszczepienie fenotypów w stosunku 9:3:3:1.
Allele wielokrotne- allele występujące w więcej niż dwóch postaciach; w danym organizmie mogą występować tylko dwa allele, natomiast w puli genowej populacji może być ich wiele.
Chromosomowa teoria dziedziczenia (Thomas Morgan) Jest ona b. dobra do badań genetycznych ponieważ:
- Wykazuje dużą zmienność genetyczną
- Posiada małą liczbę chromosomów 2n=8
- Wykazuje dużą płodność
- Krótki cykl życiowy - rozwój następnego pokolenia od 7 do 14 dni
Założenia chromosomowej teorii dziedziczenia
- Geny zlokalizowane są liniowo, każdy gen zajmuje ściśle określone miejsce - locus
- Allele zajmują to samo miejsce w chromosomach homologicznych
- Geny zlokalizowane w tych samych chromosomach tworzą tzw. grupę sprzężenia tzn. że dziedziczą się razem, czyli niezgodnie z II prawem Mendla.
- Rozprzęganie genów zachodzi w procesie crossing- over
- Crossing- over - zachodzi między dwoma genami tym częściej im dalej są one od siebie położone w chromosomie.
- Częstość procesów crossing over może być więc względna miarą długości genów w chromosomie. Jednostką odległości między genami jest 1 centimorgan 1 j.m.
- Rekombinanty powstają z gamet w których doszło do crossing- over miedzy genami. Ilość tych rekombinantów jest więc miarą częstości crossing- over.
- Znając odległość między genami w chromosomie, można ustalić kolejność genów na chromosomie.
Geny kumulatywne (poligeny)- geny te dziedziczą się zgodnie z prawami Mendla, warunkują powstawanie cech ilościowych, są to geny, które należą do różnych par alleli, których działanie sumuje się kształtując w ten sposób fenotyp.
Dziedziczenie cech ilościowych- są to cechy zależne od większej liczby par genów nieallelicznych. Im większa liczba par genów uczestniczy w dziedziczeniu cechy tym rozkład fenotypów u potomstwa jest bardziej zbliżony do normalnego tzn. zgodny z przebiegiem krzywej populacyjnej. Krzywa populacyjna zwana krzywą Gausa przedstawia zależność liczby osobników o określonym natężeniu badanej cechy. Pokazuje ona, że w populacji zawsze jest najwięcej osobników o średnim natężeniu badanej cechy. Przykładem cech ilościowych jest wzrost, waga, inteligencja i wynikające z niej szczególne zdolności np. matematyczne, muzyczne, barwa skóry.
Epistaza- dotyczy dwóch lub więcej par alleli. Zach. gdy ekspresja genu dominującego jest tłumiona działaniem produktu innego genu znajdującego się w odmiennym locus lub w innym chromosomie. Gen hamujący nazywamy epistatycznym, a hamowany hipostatycznym. W zależności od wł. genu tłumiącego mówimy o epistazie dominującej (barwa owoców dyni) lub recesywnej (fenotyp bombajski).
Penetracja- częstość ekspresji określonego genu wyrażona w procentach lub wartościach liczbowych.
Ekspresja- ujawnienie się produktu danego genu, odpowiedzialnego za dana cechę.
Plejotropia- polega na warunkowaniu przez jeden zmutowany gen kilku pozornie niezwiązanych z sobą cech genotypowych (zespól Marfana).
Komplementacja- polega na dopełniającym działaniu produktów różnych genów. Geny komplementarne działają wspólnie. Produkt jednego genu na wpływ na produkt drugiego.
12. Dziedziczenie u człowieka - układ grupowy ABO, układ grupowy Rh, dziedziczenie sprzężone z płcią.
Występowanie grup krwi związane jest z obecnością na powierzchni komórek (prócz układu nerwowego) antygenów, czyli substancji wywołujących odpowiedz immunologiczna. Antygeny występują w postaci oligosacharydów, glikolipidów i glikoprotein. Ich specyficzność jest wynikiem obecności reszt cukrowych zajmujących najbardziej powierzchniowe części makrocząsteczek. Wyróżniamy 4 grupy krwi: A, B, AB i 0. Allele warunkujące występowanie grup znajdują sie w tym samym loci (allelizm wielokrotny) ramienia długiego chromosomu 9. Allele A i B są kodominujace względem siebie i dominujące w stosunku do allelu 0. Zapis wykorzystywany w krzyżówkach genetycznych wygląda następująco: IA, IB, i0. Trzeba pamiętać, ze istnieje kilka wariantów antygenu A i B.
Grupę A1 posiadają osoby o genotypie: A10, A1A1, A1A2
Grupę A2 posiadają osoby o genotypie: A2A2 lub A20
Grupę B posiadają osoby o genotypie: BB lub B0
Grupę A1B posiadają osoby o genotypie: A1B
Grupę A2B posiadają osoby o genotypie: A2B
Grupę 0 posiadają osoby o genotypie: 00
Poza antygenami każda grupa krwi charakteryzuje się odpowiednim zestawem naturalnych przeciwciał należących do klasy IgM (występują również w płynach ustrojowych i wydzielinach):
Grupa A1: anty-B
Grupa A2: anty-B i niekiedy anty-A1
Grupa B: anty-A
Grupa 0: anty-A i anty-B
Grupa A1B: brak naturalnych przeciwciał
Grupa A2B: mogą wystąpić anty-A1
Występowanie danej grupy krwi zależy od genu, którego enzym odpowiada za dołączanie odpowiednich reszt cukrowych. Jednak bardziej podstawowy jest gen H (chromosom 19), który odpowiada za syntezę enzymu dołączającego substancje podstawowa (antygen) H (cukier).
H - kodujący enzym
h - allel niemy (brak produktu białkowego)
Osoby z genotypem hh pomimo posiadania genów kodujących grupę krwi A lub B nie posiadają tych antygenów na erytrocytach, choć w ich surowicy występują odpowiednie naturalne przeciwciała dla danej grupy AB0. Jest to tzw. grupa krwi "Bombay". Fenotypy: 0hA,0hB,0hAB. Nie wykształcana jest substancja podstawowa i mimo obecności reszty odpowiadającej danej grupie brak jej "uwidocznienia".
Układ Rh
Układ Rh dziedziczy sie niezależnie od układu AB0. Na krótkim ramieniu chromosomu 1 znajdują sie 3 par genów sprzężonych ze sobą: Cc, Dd i Ee. Antygen D ma największą moc do pobudzania reakcji immunologicznej (wytwarzania przeciwciał) i on właśnie determinuje fenotyp Rh+ lub Rh-. Przeciwciała tego układu powstają w ustroju na skutek przetaczania krwi bądź wystąpienia konfliktu serologicznego. Oznacza to, ze antyRh nie występują naturalnie w osoczu. Przeciwciała te należą do immunoglobulin IgG i maja zdolność przechodzenia przez łożysko. W konflikcie serologicznym matka zawsze jest Rh-, a dziecko Rh+ (ojciec Rh+). Krwinki płodu przedostają sie do organizmu matki i aktywują wytwarzanie antyRh. Kiedy antyRh przedostają sie do ustroju dziecka powodują chorobę hemolityczna. 90% kobiet nie wytwarza tych przeciwciał w czasie pierwszej ciąży. Ryzyko wystąpienia konfliktu wzrasta wraz z kolejnymi ciążami. Zapobiegawczo podaje sie kobiecie gammaglobuline antyRh, która niszczy erytrocyty dziecka. Organizm matki nie produkuje wtedy własnych antyRh.
