
niemo˝liwe wnikni´cie do nich obcego py∏ku. Wynikajàca z takiej budowy samopyl-
noÊç u∏atwi∏a przeprowadzanie eksperymentów krzy˝ówkowych.
Mendel rozpoczà∏ doÊwiadczenia od wyhodowania roÊlin grochu genetycznie
czystych, tzw.
linii czystych, to znaczy jednolitych pod wzgl´dem danej cechy, np.
8
1.1.
P
rzedmiotem badaƒ genetycznych sà zarówno zjawiska dziedzicznoÊci, jak i zmien-
noÊci.
ISTOTA DZIEDZICZENIA.
ZASADY DZIEDZICZNOÂCI
Genetyka
to nauka
o przekazywaniu cech
potomstwu, zajmujà-
ca si´ badaniem praw
dziedzicznoÊci i zmien-
noÊci organizmów.
ZmiennoÊç – zró˝nico-
wanie cech w obr´bie
jednego gatunku; mo-
˝e byç uwarunkowana
czynnikami genetycz-
nymi i Êrodowiskowy-
mi. ZmiennoÊç przeja-
wia si´ ró˝nicami
w budowie, fizjologii
i zachowaniu poszcze-
gólnych osobników.
DEFINICJA
Fot. 1.
Grzegorz Mendel
(1822–1884)
DziedzicznoÊç polega na przekazywaniu w pro-
cesie rozmna˝ania cech z jednego pokolenia na
drugie, dzi´ki czemu pszenica rodzi pszenic´,
a cz∏owiek wydaje na Êwiat cz∏owieka. Jest uwa˝a-
na za jednà z podstawowych w∏aÊciwoÊci organi-
zmów ˝ywych, dzi´ki której poszczególne gatunki
zachowujà swoje istnienie i odr´bnoÊç. Jest to cecha
o charakterze zachowawczym. Natomiast
zmien-
noÊç obejmuje procesy prowadzàce do wytworze-
nia ró˝nic w wyposa˝eniu genetycznym mi´dzy for-
mami rodzicielskimi a potomstwem. Ta w∏aÊciwoÊç
organizmów jest w pewnym sensie przeciwieƒ-
stwem dziedzicznoÊci. Du˝e zró˝nicowanie gene-
tyczne ma charakter twórczy. Dzi´ki niemu roÊliny
i zwierz´ta podlegajà zmianom, ulegajà ewolucji.
Poczàtki nauki o dziedzicznoÊci wià˝à si´ z eks-
perymentami
Grzegorza Mendla (fot. 1.), zakonnika,
opata klasztoru Augustianów w Brnie na Morawach.
Dzi´ki nowatorskim pomys∏om, w∏aÊciwemu do-
borowi materia∏u badawczego i zdolnoÊciom Men-
dlowi uda∏o si´ przedstawiç pierwszy statystyczny
(iloÊciowy) model dziedziczenia cech. By∏o to tym
bardziej cenne, ˝e w czasie gdy opublikowa∏ swojà
najwa˝niejszà prac´, nic jeszcze nie wiedziano
o chromosomach i podzia∏ach komórkowych.
Grzegorz Mendel prowadzi∏ badania nad dzie-
dziczeniem cech grochu zwyczajnego. RoÊlina ta
okaza∏a si´ prawie idealna do prowadzonych do-
Êwiadczeƒ. Ma ona liczne odmiany o sta∏ych, wy-
raênie odró˝niajàcych si´ od siebie cechach. Ist-
niejà na przyk∏ad odmiany o kwiatach czerwonych
i inne o kwiatach bia∏ych, o nasionach ˝ó∏tych i na-
sionach o barwie zielonej. Sà te˝ odmiany o na-
sionach g∏adkich i inne – o nasionach pomarsz-
czonych. Groch wytwarza kwiaty obup∏ciowe (tzn.
majàce organy p∏ciowe m´skie – pr´ciki – i ˝eƒskie
– s∏upki). Pr´ciki i s∏upek sà szczelnie otoczone
przez p∏atki korony kwiatu, dzi´ki czemu jest
wy∏àcznie roÊliny o kwiatach czerwonych. Pos∏u-
giwanie si´ liniami czystymi w eksperymentach
krzy˝ówkowych daje wi´c pewnoÊç, co wnosi do
konkretnej krzy˝ówki ka˝dy z organizmów rodzi-
cielskich. Wielokrotne powtarzanie ka˝dego eks-
perymentu umo˝liwi∏o Mendlowi statystyczne
opracowanie wyników, które do dziÊ stanowi pod-
staw´ wszelkich eksperymentów krzy˝ówkowych.
