
ELONGACJA
TRANSLACJI
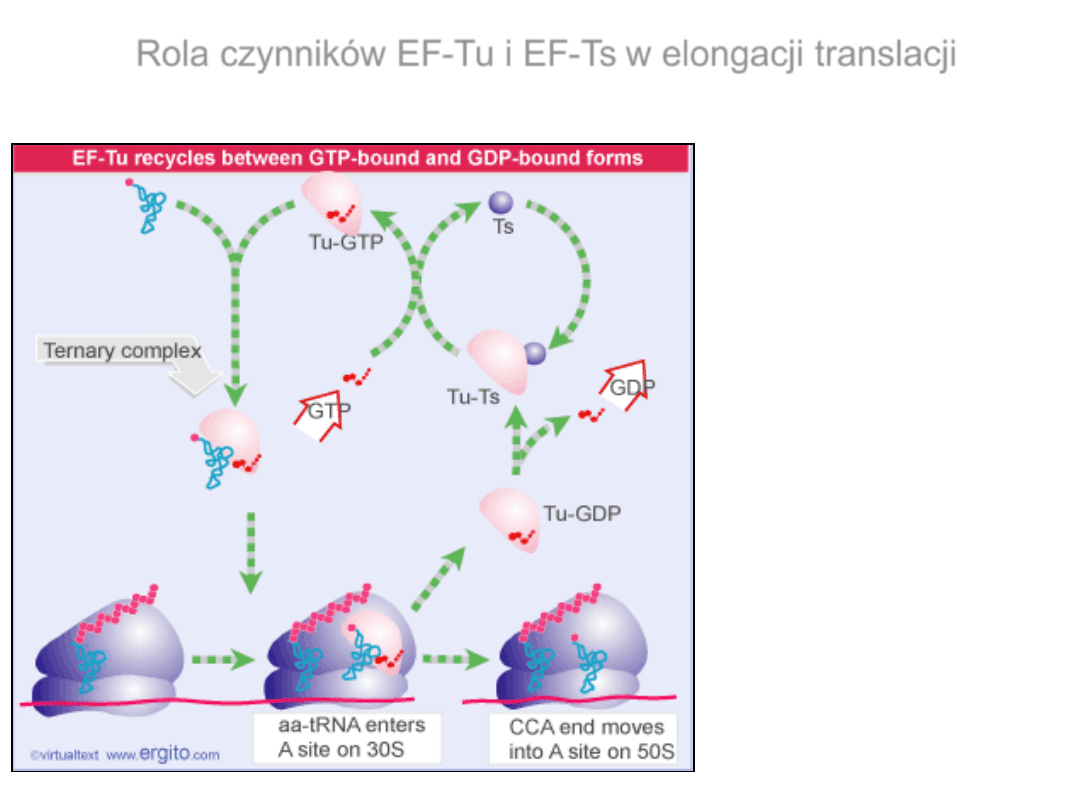
EF-Tu – bakteryjny czynnik
elongacyjny wiążący
aminoacylo-tRNA i
przenoszący je w miejsce A
rybosomu
EF-Ts –
czynnik powodujący
przekształcenie nieaktywnego
EF-Tu
●GDP w aktywny EF-
Tu
●GTP
eEF1
α – eukariotyczny
analog EF-Tu
eEF1
βγ -eukariotyczny
analog EF-Ts
Rola czynników EF-Tu i EF-Ts w elongacji translacji
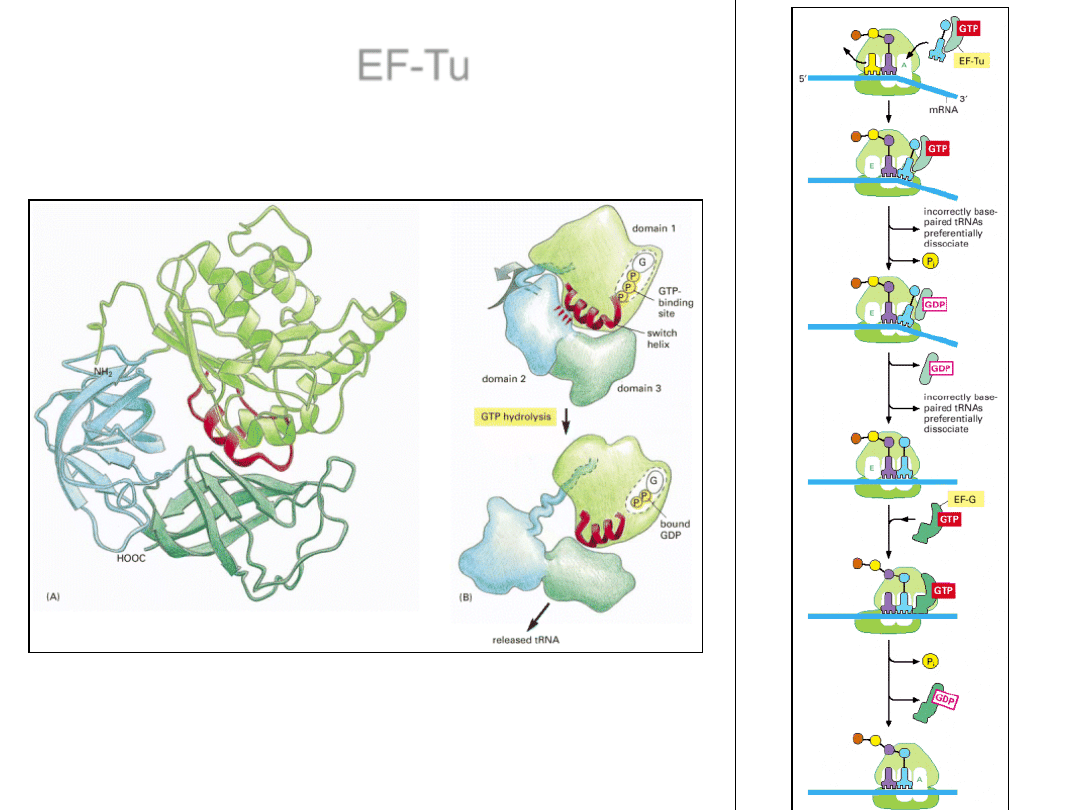
EF-Tu
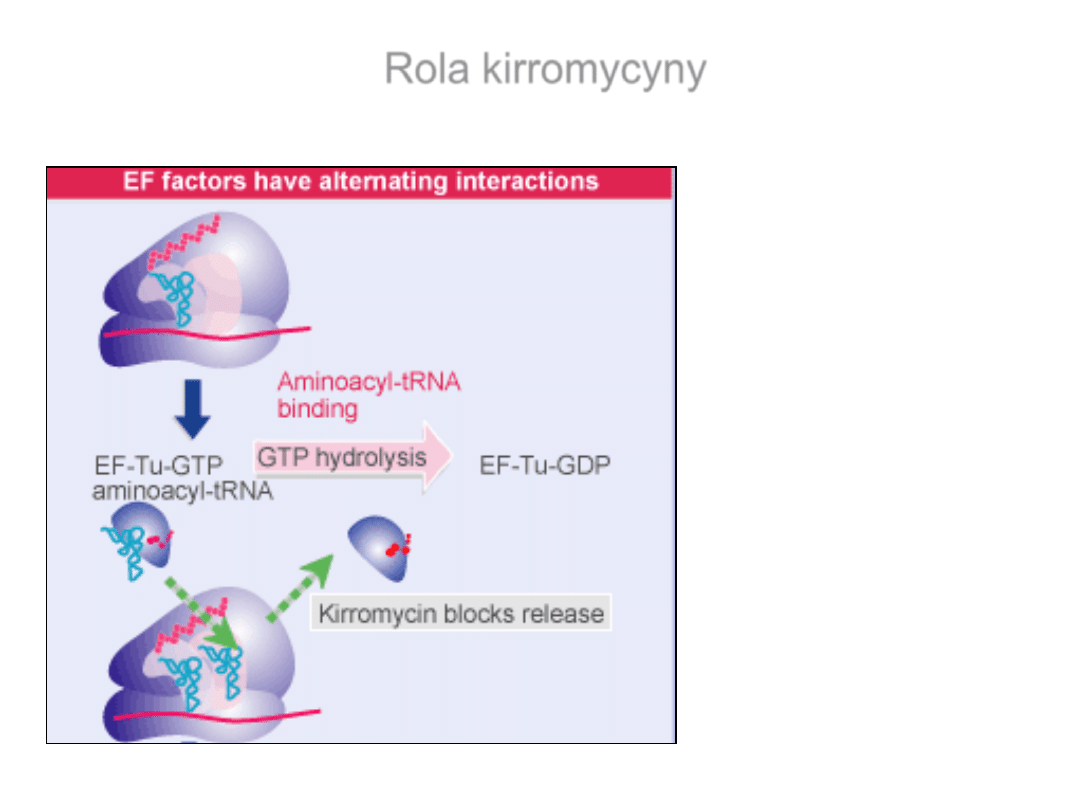