Układ grupowy MNSs
Występują w erytrocytach dodatkowe antygeny M i N. Locus L genu M i N- chromosom 4. Miedzy antygenami występuje kodominacja - grupa M, N
i MN. Obecny jest również antygen S.
Sposób przekazywania genów związany z ich obecnością w chromosomach płci. Należy pamiętać, ze Y jest pusty. Mężczyzna jest w tym wypadku hemizygota.
Chorobami sprzężonymi z płcią są: daltonizm, hemofilia, dystrofia Duchenne'a i wiele innych.
13. Zmienność mutacyjna i rekombinacyjna.
Zmienność- występowanie dziedzicznych lub niedziedzicznych różnic pomiędzy komorami danego organizmu(zmienność wewnątrzosobnicza), miedzy osobnikami tej samej populacji(zmienność osobnicza) lub miedzy populacjami(zmienność grupowa)
Zmienność może mieć charakter:
-ciągły-zmienność fluktuacyjna(wzrost, pigmentacja skóry)
-nieciągły-zmienność alternatywna(układ grupowy RH)
Rozwój organizmu zależy zarówno od genotypu jak i od warunków środowiska. Dokonano podziału chorób ze względu na te czynniki:
-choroby uwarunkowane tylko genotypem(hemofilia, zespół Downa)
-choroby uwarunkowane genotypem i warunkami środowiska(cukrzyca, choroba wieńcowa)
-choroby uwarunkowane głównie warunkami środowiska(dur brzuszny, pełzakowica)
Rekombinacja-proces wymiany fragmentów DNA miedzy chromosomami homologicznymi lub dwuniciowymi helisami DNA. Nie prowadzi ona do wytwarzania nowych alleli, lecz do ich przetasowywania. Wyróżniamy rekombinację:
-homologiczna
-zlokalizowana
-transpozycyjna
U organizmów prokariotycznych zachodzi w wyniku:
-losowego doboru koniugantów
-rekombinacji zlokalizowanej, transpozycyjnej lub homologicznej podczas koniugacji
U organizmów eukariotycznych zachodzi w wyniku:
- losowej segregacji chromosomów podczas tworzenia gamet
- losowego doboru rodziców
- losowego łączenia się gamet
- rekombinacji zlokalizowanej, transpozycyjnej i homologicznej.
Rekombinacja homologiczna (crossing-over)
- zachodzi w wyniku symetrycznych pęknięć i ponownego połączenia się odcinków chromatyd chromosomów homologicznych po ich wzajemnej wymianie w pachytenie i/lub diplotenie mejozy. Mitotyczny crossing over występuje bardzo rzadko. Na podstawie częstości crossing over można ustalić lokalizację genów w nici DNA i określić ich wzajemne położenie.
Rekombinacja zlokalizowana
- dotyczy wymiany niehomologicznych, ale specyficznych fragmentów. Katalizowana jest przez białka rozpoznające specyficzne sekwencje zasad. Jej przykładem jest tworzenie przeciwciał i receptorów limfocytów T.
Rekombinacja transpozycyjna
- zachodzi podczas wbudowywania transpozonów w nowe miejsce genomu. Transpozony są ruchomymi elementami genomu zawierającymi geny kodujące transpozazę, czyli enzym o właściwościach nukleazy(np. sekwencje inercyjne). Oprócz genów transpozazy transpozon może zawierać inne geny np. geny oporności na antybiotyki.
Mutacje
- zmiany sekwencji nukleotydów w obrębie genu na inną od sekwencji nukleotydów genu wyjściowego
Punktowe
Tranzycja
zmiana puryny na purynę, lub pirymidyny na pirymidynę
Transwersja
zmiana putyny na pirymidynę lub pirymidyny na purynę
Delecja
wypadnięcie pojedynczej lub większej liczby par nukleotydów z danego genu
Inwersja
wstawienie pojedynczej lub większej liczby par nukleotydów do danego genu
Mogą prowadzić one do:
-zmiany sekwencji aminokwasów w białku
-przerwania syntezy łańcucha polipeptydowego jeśli powstanie triplet nonsensowny
-mutacji niemej jeśli powstanie triplet synonimiczny
Wynikiem mutacji genowych u człowieka są choroby monogenowe (np. mukowiscydoza, fenyloketonuria, albinizm)
Chromosomowe
- wszelkie zmiany w liczbie i strukturze chromosomów
- mogą występować w komórkach somatycznych lub gametach
- mogą być dziedziczone lub powstawać de Novo
14. Zespoły aberracji chromosomowych.
Aberracje strukturalne:
Delecja
utrata fragmentu chromosomu
- terminalna- utrata końcowej części chromosomu
- interstycjalna- utrata środkowego fragmentu chromosomu
Odcinki niektórych chromosomów wykazują szczególną łamliwość, są to np. odcinek 13q chromosomu 2, odcinek 27q chromosomu X oraz chromosomy 6,9,12,20
Inwersja
odwrócenie odcinka
- pericentryczna(eucentryczna)- inwersja obejmująca odcinek chromosomu wraz z centromerem
- paracenteryczna(acentryczna)- inwersja obejmująca odcinek chromosomu bez centromeru
Translokacja
przemieszczenie części chromosomu w inne miejsce tego samego chromosomu bądź w obręb innego chromosomu
Duplikacja
podwojenie fragmentu chromosomu
Utworzenie chromosomu kolistego
w wyniku pęknięcia, a następnie połączenia się końców chromosomu.
