2007 by nixon.SUM@gmail.com
1
RECEPTORY cz1
receptory to wyspecjalizowane struktury nerwowe, których podstawową funkcją jest odbieranie
i kodowanie informacji
przyjmujemy z otoczenia ok 10
9
bitów informacji (które są następnie obrabiane, część
przechodzi przez CNS) a generujemy „do” otoczenia ok 10
7
bitów (mowa, ruch etc.)
PRAWO SWOISTEJ ENERGII MULLERA - każdy receptor jest wyspecjalizowany w
odbiorze określonego rodzaju bodźca, choć może odpowiadać na działanie innego,
nieadekwatnego bodźca, jeśli jego siła jest dostatecznie duża (np. uderzenie w tył głowy daje
efekt w postaci jasnych plam przed oczami)
ślad pamięciowy
poziom świadomości
droga czuciowa
receptor
świat
Przetwarzanie energii bodźca zachodzi dwufazowo („analogowo i cyfrowo”)
Potencjał generujący powoduje miejscowy przepływ prądu i po osiągnięciu pewnej wielkości
zwanej potencjałem progowym wyzwala w pierwszej cieśni węzła nerwu dośrodkowego potencjał
czynnościowy, który jako pojedynczy impuls lub salwa dociera do CNS. Siła bodźca jest
zakodowana w częstotliwości pot. czynnościowych.
przetwornik
bodziec
generator iglic
amplituda
odkształceń
bodziec
(um)
potencjał -50
(mV)
potencjał generujący (lub receptorowy) – przetwarzanie „analogowe”
-80
potencjał +30
(mV)
salwa potencjałów iglicowych (czynnościowych)
-80

2007 by nixon.SUM@gmail.com
2
Klasyfikacja receptorów:
1) Mechanoreceptory (dotyk, ucisk, wibracja)
2) termoreceptory (temperatura)
3) nocyceptory (bólowe)
4) chemoreceptory
5) fotoreceptory
Klasyfikacja wg Sherringtona:
1) interoreceptory
2) eksteroreceptory
3) telereceptory
4) proprioreceptory
Adaptacja
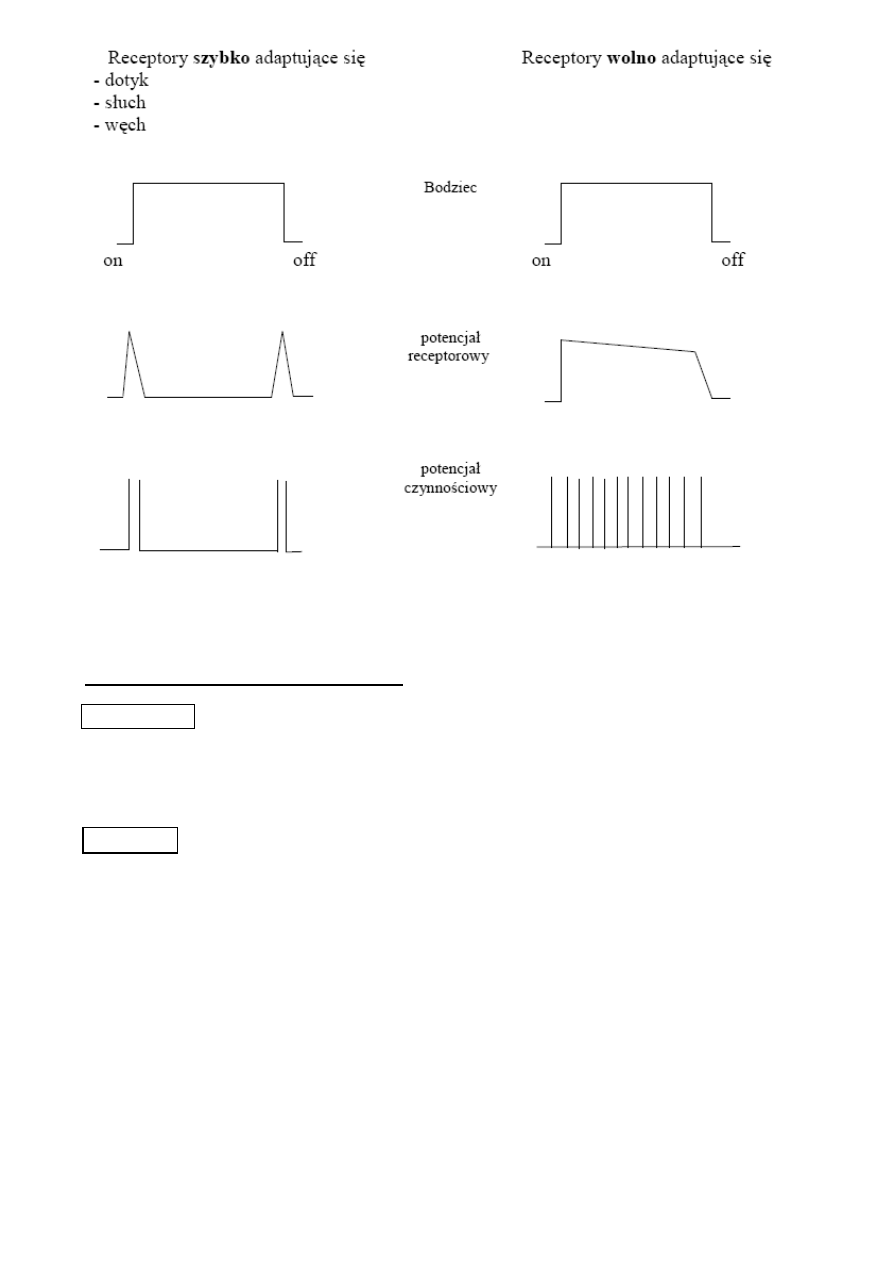
Każdy receptor charakteryzuje się dodatkowo szybkością adaptacji, możemy zatem wyróżnić te,
które
adaptują się wolno (toniczne, statyczne) – wysyłają one impulsy przez cały czas trwania
bodźca
adaptują się szybko (fazowe, dynamiczne)- wysyłają impulsy ze zmniejszającą się