Kirromycyna- blokuje
funkcje EF-Tu
Rola kirromycyny
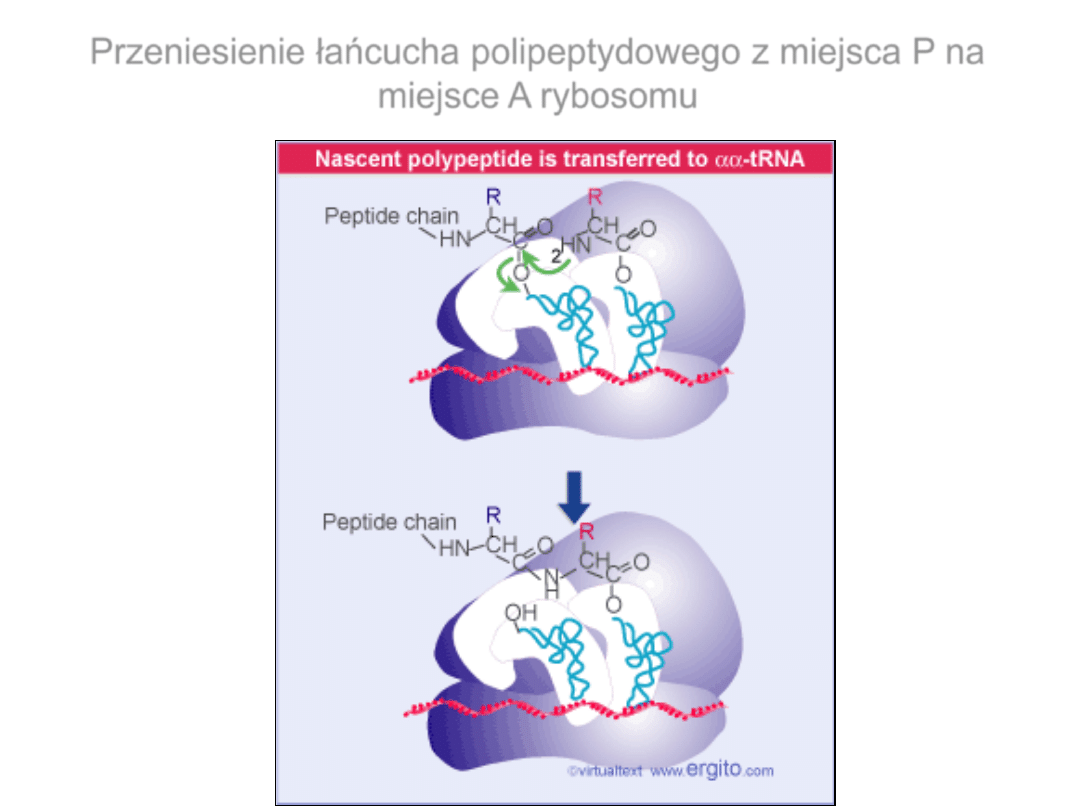
Przeniesienie łańcucha polipeptydowego z miejsca P na
miejsce A rybosomu
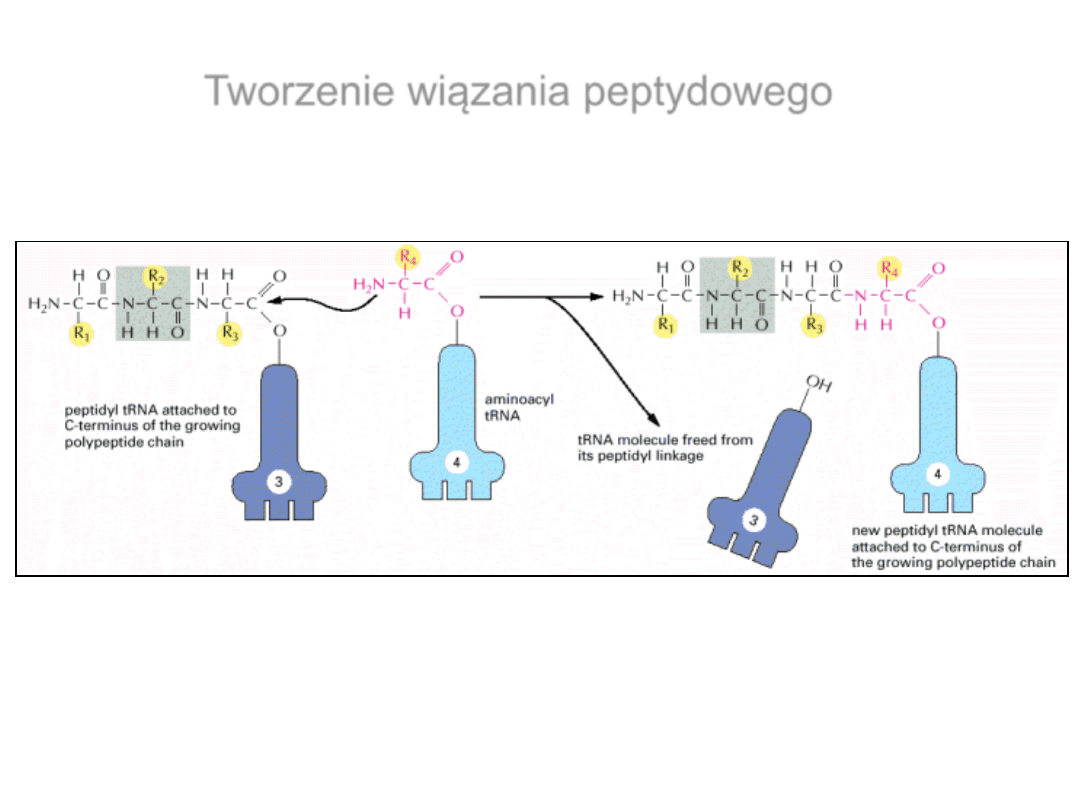
Tworzenie wiązania peptydowego
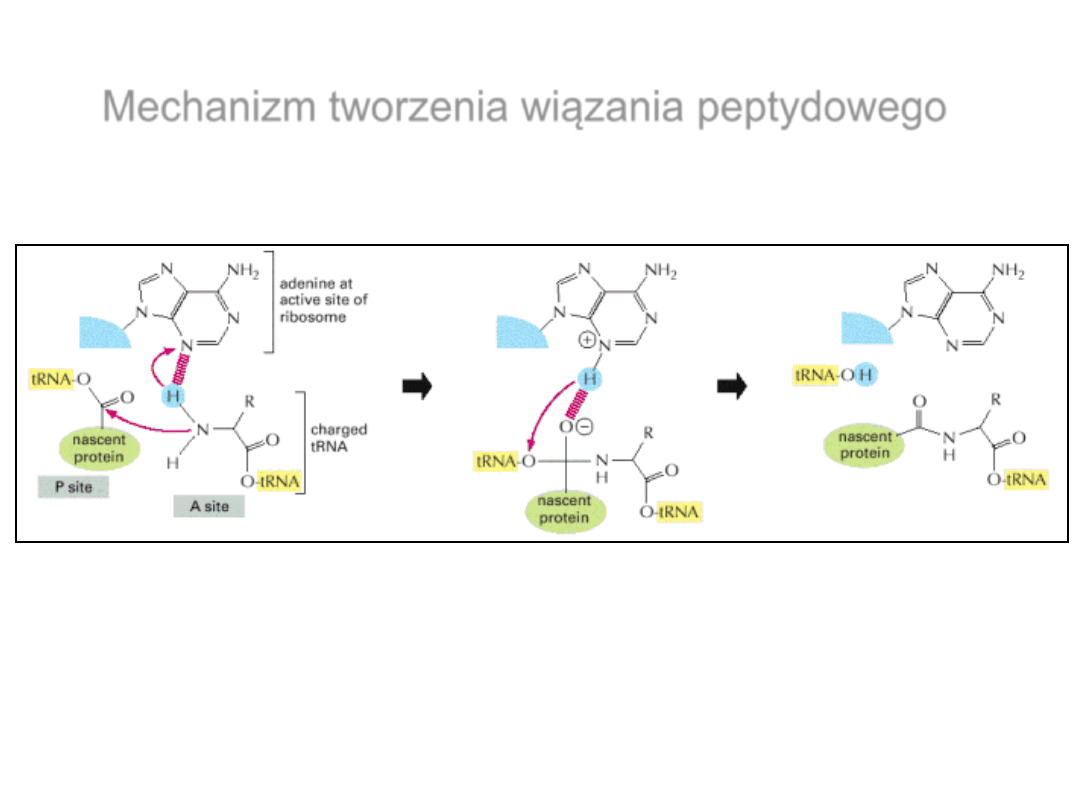
Mechanizm tworzenia wiązania peptydowego
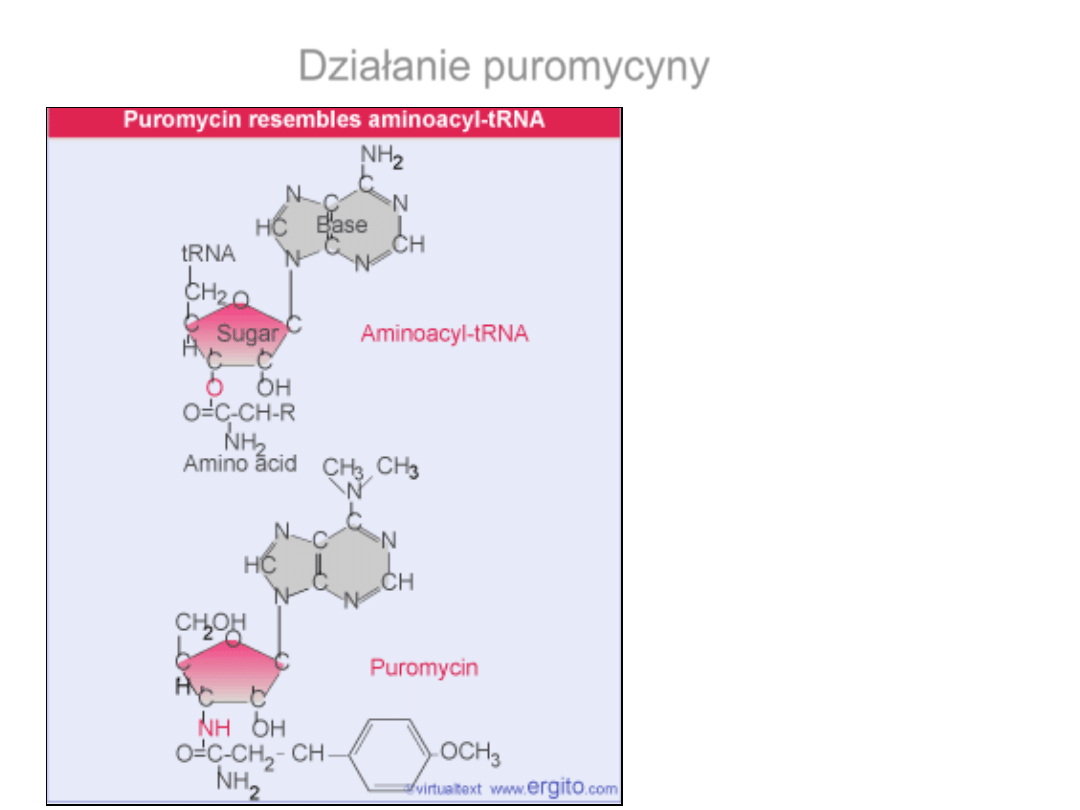
Puromycyna blokuje
syntezę białek. Naśladuje
ona tRNA i zostaje