- najczęściej zjawisko to zachodzi w chromosomach: 4, 13, 18, X
Utworzenie chromosomu dwucentromerowego
Utworzenie izochromosomu
w wyniku poprzecznego podziału centromeru chromosomu metafazowego
- składa się on tylko z połączonych ramion krótkich lub długich
Aberracje liczbowe:
- powstają w wyniku nierozdzielenia się(nondysjunkci) par chromosomów homologicznych w czasie podziałów. Dzielą się na:
-aneuploidie- powstają w wyniku zwiększenia lub zmniejszenia diploidalnej liczny chromosomów o pojedyncze chromosomy np. zespół Patau, zespół Downa, zespół Turnera, zespół Klinefeltera
-euploidie- polegają na zwielokrotnieniu całego haploidalnego zestawu chromosomów(np. 3n, 4n, 5n)
a)autopoliploidie- garnitur chromosomowy jest zwielokrotniony o ten sam zestaw genów, u człowieka są letalne
b)allopoliploidie- występowanie dwóch lub więcej niehomologicznych zespołów chromosomów, nie występują u człowieka
CHOROBA |
PRZYCZYNA |
CZĘSTOŚĆ WYSTĘP. |
OBJAWY |
INFORMACJE DODATKOWE |
Zespół Downa |
Trisomia 21 pary chromosomów |
1:700, wzrasta wraz z wiekiem rodziców |
Upośledzenie umysłowe, niski wzrost, przedwczesne starzenie się, większa zapadalność na białaczkę i chorobę Alzheimera, bezpłodność w przypadku mężczyzn, skośne ustawienie szpar powiekowych, obniżone napięcie mięśniowe, opuszczone kąciki ust, szeroka przestrzeń między I i II palcem u stopy, otwarte usta zapadnięty grzbiet nosa,krótka szyja, zmarszczka nakątna krótkogłowie, wąskie podniebienie, krótkie i szerokie dłonie, małpia bruzda, zniekształcone małżowiny uszne, duży i pobrużdżony język, plamki Brushfielda na tęczówce, wrodzone wady serca |
Ponad 60% zarodków i płodów z trisomią 21 ulega samoistnemu poronieniu we wczesnym okresie rozwoju |
Zespół Pataua |
Dodatkowy chromosom pary 13; |
1:8000 -1:10 000 |
Mikrocefalia, ubytki skóry na głowie, wystające czoło, rozszczep wargi i podniebienia, wady gałek ocznych, hipoteloryzm, nisko osadzone uszy, polidaktylia, syndaktylia, nerki torbielowate, wodonercze, ubytki w przegrodach serca, macica dwurożna, złączenie płatów czołowych mózgowia, brak opuszek węchowych, niedorozwój móżdżku, wodogłowie, hipotonia mięśniowa, głuchota, hipersegmentacja jąder granulocytów |
70% dzieci z tym zespołem umiera w ciągu pierwszego półrocza życia. 10% dzieci przeżywa do 1 roku życia |
Zespół Edwardsa |
Trisomia chromosomu 18
|
1:5000 |
Niska waga urodzeniowa, małogłowie, wystająca potylica, nisko osadzone i zniekształcone uszy, hiperteloryzm, zmarszczka nakątna, krótka szyja z widocznym fałdem skórnym, zaciśnięte w pięści dłonie, nakładanie się palców ręki, niedorozwój płytki paznokciowej, stopa „cepowata” z wystającą kością piętową, krótkim paluchem i zrostami palców, ubytki w przegrodach serca, podkowiasta nerka, niedorozwój zewnętrznych narządów płciowych u dziewczynek i niezstąpienie jąder u chłopców, zaburzenie rozwoju psychoruchowego |
W 30% przypadków następuje zgon w okresie noworodkowym. 10% dzieci przeżywa 1 rok |
Trisomia chromosomu 8 |
Trisomia chromosomu 8
|
|
Zahamowanie wzrostu, zaburzenia w budowie twarzoczaszki, anomalie kostno-stawowe, wysokie czoło, mała i cofnięta żuchwa, duże, odstające i nisko osadzone uszy, duży i zadarty nos, hiperteloryzm, skrzywienia kręgosłupa, dodatkowe kręgi i żebra, rozszczep kręgów, brak rzepki, wodonercze, spodzectwo i niezstąpienie jąder u chłopców, bruzda w obrębie dłoni i stóp, upośledzenie umysłowe |
|
Delecja długich ramion Chromosomu 13 |
Delecja długich ramion Chromosomu 13 |
|
Mikrocefalia, zniekształcenie twarzoczaszki, wąskie szpary powiekowe, zmarszczka nakątna, wady tęczówki, zaćma, szeroki grzbiet nosa, duże i nisko polożone uszy, krótka szyja, zarośnięcie odbytu, spodziectwo, niezstąpienie jąder, zaburzenia budowy moszny, wrodzone zwichnięcie stawów biodrowych, stopy końsko-szpotawe, niedorozwój kciuka, wytępowanie siatkówczaka |
|
Zespół Pradera-Williego |
Delecja interstycjalna długiego ramienia chromosomu 15 |
1:10 000 - 1:15 000 |
Hipotonia mięśniowa, opóźniony rozwój psychoruchowy, hipogonadyzm hipogonadotropowy, nadmierne łaknienie, węższe czoło, antymongoloidalne ustawienie szpar powiekowych, "migdałowaty" kształt szpar powiekowych, skierowane do dołu kąciki ust, małe i trójkątne usta ("rybie"), wąska czerwień wargowa górnej wargi, małe dłonie i stopy (akromikria), zwężające się ku końcom palce; zez zbieżny; niezwykle gęsta ślina, zwiększająca podatność na próchnicę |
|
Zespół Angelmana |
Delecja fragmentu chromosomu 15 w regionie 15q11-q13 |
|
upośledzenie umysłowe, ataksja, padaczka, charakterystyczne ruchy przypominające marionetkę i napady śmiechu, duże usta, wystający język szeroko rozstawione zęby, tęczówki prawie zawsze mają kolor niebieski znaczne zaburzenia rozwoju mowy, |
|
Zespół Di George'a |
delecja 22q11.2 |
1:4 000 |
atrezja tętnicy płucnej z ubytkiem przegrody międzykomorowej, przerwanie łuku aorty typu B (30%), ubytek przegrody międzyprzedsionkowej, zaburzenia rozwoju podniebienia (69%), trudności w uczeniu (70-90%), pierwotny niedobór odporności (77%) spowodowany aplazją grasicy, hipokalcemia, wrodzone wady nerek, choroby autoimmunologiczne, większe prawdopodobieństwo wystąpienie chorób psychicznych |
|
Zespół Turnera |
Monosomia chromosomu X lub zmiany strukturalne jednego z chromosomów X u płci żeńskiej |
1:3000 |
Waga i wzrost przy urodzeniu niższe od średniej, brak skoku pokwitaniowego, obrzęki limfatyczne rąk i stóp, wysokie czoło, szeroka nasada nosa, szerokie szpary powiekowe, hiperteloryzm, zmarszczka nakątna, krótka szyja płetwistość szyi, niski wzrost, krępa budowa ciała, brak talii, koślawość łokci, klatka piersiowa szeroka, uwypuklona na boki, zewnętrzne narządy płciowe niedorozwinięte, skąpe owłosienie łonowe i pachowe, wczesne występowanie zmarszczek, zwężenie łuku aorty, zmiany w kościach czaszki i wczesne występowanie osteoporozy, nerka podkowiasta, podwójna nerka i podwójne tętnice nerkowe, mała i spłaszczona macica, dysgenetyczne gonady, brak miesiączki, pierwotna bezpłodność, trudności uczenia się, bezpłodność, |
|
Zespół kobiety, 47, XXX |
Trisomia chromosomu X |
1:1000 |
Upośledzenie umysłowe w stopniu lekkim, wcześniejsza menopauza, zaburzenia miesiączki. |
|
Zespół Klinefeltera |
Obecność dodatkowego chromosomu X u osoby płci męskiej |
1:1000 |
Wysoki wzrost, słaby zarost, chłopięcy wygląd, linie włosów półkolista, sylwetka typu kobiecego, ginekomastia, skąpe owłosienie pachowe i łonowe, brak zębów ósmych, nieznaczne upośledzenie umysłowe, prącie rozmiarów od małych do normalnych, słabo rozwinięta moszna, małe jądra, pierwotna bezpłodność, stopniowy zanik potencji, |
|
Zespół mężczyzny, 47, XYY |
Obecność dodatkowego chromosomu Y u osoby płci męskiej |
1:1000 |
Wysoki wzrost, typowo męska budowa ciała, bliznowate zmiany na skórze twarzy i pleców, nadmierna agresywność, przewaga długości śródręcza nad paliczkami, zmniejszenie kąta czaszki, progenia |
|
15. Genom mitochondrialny.
Genom mitochondrialny
- najwięcej mitochondriów występuje w tkance mięśniowej poprzecznie prążkowanej, mięśniu sercowym, nerkach, OUN
- DNA mitochondrialny nie ulega rekombinacji, nie ma systemów naprawczych
- kumulacja zmian mutacyjnych- dużo mutacji jest wynikiem powstawania wolnych rodników podczas oddychania wewnątrzkomórkowego
- dziedziczy się po matce
-dwuniciowa kolista cząsteczka DNA - nić ciężka i lekka
-22 geny kodujące tRNA, 2 geny rRNA i 13 regionów kodujących białka
- brak intronów
-brak kodu terminacyjnego - ramki odczytu kończą się na U lub UA, a kodon ochra jest tworzony przez poliadenylacje transkryptu
16. Mutacje u Drosophila melanogaster.
Mutacja recesywna w stosunku do typu dzikiego.