częstotliwością mimo działania bodźca
Ogólnie rzecz ujmując adaptacja jest to obniżenie odpowiedzi receptora pomimo stałej obecności
bodźca. Co ma takie działanie na celu?
1) adaptacja czuciowa dostosowuje częstotliwość odpowiedzi do szybkości narastania i
stosowania bodźca
1. w przypadku receptorów szybko adaptujących się częstość wyładowań wzrasta,
jeśli wzrasta częstość stosowania bodźca – czyli nasz receptor zostanie
pobudzony tylko wtedy jeśli dany bodziec będzie działa dostatecznie szybko
(zapobiega to wytwarzaniu potencjału w odpowiedzi na wolno narastające
bodźce)
2. w przypadku receptorów wolno adaptujących się wyładowania występują ze
stałą częstotliwością tak długo, jak długo działa bodziec
2) w pewnych okolicznościach zmniejsza ilość informacji które dochodzą do mózgu.
Mechanizmy adaptacji:
1) w pierwszym przypadku mechanizm przetwarzania energii jest niezdolny do podtrzymania
potencjału generacyjnego pomimo stałego stosowania bodźca
2) w drugim przypadku generator potencjałów iglicowych staje się niezdolny do utrzymania
ciągu potencjałów – może to być spowodowane np. wzrostem przewodności błony dla
jonów K+, wzrostem aktywności Na/K-ATPazy lub inaktywacją kanałów dla Na
2007 by nixon.SUM@gmail.com
3
warto pamiętać, że receptory odpowiedzialne za podstawowe parametry organizmu nie posiadają
zdolności adaptacji (pH, ból itp)
Intensywność, lokalizacja i jakość bodźca
Intensywność bodźca jest kodowana przez częstotliwość potencjałów czynnościowych oraz przez
liczbę zaktywowanych neuronów czuciowych. Częstotliwość ta jest proporcjonalna do wielkości
potencjału generacyjnego. Jeśli wzrasta intensywność bodźca to również wzrasta wielkość
potencjału generacyjnego (patrz wykres str 7 – siła bodźca jest proporcjonalna do wielkości
potencjału generacyjnego)
Lokalizacja bodźca odbywa się głównie w korze mózgu i jest nazywany prezentacją topograficzną.
