
METABOLIZM LIPIDÓW
cz. I
dr Aleksandra Król
Zakład Biochemii Medycznej

Lipidy
– heterogenna grupa związków, pokrewnych kwasom tłuszczowym,
cechuje je: względna nierozpuszczalność w wodzie oraz rozpuszczalność w
rozpuszczalnikach niepolarnych, takich jak eter, chloroform i benzen.
Znaczenie biomedyczne lipidów:
• są wydajnym źródłem energii
• stanowią izolator termiczny
• utrzymują narządy w stałym położeniu
• są składnikiem budulcowym błon biologicznych
• biorą udział w przekaźnictwie sygnałów (hormony steroidowe, wtórne
przekaźniki– diacyloglicerol, fosfatydyloinozytol)
• biorą udział w zjawiskach immunologicznych (eikozanoidy)
• są substratem w biosyntezie niektórych witamin
• połączenia tłuszczów z białkami (lipoproteiny) służą jako środek transportu
lipidów w osoczu
• karotenoid – retinol, wrażliwy na światło, odgrywa główną rolę w procesie
widzenia
LIPIDY

Lipidy proste – estry kwasów tłuszczowych z różnymi
alkoholami:
•
tłuszcze właściwe
(glicerydy) – estry kwasów tłuszczowych z
glicerolem
•
woski
- estry kwasów tłuszczowych z długołańcuchowymi
alkoholami monohydroksylowymi
Lipidy złożone - estry kwasów tłuszczowych, zawierające
dodatkowe grupy funkcyjne:
•
fosfolipidy
– zawierają resztę kwasu fosforowego
glicerofosfolipidy – alkoholem jest glicerol
sfingofosfolipidy – alkoholem jest sfingozyna
•
glikolipidy
– zawierają węglowodany i sfingozynę
cerebrozydy
gangliozydy
•
inne lipidy złożone
– sulfolipidy, aminolipidy, lipoproteiny, lipidy
izoprenoidowe (sterydy i karotenoidy)
PODZIAŁ LIPIDÓW

Kwasy tłuszczowe (KT)
- są łańcuchami węglowodorowymi o różnej
długości i stopniu nasycenia, zakończonymi grupami karboksylowymi.
•
Posiadają zwykle parzystą liczbę atomów węgla (14-24), powstają z
jednostek dwuwęglowych.
•
Cząsteczka KT ma charakter amfipatyczny – posiada fragment
hydrofobowy (łańcuch węglowodorowy) i hydrofilny (grupa
karboksylowa).
• Łańcuch węglowodorowy może być nasycony lub nienasycony (kwasy
jednonienasycone – monoenowe, wielonienasycone – polienowe).
•
W miarę wzrostu liczny atomów węgla w cząsteczce rośnie
temperatura topnienia; obecność i wzrost liczby wiązań podwójnych
obniża temperaturę topnienia. KT są tym bardziej płynne im krótsze
mają łańcuchy i im są bardziej nienasycone.
•
Niemal wszystkie KT mają łańcuchy proste (wyjątek: kwas fitonowy).
•
Nienasycone kwasy tłuszczowe (NKT) mogą występować w dwóch
formach stereoizomerycznych: cis i trans.
•
Wszystkie podwójne wiązania w naturalnie występujących kwasach
tłuszczowych są w konfiguracji
cis
cis
.
.
KWASY TŁUSZCZOWE
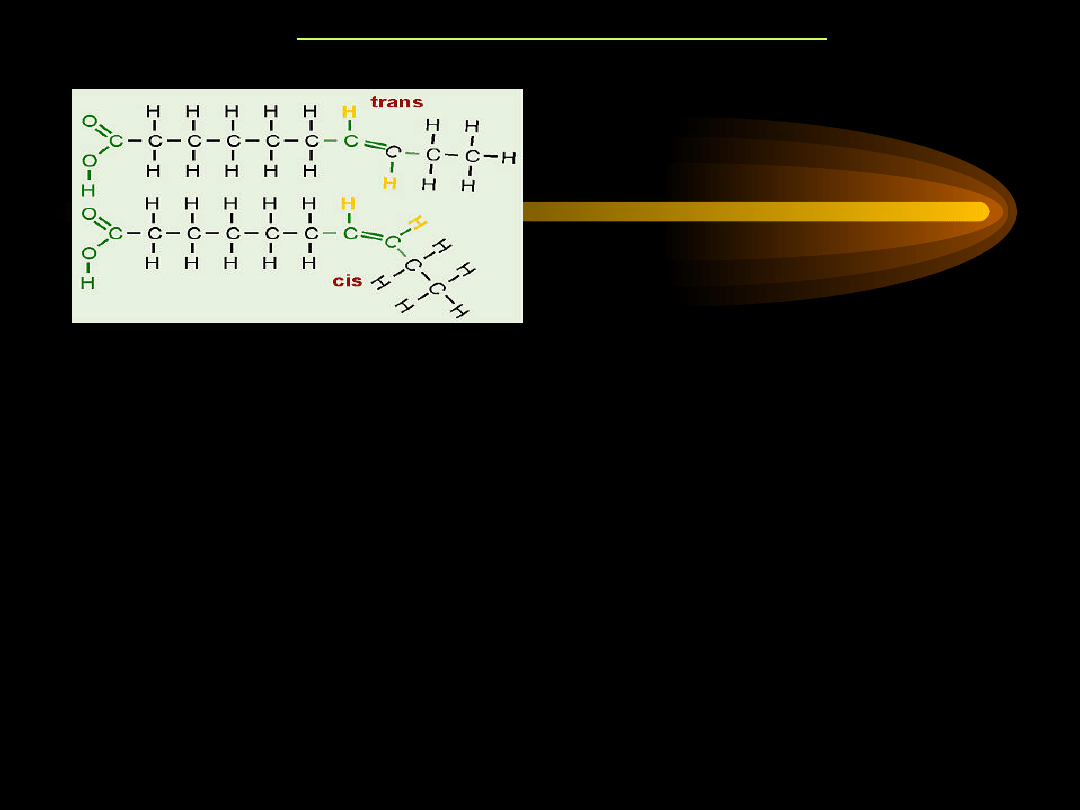
W naturalnych tłuszczach zwierzęcych
izomery trans występują w ok. 3-5% w
stosunku do izomerów cis. Izomery trans
powstają w wyniku przemysłowej obróbki
tłuszczu. Występują one w wielu
półsyntetycznie otrzymywanych stałych
tłuszczach roślinnych - takich jak margaryna i
masło roślinne, które są otrzymywane w
wyniku częściowego uwodorniania
naturalnych tłuszczów roślinnych.
IZOMERY CIS I TRANS NKT
Zmiana konfiguracji z cis na trans powoduje, że zmienia się kształt
cząsteczki. Reszty kwasów tłuszczowych o konfiguracji trans mają
kształt liniowy, zaś o konfiguracji cis mają kształt litery "V", co
radykalnie wpływa na ich metabolizm.
Tłuszcze zawierające postać trans kwasów tłuszczowych są szkodliwe
dla zdrowia, doprowadzają do miażdżycy tętnic, należy ograniczyć ich
spożycie.
Izomery trans wpływają niekorzystnie na profil lipidowy człowieka -
powodują spadek poziomu „dobrego cholesterolu” HDL i podnoszą
poziom tzw. „złego cholesterolu” LDL.

Numeracja atomów węgla w kwasach tłuszczowych:
KWASY TŁUSZCZOWE
H
3
C – (CH
2
)
n
– CH
2
– CH
2
– C
O
H
O
ω
3
2
1
β
α
—
— —
Cząsteczka kwasu tłuszczowego ma charakter amfipatyczny –
posiada zarówno fragment hydrofobowy (łańcuch węglowodorowy),
jak i hydrofilowy (grupa karboksylowa zdolna do dysocjacji).

Nazwa
zwyczajow
a kwasu
Nazwa
systematycz
na kwasu
Symbol
kwasu
Wzór
chemiczny
kwasu
Temperatur
a topnienia
Masłowy
Butanowy
C 4:0
CH
3
(CH
2
)
2
CO
OH
- 7,9
Laurynowy
Dodekanowy
C 12:0
CH
3
(CH
2
)
10
COO
H
44,2
Mirystyno
wy
Tetradekano
wy
C 14:0
CH
3
(CH
2
)
12
COO
H
53,9
Palmityno
wy
Heksadekano
wy
C 16:0
CH
3
(CH
2
)
14
COO
H
63,0
Stearynow
y
Oktadekanow
y
C 18:0
CH
3
(CH
2
)
16
COO
H
69,6
Arachidow
y
Eikosanowy
C 20:0
CH
3
(CH
2
)
18
COO
H
75,3
Behenowy
Doeikosanow
y
C 22:0
CH
3
(CH
2
)
20
COO
H
79,9
NASYCONE KWASY TŁUSZCZOWE

Ważniejsze nienasycone kwasy tłuszczowe to:
•
kwasy jednonienasycone (monoenowe):
• kwasy dwunienasycone (dienowe):
• kwasy trójnienasycone (trienowe):
•
kwasy czteronienasycone (tetraenowe):
•
kwasy pięcionienasycone (pentaenowe):
NIENASYCONE KWASY TŁUSZCZOWE
kw
a
s
p
a
lm
it
o
o
le
in
o
w
y
1
6
C
(
ω
-
7
)
kw
a
s
o
le
in
o
w
y
1
8
C
(
ω
-9
)
kw
a
s
lin
o
lo
w
y
1
8
C
(
ω
-6
)
kw
a
s
α
-l
in
o
le
n
o
w
y
1
8
C
(
ω
-
3
)
kw
a
s
γ
-l
in
o
le
n
o
w
y
1
8
C
(
ω
-
6
)
k
w
a
s
a
ra
ch
id
o
n
o
w
y
2
0
C
(
ω
-6
)
kw
a
s
ik
o
za
p
e
n
ta
e
n
o
w
y
2
0
C
(ω
-3
)

N
ie
n
a
sy
co
n
e
k
w
a
sy
tł
u
sz
cz
o
w
e
d
zi
e
lim
y
n
a
g
ru
p
y
n
ie
w
ie
le
ró
żn
ią
ce
si
ę
w
ła
śc
iw
o
śc
ia
m
i
b
io
lo
g
ic
zn
y
m
i.
P
o
d
zi
a
ł
za
le
ży
o
d
b
u
d
o
w
y
c
h
e
m
ic
zn
e
j
–m
ie
js
ca
w
y
st
ą
p
ie
n
ia
p
ie
rw
sz
e
g
o
w
ią
za
n
ia
p
o
d
w
ó
jn
e
g
o
.
W
y
ró
żn
ia
m
y
k
w
a
sy
z
g
ru
p
y:
•
O
m
e
g
a
3
–
w
ią
za
n
ie
p
o
d
w
ó
jn
e
n
a
m
ie
js
cu
3
,
•
O
m
e
g
a
6
–
w
ią
za
n
ie
p
o
d
w
ó
jn
e
n
a
m
ie
js
cu
6
,
•
O
m
e
g
a
9
–
w
ią
za
n
ie
p
o
d
w
ó
jn
e
n
a
m
ie
js
cu
9
.
NIENASYCONE KWASY TŁUSZCZOWE
Kwasy omega 6 odznaczają się wyższą aktywnością biologiczną.

Niezbędne nienasycone kwasy tłuszczowe (NNKT)
–
egzogenne kwasy tłuszczowe, niezbędny element diety
człowieka, gdyż nie są syntetyzowane przez organizm,
muszą być przyjmowane z pożywieniem. Są potrzebne do
tworzenia ważnych związków (np.prostaglandyn).
NNKT z rodziny omega-3:
kwas α-linolenowy -
źródłem są tłoczone na zimno oleje: lniany i
rzepakowy, nasiona lnu i rzepaku, siemię lniane, orzechy
włoskie, kiełki pszenicy.
kwas dokozaheksaenowy (DHA)
kwas eikozapentaenowy (EPA)
–
oba kwasy nasz organizm może
wytworzyć z kwasu α-linolenowego. Źródłem DHA i EPA są ryby
morskie, tj. makrela, łosoś, halibut, dorsz, śledź, sardynka.
NNKT z rodziny omega-6:
kwas linolowy
–
żródłem są
tłoczone na zimno oleje: sojowy i
kukurydziany, nasiona słonecznika, dyni, sezamu i większość
orzechów.
kwas γ-linolenowy
kwas arachidonowy
– oba kwasy organizm może wytwarzać z
kwasu linolowego.
NIEZBĘDNE NIENASYCONE KWASY TŁUSZCZOWE

Niezbędne
nienasycone
kwasy
tłuszczowe
(niezbędne
dla
prawidłowego rozwoju młodych organizmów i zachowania zdrowia u
osób dorosłych):
•
są składnikiem budulcowym wszystkich
błon komórkowych
;
•
są prekursorem w syntezie
eikozanoidów
(kwas arachidonowy), do których
należą:
prostaglandyny
(hormony tkankowe o szerokim działaniu fizjologicznym);
tromboksany
(syntetyzowane w płytkach krwi, powodują skurcz naczyń i
agregację płytek);
leukotrieny
(mediatory prozapalne);
lipoksyny
(silne
właściwości przeciwzapalne - efekt antagonistyczny do leukotrienów);
•
biorą udział w
metabolizmie cholesterolu
i jego transporcie - ponad połowa
estrów cholesterolu występuje w postaci połączeń z kwasem linolowym, co
ułatwia ich rozprowadzanie w organizmie, obniżając poziom cholesterolu we
krwi;
•
biorą udział w
transporcie wody i elektrolitów
przez błony komórkowe;
•
regulują
wydalanie jonów sodu
z organizmu.
Największą wartością i aktywnością biologiczną odznaczają się kwasy
tłuszczowe należące do rodziny omega-3. Prawidłowy stosunek kwasów
tłuszczowych z rodziny omega-6 do kwasów z rodziny omega-3 powinien
wynosić (<5:1).
ROLA NIENASYCONYCH KWASÓW TŁUSZCZOWYCH
Większość lipidów spożywamy w postaci triacylogliceroli, które muszą
być rozłożone do kwasów tłuszczowych, aby mogły być wchłonięte
przez nabłonek jelita. Jelitowe enzymy zwane lipazami rozkładają
trójglicerydy do wolnych kwasów tłuszczowych i
monoacyloglicerolu.
• Proces trawienia tłuszczów rozpoczyna się w
jamie ustnej
. Gruczoły
Ebnera, położone na grzbiecie języka, wytwarzają
lipazę językową
.
•
Trawienie kontynuowane jest w żołądku.
Żołądek
wydziela
lipazę
żołądkową
, aktywną w obojętnym pH. Tu działa też
lipaza ślinowa
.
Obie lipazy hydrolizują
triacyloglicerole
zawierajace krótko- (lipaza
ślinowa)
i
długołańcuchowe
kwasy
tłuszczowe
do
1,2-
diacyloglicerolu
i
wolnych kwasów tłuszczowych
. Są szczególnie
potrzebne w okresie noworodkowym, w którym aktywność lipazy
trzustkowej jest mała.
TRAWIENIE LIPIDÓW
a
c
y
l
a
c
y
l
a
c
y
l
a
c
y
l
a
c
y
l
O
H
li
p
a
z
a
H
2
O
K
T
tr
ia
c
y
lo
g
li
c
e
r
o
l
1
,2
-
d
ia
c
y
lo
g
li
c
e
ro
l