1. Yellow - y - zmutowany gen warunkujący barwę ciała, barwa ciała żółta.
2. Ebon -e- zmutowany gen warunkujący barwę ciała, barwa ciała ciemnoszara.
3. Vestigal - vg- zmutowany gen warunkujący wielkość skrzydeł, skrzydła szczątkowe.
4. White -w- zmutowany gen warunkujący barwę oka, barwa oka biała.
5. Bar -B- mutacja chromosomowa strukturalna, podwojenie odcinka chromosomu pierwszego, w którym znajduje się gen na kształt oka.
17. Podstawowe pojęcia w biologii rozwoju: determinanty cytoplazmatyczne, indukcja, kompetencja, totipotencja, morfogeneza, amplifikacja, chromosomy olbrzymie.
Totipotencja (totipotencjalność, ekwipotencjalność rozwojowa)
zdolność pojedynczej komórki do zróżnicowania się w każdy typ komórkowy organizmu. Z definicji komórką totipotencjalną jest zygota. U ssaków totipotipotencją charakteryzują się komórki macierzyste i komórki zarodkowe węzła zarodkowego blastocysty. U roślin własność totipotencji mają komórki merystematyczne, choć potencjalnie każda komórka roślinna może ulec odróżnicowaniu w procesie regeneracji do kallusa i osiągnąć stan toti- lub pluripotencji.
Amplifikacja
- dotyczy szczególnie komórek jajowych z których po zapłodnieniu rozwijają się zarodki stale potrzebujące podstawowych białke związanych z aparatem mitotycznym
- polega na zwielokrotnieniu odcinków rDNA - genów rRNA znajdujących się w chromosomach jąderkotwórczych
- strategie amplifikacyjne:
Jajniki panoistyczne - wszystkie komórki przekształcają się w okresie dojrzewania w gamety
Jajniki meroistyczne - część komórek funkcjonuje jako gamety a pozostałe odżywiają komórki jajowe i produkują rybosomy są połączone z k.jajowa za pomocą mostków plazmatycznych (trofocyty), u stawonogów i pierścienic
Strategia tylko dla jajnika meroistycznego:
k. jajowa zapotrzebowanie na rybosomy genomy ulegają poliploizacji w procesie endoreplikacji (wielokrotny podział chromosomów bez ich rozdziału) amplifikacja genu rRNA
Strategia dla jajnika heroistycznego i panoistycznego:
profaza mejotyczna (pachyten) koliste cząsteczki uwalniają się z rejonu organizatora jąderka NOR replikacja uwolnionego rDNA przez dodawania zasad DNA do otwartego końca 3' jej nici amplifikacja
Chromosomy olbrzymie
- politeniczne - powstają w efekcie wielu endoreplikacji chromatyd po których nie następuje podział mitotyczny; mają prążkowaną strukturę (wynik periodycznego ułożenia obok siebie chromomerów) i obecność jednego lub kilku charakterystycznych pierścieniowatych obrzmień zwanych pierścieniami Balbiniego; rejony nie wybrawiające się to rejony międzyprażkowe
- szczoteczkowe - występują w I profazie mejotycznej w diplotenie; homologi połączone są w miejscach określanych chiazmami których chromatydy siostrzane krzyżują się dochodzi do przejściowej dekondensacji tworzy się pętla chromatynowa a chromosomy przybierają formy szczoteczkowate, aktywne chromosomy szczoteczkowe prowadzą wzmożoną syntezę rRNA
Morfogeneza
biologiczne procesy rozwojowe, w wyniku których jest determinowany kształt zarodka w kolejnych stadiach rozwojowych i ostatecznie kształt dorosłego organizmu. Pojęcie obejmuje także kształtowanie się ostatecznej formy narządów i części organizmów, a w grupie protista kształtowanie się właściwych dla nich organelli. Bruzdkowanie (ang. cleavage), segmentacja, pierwszy etap rozwoju zarodkowego zwierząt tkankowych; seria mitotycznych podziałów zapłodnionej (lub rozwijającej się partenogenetycznie) komórki jajowej na coraz mniejsze komórki - blastomery; w wyniku bruzdkowania powstaje morula, a następnie blastula. W czasie bruzdkowania ani masa, ani objętość, ani zasadniczy kształt zarodka nie ulegają zmianie. Przebieg bruzdkowania jest charakterystyczny i stały dla danej grupy zwierząt.
Typ bruzdkowania oraz budowa blastuli zależy od ilości i rozmieszczenia żółtka w jaju:
- bruzdkowanie całkowite - w jajach o małej i średniej ilości żółtka; podziałom ulega cała komórka:
- bruzdkowanie całkowite równomierne - jaja z małą ilością żółtka (alecytalne i oligolecytalne); dzielą się w całości na jednakowe blastomery. Występuje u jeżowców, osłonic, lancetnika, większości ssaków
- bruzdkowanie całkowite nierównomierne - jaja o średniej ilości żółtka (mezolecytalne), nagromadzonego w półkuli wegetatywnej (telolecytalne); dzielą się na większe komórki (makromery) i mniejsze komórki (mikromery). Mikromery, w przeciwieństwie do makromerów, nie zawierają prawie w ogóle żółtka, więc dzielą się szybciej. Tak dzielą się zygoty płazów.
- bruzdkowanie częściowe - w jajach o dużej ilości żółtka (polilecytalnych); podziałom podlegają jądra komórkowe wraz z cytoplazmą nie zawierającą żółtka:
- bruzdkowanie częściowe powierzchniowe - przy dużej ilości żółtka zgrupowanego w środku jaja (jajo centrolecytalne); dzieli się tylko cytoplazma na powierzchni jaja; występuje u wielu stawonogów.
- bruzdkowanie częściowe tarczkowe - w jajach telolecytalnych; podziałowi ulega tylko tarczka cytoplazmy na biegunie nie zawierającym żółtka; występuje u ptaków, ryb kostnoszkieletowych, stekowców.
Kompetencja
- zdolność reagowania zespołów komórek na bodziec morfogenetyczny, a tym samym występowanie komórek na określona drogę różnicowania
- zdolność komórek bakteryjnych do ulegania transformacji.
Indukcja
- wyzwolenie określonego procesu w organizmie przez proces go poprzedzający np.: bodziec zewnętrzny lub czynnik wewnętrzny.
18. Rola heterochromosomów w determinacji płci u człowieka.
X i Y są odpowiedzialne za prawidłowy rozwój cech płciowych - biosyntezę białek strukturalnych i regulatorowych niezbędnych do:
- prawidłowego różnicowania się pierwotnej gonady w kierunku jądra lub jajnika
- rozwoju wewnętrznych i zewnętrznych narządów płciowych.
