
Martwe geny na emalię zębów
Autor tekstu: Jerry Coyne
Tłumaczenie: Małgorzata Koraszewska
Narządy szczątkowe przywołuje się często jako dowód ewolucji; sam napisałem niemal cały
rozdział o nich w książce Why Evolution Is True. A chociaż niektóre z tych narządów mogą mieć
jakąś funkcję (na przykład uważa się obecnie, że wyrostek robaczkowy odgrywa pewną rolę
w układzie odpornościowym), fakt funkcjonalności nie umniejsza ich szczątkowości — lub ich
pożyteczności jako dowodu na wspólne pochodzenie. Fakt, że „skrzydła" pingwinów służą teraz jako
płetwy, nie oznacza, iż nie mówią niczego o pochodzeniu pingwinów od fruwających ptaków. (Moja
definicja cechy szczątkowej brzmi luźno: „cecha, która obecnie służy funkcji zupełnie innej niż ta, do
której pchała ją początkowo ewolucja i której ślady oryginalnej budowy nadal istnieją".)
A poza tym pewne narządy szczątkowe wydają się nie mieć w ogóle żadnej funkcji. Miałbyś
trudności z twierdzeniem, że istnieje jakaś funkcja dla maleńkich, podobnych do guziczków skrzydeł
kiwi, zagrzebanych głęboko w piórach na jego bokach.
Ale miałbyś jeszcze większe trudności z argumentowaniem, że martwe geny mają jakąś
funkcję.
Jednym z przewidywań ewolucji jest, że jeśli istnieją narządy szczątkowe, to z pewnością
muszą istnieć geny szczątkowe, ponieważ cechy, które kurczą się lub znikają, z pewnością muszą
być oparte, przynajmniej czasami, o geny, które znikają. Ale geny nie są po prostu wycinane
w całości z DNA, kiedy nie są już dłużej potrzebne: dobór naturalny wyłącza je, przez wycięcie
odcinka DNA (delecja), wstawienie „kodonu STOP" (zmiana w DNA, która przedwcześnie kończy
odczytywanie białka z kodu DNA, tworząc "mutację zmiany odczytu ramki" - zmieniając trójkowy
kod tak, że przesuwa się cała rama kodowa sekwencji DNA) lub tak zmieniając regulacyjny region
genu, że białko już nie powstaje. Resztki genu pozostają jednak w DNA, świadcząc o jego
pochodzeniu od genu, który był aktywny u przodka. Te martwe geny nazywają się pseudogenami.
Obecnie, kiedy sekwencjonowanie genomu jest sprawą rutynową, ewolucjoniści mogą
prowadzić poszukiwania martwych genów na dużą skalę. I, jak przewidziano, są wszędzie —
w każdym niemal zbadanym gatunku. Niektóre z tych „martwych" genów omawiałem w mojej
książce, włącznie z ludzkimi genami, które dawniej wytwarzały witaminę C lub żółtka jajeczek
u naszych przodków, ale obecnie są nieme. Trudno byłoby twierdzić, że te geny nadal mają
„funkcję", ponieważ w ogóle nie produkują żadnego białka. Ich istnienie jest potężnym argumentem
na korzyść ewolucji i przeciwko kreacjonizmowi.
(http://www.plosgenetics.org/article/info:doi/10.1371/journal.pgen.1000634)
w „PLoS Genetics" kontynuuje te poszukiwania przewidzianych martwych genów — tym razem
szuka genów, które kiedyś dawały emalię zębów — i znajduje mnóstwo tych wraków. Są dokładnie
tam, gdzie można się ich spodziewać — u bezzębnych zwierząt, o których od dawna uważano, że
pochodzą od zwierząt z zębami. Tak więc „teoria" ewolucji jest potwierdzona raz jeszcze, choć
doprawdy nie potrzebujemy więcej potwierdzeń. Ten artykuł jednak wychodzi poza jedynie zbędny
już dowód wspólnego pochodzenia. Autorzy tworzą model degeneracji „emalii" i dzięki wyliczeniom,
kiedy ta degeneracja zaszła, mogą przewidzieć, kiedy w zapisie kopalnym powinni zobaczyć albo
bezzębnych, albo zębatych przodków.