Proces trawienia kontynuowany jest w
jelitach
pod wpływem
lipazy
i
fosfolipazy
; enzymy te działają jedynie na powierzchni tłuszczu. W celu
zwiększenia powierzchni tłuszczu dostępnej dla enzymu, woreczek
żółciowy wydziela
żółć
.
Sole kwasów żółciowych
– biologiczne
detergenty, zmniejszają napięcie powierzchniowe kulek tłuszczu, co
prowadzi do ich emulsyfikacji (duże kulki rozpadają się na mniejsze,
wzrasta powierzchnia kontaktu lipidu z enzymem).
Żółć:
•
wytwarzana w hepatocytach wątroby w sposób ciągły
•
magazynowana w pęcherzyku żółciowym
•
emulguje tłuszcze
•
zobojętnia miazgę żołądkową
(pH bardziej zasadowe)
•
z żółcią wydalane są: cholesterol, kwasy żółciowe, leki, toksyny, miedź,
rtęć, cynk, barwniki żółciowe.
Jeśli produkcja soli żółciowych jest niewystarczająca z powodu
choroby wątroby, spore ilości tłuszczu są wydalane z kałem. Taki
stan to biegunka tłuszczowa.
TRAWIENIE LIPIDÓW

Sok trzustkowy
zawiera liczne enzymy:
•
lipazy trzustki
(rozbijają triacyloglicerole na glicerol i
kwasy tłuszczowe)
•
esterazę cholesterolową
(hydrolizuje estry
cholesterolu do cholesterolu i kwasów tłuszczowych)
•
fosfolipazę A
2
(hydrolizuje glicerofosfolipidy do
lizofosfolipidów)
Sok jelitowy
wydzielany przez gruczoły dwunastnicze i
jelitowe zawiera:
•
fosfolipazę soku jelitowego
(rozkłada fosfolipidy do
glicerolu, kwasów tłuszczowych, kwasu fosforowego i
zasady, takiej jak cholina)
TRAWIENIE
LIPIDÓW
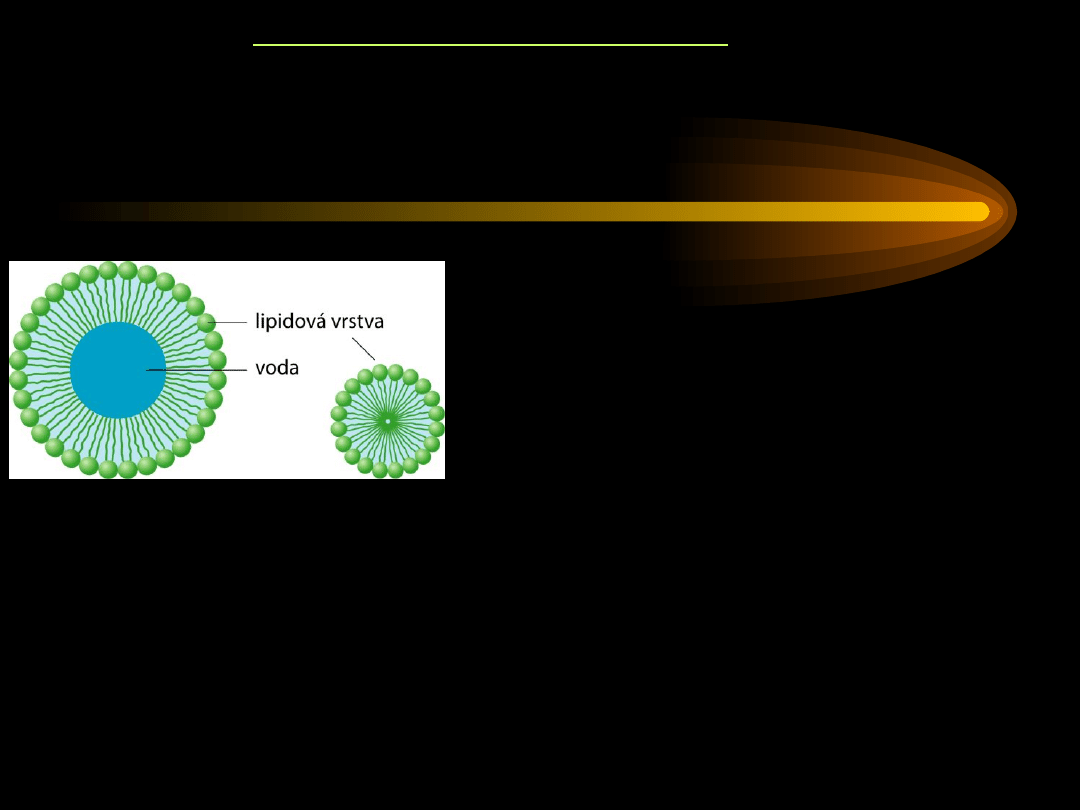
Wchłanianie odbywa się w jelicie cienkim.
Produkty trawienia lipidów wchłaniane są z
miceli
utworzonych
przez sole kwasów żółciowych (cząsteczki lipidów otoczone przez
cząsteczki kwasów żółciowych).
micela
W micelach znajdują się: 1-monoacyloglicerole,
2-monoacyloglicerole, kwasy tłuszczowe,
cholesterol, fosfatydylocholina, sole kwasów
żółciowych.
Z rozpuszczalnymi w wodzie micelami, produkty trawienia lipidów dostają się
do wnętrza
enterocytów
– komórek jelita cienkiego, które biorą udział we
wchłanianiu z jelita cienkiego do krwi.
W warunkach fizjologicznych ponad 98% lipidów zawartych w pokarmach jest
wchłanianych w jelitach.
WCHŁANIANIE LIPIDÓW

• Sole kwasów żółciowych
– są wchłaniane w jelicie i przechodzą
do krążenia jelitowo-wątrobowego.
• Fosfolipidy
– są rozkładane przez trzustkową fosfolipazę A2 do
kwasów tłuszczowych i lizofosfolipidów, które są resorbowane
przez nabłonek jelita.
• Estry cholesterolu
– są hydrolizowane przez trzustkową
hydrolazę estrów cholesterolu, uwolniony cholesterol przedostaje
się do rąbka szczoteczkowego.
• 1-monoacyloglicerole
– są hydrolizowane przez lipazę do
glicerolu i kwasów tłuszczowych.
• 2-monoacyloglicerole
– ulegają przemianie do triacylogliceroli
• Długołańcuchowe kwasy tłuszczowe
mogą być użyte do
resyntezy triacylogliceroli.
• Wolny glicerol
– ulega wchłonięciu do krążenia wrotnego lub
wykorzystany jest do resyntezy triacylogliceroli.
W komórkach jelitowych zachodzi
resynteza lipidów
, które po
połączeniu z białkiem, są wydzielane w formie
lipoprotein
(chylomikronów) do układu chłonnego i krwiobiegu.
WCHŁANIANIE LIPIDÓW

Tłuszcze wchłonięte z pokarmów oraz syntetyzowane w wątrobie
i tkance tłuszczowej muszą być transportowane między
różnymi tkankami i narządami, aby mogły być zużywane i
magazynowane.
Ze względu na hydrofobowy charakter lipidów mogą one być
transportowane w środowisku wodnym, jakim jest krew, dzięki
tworzeniu rozpuszczalnych kompleksów z białkami.
Długołańcuchowe kwasy tłuszczowe wiążą się z albuminami,
krótkołańcuchowe rozpuszczają się w osoczu, natomiast inne
lipidy transportowane są w
kompleksach lipoproteinowych
,
które w osoczu krwi występują w kilku rodzajach różniących się
wielkością i składem.
LIPOPROTEINY
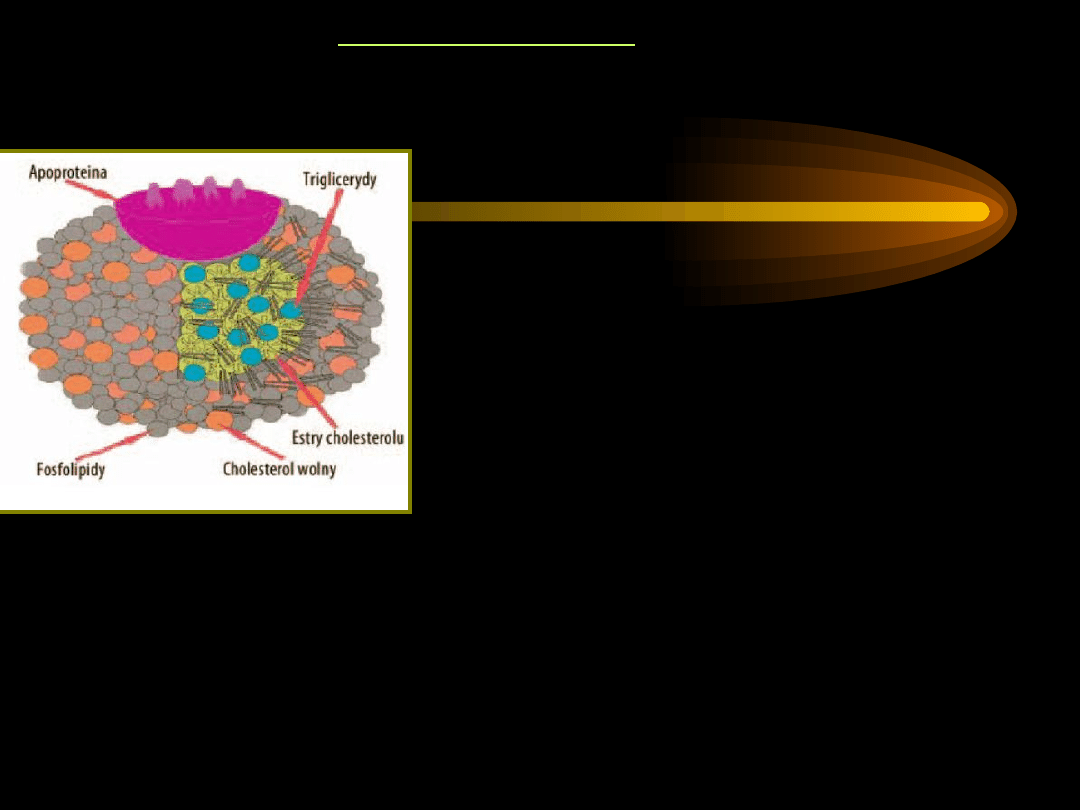
LIPOPROTEINY
Lipoproteiny
mają budowę sferyczną.
Wnętrze to
silnie hydrofobowy rdzeń złożony z
niepolarnych
triacylogliceroli i estrów cholesterolu. Rdzeń
otoczony jest hydrofilną powłoką z lipidów,
posiadających grupy polarne oraz białek
zwanych
apoproteinami
.
Apoproteiny zapewniają lipoproteinom
rozpusz -
czalność.
Apoproteiny mogą być ściśle związane z
częścią
lipidową, nie można ich wtedy usunąć, lub
mogą
być swobodnie przenoszone do innych
lipoprotein.
Ultrawirowanie osocza rozdziela lipoproteiny na cztery frakcje o różnej gęstości:
• chylomikrony (CM)
• lipoproteiny o bardzo niskiej gęstości (VLDL)
• lipoproteiny o niskiej gęstości (LDL)
• lipoproteiny o wysokiej gęstości (HDL)
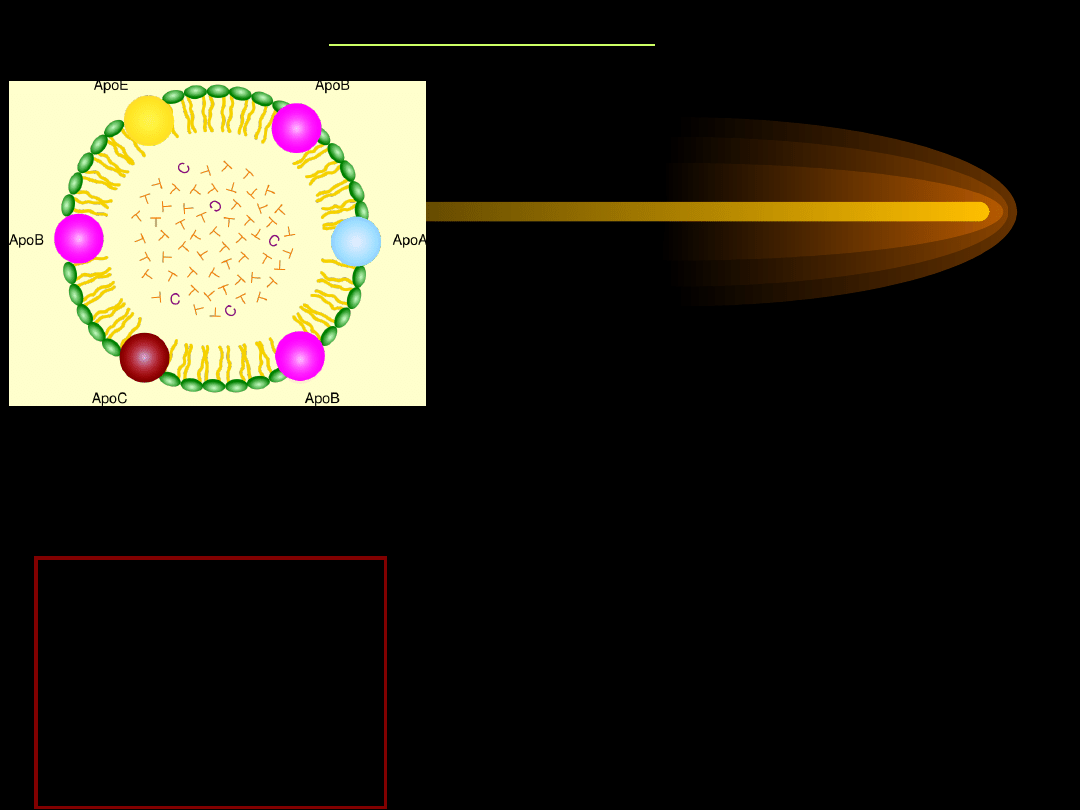
CHYLOMIKRONY
fosfolipidy
T
–
tr
ia
c
y
lo
g
li
c
e
ro
l
C
–
c
h
o
le
s
te
ro
l
S
k
ła
d
c
h
y
lo
m
ik
ro
n
ó
w
:
•
b
ia
łk
a
2
%
•
fo
s
fo
li
p
id
y
3
-8
%
•
e
s
tr
y
c
h
o
le
s
te
ro
lu
1
-
3
%
•
c
h
o
le
s
te
ro
l
1
-2
%
•
tr
ó
jg
li
c
e
ry
d
y
8
5
-9
4
%
•
C
h
y
lo
m
ik
ro
n
y
t
o
l
ip
o
p
ro
te
in
y
z
b
u
d
o
w
a
n
e
z
rd
ze
n
ia
tr
ig
lic
e
ry
d
o
w
e
g
o
(o
k.
9
0
%
)
o
ra
z
o
to
cz
k
i
zł
o
żo
n
e
j
z
fo
sf
o
lip
id
ó
w
,
n
ie
w
ie
lk
ie
j
ilo
śc
i
ch
o
le
st
e
ro
lu
i
a
p
o
lip
o
p
ro
te
in
y
(
a
p
o
B
-4
8
,
a
p
o
A
,
a
p
o
C
II
I
o
ra
z
a
p
o
E
).
•
S
ą
lip
o
p
ro
te
in
a
m
i
n
a
jn
iż
sz
e
j
g
ę
st
o
śc
i
i
n
a
jw
ię
k
sz
e
j
śr
e
d
n
ic
y
,
za
w
ie
ra
ją
n
a
jw
ię
ce
j
lip
id
ó
w
i
n
a
jm
n
ie
j
a
p
o
p
ro
te
in
.
•
Po
w
st
a
ją
w
ś
ci
a
n
ie
j
e
lit
a
c
ie
n
ki
e
g
o
.
•
S
ą
t
ra
n
sp
o
rt
o
w
a
n
e
p
rz
e
z
lim
fę
d
o
k
rw
i,
a
z
n
ią
d
o
t
ka
n
k
i
tł
u
sz
cz
o
w
e
j
i
m
ię
śn
i.
•
P
rz
e
n
o
sz
ą
l
ip
id
y
p
o
ch
o
d
ze
n
ia
p
o
ka
rm
o
w
e
g
o
:
tr
ig
lic
e
ry
d
y
,
ch
o
le
st
e
ro
l
i
je
g
o
e
st
ry
z
j
e
lit
a
d
o
tk
a
n
e
k
o
b
w
o
d
o
w
y
ch
.
•
N
a
p
o
w
ie
rz
ch
n
i
śr
ó
d
b
ło
n
ka
n
a
cz
y
ń
w
ło
so
w
a
ty
ch
t
ka
n
e
k
p
o
za
w
ą
tr
o
b
o
w
y
ch
(
m
ię
śn
i
i
tk
a
n
ki
tł
u
sz
cz
o
w
e
j)
,
lip
a
za
lip
o
p
ro
te
in
o
w
a
h
y
d
ro
liz
u
je
w
ię
k
sz
o
ść
t
ri
a
cy
lo
g
lic
e
ro
li.
•
U
w
o
ln
io
n
e
kw
a
sy
tł
u
sz
cz
o
w
e
i
g
lic
e
ro
l
w
ch
ła
n
ia
n
e
są
d
o
ko
m
ó
re
k
,
a
ch
y
lo
m
ik
ro
n
y
p
rz
e
ch
o
d
zą
st
o
p
n
io
w
o
w
ch
y
lo
m
ik
ro
n
y
re
sz
tk
o
w
e
–
r
e
m
n
a
n
ty
,
u
su
w
a
n
e
z
k
rw
i
p
rz
e
z
w
ą
tr
o
b
ę
.