PAR - region pseudoautosomalny - końce ramion krótkich od Yp11.3 i Xp22.3 - podczas mejozy u faceta te regiony tworzą tzw. pęcherzyk płciowy i zachodzi między nimi c.-o.
U kobiet niezbędna jest obecność 2×X (XX), przy jednym tw. się dysplastyczne jajniki bez gonocytów.
Na Y są geny odpowiedzialne za wykształcenie przez zarodek cech męskich, osobnik z Y zawsze wykazuje cechy męskie
- jest tam pojedynczy gen kodujący białko odpowiedzialne za rozwój jąder - biosyntezę TDF (Testis Determining Factor) czyli czynnika determinującego rozwój jądra;
- Yp11.3 jest gen SRY (Sex determining Region of Y) kodujący białko SRY wykazujące pokrewieństwo z kilkoma czynnikami transkrypcyjnymi.
SRY:
- gł. msc transkrypcji SRY są prekursory komórek Sertolego, na namnażanie których w okresie płodowym wpływa gonadotropina komówkowa (hCG) i FSH (zwiększa też w kom. Leydiga syntezę receptorów dla LH pobudzającego w nich syntezę testosteronu)
- aktywator transkrypcji genu kodującego czynnik antymüllerowski (AMH) odp. za zanik przewodów przyśródnerczowych Müllera
- negatywny regulator transkrypcji genu aromatazy P450, która katalizuje przemianę testosteronu w estradiol
Ekspresja genów
- wieloetapowy proces prowadzący do ujawnienia zawartej w genie informacji o biosyntezie białka lub RNA
Pierwszym etapem e. g. jest transkrypcja, jej produktem są prekursory RNA(transportujące tRNA, rybosomalne rRNA, informacyjny mRNA, mało jądrowy snRNA), następnie dochodzi do posttranskrypcyjnego dojrzewania RNA, dojrzale cząstki pełnio swoją funkcję biologiczną jako cząsteczki RNA.
Kolejnym etapem jest translacja , cząsteczki mRNA zostają przetłumaczone na sekwencję aminokwasową łańcucha polipeptydowego, one również muszą ulec obróbce postranslacyjnej- dojrzewanie postranslacyjne.
GEN---mRNA---polipeptyd
U eukariontów oba procesy są rozdzielone w czasie i przestrzeni.
--== ROZWÓJ ==--
19. Zróżnicowanie gonad i komórek płciowych.
Oogeneza:
- powstają 1 komórka jajowa i 3 ciałka kierunkowe II rzędu, które ulegają degeneracji
- poza zmianami morfologicznymi w procesie oogenezy zachodzą też zmiany funkcjonalne:
-rekombinacja genetyczna
-amplifikacja genów (część genomu- organizator jąderko twórczy, ulega poliploidyzacji)
-pojawienie się chromosomów szczoteczkowych
-synteza mRNA i rRNA (warunkują prawidłowy przebieg wczesnych etapów rozwoju zarodka
Spermatogeneza:
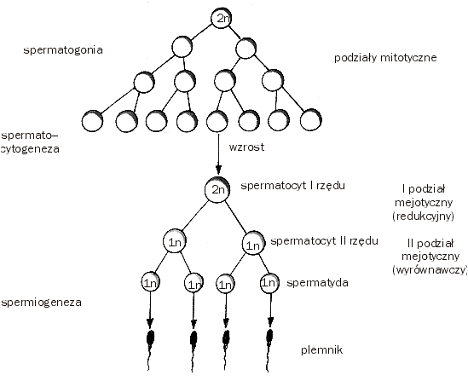
Porównanie spermatogenezy i oogenezy:
Cechy wspólne:
- proces obejmuje mejoze, w której liczba chromosomów ulega redukcji
- obejmują zmiany morfologiczne polegające na wyposażeniu gamet do pełnienia określonej roli w zapłodnieniu
- gameta może przeżyc długo jeżeli nie zachodzi zapłodnienie
Różnice:
- podziały mejotyczne w spermatogenezie są równomierne prowadzą do wytworzenia 4 równorzędnych spermatyd, natomiast w oogenezie podziały są niesymetryczne i powstaje tylko jedno jajo oraz ciałka kierunkowe
- mejoza w spermatogenezie nie ulega zahamowaniu a w oogenezie jest blokowana przynajmniej w jednym stadium (profaza 1 podziału do osiągnięcia dojrzałości płciowej i oocyty czekające na zapłodnienie)
- jajo rozwija się stopniowo, a plemiki rozwijają się ze spermatyd gwałtownie
- zmiany morfologiczne w plemniku polegaja na wykształceniu witki, kondensacji jądra, odrzuceniu cytoplazmy, powstaniu akrosomu, w jaju następuja wzrost rozmiarów, gromadzenie materiałów zapasowych, wydzielanie białek tworzących zewnętrzne warstwy ochronne
Budowa plemnika:
Budowa komórki jajowej:
Od zewnątrz kolejno znajduje się:
-otoczka galaretowata u jeżowców i żab lub wieniec promienisty u ssaków
-osłonka żółtkowa (u ssaków przejrzysta)
-cytoplazma korowa z ziarnami korowymi
-pęcherzyk zarodkowy (jądro)
20. Kontakt i zapłodnienie: przedjądrze męskie i żeńskie, zapobieganie polispermii.
-dostanie się plemnika do żeńskiego układu rozrodczego
-proces kapacytacji plemnika (usunięcie białek i cukrów blokujących połączenia plemnika z osłonką przejrzystą)
-reakcja akrosomowi (uwalnianie enzymów uszkadzających osłonkę przejrzystą)
-fuzja (bezpośredni kontakt błon cytoplazmatycznych jaja i plemnika)
-zapłodnienie: jądro plemnika i jądro jaja tworzą przedjądrze męskie i żeńskie, które ulegają fuzji
Zapobieganie polispermii:
Po wniknięciu pierwszego plemnika wejście następnych do komórki jajowej jest blokowane.
U jeżowców występują dwa mechanizmy:
-szybkiego blokowania, w którym plemnik powoduje depolaryzację błony jaja
-mechanizm powolny, w którym ziarna korowe zlewają się z błoną cytoplazmatyczną uwalniając substancje zmieniające właściwości osłonki żółtkowej
U ssaków reakcja ziaren korowych prowadzi do odczepienia pozostałych plemników do osłonki przejrzystej.
21. Bruzdkowanie: przejście poblastulacyjne, sposoby bruzdkowania, wpływ żółtka, typy blastuli, tworzenia blastocelu.
Bruzdkowanie- seria szybko po sobie zachodzących podziałów mitotycznych występujących zaraz po zapłodnieniu . Termin ten wywodzi się od pojawiających się na powierzchni zygoty charakterystycznych bruzd będących wynikiem cytokinezy. Powstałe w wyniku tego procesu komórki nazywamy blastomerami.