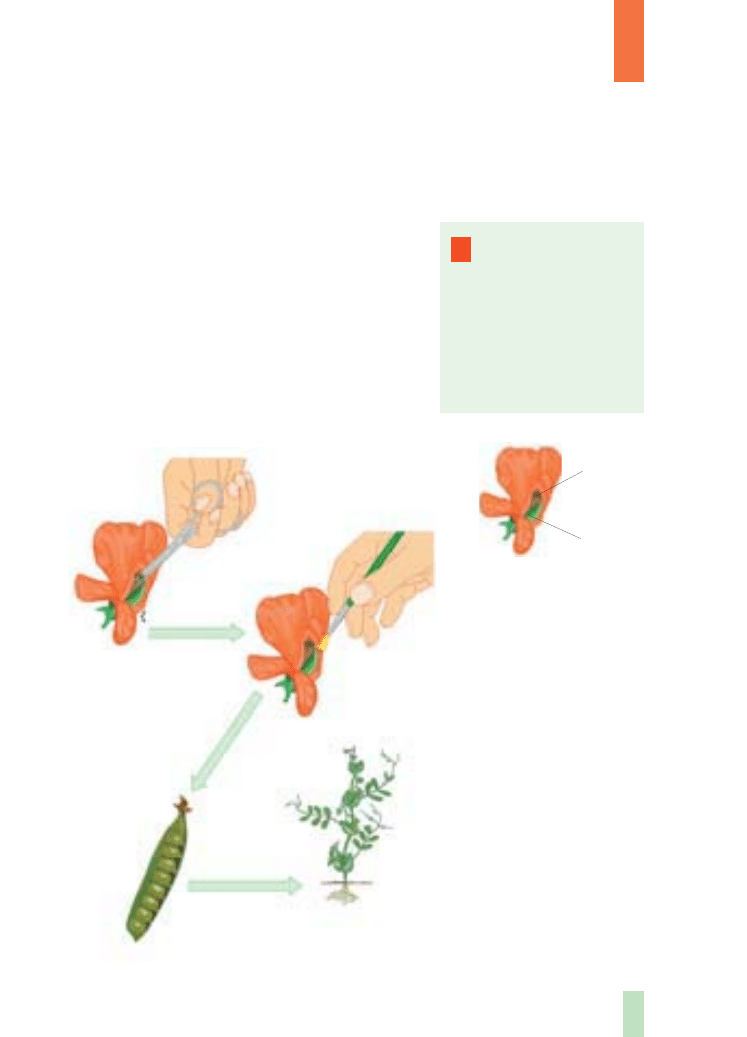
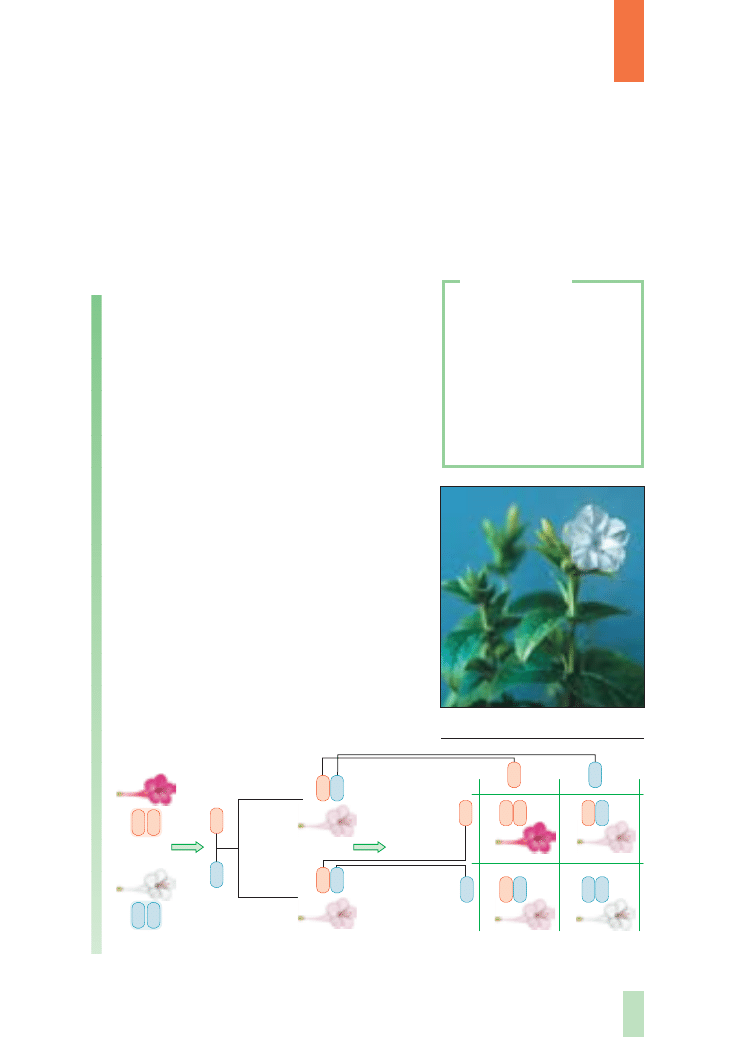
Praca badawcza Mendla polega∏a na sztucz-
nym krzy˝owaniu roÊlin (rys. 3.) ró˝niàcych si´
okreÊlonà cechà i analizowaniu wyst´powania tej
cechy u potomstwa.
