339 (22)

iego ładunku po zewnętrznej stronic i ujemnego po wewnętrznej, co z koki rosnącym siłom odpychania elektrostatycznego powoduje zmniejszenia in-aywnośct dyfuzji. Tak więc konkurują ze sobą dwa przeciwstawne bodźce: grani stężenia napędzający dyfuzję jonów K* z komórki i różnica potencjałów ciek-iznyctl hamująca ją. Potencjał elektryczny wzrasta do chwili wyrównania jego iw u z wpływem gradientu stężenia. Dla więkx/x>ści komórek glejowych war-Md potencjału ekktrycznego błony, przy którym następuje zatrzymanie dy fuzji jonem K* z komórki, wynosi -75 mV. Ponieważ błona komórkowa jest przepuszczalna tylko dla jonów K\ osiągnięty w ten sposób potencjał równowagowy jest czelnie potencjałem spoczynkowym tej komórki. W przypadku błony komór-sej przepuszczalnej tylko dla jonów K* do utrzymania gradientu stężeli nic jest czne wydatkowanie energii. Potencjał błony komórkowej automatycznie przyjmuje taką wartość. by jony K* były w sunie równowagi, czyli aby wpływ gra-i stężeń jonów K* by ł niwelowany przez wytworzoną na błonie różnicę poten-f elektrycznych.
Vkartość potencjału błonowego AVK. przy który m jony K* są w stanic równowa-. można obliczyć z równania wyprowadzonego w 1888 roku przez Nemsta (patrz ział 7.11.6):
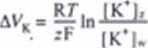
par* |K*|. 1 IK*I| - uętoue jooó» wewnątrz i m zewnątrz kootórii.
Wzór Ncrnsta może być również wykorzystany do obliczenia potencjału równowagowego dla dowolnego rodzaju jonów obecnych po obydwu stronach błony komórkowej (lab. 13.3). Porównując zmierzoną różnicę potencjałów elektrycznych w poprzek błony komórki glejowej, otrzymujemy warfo<ci zgodne z obliczonymi aa podstaw ie wzoru Ncrnsta W przypadku neuronów dla małych stężeń jonów K* m zewnątrz komórki rzeczywista wartość potencjału maleje wolniej, niż to wynika ar wzoru Nemsta. Świadczy to o tym. że błona komórkowa neuronu jest przepusz-cżalna nie tylko dla jonów K#. Błony komórek nerwowych są przepuszczalne zarówno dla jonów K\ jak i dla jonów Na* i G". Z najważniejszych jonów obecnych w komórce nerwowej tylko wymienione wcześniej aniony organiczne nic przenikają przez błonę komórkową. Powraca zatem postaw ione wcześniej pytanie, w jaki 9pcnób utrzymywane są stałe wartości gradientów stężeń poszczególnych jonów i jaki w pływ wywierają one na wartość potencjału spoczynkowego błony komórko* •cj
Analizując wartości zawarte w tabeli 13.3, można zauważyć, że potencjał spoczynkowy jest bardzo bliski wartości potencjału równowagowego dla jonów potasu obliczonemu na podstawie wzoru Nemsta. Jeżeli błona komórkowa oprócz kanałów dla jonów K* będzie posiadać otwarte kanały dla jonów Na*, to dzięki istniejącemu gradientowi stężeń będą one próbować dy fundować do w nętrza komórki Lw/ględniając obliczoną wartość potencjału równowagowego dla jonów sodu (AV'Sł) +55 mV oraz przyjmując, żc potencjał spoczynkowy błony w ty m momen-
339
Wyszukiwarka
Podobne podstrony:
img053 (42) zakończona u wierzchu guzikiem płaskim. Po zewnętrznej stronic kolanka są ślady odłamane
TRENING NA BRYCZESY zcodzienniefit.pl BRYCZESY TŁUSZCZ PO ZEWNĘTRZNEJ STRONIE UD ORAZ
Dokument (28) W PARKU ROSNĄ DĘBY UMYJ DOKŁADNIE ZĘBY. Dziecko przesuwa język po: -zewnętrznej stroni
82773 JAK25,2 3. Wszystkie części należy wycinać dokładnie po zewnętrznej stronie
Przeszkody: sztywna ściana komórkowa, ujemny potencjał po zewnętrznej stronie błony komórkowej. Fuzj
klstidwa054 102 l. MOSZYŃSKI: KULTURA LUDOWA SŁOWIAN dym razie i tam rachowano po zewnętrznej stroni
img053 (42) zakończona u wierzchu guzikiem płaskim. Po zewnętrznej stronic kolanka są ślady odłamane
img053 (42) zakończona u wierzchu guzikiem płaskim. Po zewnętrznej stronic kolanka są ślady odłamane
Bez nazwy? Rys. 16.4. Rozkłady naprężeń po zewnętrznej stronie ścianki zbiornika Tablica 16.1. Warto
img012 (65) we w. Ryc. 5.22. Pompa Ha /K - ATPazowa. Sod i ATP zbliżają się do kompleksu ATPazy po w
img012 (65) we w. Ryc. 5.22. Pompa Ha /K - ATPazowa. Sod i ATP zbliżają się do kompleksu ATPazy po w
więcej podobnych podstron