Sposoby bruzdkowania
bruzdkowanie promieniste - pierwsze dwa podziały w płaszczyźnie południkowej (pionowej) , wzajemnie prostopadłe do siebie, następne podziały naprzemiennie: południkowo i równoleżnikowo (poziomo)
bruzdkowanie spiralne - podłużne płaszczyzny podziałowe odchylone są pod pewnym kątem do głównej osi jaja, a blastomery układają się naprzemianlegle. Pierwsze dwa podziały są południkowe trzeci natomiast przebiega skośnie, ponieważ pozycja wrzeciona odchylona jest o pewien kąt.
bruzdkowanie rotacyjne (charakterystyczne min dla zarodka ssaków) -pierwsza płaszczyzna podziałowa jest pionowa i dzieli jajo na dwa blastomery nierównej wielkości, natomiast w drugim podziale dzieli się tylko jeden większy elastomer, a bruzda podziałowa przebiega poziomo. Potem pionowo dzieli się drugi elastomer - w tym momencie zarodek składa się z 4 blastomerów ułożonych na krzyż.
bruzdkowanie dwubocznie symetryczne (zarodki żachw) -pierwszy podział wyznacza lewą i prawą stronę zarodka, a wszystkie późniejsze podziały każdej ze stron są lustrzanym odbiciem drugiej.
LUB
bruzdkowanie całkowite (holoblastyczne) - podziałom ulega cała komórka (wys. u jeżowców ssaków)
bruzdkowanie częściowe (meroblastyczne)-proces podziału ogranicza się do niewielkiego pola cytoplazmy na biegunie animalnym jaja zwanym tarczą zarodkową. (wyst. u ryb, gadów, ptaków )
Wpływ żółtka na przebieg bruzdkowania
Istotny wpływ na sposób bruzdkowania ma ilość żółtka i jego rozmieszczenie w cytoplazmie jaja. Wpływa na tempo podziałów komórkowych zarodka, przebieg bruzd podziałowych oraz na kształt i wielkość powstających blastomerów. Ogólnie obecność żółtka w jaju opóźnia lub wręcz hamuje podziały komórkowe, dlatego blastomery w różnych częściach zarodka u wielu gatunków zwierząt mogą różnić się wielkością.
Rodzaje jaj w zależności od zawartości żółtka:
Alecytalne- bezżółtkowe
Oligolecytalne- małożółtkowe
Komórki jajowe typu a i b - robaki płaskie jamochłony a także ssaki
Mezolecytlne- średniożółtkowe : płazy
Polilecytalne- bogato żółtkowe : owady, gady, ptaki, ryby chrzęstnoszkieletowe, ryby kostnoszkieletowe
Rodzaje jaj pod względem rozmieszczenia żółtka:
Izolecytalne- z żółtkiem rozmieszczonym równomiernie
Anizolecytalne- z żółtkiem rozmieszczonym nierównomiernie
Centrolecytalne- żółtko skupione centralnie a cytoplazma na obwodzie (owady)
Telolecytalne- z żółtkiem na jednym biegunie a z cytoplazmą na przeciwległym (ptaki)
Bruzdkowanie holoblastyczne (całkowite)- zachodzi w komórkach jajowych alecytalnych, oligolecytalnych i mezolecytalnych. Pierwsza bruzda podziałowa ma przebieg południkowy tzn. łączy biegun animalny z wegetatywnym i dzieli zarodek na dwa blastomery jednakowej wielkości. Druga bruzda podziałowa ma również przebieg południkowy i następuje pod kątem 90 stopni stosunku do płaszczyzny pierwszej bruzdy , dając zarodek 4 blastomerowy. Trzecia bruzda podziałowa przebiega:
W płaszczyźnie równika dając 8 jednakowych blastomerów- bruzdkowanie całkowite równomierne
W płaszczyźnie równoleżnikowej( bliżej bieguna animalnego), dając 4 większe makromery na biegunie wegetatywnym i 4 mniejsze mikromery na biegunie animalnym
Bruzdkowanie całkowite nierównomierne- występuje np. u płazów u których żółtko nie jest rozmieszczone równomiernie a bruzdkowanie ulega hamowaniu w regionie bogatym żółtko ( w jajach płazów żółtko jest zmagazynowane na biegunie wegetatywnym jaja natomiast jadro zepchnięte zostaje na biegun animalny czyli przeciwległy który jest pozbawiony żółtka i choć zarodek ulega całkowitym podziałom to na biegunie animalnym zachodzą one szybciej niż na biegunie obarczonym żółtkiem .
Kolejne bruzdy mają przebieg prostopadły do poprzedniej i dzielą się równocześnie w sposób synchroniczny tzn. wszystkie blastomery na coraz to mniejsze komórki.
Bruzdkowanie meroblastyczne (niecałkowite) - zachodzi w komórkach jajowych polilecytalnych w tej części jaj gdzie występuje większe nagromadzenie cytoplazmy
Polilecytalne- telolecytalne u ptaków bruzdkuje tylko tarcza zarodkowa zlokalizowana na powierzchni kuli żółtka - bruzdkowanie meroblastyczne tarczowe
Polilecytalne- centrolecytalne u owadów ma charakter mezolecytalny powierzchniowy - w jego wyniku powstaje wielokomórkowa morula, która po przegrupowaniu komórek przekształca się w blastulę. Proces ten polega najogólniej na tym, że układają się one powierzchniowo tworząc jedną lub kilkuwarstwową blastodermę okalającą jamę czyli blastocel.
Przejście poblastulacyjne (MBT) - jest to proces wyłączania genomu matczynego i uaktywniania się genomu zarodka, jest to kluczowy moment w rozwoju wszystkich zwierząt. (wczesny rozwój zarodka kontrolują produkty genów matczynych, zgromadzone w jaju w oogenezy, lecz robią to tylko do określonego momentu, gdyż stają się nieaktywne bądź zostają zużyte. Brak produktów genów matczynych, sprawia że zostają uruchomiony genom zarodka i wszystkie procesy rozwojowe zachodzą pod kontrolą genów zygotycznych. Przejście poblastulacyjne może być nagłe i zbiegać się z gwałtownym spowolnieniem podziałów komórkowych, zanikiem synchronii podziałów i zmianami zachowania się komórek, np. stają się ruchliwe i zaczynają się przemieszczać.
U zarodków Drosophila stadium MBT występuje dopiero po 14 podziale jądra komórkowego, kiedy to zostają wyczerpane zapasy białka String które jest matczynym regulatorem cyklu komórkowego blastomerów. Wyczerpanie owego białka powoduje uaktywnienie genomu zarodka i dalsze podziały bruzdkowania zachodzą pod kontrolą zygotycznej transkrypcji genu String.
U zarodków Danio i Xenopus MBT występuje po tym jak produkty genów matczynych zostają związane z wzrastającą ilością DNA. W cytoplazmie jaja zgromadzony jest czynnik który hamuje ekspresje genów zygotycznych. U zarodków Dnio wystarcza na hamowanie transkrypcji zygotycznej w 9 kolejnych podziałach, natomiast u Xenopus w 12 podziałach i po tym ostatnim podziale uaktywniony zostaje genom zarodka. Prawdopodobnie dzieje się tak ponieważ wszystkie wolne inhibitory obecne w zarodku zostały już związane z DNA. Synchronizację MBT można kontrolować poprzez modyfikację zawartości DNA w zygocie.