9
A – U grochu ∏atwo mo˝na
przeprowadziç sztuczne
zapylenie. Usuwa si´ pr´ciki
wybranego kwiatu, aby
zapobiec samozapyleniu.
Na znami´ s∏upka takiego
kwiatu trzeba nanieÊç py∏ek
pochodzàcy z innego kwiatu
grochu. Potem nale˝y poczekaç
na pojawienie si´ stràków
z nasionami wewnàtrz.
Z takich nasion wyrosnà roÊliny
stanowiàce pierwsze pokolenie
potomne (F
1
). Natomiast
drugie pokolenie potomne (F
2
)
mo˝na uzyskaç, poddajàc
samozapyleniu roÊliny
pokolenia F
1
lub je krzy˝ujàc.
B – Budowa kwiatu grochu
– aby pokazaç pr´ciki i s∏upek,
odchylono p∏atki korony.
Rys. 3.
Schemat
eksperymentów krzy˝ówkowych
przeprowadzonych u grochu
zwyczajnego przez G. Mendla
usuni´cie
pr´cików
przeniesienie
py∏ku na
s∏upek roÊliny
przyjmujàcej
wytworzenie
stràków
z nasionami
wzrost
i rozwój
wysianie nasion
A
P
P
F
1
barwy nasion (zielone i ˝ó∏te), barwy kwiatów (bia∏e i czerwone) czy wysokoÊci p´-
dów (wysokie i niskie). Linie czyste charakteryzujà si´ tym, ˝e jeÊli skrzy˝uje si´ osob-
niki nale˝àce do jednej linii, to potomstwo powsta∏e w wyniku krzy˝owania powtó-
rzy cechy rodziców (b´dzie jednolite). Na przyk∏ad ze skrzy˝owania ze sobà roÊlin
grochu o kwiatach czerwonych, stanowiàcych lini´ czystà, w potomstwie uzyska si´
B
s∏upek
pr´ciki
Zastanów si´
i odpowiedz
na pytanie
Dlaczego podczas
sztucznego zapylenia
na kwiaty z naniesionym
py∏kiem dodatkowo
nak∏ada si´ papierowe
torebki?
?

Podda∏ on analizie 7 cech wyst´pujàcych u grochu, badajàc oddzielnie, jak dzie-
dziczy si´ ka˝da z nich (rys. 4). JeÊli w serii doÊwiadczeƒ wzià∏ pod uwag´ d∏ugoÊç
∏odyg, to nie analizowa∏ innych cech, którymi roÊliny rodzicielskie mog∏y si´ ró˝niç.