Dodatkowo możemy mówić o:
1) polu recepcyjnym – każdy neuron czuciowy otrzymuje inf. z określonego pola czuciowego
nazywanego polem recepcyjnym. Im mniejsze jest pole tym precyzyjniejsze jest kodowanie
lokalizacji bodźca (najmniejsze pole jest w dołku środkowym oka, podobnie na opuszkach
palców. Na rękach i plecach ciężko umiejscowić punkt mechanicznej stymulacji)
2) hamowaniu obocznemu, dzięki któremu lokalizacja bodźca może być bardziej precyzyjna
bez hamowania obocznego dwa bodźca są rozpoznawane jako oddzielne jedynie wtedy, gdy
działają na dwa pola recepcyjne oddzielone od siebie przez niepobudzone pole
2007 by nixon.SUM@gmail.com
4
definicje:
jednostka czuciowa – 1 neuron i wszystkie receptory które unerwia
pole recepcyjne – obszar z którego informacja czuciowa jest odbierana przez jedno włukno
Jakość bodźca jest kodowana przez:
1) mechanizm oznaczonej linii - bodziec jest zakodowany przez określoną drogę neuronalną,
która jest pobudzana. Dlatego drażnienie drogi w dowolnym miejscu lokalizowane jest w
receptorze (tam skąd „powienien pochodzić” dany bodziec – np. bóle kończyn
fantomowych). Nosi to nazwę Prawa Projekcji
2) bardziej złożony jest mechanizm wykorzystujący wzorzec aktywności w obrębie drogi
nerwowej przenoszącej inf. do mózgu
1. wzorzec kodowania czasowego – ten sam neuron może przewodzić dwa rodzaje
informacji w zależności od tego czy są one w postaci salw potencjałów
czynnościowych czy w sposób ciągły
2. wzorzec kodowanie przestrzennego – np. trzy neurony mogą być potrzebne do
zakodowanie różnych wrażeń smakowych – jeśli pobudzimy wszystkie poczujemy
smak kwaśny, a jeśli tylko dwa poczujemy smak słony
Czucie Skórne
Skóra zawiera receptory przystosowane do odbierania dotyku, bólu i temperatury. W większości są
to włókna bez osłonki mielinowej, przenoszące surowe somatosensoryczne wrażenia mechaniczne.