• VLDL są produkowane przez wątrobę. Zawierają
triacyloglicerole pochodzenia endogennego. Przenoszą
lipidy z wątroby do tkanek peryferyjnych. Wydzielone do
krwi wymieniają apoproteiny i lipidy z innymi
lipoproteinami. Lipidy w nich zawarte są trawione przez
lipazę lipoproteinową, co powoduje zmniejszenie ich
średnicy i zwiększenie gęstości, przez co następuje
przemiana VLDL do LDL.
•
LDL transportują cholesterol z wątroby do innych
narządów: nerek, mięśni, kory nadnerczy. Zawierają
większość cholesterolu osoczowego. Odkładają wolny
cholesterol na powierzchni błon komórkowych.
•
HDL syntetyzowane są w wątrobie i ścianie jelita. Pełnią
rolę czynnika oczyszczającego osocze z cholesterolu.
Głównymi lipidami HDL są fosfolipidy.
POZOSTAŁE LIPOPROTEINY OSOCZA
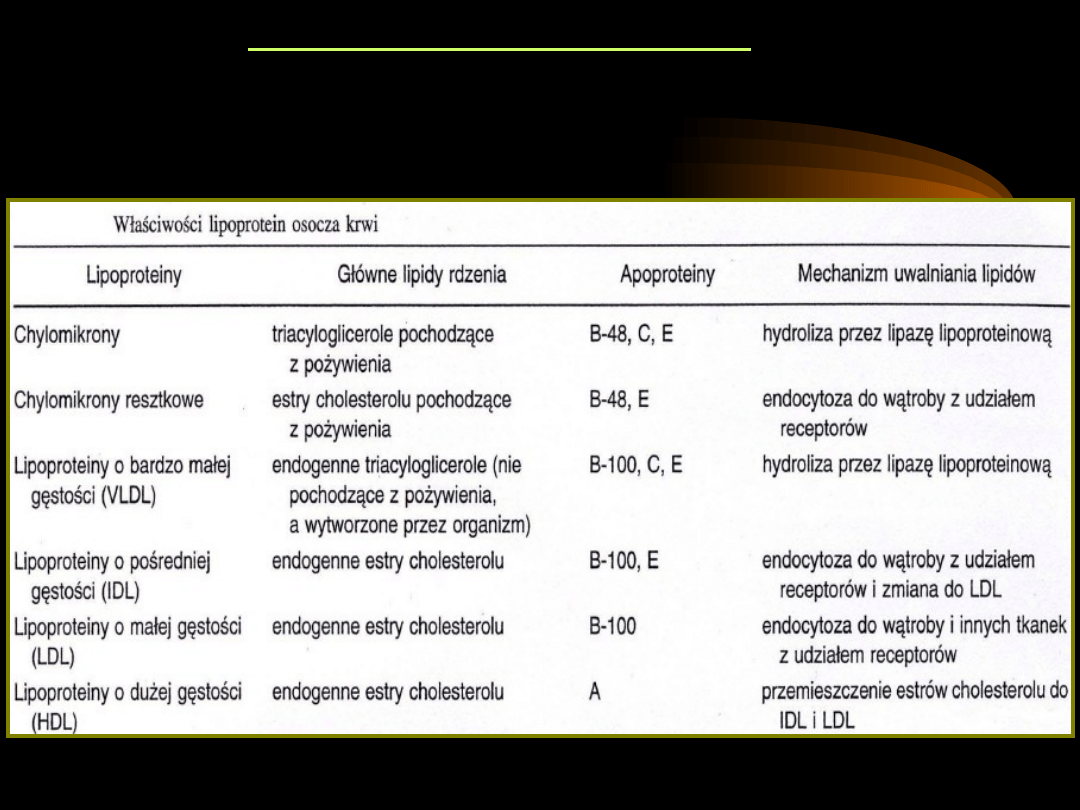
LIPOPROTEINY OSOSCZA

Prawidłowe wartości stężenia lipidów w osoczu:
• TC (cholesterol całkowity) < 200 mg/dl
• LDL < 135 mg/dl
• HDL > 45 mg/dl dla kobiet i > 40 mg/dl dla mężczyzn
• TG (trójglicerydy) < 200 mg/dl
Hiperlipidemie:
• hipercholesterolemia – TC ≥ 200 mg/dl i TG < 200 mg/dl
• hipertrójglicerydemia – TG ≥ 200 mg/dl i TC < 200
mg/dl
• hiperlipidemia mieszana – TC ≥ 200 mg/dl i TG ≥ 200
mg/dl
LIPIDOGRAM

Dorosły człowiek pobiera w ciągu doby 60g-150g lipidów, ponad 90% z nich to
triacyloglicerole
. Pozostałe to
cholesterol i jego estry
,
fosfolipidy
i
wolne kwasy tłuszczowe.
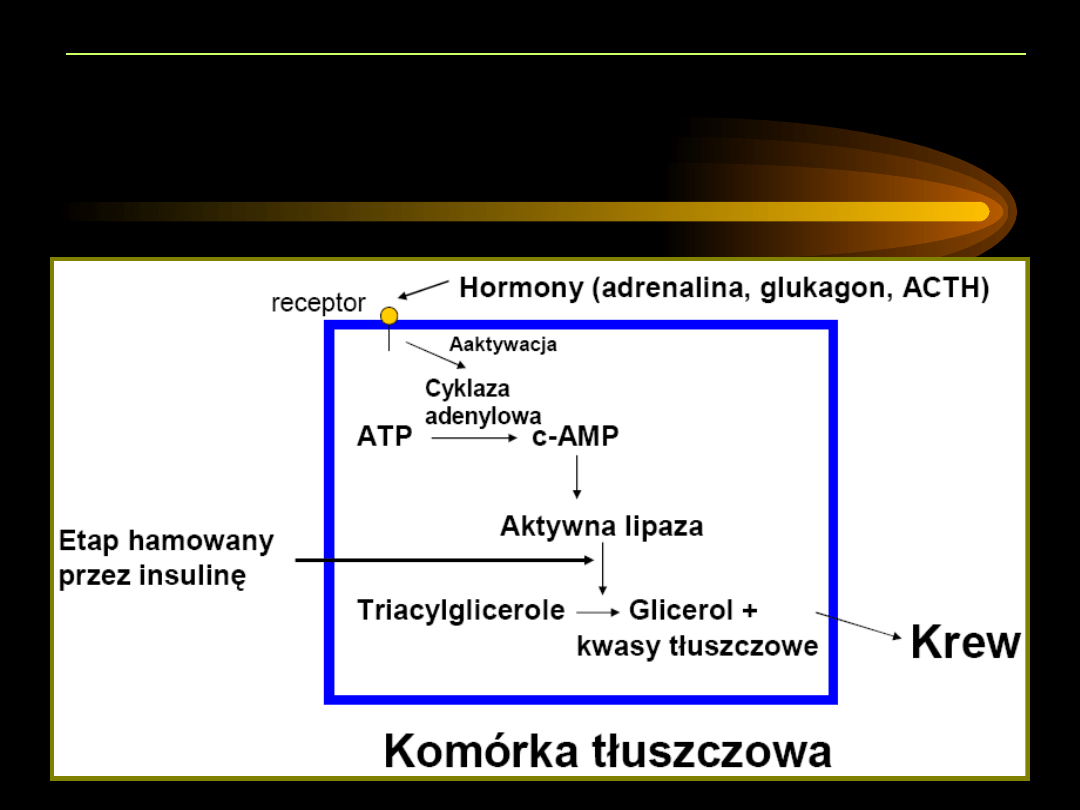
Lipoliza
to rozkład hydrolityczny triacylogliceroli i innych estrów kwasów
tłuszczowych w tkance tłuszczowej, prowadzący do powstania kwasów
tłuszczowych i glicerolu, które uwolnione do krwioobiegu wychwytywane są
przez większość tkanek (poza mózgiem i erytrocytami).
•
Katalizowana jest w
adipocytach
przez enzym:
lipazę hormonowrażliwą
.
•
Lipaza regulowana jest na drodze interkonwersji przez
adrenalinę
,
noradrenalinę
,
glukagon
i
adrenokortykotropinę
(
indukują lipolizę
)
oraz
insulinę
(
hamuje lipolizę
).
•
Uwolnione kwasy tłuszczowe przenikają przez błonę komórkową adipocytów i
docierają do krwi; nie są rozpuszczalne w osoczu, wiążą się z albuminą
osoczową (przenośnik).
•
Po utlenieniu stają się dostępnym
źródłem energii
dla tkanek obwodowych.
•
Erytrocyty i komórki nerwowe
nie mogą zużywać kwasów tłuszczowych
do celów energetycznych.
LIPOLIZA WEWNĄTRZKOMÓRKOWA
R
1
R
2
R
3
w
ią
za
n
ie
e
s
tr
o
w
e
T
R
IA
C
Y
L
O
G
L
IC
E
R
O
L
G
L
IC
E
R
O
L
li
p
a
z
y
+
3
H
2
O
+
K
W
A
S
Y
T
Ł
U
S
Z
C
Z
O
W
E
LIPOLIZA
REGULACJA LIPOLIZY WEWNĄTRZKOMÓRKOWEJ
Regulacja przez interkonwersję: aktywacja lub deaktywacja pewnych
białek poprzez reakcje fosforylacji i defosforylacji. Hormony indukują
cyklazę adenylową. Podwyższenie stężenia cAMP aktywuje przez
fosforylację lipazę hormonowrażliwą.
Tr
ia
c
y
lo
g
li
c
e
ro
le
K
w
a
s
y
tł
u
s
zc
zo
w
e
L
ip
id
y
b
ło
n
o
w
e
C
h
o
le
s
te
ro
l
C
ia
ła
k
e
to
n
o
w
e
A
c
e
ty
lo
-
C
o
A
S
y
n
te
za
k
w
a
s
ó
w
tł
u
s
zc
z
o
w
y
c
h
fo
s
fo
ry
la
c
ja
o
k
s
y
d
a
c
y
jn
a
β
-
o
k
s
y
d
a
c
ja
C
y
k
l
K
re
b
s
a
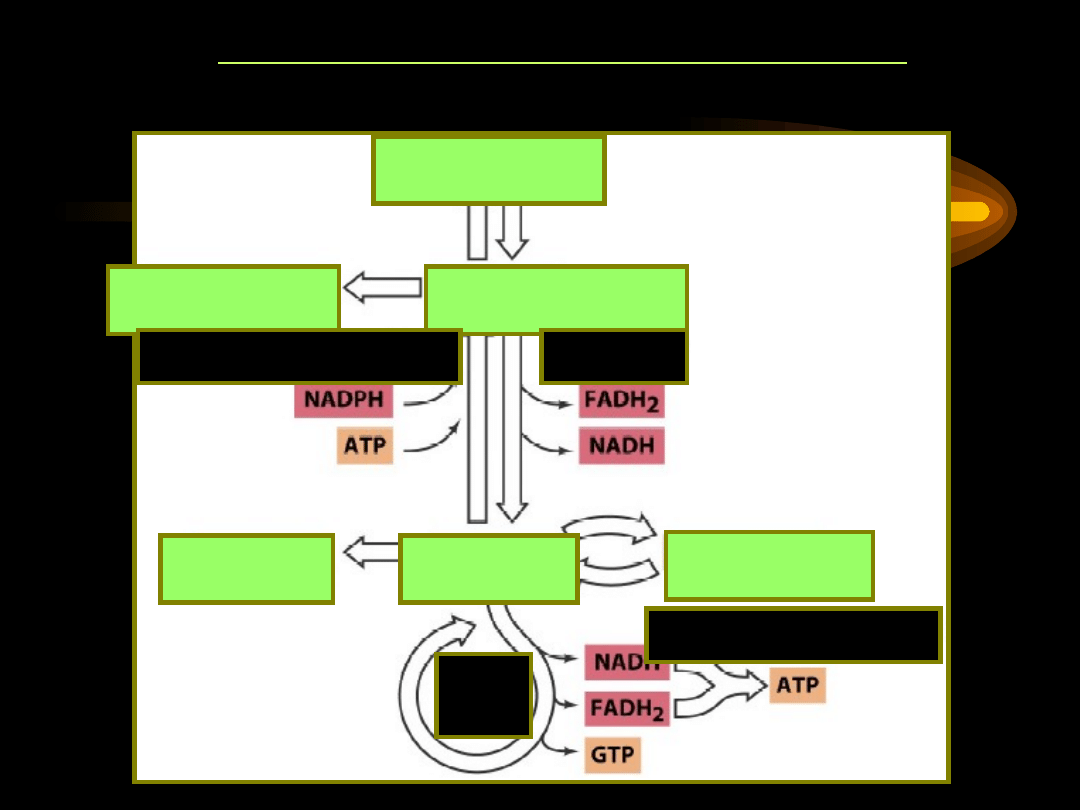
SCHEMAT METABOLIZMU LIPIDÓW