wbudowana do rosnącego
łańcucha polipeptydowego.
Działanie puromycyny
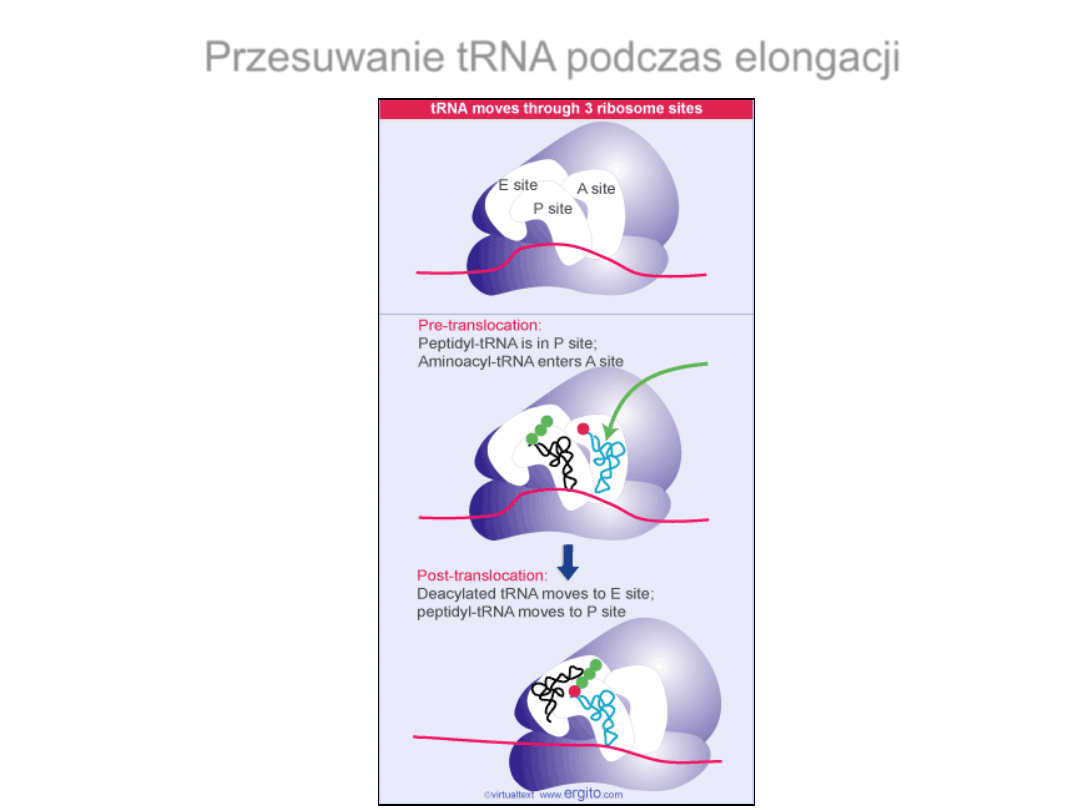
Przesuwanie tRNA podczas elongacji
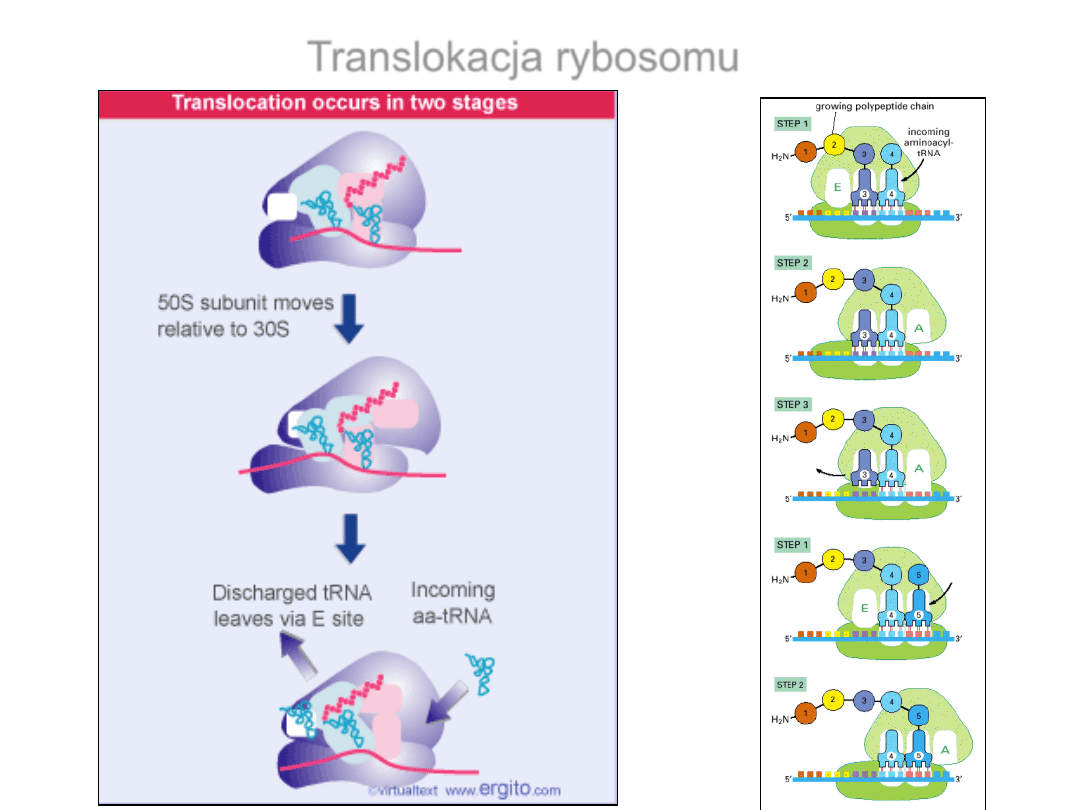
Translokacja rybosomu
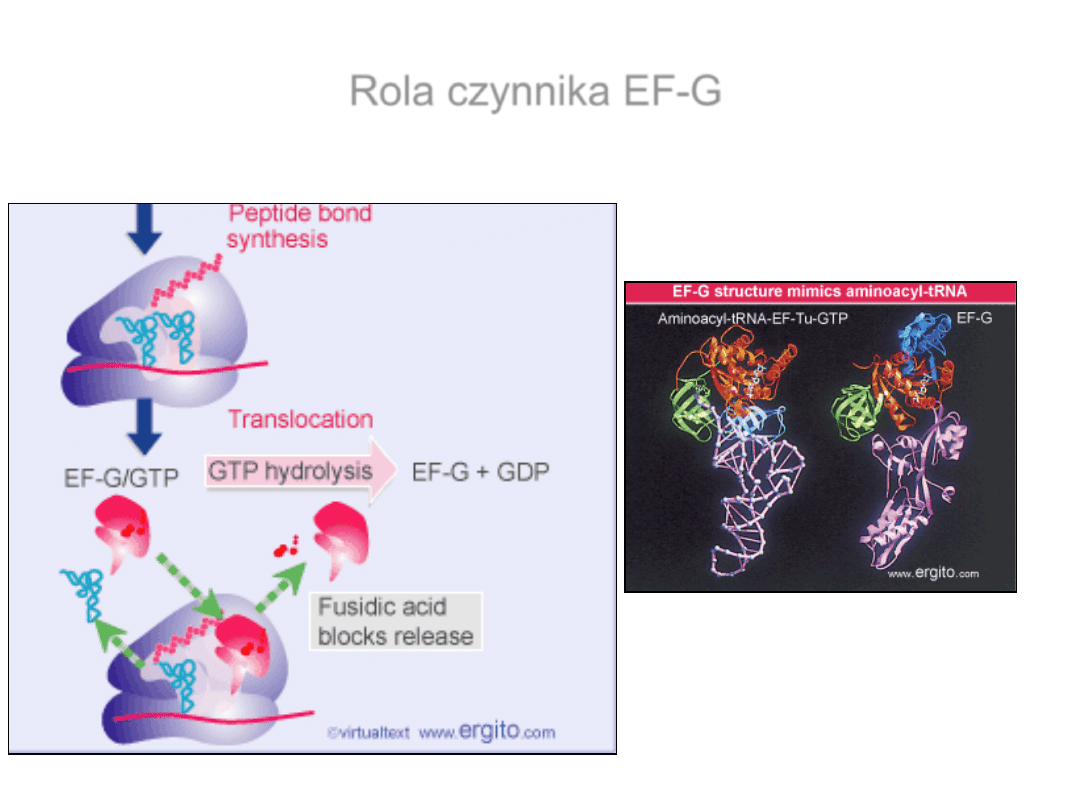
EF-G- czynnik potrzebny do
translokacji rybosomu.
Kwas fuzydowy – blokuje
uwolnienie EF-G
●GDP
eEF2 – eukariotyczny
odpowiednik EF-G
Rola czynnika EF-G
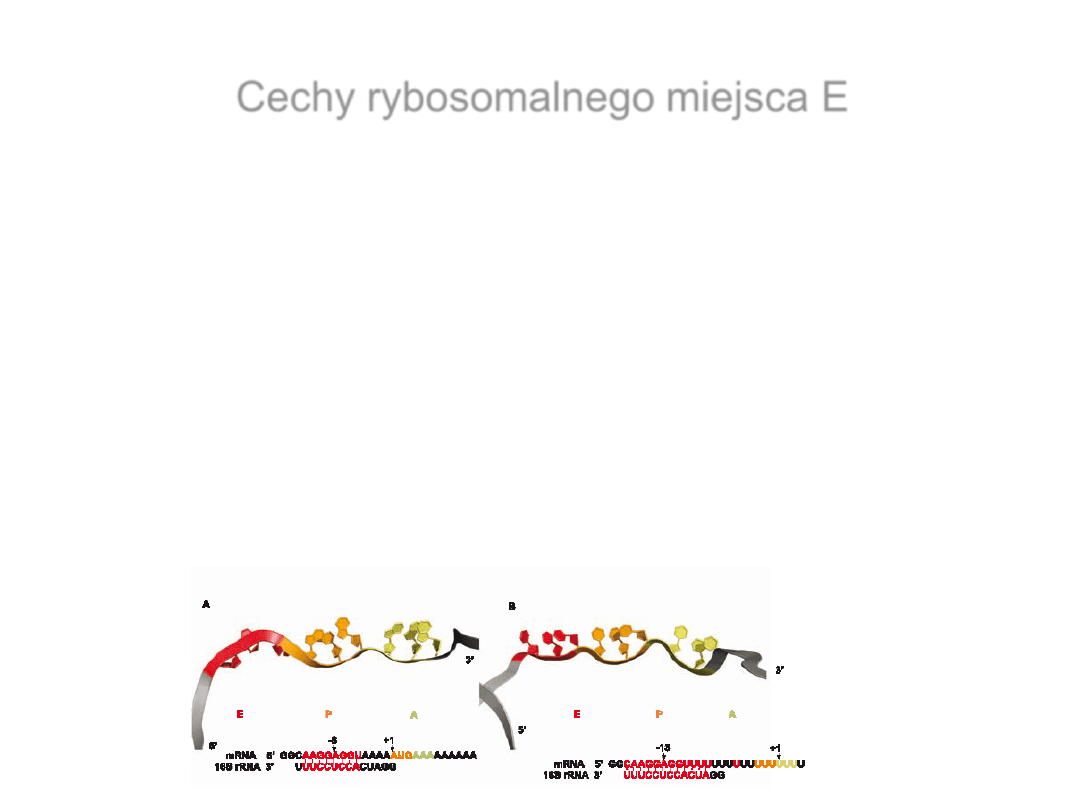
