50 51

należy łych terminów wiązać z poziomem złożoności morfologicznej. np. gąbki określane są jako niższy typ. Stawia się je na niskim poziomie w systematyce, ponieważ przyjmuje się, że wyłoniły się wcześnie w ewolucji wielokomórkowców. Nie oznacza to, że mają prostą organizację morfologiczną pod każdym względem. W porównaniu z resztą bezkręgowców, mają np. najbardziej skomplikowany szkielet wewnętrzny, a występujący u nich układ komór i kanałów wodnych, nie obserwowany u reszty zwierząt, może być krańcowo skomplikowany. Termin wyspecjalizowany odnosi się do cech najbardziej charakteryzujących grupę, będących przystosowaniem do życia w określonej niszy ekologicznej. Termin ten nie powinien być utożsamiany z „lepszą” organizacją morfologiczną, gdyż pod względem zasadniczych cech grupa może być prymitywna. Określenia pierwotny i wtórny są używane do rozróżnienia pomiędzy prostą organizacją morfologiczną, którą grupa osiągnęła w procesie ewolucji i poza nią nie wyszła a prostą organizacją nabytą w drodze uwstecz-nienia. Na przykład wirki jelitowe najprawdopodobniej pochodzą od przodków. które nie miały odbytu, u nich budowę układu pokarmowego cechuje brak drożności. U wirków brak odbytu jest więc cechą pierwotną. Wężowidła także nie mają odbytu, ale pochodzą od przodków, które miały odbyt, o czym świadczy zawiązywanie się odbytu u nich w trakcie rozwoju zarodkowego. Brak odbytu jest więc u wężowideł cechą wtórną, nabytą w trakcie ewolucji. Trudności terminologiczne w dużym stopniu eliminują graficzne ujęcia filogenezy. Systematycy od dawna próbowali przedstawiać ewolucję i powiązania rodowe jednostek systematycznych, w postaci różnorodnych wykresów. Do osiemnastego wieku przedstawiano ewolucję świata żywego w postaci „drabiny jestestw” (scala naturae). Najniższy szczebel w drabinie jestestw reprezentował formy martwe, następny — formy ożywione, dalsze szczeble reprezentowały rośliny i zwierzęta, ostatni — człowieka. Takie ujęcie graficzne sugerowało ewolucję ciągłą, od form prymitywnych do coraz to bardziej doskonałych. Pozostałością takiego ujmowania ewolucji są terminy „niższy” i „wyższy”, stosowane do dziś, ale już w innym znaczeniu, jak podaliśmy wyżej. W miarę coraz to lepszego poznawania świata żywego, sugerowana przez drabinę jestestw ciągłość stawała się coraz mniej prawdopodobna. Coraz więcej sys-
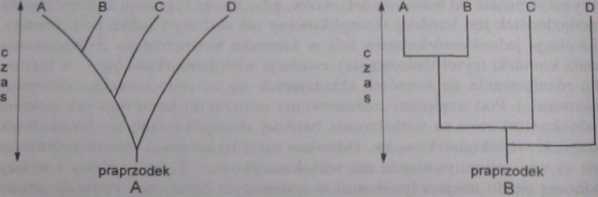
Ryc. 11. Fonogramy. Objaśnienie w tekście
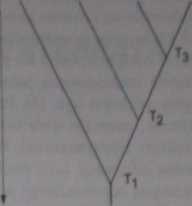
Ryt. 12. Kład ogram Objaśnienie w tekście
O
C
c
z
a
•
praprzodek