Źródłem kwasów tłuszczowych jest ich biosynteza oraz lipoliza
acylogliceroli i estrów cholesterolu. Procesy te zachodzą w
cytosolu.
Rozpad kwasów tłuszczowych jest procesem
utleniania (-oksydacji) i zachodzi w mitochondriach.
Kwasy tłuszczowe przeznaczone do utleniania muszą być więc
przetransportowane do wnętrza mitochondrium.
Etapy β-oksydacji:
•
aktywacja kwasu tłuszczowego poprzez estryfikację z CoA-SH,
•
transport acylo-Co-A przez błonę mitochondrialną,
•
reakcje procesu β-oksydacji:
-utlenienie (z udziałem FAD),
-hydratacja,
-utlenienie (z udziałem NAD
+
),
-tioliza (tiolityczne rozerwanie wiązania z udziałem drugiej
cząsteczki CoA-SH).
ROZPAD KWASÓW TŁUSZCZOWYCH (-OKSYDACJA)

Etapy β-oksydacji:
•
aktywacja
kwasu
tłuszczowego
poprzez
estryfikację z CoA-SH,
•
transport acylo-Co-A przez błonę mitochondrialną,
•
reakcje procesu β-oksydacji:
-utlenienie (z udziałem FAD),
-hydratacja,
-utlenienie (z udziałem NAD
+
),
-tioliza (tiolityczne rozerwanie wiązania z udziałem
drugiej cząsteczki CoA-SH).
ROZPAD KWASÓW TŁUSZCZOWYCH (-OKSYDACJA)
Zanim dojdzie do utlenienia kwasu tłuszczowego, musi on
zostać przekształcony w aktywny metabolit poprzez
przyłączenie CoA-SH. Proces ten wymaga energii
gromadzonej w ATP i obecności enzymu - syntetazy
acylo-CoA (tiokinazy).
R
-C
-O
H
+
C
o
A
-
S
H
O װ
O װ
s
y
n
te
ta
za
a
c
y
lo
-C
o
A
(t
io
k
in
a
za
)
A
T
P
A
M
P
+
P
P
i
2
P
i
R
-C
-S
-C
o
A
p
ir
o
fo
s
fa
ta
za
AKTYWACJA KWASÓW TŁUSZCZOWYCH

Etapy β-oksydacji:
• aktywacja kwasu tłuszczowego poprzez estryfikację z
CoA-SH,
•
transport
acylo-Co-A
przez
błonę
mitochondrialną,
•
reakcje procesu β-oksydacji:
-utlenienie (z udziałem FAD),
-hydratacja,
-utlenienie (z udziałem NAD
+
),
-tioliza (tiolityczne rozerwanie wiązania z udziałem
drugiej cząsteczki CoA-SH).
ROZPAD KWASÓW TŁUSZCZOWYCH (-OKSYDACJA)

•
Kwasy tłuszczowe o krótkim łańcuchu (do C10)
–
przenikają bezpośrednio do wnętrza mitochondrium. Są
aktywowane przez przyłączenie CoA-SH, kosztem energii z
rozpadu ATP, przy udziale mitochondrialnej syntetazy acylo~S-
CoA. Powstaje odpowiedni mitochondrialny acylo~S-CoA.
•
Kwasy tłuszczowe o długim łańcuchu (C12 i więcej)
– nie
mogą bezpośrednio wnikać do mitochondrium. Ich aktywacja
zachodzi w cytosolu, przez przyłączenie CoA-SH, pod wpływem
cytosolowej syntetazy acylo~S-CoA. Powstaje odpowiedni
cytosolowy acylo~S-CoA.
Błona mitochondrialna jest nieprzepuszczalna dla cytosolowego
acylo~S-CoA. W komórce istnieje więc system transportu reszt
acylowych z cytosolu do macierzy mitochondrialnej.
Przenośnikiem acylo~S-CoA jest karnityna, a transport nosi
nazwę czółenka karnitynowego (mostek karnitynowy).
TRANSPORT ACYLO-CoA PRZEZ BŁONĘ MITOCHONDRIALNĄ
MOSTEK KARNITYNOWY
H 3 C N C H 2 C H C H 2
C H 3
C H 3
O H
C O O
+
R
C
S C o A
O
+
H 3 C N C H 2 C H C H 2
C H 3
C H 3
O
C O O
+
C
R
O
+
H S C o A
carnitine
fatty acyl carnitine
Carnitine Palmitoyl
Transferase
k
a
rn
it
y
n
a
a
c
y
lo
k
a
rn
it
y
n
a
a
c
y
lo
tr
a
n
s
fe
ra
z
a
k
a
rn
it
y
n
o
w
a
Karnityna
–
organiczny związek
chemiczny,
syntetyzowany w
wątrobie, nerkach i
mózgu z lizyny i
metioniny. Pełni rolę
w transporcie
kwasów tłuszczowych
z cytosolu do
mitochondrium.

Mostek karnitynowy
polega na przenoszeniu reszt
acetylowych z cytosolowego acylo~S-CoA na karnitynę
z wytworzeniem acylokarnityny. Reakcja jest
katalizowana przez
acylotransferazę karnitynową I
.
Wiązanie między karnityną a grupą acylową jest
wiązaniem wysokoenergetycznym.
Acylokarnityna przy udziale
translokazy
, przenika do
macierzy mitochondrialnej, gdzie grupa acylowa zostaje
przeniesiona na mitochondrialny CoA-SH, wytworzony
zostaje mitochondrialny acylo~S-CoA. Reakcja
katalizowana jest przez
acylotransferazę
karnitynową II
. Uwolniona karnityna powraca do
cytosolu.
MOSTEK KARNITYNOWY
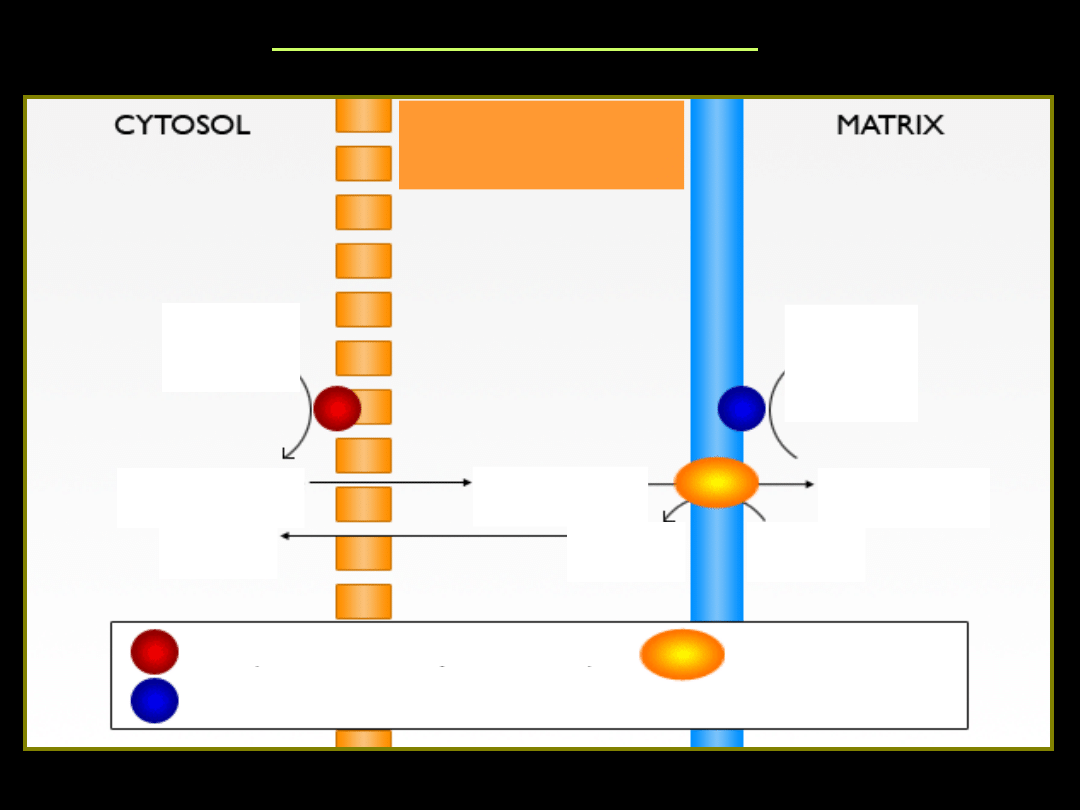
Tr
a
n
s
p
o
rt
a
c
y
lo
-C
o
A
z
c
y
to
s
o
lu
d
o
m
it
o
c
h
o
n
d
ri
u
m
a
c
y
lo
-C
o
A
+
k
a
rn
it
y
n
a
a
c
y
lo
-
C
o
A
+
k
a
rn
it
y
n
a
a
c
y
lo
k
a
rn
it
y
n
a
a
c
y
lo
k
a
rn
it
y
n
a
a
c
y
lo
k
a
rn
it
y
n
a
k
a
rn
it
y
n
a
k
a
rn
it
y
n
a
k
a
rn
it
y
n
a
tr
a
n
s
lo
k
a
z
a
a
c
y
lo
tr
a
n
s
fe
ra
za
k
a
rn
it
y
n
o
w
a
I
a
c
y
lo
tr
a
n
s
fe
ra
za
k
a
rn
it
y
n
o
w
a
I
I
MOSTEK KARNITYNOWY
z
e
w
n
ę
tr
z
n
a
b
ło
n
a
m
it
o
c
h
o
n
d
ri
a
ln
a
w
e
w
n
ę
tr
zn
a
b
ło
n
a
m
it
o
c
h
o
n
d
ri
a
ln
a

Etapy β-oksydacji:
• aktywacja kwasu tłuszczowego poprzez estryfikację z
CoA-SH,
• transport acylo-Co-A przez błonę mitochondrialną,
•
reakcje procesu β-oksydacji:
-utlenienie (z udziałem FAD),
-hydratacja,
-utlenienie (z udziałem NAD
+
),
-tioliza (tiolityczne rozerwanie wiązania z udziałem
drugiej cząsteczki CoA-SH).
ROZPAD KWASÓW TŁUSZCZOWYCH (-OKSYDACJA)