Chociaż znacznie mniej liczne, grube, zmielinizowane włókna czuciowe kodują ważne informacje
dotyczące czucia dotyku,wibracji i ciśnienia. Czucie temp i bólu jest kodowane przez cienkie
zmielinizowane włókna Aδ i niezmielinizowane włókna C
2007 by nixon.SUM@gmail.com
5
mechanoreceptory:
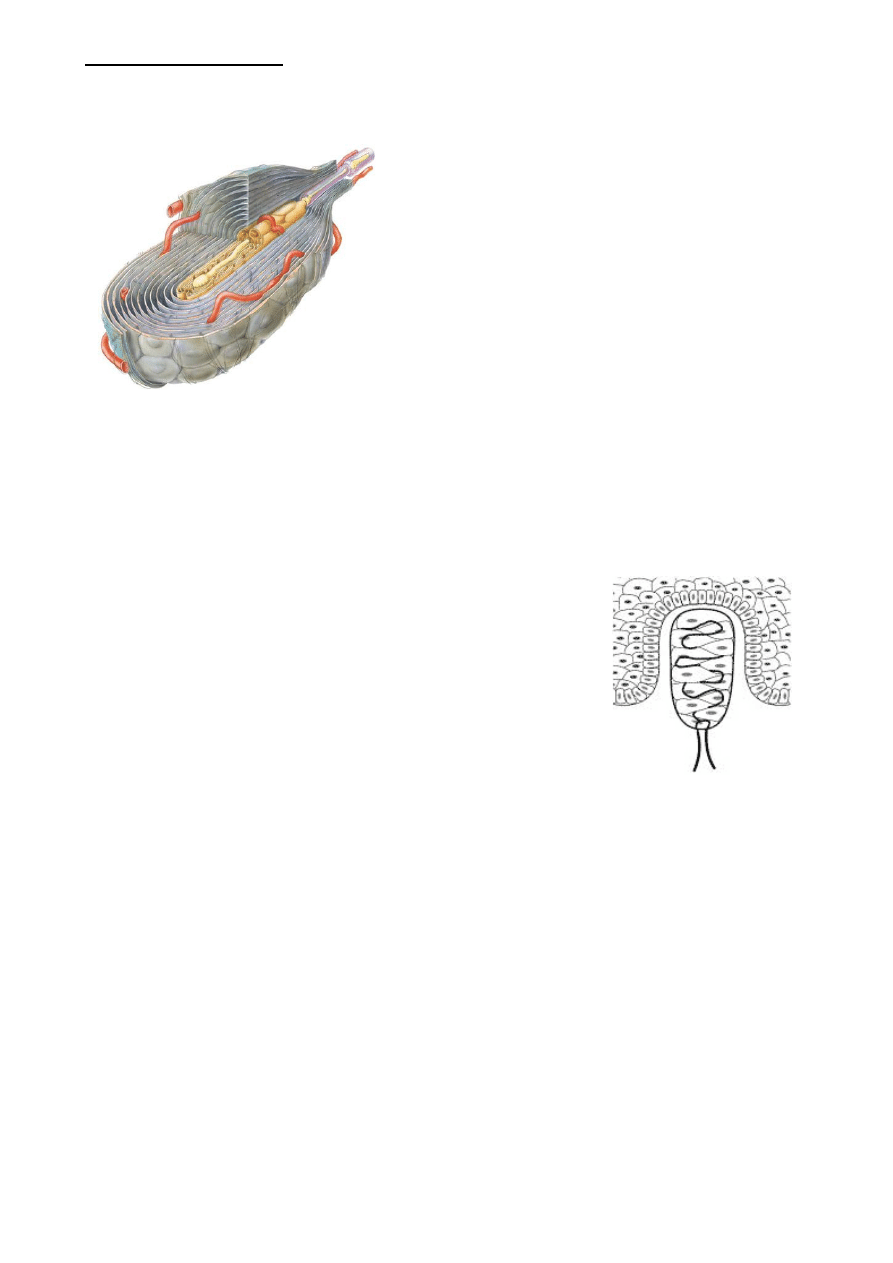
1) ciałko Paciniego = dotyk i wibracje
szybko adaptujące się
koduje czucie wibracji
kiedy bodziec mechaniczny odkształca cebulowatą
torebkę, to odkształcenie jest przenoszone na
zakończenie nerwowe i powoduje depolaryzację.
Wielkość potencjału generacyjnego jest proporcjonalna
do wielkości odkształcenia.