Istnieją dwa rodzaje ssaków, które nie mają emalii zębów: te, które są bezzębne (np.
pancerniki, pangoliny, mrówkojady, fiszbinowce), i te, które mają zęby, ale nie mają emalii (np.
kaszaloty małe, leniwce dwupalczaste). Na podstawie innych danych, włącznie ze skamielinami
i morfologią porównawczą, naukowcy z zaufaniem przewidzieli, że każdy z tych gatunków pochodzi
od przodków, którzy mieli emalię na zębach. U wielu gatunków ssaków naukowcy zsekwencjonowali
krytyczny gen enameliny (ENAM), gen, który pomaga odkładać hydroksyapatyt w zębach. Jeśli
wyłączy się ENAM u myszy, nie formuje się emalia.
Rzeczywiście, w każdym gatunku, który nie ma zębów albo emalii na zębach, ENAM był
niefunkcjonalny, albo z powodu akumulacji mutacji zmiany ramki odczytu, albo kodonów STOP.
Autorzy pokazują bardzo ładny wykres tego, jak geny degenerowały się w każdej linii rodowej, który
przedstawiam poniżej (pełen podpis znajduje się na końcu mojego tekstu, jeśli jesteś genetykiem)
(a tłumaczka zostawia go po angielsku, bo genetykiem nie jest. MK)
Racjonalista.pl
Strona 1 z 4
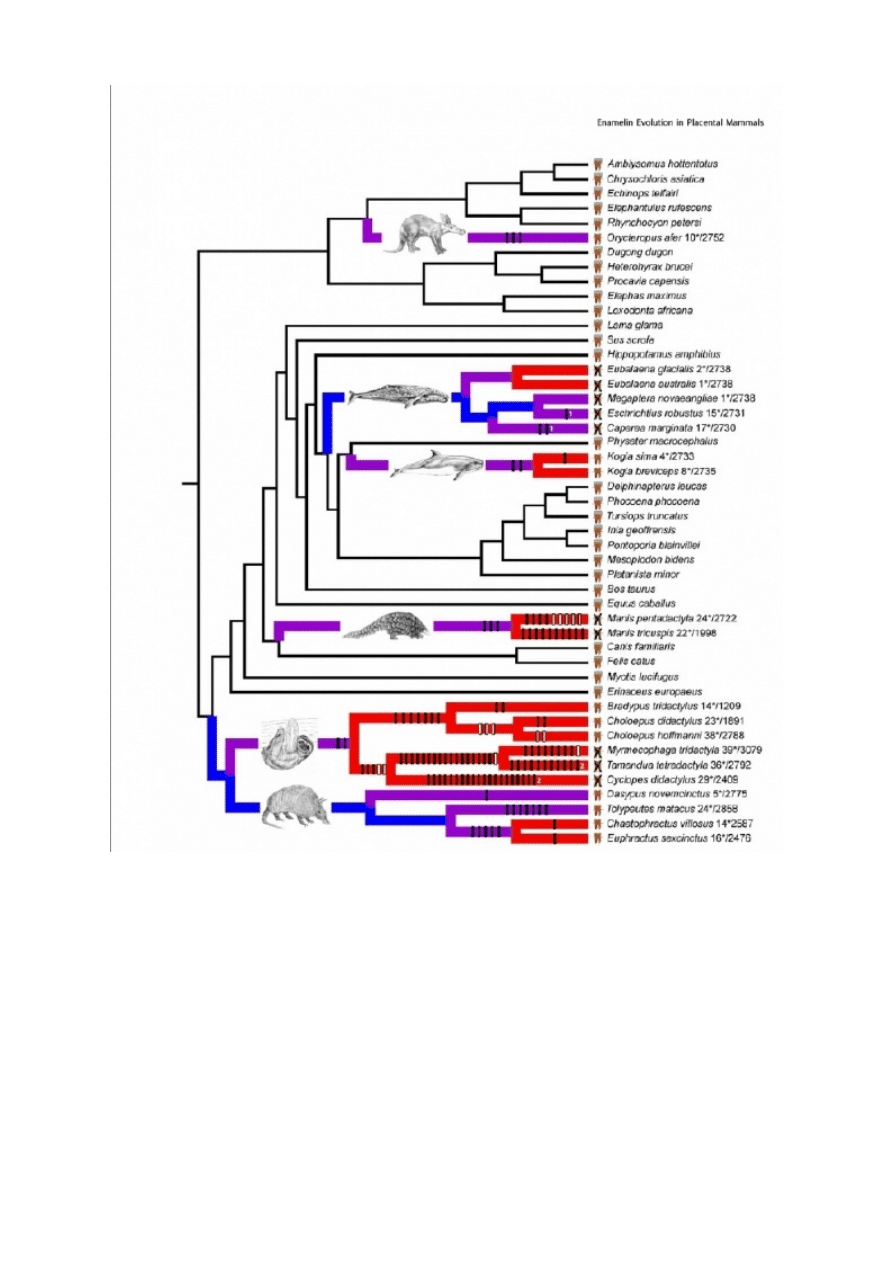
Filogeneza genu enameliny dla gatunków ssaków bez zębów, z zębami bez emalii lub
z zębami z emalią (typ zęba zilustrowany po prawej stronie, przed nazwą gatunku).
