107903
ponownie przez sekwencję start-transfer itp. Po odcięciu peptydu sygnałowego białko pozostaje w błonie.
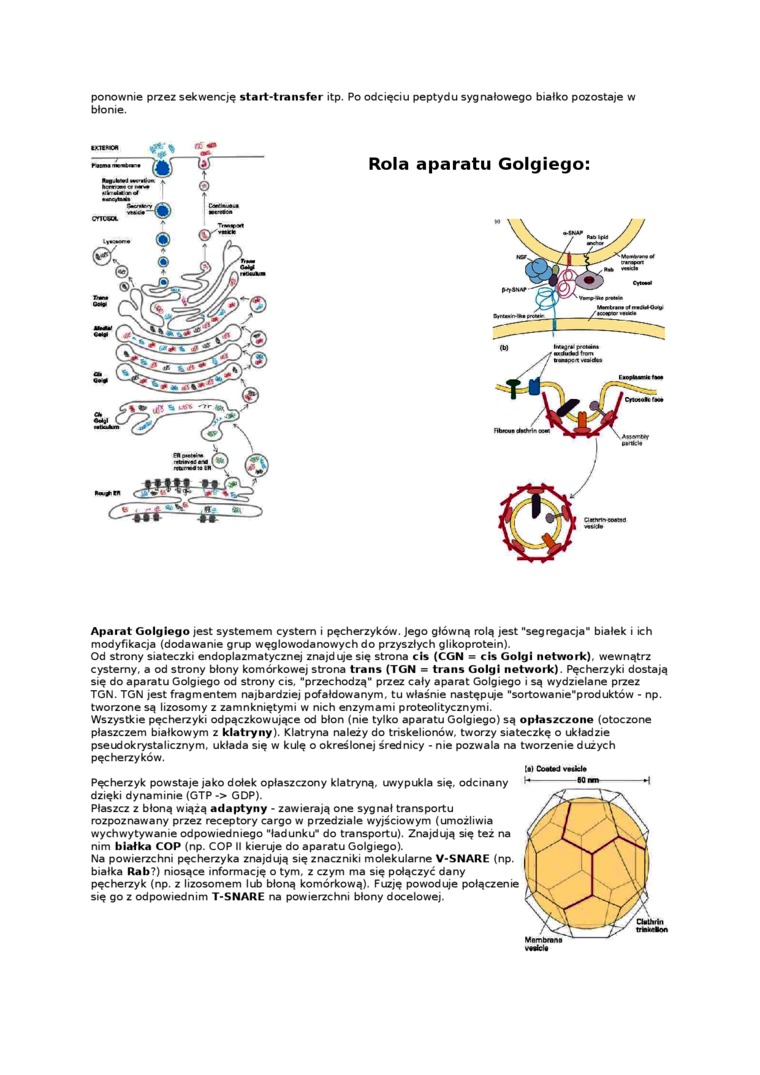
Aparat Golgiego jest systemem cystern i pęcherzyków. Jego głównę rolę jest “segregacja" białek i ich modyfikacja (dodawanie grup węglowodanowych do przyszłych glikoprotein).
Od strony siateczki endoplazmatycznej znajduje się strona cis (CGN = cis Golgi network). wewnętrz cysterny, a od strony błony komórkowej strona trans (TGN = trans Golgi network). Pęcherzyki dostaję się do aparatu Golgiego od strony cis. “przechodzę" przez cały aparat Golgiego i sę wydzielane przez TGN. TGN jest fragmentem najbardziej pofałdowanym, tu właśnie następuje “sortowame"produktów - np. tworzone sę lizosomy z zamokniętymi w nich enzymami proteolitycznymi.
Wszystkie pęcherzyki odpęczkowujęce od błon (nie tylko aparatu Golgiego) sę opłaszczone (otoczone płaszczem białkowym z klatryny). Klatryna należy do triskelionów, tworzy siateczkę o układzie pseudokrystalicznym, układa się w kulę o określonej średnicy - nie pozwala na tworzenie dużych pęcherzyków.
[•) Co«(«ł vMkł«
h-Mm-H
Mwnbrant
Pęcherzyk powstaje jako dołek opłaszczony klatrynę, uwypukla się, odcinany dzięki dynaminie (GTP -> GDP).
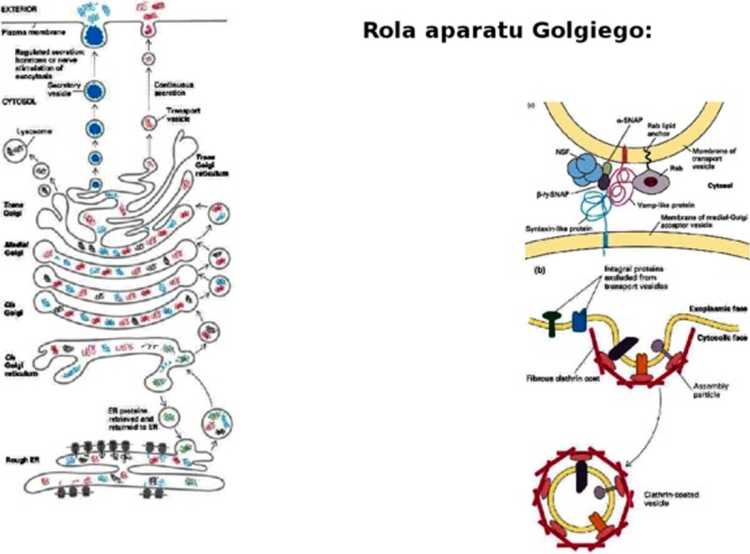
Płaszcz z błonę więżę adaptyny - zawieraję one sygnał transportu rozpoznawany przez receptory cargo w przedziale wyjściowym (umożliwia wychwytywanie odpowiedniego “ładunku* do transportu). Znajduję się też na nim białka COP (np. COP II kieruje do aparatu Golgiego).
Na powierzchni pęcherzyka znajduję się znaczniki molekularne V-SNARE (np. białka Rab?) niosęce informację o tym, z czym ma się połęczyć dany pęcherzyk (np. z lizosomem lub błonę komórkowę). Fuzję powoduje połęczenie się go z odpowiednim T-SNARE na powierzchni błony docelowej.
Wyszukiwarka
Podobne podstrony:
f9 TRANSFORMATA OBRAZU ORYGINALNEGO TRANSFORMATA OBRAZU PO ZMNIEJSZENIU ROZDZIELCZOŚCI
PrepOrg II118 (2) 121 - olejowej przez 30 minut w temperaturze 200° i po stopieniu (uwaga 2) wlewa s
PICT0107 ->U określane przez scałkowanie gęstości strumienia cieplnego po powierzchni komponentu
ScannedImage 45 stację w tej pamiętnej podróży do nieba, stała się przez to trzecim miastem kultu po
IMG 63 (2) ł. WIADOMOŚCI OGÓLNE uzwojeń transformatora, który po wyłączeniu jednego‘uzwojenia może n
IMGw87 129 dwa kroki na wprost, krok w lewo, krok na wprost, pół obrotu w prawo itp. Po odejściu do
page0585 577Śledź przez kości między-szczękowe, zaś po bokach przez kości szczęki górnej wątłe, płas
AGHPrzekrój przez koryto rzeki a) w stanie naturalnym b) po wybudowaniu zapory.
Jeżeli licznik przechodzi przez wszystkie stany cyklicznie (tzn. po przejściu N stanów cykl jest pow
1Cc Przez B i S prowadzimy prostą i odkładamy długość B S po drugiej stronie, z czego wychod
więcej podobnych podstron