bodziec wibracyjny powoduje stałe wyładowania
ciałka. Za każdym razem kiedy bodziec zanika i
ponownie się pojawia jest generowany potencjał
występuje tu zjawisko adaptacji (wynika to z
elastyczności torebki)
jedynie silny bodziec może skutecznie przenieść deformację do rdzenia torebki
częstotliwość wyładowań koduje częstotliwość wibracji (a nie jej intensywność)
intensywność jest kodowana przez liczbę potencjałów czynnościowych generowanych przez
każdą deformację a także przez liczbę pobudzonych ciałek
2) Ciałko Meissnera = częstość
szybko adaptujące się
małe pole recepcyjne
informuje o częstości stosowania bodźca
3) Ciałko Merkela
wolno adaptujące się
małe pole recepcyjne
służy do kodowania lokalizacji bodźca
przetwornikiem energii bodźca nie jest zakończenie nerwowe lecz są nim komórki nabłonka
tworzące tarczkę
2007 by nixon.SUM@gmail.com
6
4) Ciałko Ruffiniego
zakończenie włókna otoczone torebką
kolagenową wypełnioną płynem
duże pole recepcyjne – każde
odkształcenie lub rozciągnięcie skóry
powoduje depolaryzację
wolno adaptujący się receptor
Termoreceptory:
czucie temperatury jest kodowane przez termoreceptory występujące w zakończeniach małych
włókien osłoniętych mieliną ( Aδ ) i włókien bez osłonki - C. Kodowanie ciepła i zimna odbywa
się przez osobne receptory z oddzielnymi polami recepcyjnymi
Odbieranie ciepła
receptory wrażliwe na wzrost temperatury skóry w zakresie 38-44 C
odbierają różnicę temp. Rzędu 0,001 C/sek.
Jest ich 4-10 razy mniej niż receptorów zimna
jeśli temperatura wzrośnie powyżej 45 dochodzi do jednoczesnego pobudzenia receptorów
zimna i nocyceptorów (tzw. „paradoks uczucia chłodu”)
szybko się adaptują
Odbieranie zimna
wrażliwe na spadek temperatury skóry w zakresie 27-38 C
odbierają tempo zmian rzędu 0,004 C/sek
jeżeli temperatura spadnie poniżej 25 C wyładowania stopniowo zanikają, a dalszy spadek
temp. działa anestetyczne (przeciwbólowo)
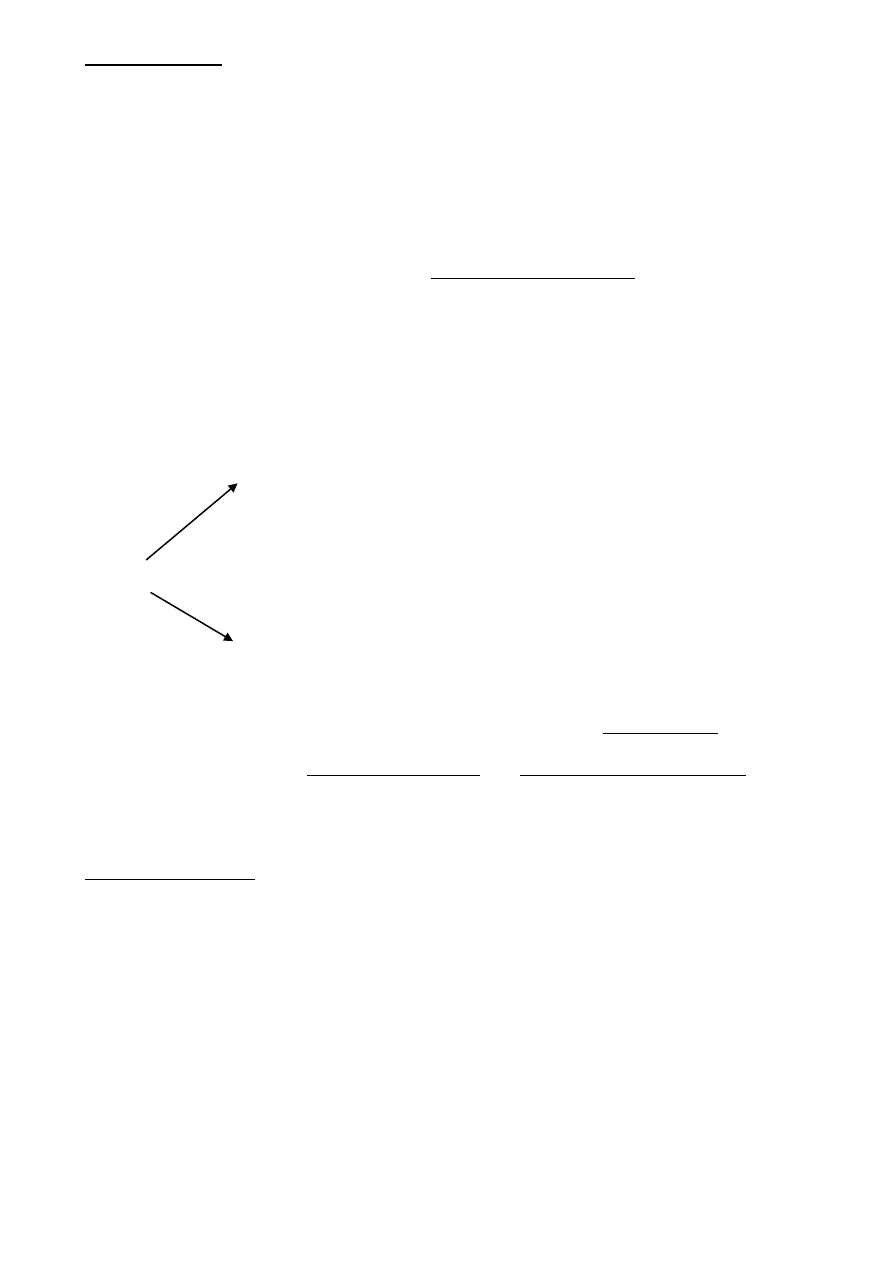
Występuje 6 różnych kanałów Trp odpowiedzialnych za wrażliwość na różne zakresy temperatury