No cóż, to jest dowód na rzecz ewolucji i jest on elegancki. Ale doprawdy nie potrzebujemy
więcej dowodów tego typu, żeby poprzeć Darwina, ponieważ jest wiele danych o takich martwych
genach. Tym, co powoduje, że ta praca jest wyjątkowa, jest model autorów jak i kiedy ENAM był
dezaktywowany w wielu liniach rodowych.
Szczegóły są skomplikowane, ale zasadniczo autorzy modelowali dwustopniowy proces ewolucji
genu: przed mutacją dezaktywującą ENAM (której zajście można oszacować na podstawie drzewa
genu) i po niej. Zakłada się, że przed dezaktywacją gen ewoluował powoli, ponieważ zmiany genu
były na ogół szkodliwe. Po dezaktywacji gen ewoluował szybciej — uważa się, że ponieważ nie
wytwarzał już dłużej pożytecznego produktu, zmiany akumulowały się „neutralnie" (tj. wszystkie
zmiany ani nie powodowały selekcyjnej kary, ani nie dawały korzyści). Ta analiza stawia kilka
przewidywań. Jednym z nich jest, że pra-szczerbaki (pancerniki, leniwce i mrówkojady) miały zęby
z emalią, mimo że ich żywi reprezentanci ich nie mają. Jest to przewidywanie, które daje się
przetestować: znajdź wczesne „podstawowe" szczerbaki i zajrzyj im w zęby.
Jak zauważyliście, przyjęto założenie, że po dezaktywacji genu degenerował on się z powodu
akumulacji losowych, neutralnych mutacji. Jest to sensowne założenie dla dezaktywacji genu, ale nie

dla zniknięcia lub skurczenia się całej cechy. Jeśli cecha nie jest już dłużej użyteczna, są trzy
sposoby, na jakie może się degenerować:
1.Jak założono dla genów, przez akumulację dezaktywujących mutacji, które nie niosą żadnej
kary, skoro zachodzą w cesze w żaden sposób nie przyczyniającej się do rozmnażania.
2. Cecha może degenerować się przez pozytywny dobór naturalny. Jednym sposobem jest po prostu
to, że na zwierzę (lub roślinę) można patrzeć jak na ekonomiczny kompromis między biologicznymi
materiałami i jeśli nie potrzebujesz jakiejś cechy, dobór może faworyzować zamianę jej na cegiełki
innych cech. Na przykład, materiał używany do zbudowania skrzydła nielotnego ptaka może być
skierowany na tworzenie lub wzmocnienie kości w innych częściach ciała.
3. Podobnie, cechy, które nie są już dłużej użyteczne, mogą łatwo ulegać uszkodzeniom. Dobór
będzie wtedy faworyzował degenerację tej cechy w taki sposób, by zapobiec uszkodzeniu. To na
przykład może wyjaśnić redukcję oczu fauny jaskiniowej lub zwierząt drążących pod ziemią, takich
jak kret.
Meredith RW, Gatesy J, Murphy WJ, Ryder OA, Springer MS (2009). Molecular decay of the
tooth gene enamelin (ENAM) mirrors the loss of enamel in the fossil record of placental mammals.
PLoS Genet 5(9): e1000634. doi:10.1371/journal.pgen.1000634
Podpis pod wykres 1 (powyżej), z Meredith i in.: Figure 1. Species tree with
frameshift mutations and dN/dS branch coding. Symbols next to taxon names denote
taxa having teeth with enamel, taxa having teeth without enamel, and edentulous
taxa. Branches are functional (black), pre-mutation (blue), mixed (purple), and
pseudogenic (red). Vertical bars on branches represent frameshift mutations (see
Table S1). Frameshifts that map unambiguously onto branches are shown in black.