tematyków proponowało dodawanie do drabiny jestestw bocznych odgałęzień (m. in. Buffon, Lamarck). Wprowadzenie przez Cuviera typów, jako kategorii systematycznych i rozdzielenie zwierząt pomiędzy cztery niezależne typy. ostatecznie pogrzebało drabinę jestestw. W 1859 r. Ernest Haeckef przedstawił pierwszą próbę graficznego ujęcia filogenetycznych pokrewieństw organizmów żywych, które nie miało postaci ciągłej, miało wygląd drzewa, z pniem i odgałęzieniami. Od tego czasu, jakiekolwiek graficzne ujęcie ewolucyjnych pokrewieństw nazywa się tradycyjnie drzewem filogenetycznym (dendrogra-mem). Obecnie proponowanych jest wiele metod graficznego przedstawiania podobieństw i pokrewieństw jednostek systematycznych. Stosuje się różne kryteria dla ich opracowania. Ujęcia graficzne zaznaczające tylko pokrewieństwa oraz względny czas pojawienia się jednostek systematycznych w ewolucji nazywane są fenogramami. mogą one być złożone z nieregularnych linii (odgałęzień, ryc. IIA) lub regularnych (ryc. 1 IB) W kladogramie na osi rzędnych podawana jest ocena czasu w pojawieniu się jednostki systematycznej, na osi odciętych stopień różnicy cech. Stopień pokrewieństwa jest określany za pomocą położenia punktów odgałęzień. Ilustruje to ryc. 12. Widać, że jednostka B jest bliżej spokrewniona z jednostkami C i D niż z jednostką A. ponieważ jednostki B, C i D oddzieliły się w tym samym punkcie T2, podczas gdy jednostka A oddzieliła się wcześniej (punkt T,). Do zaznaczenia w klado-
A B C O
c
z
a
s
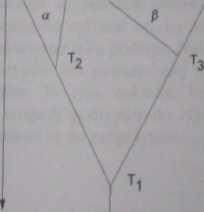
praprzodek
Ryc. 13. Filogram. Objaśnienie w tekście
51
Wyszukiwarka
Podobne podstrony:
50 51 (23) 50 URAZY l USZKODZENIA SPORTOWE gicznych wymienić należy zapalenie wielostawowe, dnę i ch
50,51 (10) _Ćwiczenia fnetrorytnuczne Podobnie jak przy Ćwiczeniach poprzcun należy wyzyskać natural
50 51 stanowić się również nad bardziej prozaicznymi, r2e, mieślniczymi błędami w sztuce. Na pewno n
50,51 3 2. Ćwiczenia metroryttniczne Podobnie jak przy ćwiczeniach poprzcdnich^B należy wyzyskać nat
50 51 (2) 50 Depolaryzację ideologiczną należy przypisywać przede wszystkim przesunięciu się na praw
CCI00137 (2) KWESTIONARIUSZ DOTYCZĄCY ZUP I. Do jakiej grupy wiekowej pan(i) należy? 13-30 _ 31-40 _
SZEFOSTWO SŁUŻBY HYDROMETEOROLOGICZNEJ SZ RP(fatamorgana). Należy pamiętać, że zasięg poziomy PR
51 3.2.2 Journal de termin Nous avons vecu exactement rexpćrience que nous voulons
Pmw W PKP Urząd Pocztowy Poczta 50 51 00-24 105-6 Pmw IS3 50 51 00-24 102-3 50 51 00-24
S 50+51 srir Vorlage siehe Seite 112ćfesckirrwotiyPps wjrrfgebnmcAt Konturenfarben ✓
artykuły W zależności od koncentracji tych związków, gaz należy oczy ścić do poziomu wymaganego
M-14.01.02 45 46 47 48 49 50 51 52 53 PN-EN ISO 1461:2000 PN-EN 20898-2:1998 PN-EN
Terminy Wiążące organ prowadzący postępowanie Wiążące stronę / uczestników
S6302490 IT. Inne Lp. 48 49 50 51 52 55 54 55 56 Bazwa Oznaczenie Hec bani za maltański Hechaaiza
str300 301 fc90d — 5,7 0,8/1,3 = 3,51 MPa, l = 1993/(50-3,51) = 11,36 mm-» przyjęto dla obu belek /j
50 51 Montaż: A. Prevlećiemc lanko s lanovodom cez prave riadidlo a kryt svet
więcej podobnych podstron