Kanały dla kationów, rodzina białek TRP
(transient receptor potential):
1. TRPV1 – aktywacja 43
o
C i kapsaicyna
(papryka chili)
2. TRPV2 – aktywacja 53
o
C
3. TRPV3 – aktywacja 33
o
C
4. TRPM8 – aktywacja 25
o
C i mentol
5. TRPA1 – aktywacja 17
o
C
Niektóre receptory zimna posiadają również Trp1 wrażliwe na T>43 C – „paradoksalne uczucie
chłodu”
2007 by nixon.SUM@gmail.com
7
Nocyceptory:
receptory bólu znajdują się na wolnych zakończeniach nerwowych cienkich włókien Aδ lub C
bodźce adekwatne dla nocyceptorów to każdy rodzaj bodźca o odpowiednio dużej sile (np.
bodziec cieplny 100x silniejszy od bodźca adekwatnego dla termoreceptora.
Nie adaptują się
Rozmieszczenie receptorów :
skóra
błony śluzowe i surowicze
rogówka
torebki narządów miąższowych (nie bolą narządy miąższowe tylko ich torebki)
przewody wyprowadzające
opony mózgowia (nie boli nas głowa tylko zazwyczaj albo naczynia albo opony)
okostna, naczynia, mięśnie
BRAK w śluzówce przewodu pokarmowego, w mózgu i na pow. stawowych
Dwa rodzaje czucia bólu wynikają z zastosowania silnego szkodliwego bodźca na skórę:
szybki (ostry)
– małe pola recepcyjne
- impulsy przewodzone przez Aδ grupy III (15-30m/s)
- dochodzą do kory SI przez szlak rdzeniowo-wzgórzowy i są
reprezentowane topograficznie
ból
wolny (rozlany)
– impulsy przewodzone przez włókna C grupy IV
- docierają do kory SII przez układ
rdzeniowo-siateczkowo-wzgórzowy
- kolaterale tego układu przechodzą przez twór siatkowaty pobudzając
włókna szlaku, który wywołuje wrażenia emocjonalne,
pokrywę śródmózgowia oraz istotę szarą okołowodociągową
- Drogi te są odpowiedzialne za intensywne, nieprzyjemne odbieranie
powolnego bólu
Mechanizm działania:
bodziec uszkadzający uwalnia z tkanek histaminę i ciała histaminopodobne oraz bradykininę
powstającą w tkankach pod wpływem enzymów proteolitycznych zwanych kalikreinami
tkankowymi, które z kolei odszczepiają z globulin tkankowych (czyli kininogenów) kininy, głównie
wspomnianą bradykininę.