Frameshifts shown in white are unique, but occur in regions where sequences are
missing for one or more taxa (Figure S7) and were arbitrarily mapped onto the
youngest possible branch. Homoplastic frameshifts (deltran optimization) are marked
by numbers. Numbers after taxon names indicate the minimum number of stop
codons in the sequence (before slashes) and the length of the sequence (after
slashes).
Why Evolution is true, 10 września 2009
Jerry Coyne
Profesor na wydziale ekologii i ewolucji University of Chicago,
niedawno wydawnictwo Viking wydało jego książkę pt: Why
Evolution is True.
(Publikacja: 14-09-2009 Ostatnia zmiana: 15-09-2009)
(http://www.racjonalista.pl/kk.php/s,6787)
Contents Copyright
©
2000-2009 Mariusz Agnosiewicz
Programming Copyright
©
2001-2009 Michał Przech
Autorem portalu Racjonalista.pl jest Michał Przech, zwany niżej Autorem.
Właścicielami portalu są Mariusz Agnosiewicz oraz Autor.
Żadna część niniejszych opracowań nie może być wykorzystywana w celach
komercyjnych, bez uprzedniej pisemnej zgody Właściciela, który zastrzega sobie
niniejszym wszelkie prawa, przewidziane
w przepisach szczególnych, oraz zgodnie z prawem cywilnym i handlowym,
Racjonalista.pl
Strona 3 z 4

w szczególności z tytułu praw autorskich, wynalazczych, znaków towarowych
do tego portalu i jakiejkolwiek jego części.
Wszystkie strony tego portalu, wliczając w to strukturę katalogów, skrypty oraz inne
programy komputerowe, zostały wytworzone i są administrowane przez Autora.
Stanowią one wyłączną własność Właściciela. Właściciel zastrzega sobie prawo do
okresowych modyfikacji zawartości tego portalu oraz opisu niniejszych Praw
Autorskich bez uprzedniego powiadomienia. Jeżeli nie akceptujesz tej polityki możesz
nie odwiedzać tego portalu i nie korzystać z jego zasobów.
Informacje zawarte na tym portalu przeznaczone są do użytku prywatnego osób
odwiedzających te strony. Można je pobierać, drukować i przeglądać jedynie w celach
informacyjnych, bez czerpania z tego tytułu korzyści finansowych lub pobierania
wynagrodzenia w dowolnej formie. Modyfikacja zawartości stron oraz skryptów jest
zabroniona. Niniejszym udziela się zgody na swobodne kopiowanie dokumentów
portalu Racjonalista.pl tak w formie elektronicznej, jak i drukowanej, w celach innych
niż handlowe, z zachowaniem tej informacji.
Plik PDF, który czytasz, może być rozpowszechniany jedynie w formie oryginalnej,
w jakiej występuje na portalu. Plik ten nie może być traktowany jako oficjalna
lub oryginalna wersja tekstu, jaki zawiera.
Treść tego zapisu stosuje się do wersji zarówno polsko jak i angielskojęzycznych
portalu pod domenami Racjonalista.pl, TheRationalist.eu.org oraz Neutrum.eu.org.
Wyszukiwarka
Podobne podstrony:
Biofilm, tworzenie płytki bakteryjnej na powierzchni zębów
Materiały na wypełnienia zębów tylnych 2
Nowe geny na chromosomie Y
po stresach-cw, Metoda indygokarminowa - barwnik indygokarminowy barwi martwe tkanki na niebiesko, n
Materiały na wypełnienia zębów tylnych
Ballada o martwej naturze na emeryturze
Materiały na wypełnienia zębów tylnych
Biofilm, tworzenie płytki bakteryjnej na powierzchni zębów
Identyfikacja sprawcy zabójstwa na podstawie śladów zębów na ciele ofiary
genetyka, ćw 6 geny, 6 Techniki oparte na PCR do diagnozowania chorów genetycznych i uchwycenia zmie
Przepisy na naturalne pasty do zębów
Geny wczesnych odpowiedzi na auksynę
Przebarwienia zębów i czynniki na nie wpływające
Przepisy na naturalne pasty do zębów
Drobnoustroje znajdujące się na szczoteczkach do zębów
Identyfikacja sprawcy zabójstwa na podstawie śladów zębów na ciele ofiary
Chiny Nowy biznes – martwe dzieci przerobione na tabletki
więcej podobnych podstron