-oksydacja
to dwuetapowy proces utleniania kwasów
tłuszczowych.
Każdy etap składa się z wielu reakcji.
•
Pierwszy etap polega na wielokrotnie powtarzanych
reakcjach utleniania łańcucha węglowodorowego kwasu
tłuszczowego przy węglu i na rozpadzie utlenianego
substratu na fragmenty dwuwęglowe. Łańcuch węglowy jest
rozrywany pomiędzy atomami węgla i (stąd -oksydacja).
Każdy cykl -oksydacji powoduje skrócenie łańcucha kwasu
tłuszczowego o kolejny fragment dwuwęglowy.
Produktem -oksydacji jest wiele cząsteczek acetylo~S-CoA.
•
Drugi etap to utlenianie reszt acetylowych w cyklu kwasów
trikarboksylowych do CO
2
i H
2
O. Oba procesy dostarczają ATP.
-oksydacja zachodzi w macierzy mitochondrialnej.
-OKSYDACJA
p
a
lm
it
o
il
o
-
C
o
A
k
o
le
jn
e
u
s
u
w
a
n
ie
je
d
n
o
s
te
k
d
w
u
w
ę
g
lo
w
y
c
h
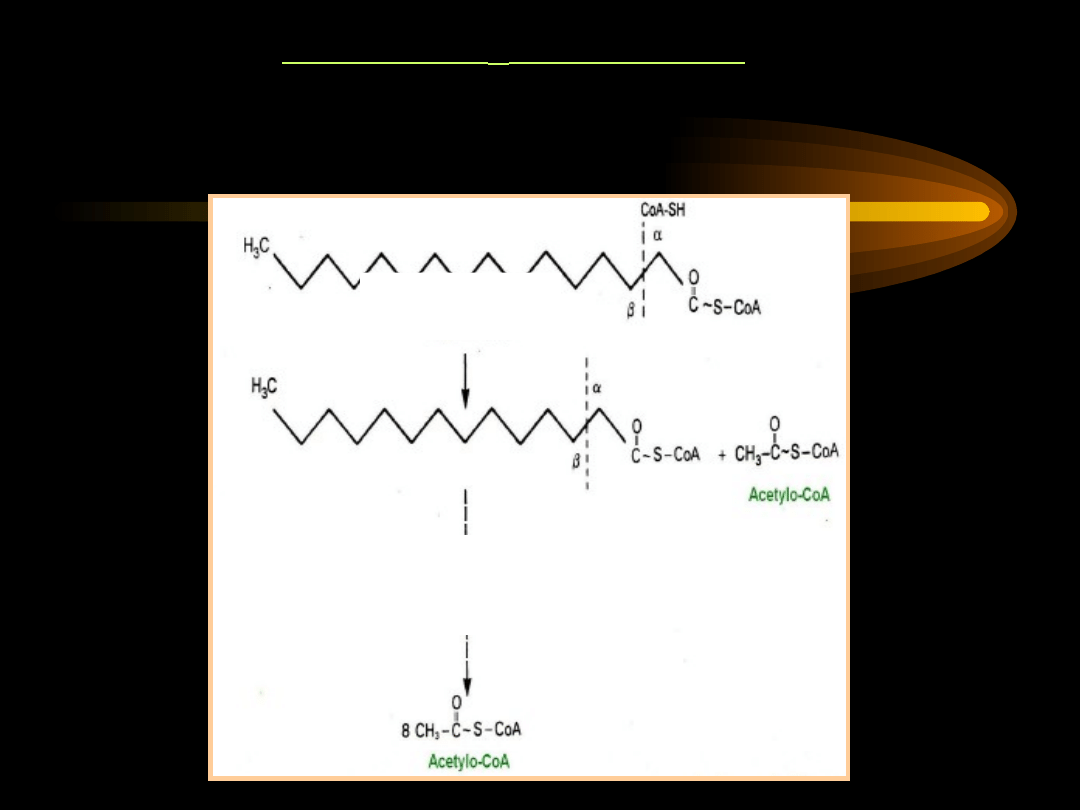
SCHEMAT -OKSYDACJI
-oksydacja kwasów tłuszczowych nasyconych o
parzystej liczbie atomów węgla
– w wyniku
zachodzących wielokrotnie czterech reakcji dochodzi do
całkowitego rozpadu kwasu tłuszczowego na n
cząsteczek acetylo~S-CoA (n=liczba par atomów węgla)
-OKSYDACJA
p
a
lm
it
o
il
o
-C
o
A
(
1
6
C
):
1
2
3
4
5
6
7
8
8 acetylo-CoA
p
a
lm
it
o
il
o
-C
o
A
+
7
F
A
D
+
7
N
A
D
+
+
7
C
o
A
+
7
H
2
O
8
a
c
e
ty
lo
-C
o
A
+
7
F
A
D
H
2
+
7
N
A
D
H
+
7
H
+
7 cykli
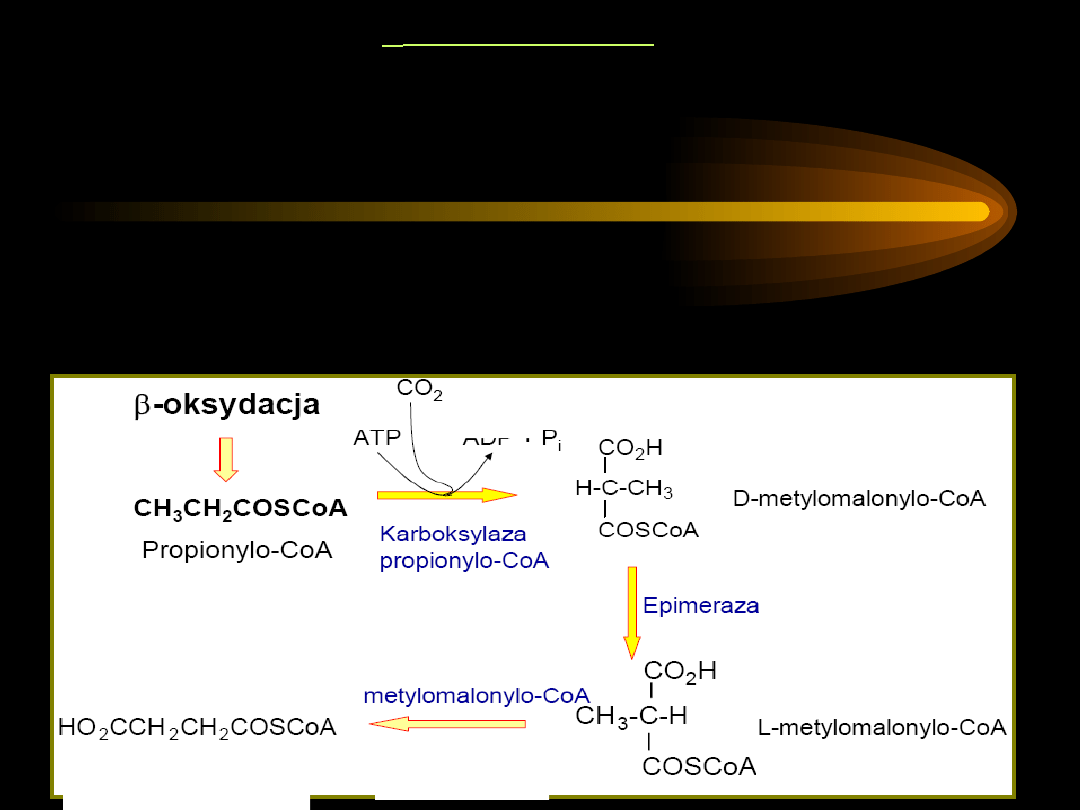
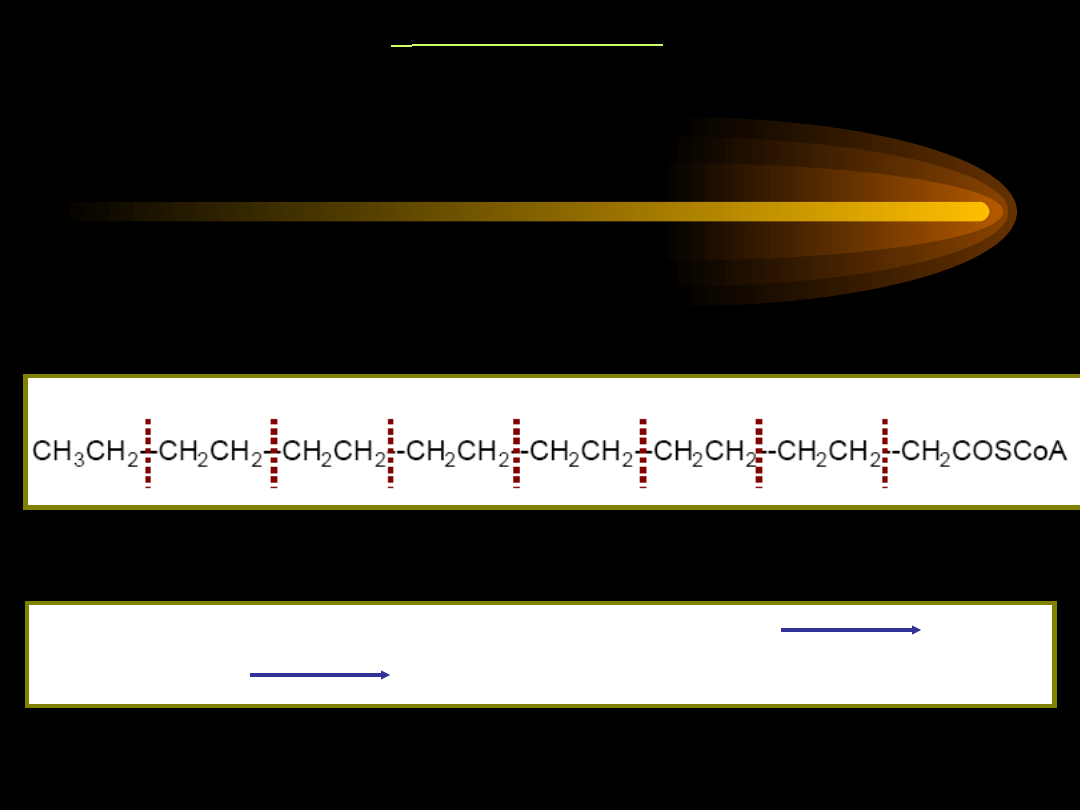
-oksydacja kwasów tłuszczowych nasyconych o nieparzystej
liczbie atomów węgla -
w wyniku utlenienia kwasu
tłuszczowego o nieparzystej liczbie atomów węgla powstają
cząsteczki acetylo-CoA i
1 cząsteczka trójwęglowa: propionylo-
CoA
.
Propionylo-CoA nie podlega dalszej -oksydacji, wymaga
specyficznych przekształceń do bursztynylo~S-CoA, który włącza
się do cyklu kwasów trikarboksylowych i ulega przekształceniu w
szczawiooctan (zużywany jako substrat w glukoneogenezie).
-OKSYDACJA
b
io
ty
n
a
w
it
a
m
in
a
B
1
2
B
u
rs
zt
y
n
y
lo
-
C
o
A
M
u
ta
za

-oksydacja kwasów tłuszczowych nienasyconych –
utlenianie kwasów tłuszczowych nienasyconych do
acetylo~CoA wymaga udziału
dodatkowych enzymów
.
Kwasy tłuszczowe z jednym podwójnym wiązaniem –
izomeraza akonitazowa.
Kwasy tłuszczowe z większą liczbą wiązań podwójnych –
reduktaza
, usuwająca niektóre wiązania podwójne drogą
redukcji.
-OKSYDACJA

Bilans energetyczny utleniania nasyconych kwasów
tłuszczowych (na przykładzie kwasu palmitynowego):
• - 2 cząsteczki ATP
potrzebne do aktywacji palmitynianu do
palmitoilo-CoA (jedna cząsteczka ATP ulega hydrolizie do
AMP i PPi, co jest równoznaczne z rozszczepieniem dwóch
wiązań wysokoenergetycznych)
• 35 cząsteczek ATP
z utlenienia FADH
2
i NADH + H
+
(powstałych w 7 obrotach cyklu β-oksydacji) w łańcuchu
oddechowym:
(2 ATP + 3 ATP) x 7 = 35 ATP
• 96 cząsteczek ATP
z utlenienia 8 cząsteczek acetylo-CoA
w cyklu Krebsa:
12 ATP x 8 = 96 ATP
EFEKT ENERGETYCZNY -OKSYDACJI
Z
y
s
k
n
e
tt
o
:
1
2
9
c
zą
s
te
c
ze
k
A
T
P
n
a
1
c
zą
s
te
c
zk
ę
p
a
lm
it
y
n
ia
n
u
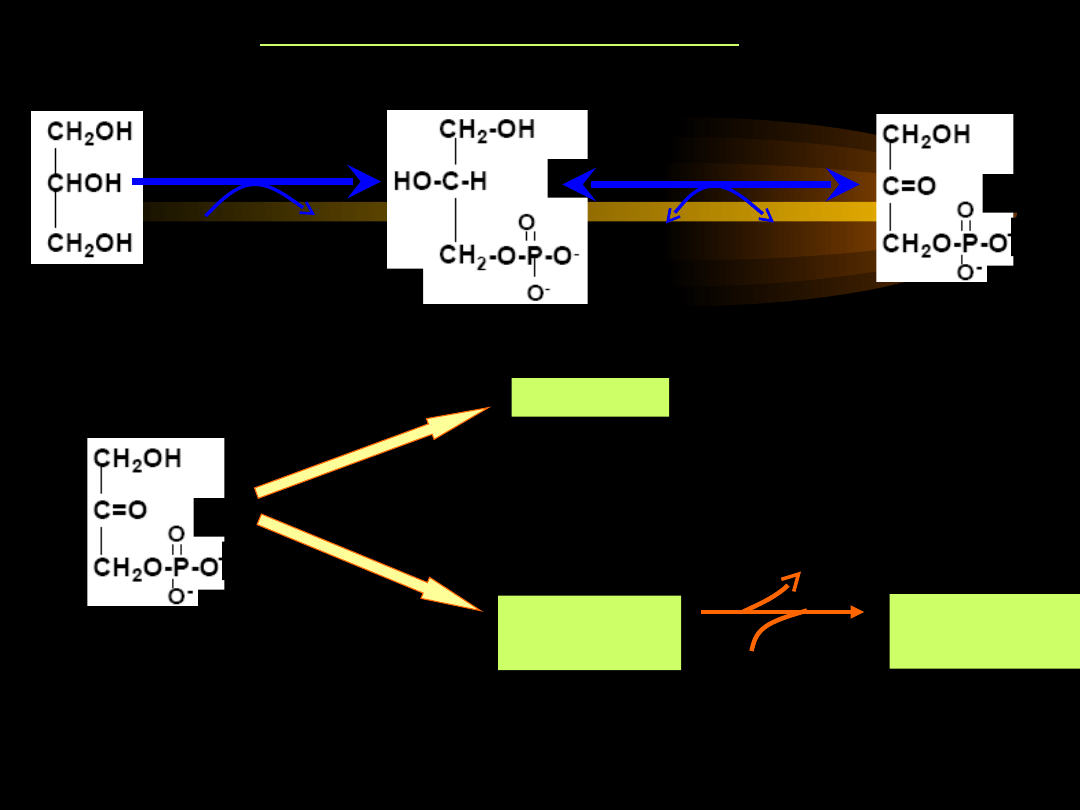
UTLENIANIE GLICEROLU
N
A
D
+
N
A
D
H
+
H
+
A
T
P
A
D
P
+
P
i
k
in
a
za
g
li
c
e
ro
lo
w
a
g
li
c
e
ro
lo
-3
-f
o
s
fo
ra
n
g
lu
k
o
z
a
p
ir
o
g
ro
n
ia
n
g
lu
k
o
n
e
o
g
e
n
e
za
g
li
k
o
li
za
a
c
e
ty
lo
-
C
o
A
C
O
2
C
o
A
-S
H
(d
o
m
in
u
ją
c
y
k
ie
ru
n
e
k
p
rz
e
m
ia
n
y
g
li
c
e
ro
lu
)
glicerol
fo
s
fo
d
ih
y
d
ro
k
s
y
a
c
e
t
o
n
dehydrogenaza
glicerolo-3-fosforanowa
fo
s
fo
d
ih
y
d
ro
k
s
y
a
c
e
t
o
n
G
li
c
e
ro
l,
j
a
k
o
p
ro
d
u
k
t
li
p
o
li
zy
p
o
w
s
ta
je
g
łó
w
n
ie
w
a
d
ip
o
c
y
ta
c
h
,
a
le
n
ie
m
o
że
b
y
ć
w
n
ic
h
m
e
ta
b
o
li
zo
w
a
n
y,
p
o
n
ie
w
a
ż
k
o
m
ó
rk
i
te
n
ie
p
o
s
ia
d
a
ją
k
in
a
zy
g
li
c
e
ro
lo
w
e
j.
T
ra
n
s
p
o
rt
o
w
a
n
y
j
e
s
t
z
k
rw
ią
d
o
w
ą
tr
o
b
y,
g
d
zi
e
j
e
s
t
s
u
b
s
tr
a
te
m
p
rz
e
d
e
w
s
zy
s
tk
im
w
g
lu
k
o
n
e
o
g
e
n
e
zi
e
.