Tworzenie blastocelu - jest to proces w którym tworzy się wypełniona płynem wewnętrzna jama zwana blastocelem, blastocel może pojawiać się na różnych etapach procesu bruzdkowania. Może zacząć się wcześnie po pierwszym lub po kilku podziałach komórkowych i osiągać coraz większe rozmiary albo pojawia się później i bardzo gwałtownie. Blastocel stanowi miejsce dla złożonych ruchów komórek zachodzących w czasie gastrulacji oraz zabezpiecza przed przedwczesnym kontaktem między grupami komórek zarodka które w późniejszych etapach rozwoju wchodzą w interakcje indukcyjne. Blastocel tworzy się z niewielkich szczelin którw powstają na skutek rozsuwania się komórek podczas podziałów bruzdkowania, a następnie powiększają się i tworzą obszerna jamę. Początkowo szczeliny tworzą się biernie jako naturalna konsekwencja rozluźnienia upakowania blastomerów.
U jeżowców wczesne promieniste podziały komórkowe prowadzą do utworzenia blastuli, w której centralna jama otoczona jest pojedynczą warstwą komórek. Wszystkie późniejsze podziały komórek zachodzą w płaszczyźnie pojedynczej warstwy co powoduje wzrost liczby komórek w Obrębie tej warstwy bez wzrostu ich wielkości. W konsekwencji komórki stają się coraz mniejsze i nie mogą wypełniać wewnętrznej przestrzeni zarodka, co inicjuje tworzenie się wewnętrznej jamy . u zarodków jeżowca i ssaków do przestrzeni międzykomórkowych bruzdkującego zarodka pompowane są również jony, które wciągają wodę na drodze osmozy. Ruch płynu powiększa rozmiary blastocelu i utrzymuje integralność zarodka poprzez wzrost ciśnienia hydrostatycznego.
Typy blastuli:
Celoblastula (blastula właściwa)- występuje np. u strzykw i lancetnika. Komórki układają się powierzchniowo tworząc jedną lub kilkuwarstwową blastodermę, okalająca różnej wielkości jamę czyli blastocel. Podobną blastulę spotykamy u płazów, przy czym w związku z ich całkowitym nierównomiernym bruzdkowaniem blastocel zlokalizowany jest niesymetrycznie , przesunięty bliżej bieguna animalnego i okolony wielowarstwowa blastodermą.
Dyskoblastula- występuje u ptaków gdzie w wyniku bruzdkowania i równoczesnego przegrupowania komórek powstaje szczelinowate światło w obrębie tarczki zarodkowej. Blastomery układają się już bardzo wcześnie w dwie warstwy: powierzchniowa epiblast i w warstwę lezącą pod spodem hypoblast. Zlokalizowane między epiblastem a hypoblastem światło jest odpowiednikiem jamy blastuli.
Periblastula- występuje u owadów nie posiada blastocelu i jej przestrzeń zajęta jest w całości przez żółtko
22. Typy regeneracji: fizjologiczna (stała i okresowa), wewnętrzna i pourazowa.
• Fizjologiczna
w zależności od poziomu
- tkankowa
(np. odnowa zużytych krwinek)
- molekularna
-fotopigmentów
(światło rozszczepia połączenie retinenu i białka opsyny, które łączą się
ponownie w serii reakcji z enzymami i ATP i odtwarzają jodopsynę w
czopkach i rodopsynę w pręcikach)
-śródkomórkowa
(np. odtwarzanie błony jądrowej w telofazie)
w zależności od częstości
- stała
(np. paznokcie, plemniki, gruczoły łojowe)
- cykliczna
(np. śluzówka macicy, gruczoły mleczne)
- jednorazowa
(wymiana uzębienia mlecznego na stałe)
w zależności od miejsca
- wewnętrzna
- zewnętrzna
• Pourazowa
- MORFALAKSJA
przestrojenie organizmu po urazie naruszającym prawidłową strukturę ciała
np. u Stylonychii (orzęsek) usunięcie jednego z trzech pól rzęsek prowadzi do
odrzucenia pozostałych
lub
odtworzenie części ciała przez zmianę istniejących tkanek bez ich wzrostu
np. u jamochłonów powstawanie całego, mniejszego osobnika z kawałka ciała
dorosłego
Uwaga! W tym przypadku regeneracja może być uznawana za jeden ze sposobów rozmnażania bezpłciowego.
- EPIMORFOZA
odtworzenie części ciała przez namnożenie komórek nowej tkanki i jej wzrost
np. u płazów ogoniastych odtworzenie kończyny
Regeneracja u bezkręgowców:
Gąbki
Izolowane komórki gąbki potrafią się łączyć, tworzyć skupienia, rosnąć i wytworzyć małego osobnika. Świadczy to o dużej zdolności regeneracyjnej i niskiej specjalizacji komórek.
Pierwotniaki
Regenerują części zawierające chociaż kawałek jądra.
Robaki (na przykładzie wypławka)
- obkurczenie mięśniówki sąsiadującej z raną by zmniejszyć jej powierzchnię
- eliminacja uszkodzonych komórek
- napełzanie nabłonka skórnego by przykryć tkanki wewnętrzne
- napływanie neoblastów (komórki niezróżnicowane) w kierunku miejsca urazu pod zgrubiały nabłonek rany
- utworzenie pączka regeneracyjnego (blastema regeneracyjna)
- wzrost, przegrupowanie i różnicowanie komórek w ostateczne tkanki
Uwaga!
Promieniowanie rentgenowskie powoduje utratę zdolności neoblastów do podziałów, więc regenerują tylko części wypławka nie poddane promieniowaniu
Pierścienice
Mają zdolność do regeneracji utraconych segmentów. Proces postępuje do momentu osiągnięcia liczby segmentów charakterystycznych dla danego gatunku. Regeneruje tylko część zawierająca głowę.
Owady
Ze względu na chitynowy pancerz możliwa jest tylko w czasie linienia.
- zabezpieczenie rany
- odróżnicowanie tkanek
- przekształcenie się komórek wyścielających chitynę zamykająca ranę w mały regenerat kończyny
- wysunięcie się zregenerowanej, mniejszej kończyny podczas kolejnej wylinki
- osiągnięcie normalnych rozmiarów kończyny po kolejnych linieniach
Szkarłupnie
Rozgwiazdy z każdego ramienia mogą odtworzyć cały organizm, ale zregenerowane ramiona są krótsze niż pierwotne.
Podrażnione strzykwy wyrzucają część swoich narządów przez otwór odbytowy i są zdolne do ich późniejszego odtworzenia.
Uwaga!
- proces samookaleczenia przez odrzucenie części ciała to autotomia
- organizmy pasożytnicze nie mają zdolności regeneracyjnych (przywry, tasiemce, nicienie, pijawki) prawdopodobnie ze względu na niskie narażenie na uszkodzenia
Płazy
Etapy procesu regeneracji kończyny u płazów ogoniastych:
- zmniejszenie powierzchni rany przez skurcz mięśniówki naczyń i powstanie skrzepu
- namnożenie komórek ektodermalnych w miejscu amputacji
- nakrycie rany i utworzenie czapeczki ektodermalnej AEC
- pobudzenie przez czapeczkę komórek leżących niżej (chrzęstnych, mięśniowych, skóry właściwej, tkanki łącznej) do odróżnicowania (dedyferencjacji) i utworzenia blastemy regeneracyjnej z niezróżnicowanych komórek mezenchymatycznych
- powstanie wszystkich struktur przyszłej dojrzałej kończyny w procesie ponownego zróżnicowania komórek (redyferencjacji)
Uwaga!