Zarówno bradykinina jak i histamina depolaryzują nagie zakończenia nerwowe
bradykinina działa na tkanki za pośrednictwem receptorów B1 i B2.
pod wpływem kinin dochodzi do miejscowego skurczu trzewnych mięśni gładkich, rozkurczu
naczyń, wzrostu przepuszczalności kapilarów i infiltracji neutrofilów.
W mechanizmie bólów mięśniowych dużą rolę przypisuje się niedokrwieniu i niedotlenieniu z
następowym nagromadzeniem produktów metabolizmu beztlenowego
2007 by nixon.SUM@gmail.com
8
Drogi przewodzenia bólu:
- z kończyn i tułowia
droga rdzeniowo – wzgórzowa boczna
WZGÓRZE (jądro brzuszne tylno – boczne)
RAS
SI
ze wzgórza impulsy biegną do okolic czuciowych pierwszorzędowych kory (SI - pola 1, 2, 3
Brodmanna), które odpowiadają za lokalizację bodźca bólowego.Oboczne połączenia do układu
siatkowatego RAS (jąder śródblaszkowych wzgórza), a także do istoty szarej środkowej
śródmózgowia
- z obszaru twarzy ból przez NC V do jądra brzusznego tylno – przyśrodkowego i stamtąd
do kory
- droga rdzeniowo – siatkowata
wielosynaptyczna sieć w rdzeniu kręgowym, skąd impulsacja przechodzi przez pień do jąder
przyśrodkowych i śródblaszkowych wzgórza a stamtąd do całej kory => reakcja wzbudzenia
w korze mózgowej
Istota szara środkowa śródmózgowia
- otrzymuje połączenia z obu dróg:
rdzeniowo – wzgórzowej bocznej
rdzeniowo – siatkowatej
- kluczowa dla skoordynowanej reakcji wegetatywnej organizmu na ból
ból głęboki
ból powierzchowny
aktywność ruchowa
wokalizacja (krzyk)
Natężenie czucia bólu zależy od ilości impulsów i od wrażliwości ośrodków czucia i bólu. Obie
te zależności są regulowane przez:
1) bramkę kontrolną na poziomie rogów tylnych rdzenia kręgowego
– hamowanie między 1 i 2 neuronem
(przez interneurony w obrębie substantia gelatinosa)
do mózgu
grube włókna
dotyk, ucisk
(rogi tylne)
I
-
+
ból

2007 by nixon.SUM@gmail.com
9
2) bramkę kontrolną na poziomie jąder wzgórza – hamowanie między 2 i 3 neuronem
(hamowanie pre i post synaptyczne)
3) okołowodociągową istotę szarą śródmózgowia
wydzielają opioidy endogenne
4) okołokomorową część wzgórza
5) istnienie systemu włókien eferentnych z pnia do rdzenia kręgowego, które hamują
przewodnictwo miedzy 1. i 2. neuronem
występują 3 typy opiatów:
- endorfiny klasy α, β, γ - pochodzą od POMC
-
enkefaliny pochodzą od proenkefaliny występują w 2 rodzajach:
o enkefalina metioninowa
o enkefalina leucynowa
-
dynorfiny pochodzą z prodynorfiny
2 sposoby działania:
1. wzrost przepuszczalności błony dla K
+
i hiperpolaryzacja
2. receptory metabotropowe – aktywacja G
i
=> cAMP↓
Tłumią ból przez receptory:
-
mi (μ)
-
delta (δ)
-
kappa (κ) (dynorfina)
Morfina wiąże się z receptorami mi
-
z μ
1
– efekt przeciwbólowy
-
z μ
2
– depresja oddechowa
Endorfiny
-
wywołują uspokojenie
-
zwężenie źrenic
-
spadek aktywności ukł. oddechowego
Ból trzewny