Jeśli poziom acetylo-CoA powstającego w mitochondriach wątroby
w wyniku β-oksydacji jest dużo wyższy niż możliwości jego
wykorzystania w cyklu kwasu cytrynowego, zostaje uruchomiony
proces
biosyntezy ciał ketonowych
, zwany
KETOGENEZĄ
.
Do ciał ketonowych zalicza się:
aceton, acetooctan i -
hydroksymaślan
.
Powstają one z
reszt acetylowych w wątrobie
, są zużywane
jako substraty energetyczne w innych narządach.
Przepływ ciał ketonowych z wątroby do tkanek jest rezultatem
aktywnego mechanizmu enzymatycznego wytwarzającego te
związki w wątrobie oraz bardzo małej aktywności enzymów
odpowiedzialnych za ich zużywanie w tym narządzie.
W warunkach fizjologicznych, przy prawidłowej diecie, ilość ciał
ketonowych powstających w wątrobie jest mała. Narasta w
okresie
głodu
i u
chorych na cukrzycę
.
KETOGENEZA

Reszty acetylowe powstałe w wyniku oksydacyjnej dekarboksylacji
pirogronianu, -oksydacji kwasów tłuszczowych oraz przemian szkieletów
węglowodorowych niektórych aminokwasów utleniają się w cyklu kwasów
trikarboksylowych do CO
2
i H
2
O, albo służą za substrat w syntezie
kwasów tłuszczowych i cholesterolu.
W okresie głodu brakuje substratu do glikolizy, zmniejsza się produkcja
pirogronianu i szczawiooctanu. Reszty acetylowe nie mogą włączyć się do
cyklu kwasów trikarboksylowych z powodu braku szczawiooctanu.
Niedobór glukozy ogranicza szlak pentozofosforanowy, zmniejsza tym
samym produkcję NADPH+ H
+
, potrzebnego do wytwarzania kwasów
tłuszczowych i cholesterolu.
W przebiegu cukrzycy spowodowanej niedoborem insuliny glukoza, mimo że
jest dostarczana, nie może być przetwarzana. Prowadzi to do niedoboru
pirogronianu, szczawioctanu i NADPH+H
+
.
W warunkach głodzenia i cukrzycy
jedynym możliwym szlakiem
przemiany acetylo~S-CoA staje się ketogeneza.
KETOGENEZA
KETOGENEZA
C
H
3
–
C
–
S
–
C
o
A
a
c
e
ty
lo
~
S
-C
o
A
a
c
e
ty
lo
~
S
-
C
o
A
a
c
e
to
a
c
e
ty
lo
~
S
-
C
o
A
C
H
3
–
C
–
S
–
C
o
A
O
+
O
C
o
A
-S
H
ti
o
la
za
a
c
e
ty
lo
~
S
-C
o
A
β
-h
y
d
ro
k
s
y
-β
-
m
e
ty
lo
g
lu
ta
ry
lo
~
S
-
C
o
A
(
H
M
G
~
S
-C
o
A
)
C
H
3
–
C
–
C
H
2
–
C
–
S
–
C
o
A
O
O
-
O
O
C
–
C
H
2
–
C
–
C
H
2
–
C
–
S
–
C
o
A
O
O
H
C
H
3
C
o
A
-S
H
s
y
n
ta
za
β
-
h
y
d
ro
k
s
y
-β
-
m
e
ty
lo
g
lu
ta
ry
lo
-
C
o
A
(
H
M
G
~
S
-C
o
A
)
C
H
3
–
C
–
S
–
C
o
A
O
H
2
O
Biosynteza ciał ketonowych z acetylo~S-CoA zachodzi w mitochondriach
wątroby.
W pierwszym etapie, katalizowanym przez tiolazę, powstaje
acetoacetylo~S-CoA.
Do niego przyłącza się kolejna reszta octanowa z acetylo~S-CoA.
Powstaje HMG~S-CoA.
KETOGENEZA
-
O
O
C
–
C
H
2
–
C
–
C
H
2
–
C
–
S
–
C
o
A
O
O
H
C
H
3
H
M
G
~
S
-C
o
A
li
a
za
H
M
G
-C
o
A
C
H
3
–
C
–
S
–
C
o
A
O
a
c
e
ty
lo
~
S
-C
o
A
-
O
O
C
–
C
H
2
–
C
–
C
H
3
O
O
H
a
c
e
to
o
c
ta
n
N
A
D
+
N
A
D
H
+
H
+
d
e
h
y
d
ro
g
e
n
a
za
β
-
h
y
d
ro
k
s
y
m
a
ś
la
n
o
w
a
β
-
h
y
d
ro
k
s
y
m
a
ś
la
n
C
O
2
s
a
m
o
is
tn
a
d
e
k
a
rb
o
k
s
y
la
c
ja
C
H
3
–
C
–
C
H
3
O
a
c
e
to
n
W kolejnych etapach, liaza HMG~S-CoA powoduje rozpad HMG~S-CoA na
acetooctan i acetylo~S-CoA. Acetooctan może zostać przekształcony w
-hydroksymaślan lub ulec samoistnej dekarboksylacji w wytworzeniem acetonu.
CH
3
- C – CH
2
–
C
H
O
O
-

• Ciała ketonowe dyfundują z mitochondriów wątroby do krwi, są transportowane do
tkanek docelowych, są rozpuszczalne w wodzie, nie wymagają przenośników.
• Ciała ketonowe służą jako wartościowy materiał energetyczny dla tkanek
pozawątrobowych. Są wychwytywane przez mięśnie szkieletowe, mięsień sercowy,
korę nerki.
• Źródłem energii dla mózgu i krwinek czerwonych w warunkach prawidłowych jest
glukoza, jednakże mózg przystosowuje się do wykorzystania acetooctanu u
cukrzyków i podczas głodu.
• Aceton jest nieprzydatny w metabolizmie i jest wydalany z moczem oraz
wydychany z powietrzem przez płuca.
• W celu włączenia ciał ketonowych w przemianę energetyczną w tkankach
pozawątrobowych, acetooctan jest przekształcany do acetylo-CoA i utleniany w
cyklu kwasu cytrynowego (ale nie w wątrobie).
• Jeśli wytwarzanie ciał ketonowych przekracza ich pozawątrobowe wykorzystanie,
dochodzi do zwiększonego stężenia tych związków w osoczu (ketonemia), a w
końcu do ich wydalenia z moczem (ketonuria).
• Oba zjawiska występują po dłuższym okresie głodzenia oraz w przypadku
nieskutecznego leczenia cukrzycy.
KATABOLIZM CIAŁ KETONOWYCH
ROZKŁAD CIAŁ KETONOWYCH
-
O
O
C
–
C
H
2
–
C
H
–
C
H
3
O
H
a
c
e
to
o
c
ta
n
N
A
D
+
N
A
D
H
+
H
+
d
e
h
y
d
ro
g
e
n
a
za
β
-
h
y
d
ro
k
s
y
m
a
ś
la
n
o
w
a
β
-
h
y
d
ro
k
s
y
m
a
ś
la
n
-
O
O
C
–
C
H
2
–
C
–
C
H
3
O
C
H
3
–
C
–
C
H
2
–
C
–
S
–
C
o
A
O
a
c
e
to
a
c
e
ty
lo
~
S
-C
o
A
O
C
o
A
-
tr
a
n
s
fe
ra
za
β
-
k
e
to
k
w
a
s
o
w
a
b
u
rs
zt
y
n
i
a
n
b
u
rs
zt
y
n
y
lo
-
C
o
A
C
H
3
–
C
–
S
–
C
o
A
O
2
x
a
c
e
ty
lo
~
S
-
C
o
A
ti
o
la
za
C
o
A
-S
H
Rozkład ciał ketonowych polega na:
• utlenieniu -hydroksymaślanu
przez dehydrogenazę -hydroksy-
maślanową do acetooctanu.
Acetooctan pod działaniem tiokinazy
zamienia się w 2 cząsteczki
acetylo~S-CoA.
• przeniesieniu z bursztynylo~S-CoA
przez transferazę CoA na acetooctan.
Powstaje acetoacetylo~S-CoA, który
rozpada się pod wpływem tiolazy
na 2 cząsteczki acetylo~S-CoA.
Reszty acetylowe są włączane do
cyklu kwasów trikarboksylowych.

•
Zachodzi przede wszystkim w
wątrobie, tkance tłuszczowej, nerce,
płucach, mózgu oraz w gruczole mlekowym w okresie laktacji
.
•
Jest zlokalizowana w
cytosolu
komórki (w przeciwieństwie do
utleniania kwasów tłuszczowych, które ma miejsce w macierzy
mitochondrialnej).
•
Substratami dla tego procesu są
acetylo-CoA
i
NADPH + H
+
,
a
dawcą energii jest
ATP
.
•
Źródłem acetylo-CoA są: oksydacyjna, dekarboksylacja pirogronianu,
β-oksydacja kwasów tłuszczowych, rozpad ciał ketonowych i szkieletów
węglowodorowych niektórych aminokwasów
.
•
Źródłem NADPH + H
+
jest szlak pentozofosforanowy i dekarboksylacja
jabłczanu przez enzym jabłczanowy.
•
Syntezę kwasów tłuszczowych de novo katalizuje
syntaza kwasów
tłuszczowych
– kompleks enzymów o siedmiu aktywnościach
enzymatycznych, połączonych w łańcuch polipeptydowy.
•
Rosnący łańcuch kwasu tłuszczowego ulega wydłużeniu przez kolejne
dobudowywanie dwuwęglowych jednostek, pochodzących z acetylo-
CoA.
•
Wydłużanie jest zahamowane po zsyntetyzowaniu palmitynianu (C16).
•
Reakcje dalszego wydłużania i wprowadzania wiązań podwójnych są
prowadzone przez inne układy enzymatyczne.
BIOSYNTEZA KWASÓW TŁUSZCZOWYCH
Procesy generujące acetylo-CoA zachodzą w mitochondrium,
natomiast biosynteza kwasów tłuszczowych w cytosolu. Ponieważ
błona
mitochondrialna
jest
nieprzepuszczalna
dla
mitochondrialnego acetylo-CoA, istnieje specjalny mechanizm
transportu acetylo-CoA, zwany
mostkiem cytrynianowym.
TRANSPORT ACETYLO-CoA DO CYTOSOLU
s
zc
za
w
io
o
c
ta
n
c
y
tr
y
n
ia
n
m
it
o
c
h
o
n
d
ri
u
m
s
y
n
ta
za
c
y
tr
y
n
ia
n
o
w
a
H
2
O
C
o
A
-S
H
c
y
tr
y
n
ia
n
c
y
to
s
o
l
a
c
e
ty
lo
-C
o
A
c
y
to
s
o
lo
w
y
A
T
P
s
zc
za
w
io
o
c
ta
n
A
T
P
-l
ia
za
c
y
tr
y
n
ia
n
o
w
a
C
o
A
-S
H
A
D
P
+
P
i
m
it
o
c
h
o
n
d
ri
a
ln
y
a
c
e
ty
lo
-C
o
A