Cechy rybosomalnego miejsca E
•
Cząsteczka deacylowanego tRNA obecna w miejscu E
(E-tRNA
) jest z nim związana w sposób stabilny
•
E-tRNA
jest połączone z cząsteczką mRNA poprzez specyficzne
odziaływania kodon-antykodon
•
Istnieje obustronna zależność funkcjonalna między miejscami E i A
– allosteryczny model trójmiejscowy elongacji peptydu
Kompleks inicjatorowy
Po inicjacji translacji
Funkcjonowanie miejsca E
•
Powiązano energię aktywacji wiązania AA-tRNA do miejsca A z istnieniem
w rybosomie E-tRNA
•
Edeina, która blokuje miejsce E, powoduje
włączenie błędnego AA-tRNA w
miejsce A
•
Mutacje w
białkach S7 i S11, które osłabiają oddziaływania pętli
antykodonowej E-tRNA z rybosomem
powodują poważne błędy w selekcji
AA-tRNA do miejsca A
•
Osłabienie lub utrata oddziaływań kodon-antykodon w miejscu E prowadzi
do
przesunięcia ramki odczytu
•
Modyfikacja
końca CCA-3’ tRNA powoduje znaczne obniżenie
powinowactwa deacylowanej
cząsteczki tRNA do miejsca E

Istnieją dowody na to, że
E-
tRNA, powodując obniżenie
powinowactwa AA-tRNA do miejsca
A, zwiększa istotność odziaływań
kodon-antykodon
Document Outline
- Slide Number 1
- Slide Number 2
- Slide Number 3
- Slide Number 4
- Slide Number 5
- Slide Number 6
- Slide Number 7
- Slide Number 8
- Slide Number 9
- Slide Number 10
- Slide Number 11
- Cechy rybosomalnego miejsca E
- Funkcjonowanie miejsca E
- Istnieją dowody na to, że E-tRNA, powodując obniżenie powinowactwa AA-tRNA do miejsca A, zwiększa istotność odziaływań kodon-antykodon
Wyszukiwarka
Podobne podstrony:
2014 BPEG część 8 regulacja translacji
2014 BPEG część 5 inicjacja translacji
2014 BPEG część 7 terminacja translacji
2014 BPEG część 1 mRNA i tRNA
2014 BPEG część 9 fałdowanie i modyfikacje białek
2014 BPEG część 3 syntetazy
2014 BPEG część 2 kod genetyczny
2014 BPEG część 4 rybosomy
Mydło - część tłumaczenie translatorem, Ekologia, Dom bez chemii
BPEG część 10 systemy degradacji i autofagia
Inzynieria materialowa czesc obliczeniowa, Elektrotechnika AGH, Semestr III zimowy 2013-2014, Inżyni
Podstawy statystyki i ekonometrii 2014 część 1
Podstawy statystyki i ekonometrii 2014 część 2
Translacja elongacja 2
2014 06 test tech czesc II
egzamin zawodowy MGR 2014 czesc Nieznany
więcej podobnych podstron