- do rdzenia kręgowego dochodzi impulsacja przez korzenie grzbietowe
- w obrębie rogu tylnego wszystkie te włókna przełączają się na II neuron w drodze
rdzeniowo – wzgórzowej bocznej
- biegną dalej do wzgórza i do kory czuciowej (pola 1-3), czyli tam, gdzie odbierane jest
czucie eksteroceptywne
- zbieżność tych włókien w rogu tylnym jest podstawą do wytłumaczenia tzw. „bólu
przeniesionego”
Ból z narządów wewnętrznych może powodować:
- przeczulicę skóry
- obronę mięśniową
Przewodzenie bólu po specyficznych drogach bólowych jest zachowane w czasie:
- głębokiej narkozy
- snu
(
endorfina, enkefalina)
2007 by nixon.SUM@gmail.com
10
Ból ścienny (parietalny)
- w odróżnieniu od trzewnego możemy go ściśle lokalizować
- pochodzi z podrażnienia opłucnej, otrzewnej lub nasierdzia
przykład:
kiedy boli nas wyrostek odbieramy rozlany ból trzewny, dopiero jeśli zapalenie przejdzie na
sąsiednią otrzewną, otrzymamy ostry ból ścienny
Ból odniesiony
- ból trzewny jest często odczuwany w częściach ciała odległych od tych narządów (np. serce boli
w okolicach barku i brody)
- teorie wyjaśniające to zjawisko mówią o konwergencji i torowaniu bólu trzewnego odbitego z
narządów trzewnych do skóry tych samych dermatonów (rogi tylne odbierają jednocześnie
impulsację bólową z receptorów skóry oraz z interoreceptorów narządów wewnętrznych
(konwergencja). W ten sposób sygnały bólowe z narządów trafiają do tych samych neuronów II
rzędu w rdzeniu. Fakt że ból odniesiony promieniuje zawsze w jednym kierunku (z obszaru
trzewnego na powierzchnię skóry) dowodzi dominacji somatycznej impulsacji aferentnej nad
trzewną
Ból projekcyjny
występuje w wyniku bezpośredniego drażnienia włókien nerwowych w drodze
(np. ból fantomowy)
Czucie trzewne
receptory:
1. wolne zakończenia nerwowe w błonie śluzowej oraz surowiczej
2. ciałka blaszkowate w krezce, wrażliwe na mechaniczne odkształcenia
3. baroreceptory w zatoce szyjnej i łuku aorty
4. chemoreceptory w kłębkach szyjnych i aortalnych
5. receptory smakowe
unerwienie czuciowe:
Głowa
NC VII, IX, X
narządy trzewne klatki piersiowej
1) biegnące we wł. współczulnych nerwów:
- trzewny,
- sercowy,
- płucny
Th1-L1
2) we wł. przywspółczulnych (głównie X)
narządy miednicy
1) wł. aferentne do S2-S4 przez:
- nerwy miednicze
2) splot podbrzuszny
nerwy trzewne
Th1-L1
Wyszukiwarka
Podobne podstrony:
CWICZENIA, Ćwiczenia 3 - 04.03.05 - Receptory, 04
Z blogu Kataryny Na łasce Anodiny cz1# 05 2010
Receptura 05
TPL WYK 13 04 05 Obliczenia recepturowe
GW Wyklad 05 cz1
TPL PRAC 13 05 13 Lek recepturowy krople
2016 01 01 pismio MZ do NIA dotyczaca kopii recept 2015 05 18
podrecznik 2 18 03 05
regul praw stan wyjątk 05
05 Badanie diagnostyczneid 5649 ppt
Podstawy zarządzania wykład rozdział 05
05 Odwzorowanie podstawowych obiektów rysunkowych
05 Instrukcje warunkoweid 5533 ppt
05 K5Z7
więcej podobnych podstron