Etapy biosyntezy kwasów tłuszczowych:
•
Karboksylacja acetylo-CoA do malonylo-CoA z udziałem
karboksylazy acetylo-CoA.
•
Cykle elongacji łańcucha kwasów tłuszczowych, prowadzone przez
syntazę kwasów tłuszczowych, na które składają się:
-
Kondensacja
acetylo-ACP i malonylo-ACP
enzym:
syntaza β-ketoacylowa
- Redukcja
acetoacetylo-ACP do β-hydroksybutyrylo-ACP
enzym:
reduktaza β-ketoacylowa
- Odwodnienie
β-hydroksybutyrylo-ACP do krotonoilo-ACP
enzym:
dehydrataza β-hydroksyacylo-ACP
- Redukcja
krotonoilo-ACP do butyrylo-ACP
enzym:
reduktaza enoilowa
BIOSYNTEZA KWASÓW TŁUSZCZOWYCH
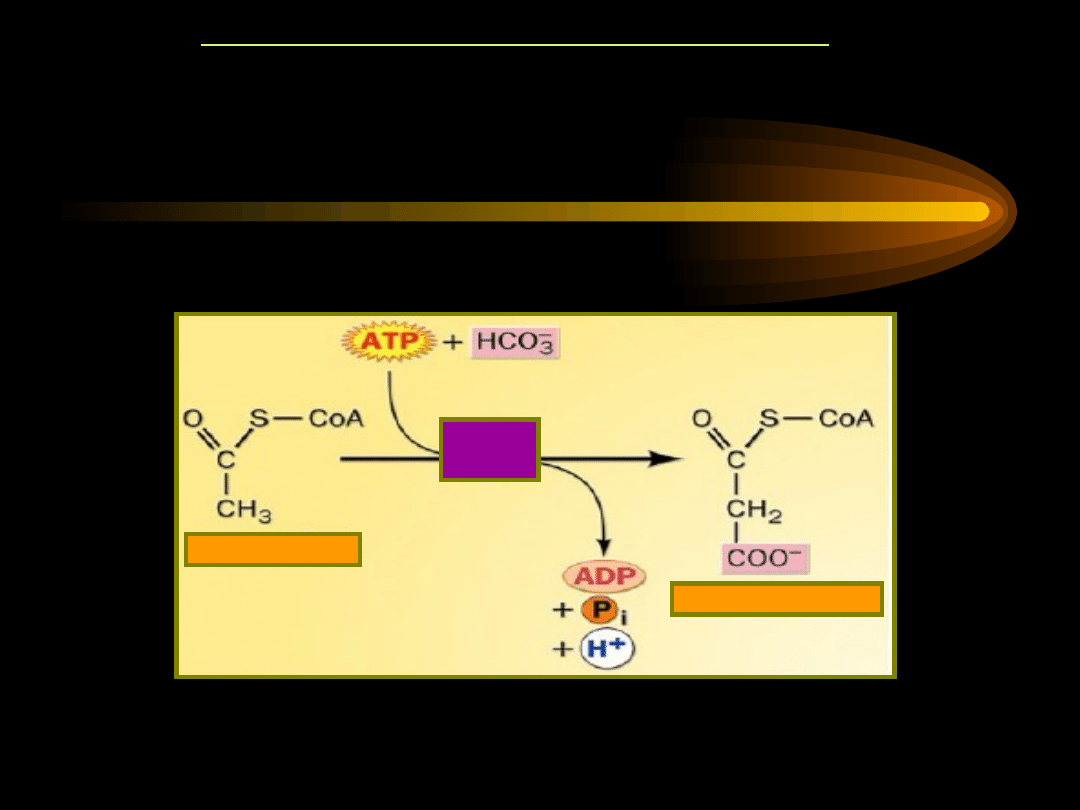
Każda (z wyjątkiem jednej) grupa acetylowa zużywana w
biosyntezie kwasów tłuszczowych jest karboksylowana
(wiąże CO
2
) z wytworzeniem malonylo~S-CoA. Reakcja
ta jest katalizowana przez karboksylazę acetylo~S-CoA
(koenzymem jest karboksybiotyna).
KARBOKSYLACJA ACETYLO-CoA
a
c
e
ty
lo
-C
o
A
m
a
lo
n
y
lo
-C
o
A
b
io
ty
n
a
POWSTAWANIE ŁAŃCUCHA KWASU TŁUSZCZOWEGO
s
y
n
ta
za
k
e
to
a
c
y
lo
w
a
a
c
e
ty
lo
-
tr
a
n
s
fe
ra
z
a
m
a
lo
n
y
lo
-
tr
a
n
s
fe
ra
za
re
d
u
k
ta
z
a
e
n
o
il
o
w
a
d
e
h
y
d
ra
ta
za
re
d
u
k
ta
z
a
k
e
to
a
c
y
lo
w
a
A
C
P
ti
o
e
s
te
ra
z
a
re
d
u
k
ta
z
a
k
e
to
a
c
y
lo
w
a
A
C
P
m
a
lo
n
y
lo
-
tr
a
n
s
fe
ra
z
a
a
c
e
ty
lo
-
tr
a
n
s
fe
ra
z
a
d
e
h
y
d
ra
ta
z
a
re
d
u
k
ta
z
a
e
n
o
il
o
w
a
s
y
n
ta
za
k
e
to
a
c
y
lo
w
a
C
y
s
S
H
S
H
P
a
n
P
a
n
S
H
S
H
C
y
s
p
o
d
z
ia
ł
p
o
d
je
d
n
o
s
te
k
1
2
S
y
n
ta
z
a
k
w
a
s
ó
w
tł
u
s
z
c
z
o
w
y
c
h
j
e
st
d
im
e
re
m
o
d
w
ó
ch
i
d
e
n
ty
cz
n
y
ch
p
o
d
je
d
n
o
st
ka
ch
p
o
lip
e
p
ty
d
o
w
y
ch
1
i
2
,
sk
ła
d
a
ją
cy
ch
si
ę
z
si
e
d
m
iu
a
k
ty
w
n
o
śc
i
ka
ta
lit
y
cz
n
y
ch
i
b
ia
łk
a
p
rz
e
n
o
sz
ą
ce
g
o
a
cy
l
-
A
C
P
(
a
n
g
.
a
cy
l
c
a
rr
ie
r
p
ro
te
in
).
R
ó
w
n
o
cz
e
śn
ie
w
y
tw
a
rz
a
n
e
s
ą
d
w
a
ł
a
ń
cu
ch
y
a
cy
lo
w
e
.
Syntaza kwasów tłuszczowych
rozpoczyna działanie od jednej
cząsteczki acetylo-CoA i wydłuża ją z użyciem reszt
malonylowych w siedmiu cyklach reakcji do palmitynianu.
Podczas każdego cyklu odłącza się CO
2
, dlatego łańcuch kwasu
tłuszczowego wydłuża się za każdym razem o jednostki
dwuwęglowe.
Pierwszym etapem elongacji kwasów tłuszczowych jest związanie
się substratów z ACP:
a
c
e
ty
lo
-
tr
a
n
s
fe
ra
za
m
a
lo
n
y
lo
-
tr
a
n
s
fe
ra
za
C
H
3
–
C
–
S
–
C
o
A
O
C
H
3
–
C
–
S
–
A
C
P
O
A
C
P
-S
H
C
o
A
-
S
H
a
c
e
ty
lo
-C
o
A
a
c
e
ty
lo
-A
C
P
-
O
O
C
–
C
H
2
–
C
–
S
–
C
o
A
O
A
C
P
-S
H
C
o
A
-
S
H
m
a
lo
n
y
lo
-
C
o
A
m
a
lo
n
y
lo
-
A
C
P
-
O
O
C
–
C
H
2
–
C
–
S
–
A
C
P
O
BIOSYNTEZA KWASÓW TŁUSZCZOWYCH

SUMARYCZNA REAKCJA BIOSYNTEZY KT
8 acetylo-CoA + 14 NADPH + 14 H
+
+ 7 ATP
kwas palmitynowy + 8 CoA-SH + 14 NADP
+
+ 7 ADP + 7 P
i
+ 7 H
2
O
•
P
ro
d
u
kt
e
m
s
y
n
ta
zy
k
w
a
só
w
t
łu
sz
cz
o
w
y
ch
j
e
st
w
p
e
łn
i
n
a
sy
co
n
y
1
6
-w
ę
g
lo
w
y
k
w
a
s
tł
u
sz
cz
o
w
y
–
k
w
a
s
p
a
lm
it
y
n
o
w
y.
•
W
sz
y
st
ki
e
–
z
w
y
ją
tk
ie
m
d
w
ó
ch
p
ie
rw
sz
y
ch
a
to
m
ó
w
w
ę
g
la
(
lic
zą
c
o
d
k
o
ń
ca
m
e
ty
lo
w
e
g
o
),
w
b
u
d
o
w
u
ją
s
ię
d
o
k
w
a
su
p
a
lm
it
y
n
o
w
e
g
o
p
o
p
rz
e
z
m
a
lo
n
y
lo
-C
o
A
.
N
a
to
m
ia
st
p
ie
rw
sz
a
p
a
ra
a
to
m
ó
w
w
ę
g
la
p
o
ch
o
d
zi
b
e
zp
o
śr
e
d
n
io
z
a
ce
ty
lo
-C
o
A
.

-OKSYDACJA A BIOSYNTEZA KWASÓW TŁUSZCZOWYCH
β-oksydacja KT
Synteza KT
Lokalizacja
macierz
mitochondrialna
cytosol
Enzymy
oddzielne
polipeptydy
kompleksy
polipeptydowe
Charakter reakcji
utlenianie
redukcja
Nośniki reszt
acylowych
koenzym A
ACP i cysteina
Akceptor/donor
elektronów
NAD
+
i FAD
NADPH + H
+
Produkt/donor
jednostek
węglowych
acetylo-CoA
malonylo-CoA
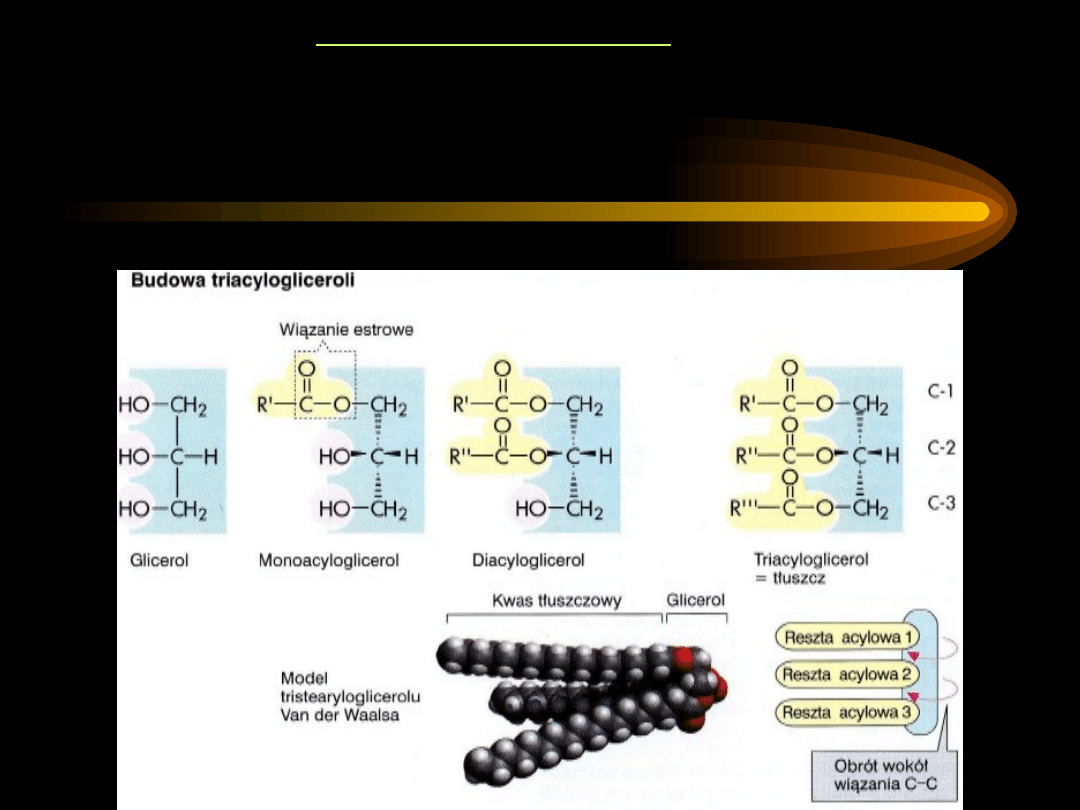
ACYLOGLICEROLE
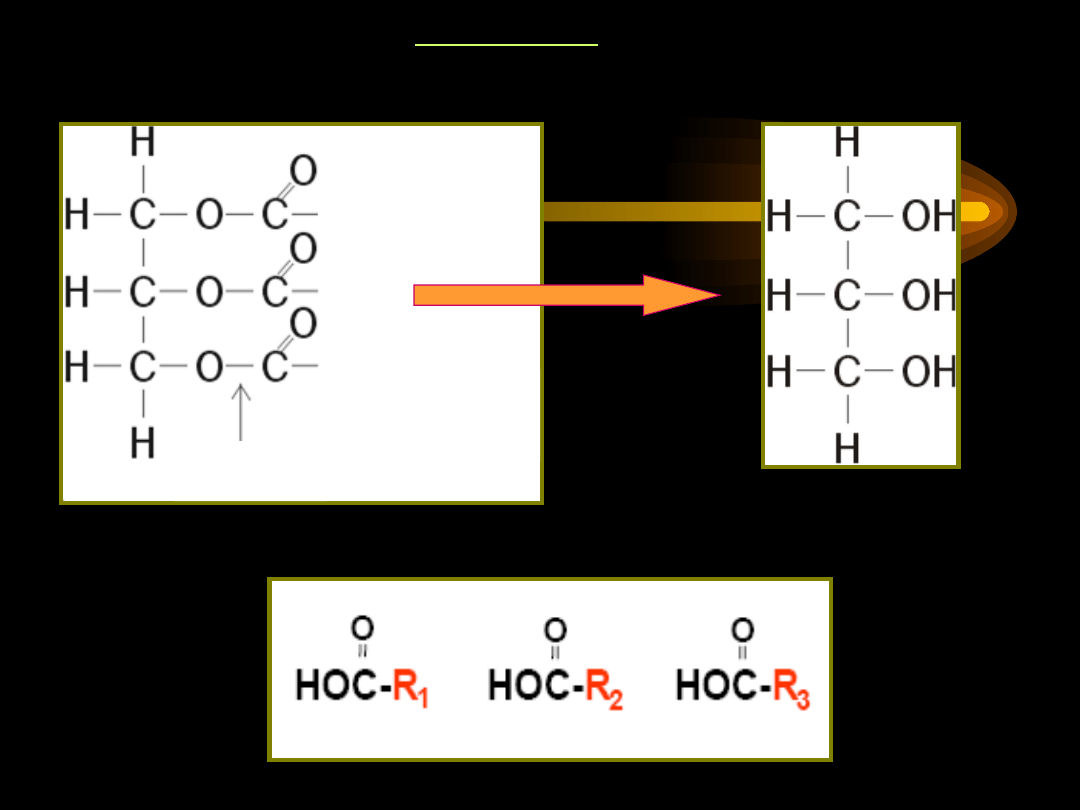
Acyloglicerole należą do tłuszczów właściwych, stanowią większość
lipidów organizmu. Ze względu na budowę należą do estrów:
• składnik alkoholowy – glicerol
• składnik kwasowy – jednokarboksylowe kwasy tłuszczowe