- tkanka mięśniowa i łączna może po odróżnicowaniu stworzyć każdy składnik kończyny
- tkanka chrzęstna po odróżnicowaniu może stworzyć tylko nową chrząstkę
- naskórek nie może utworzyć blastemy i nie może z niej powstać
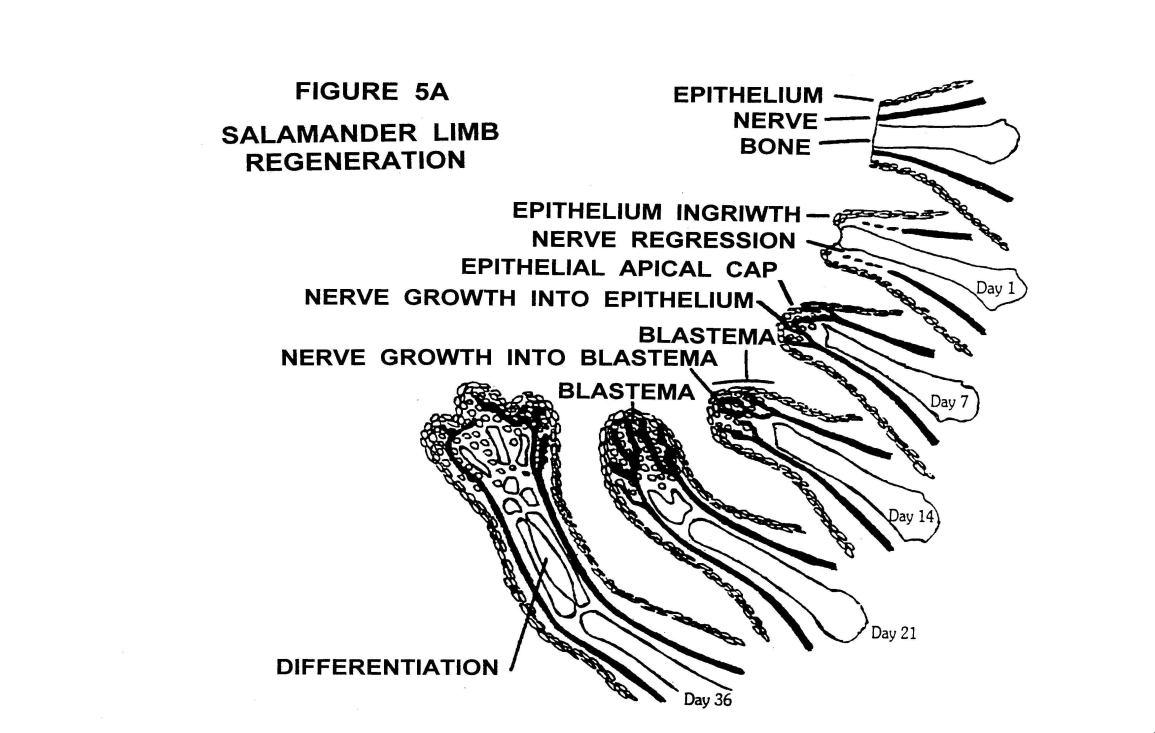
Warunki zachodzenia regeneracji:
- obecność ektodermy (pobudza powstawanie blastemy i utrzymuje jej funkcję)
- obecność nerwów (aksony uwalniają czynniki wzrostowe)
- obecność hormonów przysadki (są konieczne do rozpoczęcia procesu regeneracji)
Uwaga!
- płazy mają zdolność regeneracji zewnętrznej gdyż posiadają najgęstsze unerwienie ze wszystkich kręgowców
- regeneracja zawsze postępuje na zewnątrz (w kierunku dystalnym) niezależnie od miejsca przecięcia i orientacji kończyny prowadząc do odtworzenia brakujących tkanek
Molekularne podstawy specyfikacji regionalnej:
(wpływ substancji produkowanych w miejscu amputacji na pozycję kończyny w przestrzeni)
- kwas retinowy
Produkowany jest przez naskórek i odpowiada za utrzymanie prawidłowej osi przednio-tylnej regenerującej kończyny. W kikucie występuje różne stężenie kwasu retinowego w zależności od jego długości. Im mniej kończyny zostało ucięte, tym mniejsza dawka kwasu i mniej wyraźny efekt. Jest to konieczne be wytworzyć tylko brakujący element, a nie całą kończynę w miejscu ucięcia. W miarę odbudowy kończyny stężenie kwasu stopniowo maleje pozostaje jednak najwyższy, ale dostosowany gradient na jej dystalnym końcu. Służy to utrzymaniu wzrostu we właściwym kierunku.
- geny HoxA
Ulegają ekspresji w regenerującej blastemie krótko po jej powstaniu i pełnią funkcję regulacyjne.
Ssaki
Nie wykazują zdolności do regeneracji zewnętrznej. Zewnętrzny uraz mechaniczny w ssaków kończy się bliznowaceniem.
Są zdolne do regeneracji wewnętrznej jednak bez procesu odróżnicowania komórek (głównie w obrębie wątroby, nerek, płuc, kości i błony śluzowej macicy). Odnowa następuje w procesie hipertrofii (podziały komórek bez ich wzrostu) i hiperplazji (wzrost komórek bez podziałów). Proces ten nazywany jest regeneracją przerostowa lub przerostem regeneracyjnym.
Uwaga!
- przerost jednego z narządów parzystych (nieuszkodzonego) po usunięciu drugiego to kompensacja
- przejście jednej zróżnicowanej tkanki w drugą w zakresie tej samej tkanki np. nabłonek walcowaty w wielowarstwowy płaski to metaplazja (wyjątek stanowi tkanka mięśniowa; gładka nie może przekształcić się w poprzecznie prążkowaną i odwrotnie)
# 1 #
Wyszukiwarka
Podobne podstrony:
Genetyka rozwoju
Genetyka w medycynie regeneratywnej i reprodukcyjnej
13 Genetyka rozwoju
W4-Genetyka rozwoju, analityka medyczna UMP 2014, biologia z genetyką, wykłady
W4 Genetyka rozwoju
Genetyka rozwoju
Składniki pokarmowe pomagające w prawidłowym rozwoju i regeneracji chrząstki stawowej, Zootechnika,
genetyka a rozwój motoryczny
Genetyka rozwoju
Rozwój genetyki jako nauki
Perspektywy rozwoju genetyki
Wykład 5 - Genetyka człowieka, ⇒ NOTATKI, II semestr, Biomedyczne podstawy rozwoju (wykład)
wady genetyczne, studia, Biomedyczne podstawy rozwoju i wychowania
Biomedyczne podstawy rozwoju i wychowania Czynniki genetyczne i środowiskowe wpływające na rozwój
Kolokwium choroby gen bud i f kom, mutacje, tkanki,pasożyty,genetyka,wady,etapy rozwojowe
Rozwoj myslenia Epistemologia genetyczna J, Psychologia rozwojowa i osobowości
medycyna regeneracyjna, Dietetyka CM UMK, Biologia z genetyką
GENETYCZNE PODSTAWY ROZWOJU not Nieznany
Psychologia rozwojowa Teoplitz wykład 2 Podstawy genetyki kiedy zaczyna się człowiek ppt
więcej podobnych podstron