Triacyloglicerole
są estrami alkoholu trójwodorotlenowego –
glicerolu i trzech kwasów tłuszczowych.
Są głównymi lipidami
tłuszczu zapasowego organizmu i tłuszczu zawartego w
pokarmach.
Trzy reszty acylowe cząsteczki triacyloglicerolu mogą się różnić
długością swych łańcuchów oraz liczbą podwójnych wiązań, czego
konsekwencją
jest
duża
liczba
możliwych
kombinacji
pojedynczych cząsteczek.
Nienasycone kwasy tłuszczowe przeważnie znajdują się przy
środkowym atomie węgla glicerolu.
TRIACYLOGLICEROLE
H
2
C – OH
HO – C – H
H
2
C – OH
O
O H
2
C – O – C – R
1
R
2
– C – O – C – H
H
2
C – O – C – R
3
O
Glicerol
Triacyloglicerol

Biosynteza triacylogliceroli
zachodzi w
wątrobie, nerce,
jelicie, mięśniach, tkance tłuszczowej i w gruczole
mlekowym w okresie laktacji.
• Syntetyzowane w komórkach tłuszczowych są
przechowywane w cytosolu jako zapasowy materiał
energetyczny
• Powstałe w wątrobie wbudowywane są do kompleksów
lipoproteinowych i z krwią transportowane są do innych
tkanek
SYNTEZA TRIACYLOGLICEROLI

•
Triacyloglicerole są syntetyzowane z
glicerolo-3-fosforanu i acylo-
CoA.
•
Pierwszy etap syntezy triacylogliceroli to powstanie
glicerolo-3-
fosforanu.
•
Glicerolo-3-fosforan może powstać poprzez:
- fosforylację glicerolu – tylko w wątrobie, jelicie, nerkach oraz
gruczole mlekowym w okresie laktacji, gdyż występuje tam kinaza
glicerolowa, która katalizuje aktywację glicerolu. Wolny glicerol
powstały w wyniku lipolizy, jest transportowany do wątroby. Tam, pod
wpływem kinazy glicerolowej jest przetwarzany do glicerolo-3-
fosforanu, dawcą fosforanu jest ATP
- redukcję fosfodihydroksyacetonu - w
mięśniach i w tkance
tłuszczowej większość glicerolo-3-fosforanu pochodzi ze związku
pośredniego glikolizy – fosfodihydroksyacetonu.
SYNTEZA TRIACYLOGLICEROLI

•
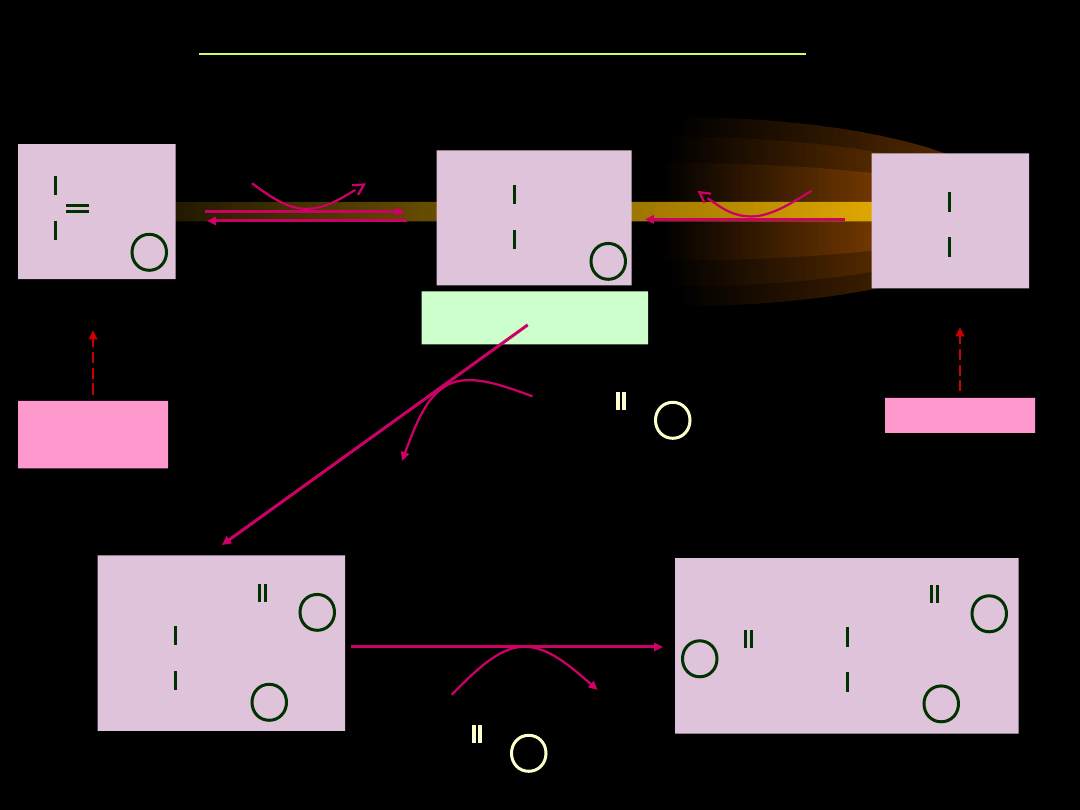
Glicerolo-3-fosforan jest estryfikowany dwiema resztami kwasów
tłuszczowych, pochodzącymi z dwóch cząsteczek acylo~S-CoA.
•
W wyniku połączenia cząsteczki acylo-CoA z glicerolo-3-
fosforanem powstaje najpierw
kwas lizofosfatydowy
(1-
acyloglicerolo-3-fosforan), a z niego
kwas fosfatydowy
(1,2-
diacyloglicerolofosforan).
SYNTEZA TRIACYLOGLICEROLI
g
li
c
e
ro
lo
-3
-
fo
s
fo
ra
n
H
2
C
–
O
H
O
H
–
C
H
H
2
C
–
O
–
P
H
2
C
–
O
H
O
H
–
C
H
g
li
c
e
ro
l
d
e
h
y
d
ro
g
e
n
a
za
g
li
c
e
ro
lo
-3
-
fo
s
fo
ra
n
o
w
a
k
in
a
z
a
g
li
c
e
ro
lo
w
a
N
A
D
+
N
A
D
H
+
H
+
A
T
P
A
D
P
H
2
C
–
O
–
C
–
R
O
H
–
C
H
H
2
C
–
O
–
P
O
a
c
y
lo
tr
a
n
s
fe
ra
z
a
g
li
c
e
ro
lo
-3
-
fo
s
fo
ra
n
o
w
a
C
o
A
–
S
–
C
–
R
O
a
c
y
lo
-C
o
A
k
w
a
s
l
iz
o
fo
s
fa
ty
d
o
w
y
C
o
A
-S
H
H
2
C
–
O
–
C
–
R
R
–
C
–
O
–
C
H
H
2
C
–
O
–
P
O
k
w
a
s
f
o
s
fa
ty
d
o
w
y
O
C
o
A
–
S
–
C
–
R
O
a
c
y
lo
-C
o
A
C
o
A
-S
H
a
c
y
lo
tr
a
n
s
fe
ra
z
a
1
-a
c
y
lo
g
li
c
e
ro
lo
-3
-
fo
s
fo
ra
n
o
w
a
G
L
IK
O
L
IZ
A
P
O
K
A
R
M
SYNTEZA TRIACYLOGLICEROLI
fo
s
fo
d
ih
y
d
ro
k
s
y
a
c
e
to
n
H
2
C
–
O
H

•
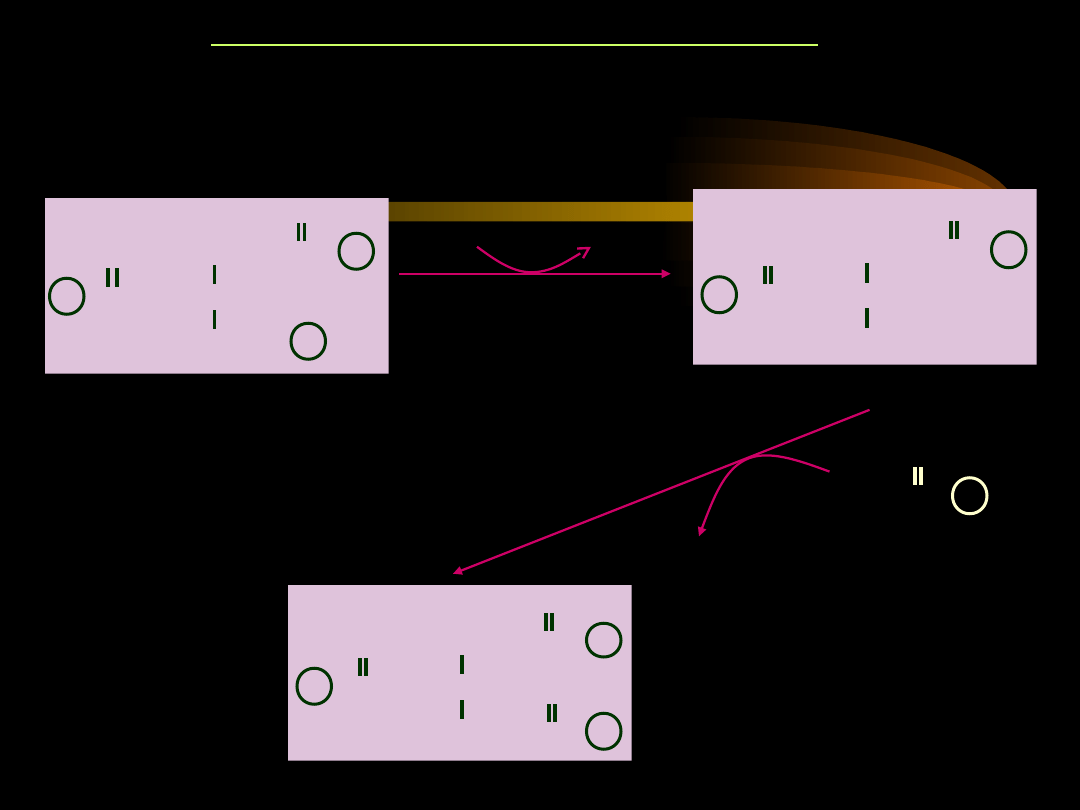
W kolejnym etapie, po usunięciu reszty fosforanowej
powstaje
1,2-diacyloglicerol
(DAG), który w reakcji
acylacji przekształca się w
triacyloglicerol
(TAG).
•
Energia niezbędna do syntezy triacyloglicerolu pochodzi
z hydrolizy wiązań tioestrowych w acylo-CoA.
SYNTEZA TRIACYLOGLICEROLI
SYNTEZA TRIACYLOGLICEROLI
H
2
C
–
O
–
C
–
R
R
–
C
–
O
–
C
H
H
2
C
–
O
–
P
O
k
w
a
s
f
o
s
fa
ty
d
o
w
y
O
H
2
C
–
O
–
C
–
R
R
–
C
–
O
–
C
H
H
2
C
–
O
H
O
1
,2
-d
ia
c
y
lo
g
li
c
e
ro
l
O
H
2
O
P
i
fo
s
fa
ta
z
a
fo
s
fa
ty
d
a
n
o
w
a
H
2
C
–
O
–
C
–
R
R
–
C
–
O
–
C
H
H
2
C
–
O
–
C
–
R
O
tr
ia
c
y
lo
g
li
c
e
ro
l
O
O
C
o
A
–
S
–
C
–
R
O
a
c
y
lo
-C
o
A
C
o
A
-S
H
a
c
y
lo
tr
a
n
s
fe
ra
z
a
d
ia
c
y
lo
g
li
c
e
ro
lo
w
a

Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
- Slide 41
- Slide 42
- Slide 43
- Slide 44
- Slide 45
- Slide 46
- Slide 47
- Slide 48
- Slide 49
- Slide 50
- Slide 51
- Slide 52
- Slide 53
- Slide 54
- Slide 55
- Slide 56
- Slide 57
- Slide 58
- Slide 59
- Slide 60
- Slide 61
- Slide 62
- Slide 63
- Slide 64
- Slide 65
Wyszukiwarka
Podobne podstrony:
METABOLIZM LIPIDOW cz II RM
METABOLIZM LIPIDÓW cz I RM
Metabolizm lipidów cz
Wykład 9 Oznaczenia lipidów cz 2
Wykład 8 Oznaczenia lipidów cz 1
cz V, wykłady, wentylacja, Minikowski, PAWEŁ KLIMA
Metabolizm lipidów (1)
IV metabolizm lipidow i sterydow
3 Bazy danych SQL cz 1 wykład
Bazy danych - podstawowe kroki w projektowaniu cz 2 - wyklady, Zajęcia z Baz Danych - MS Access, cz
Bazy danych - podstawowe kroki w projektowaniu cz 2 - wyklady, Zajęcia z Baz Danych - MS Access, cz
METABOLIZM LIPIDOW
Metabolizm lipidów 3
01 Wykłady Pisma Świętego C. T. Russella (cz. 1-4), Wykłady Pisma Świętego C. T. Russella (cz. 1-4)
3 Bazy danych SQL cz 2 wykład
3 Bazy danych SQL cz 2 wykład
Świadectwa cz.1, WYKŁADY, Zabójstwo dzieci poczętych
więcej podobnych podstron