10
Cecha dominujàca
Cecha recesywna
wysokoÊç p´du
barwa kwiatów
barwa nasion
powierzchnia
nasion
barwa m∏odych
stràków (przed
dojrzeniem)
kszta∏t stràków
rozmieszczenie
kwiatów
na p´dzie
wzd∏u˝ ca∏ego p´du
(w kàtach liÊci)
na szczycie p´du
czerwona
bia∏a
˝ó∏ta
zielona
g∏adka
pomarszczona
zielona
˝ó∏ta
sp∏aszczone bez przew´˝eƒ
przew´˝enia mi´dzy nasionami
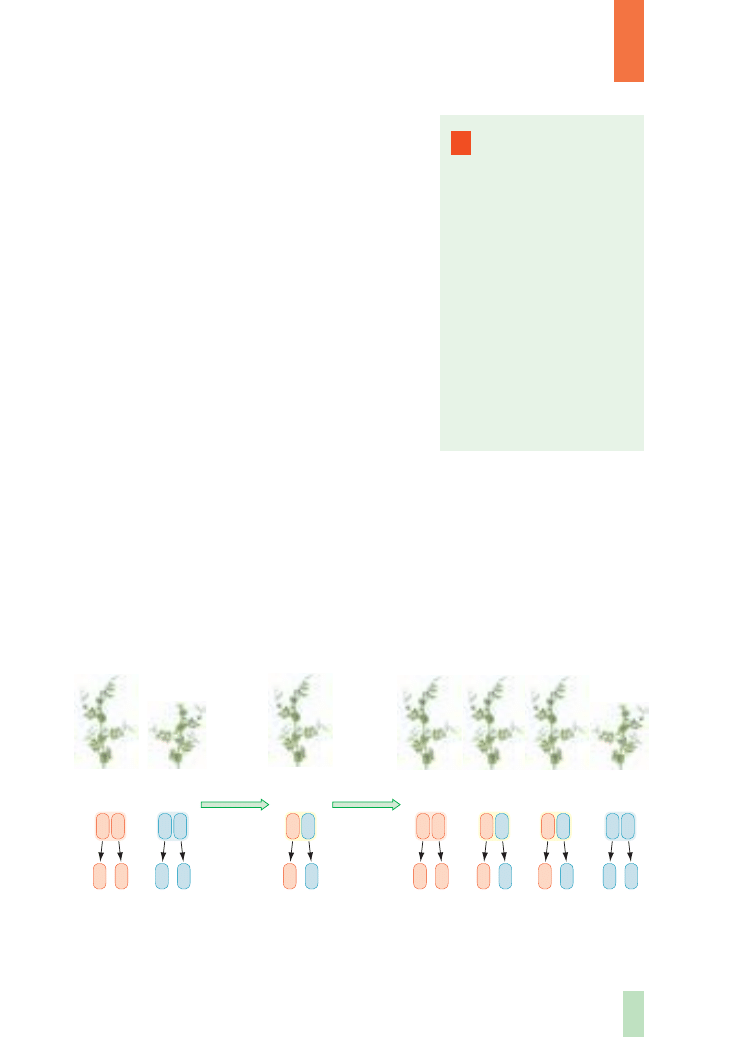
Badajàc na przyk∏ad dziedziczenie wysokoÊci p´du, w pokoleniu rodzicielskim
(P) skrzy˝owa∏ dwie linie czyste grochu, z których jedna charakteryzowa∏a si´ p´dami
wysokimi, a druga niskimi, i analizowa∏ pojawienie si´ tej cechy u potomstwa (rys. 5.).
W pierwszym pokoleniu potomnym (F
1
) otrzyma∏ wszystkie roÊliny o p´dach wysokich,
Rys. 4.
Cechy grochu zwyczajnego, których dziedziczenie bada∏ Mendel
roÊlina wysoka
roÊlina niska
(kar∏owata)
Zas∏ugà Mendla jest tak˝e wprowadzenie sposobu oznaczania alleli, stosowane-
go w genetyce do dziÊ. Du˝à literà (np. A) oznacza si´ allel dominujàcy, natomiast ma-
∏à literà (np. a) allel recesywny. Allele A oraz a sà allelami jednego genu. W przypad-
ku jednej cechy mogà wyst´powaç trzy kombinacje alleli: AA, aa i Aa. Identyczne
allele (AA lub aa) wyst´pujà u homozygot, natomiast ró˝ne allele (Aa) majà heterozy-
goty. Homozygoty mogà byç dwojakiego rodzaju: dominujàce (AA) lub recesywne
(aa). Mendel stwierdzi∏ równie˝, ˝e o przekazywaniu danego allelu decyduje przypa-
dek, to znaczy ˝e istnieje jednakowe prawdopodobieƒstwo przekazania potomkowi
ka˝dego z alleli danego genu.
czyli ta cecha by∏a
cechà dominujàcà. Cecha dru-
giego z rodziców nie ujawni∏a si´, by∏a to wi´c
ce-
cha recesywna. DoÊwiadczenia Mendla dotyczy∏y
tak˝e drugiego pokolenia potomnego (F
2
).
Drugie pokolenie potomne (F
2
) nie by∏o ju˝ jed-
nolite. Wykazywa∏o zró˝nicowanie pod wzgl´dem
ocenianej cechy. Powsta∏y roÊliny o p´dach wysokich
i niskich, przy czym wysokich by∏o oko∏o trzech razy
wi´cej ni˝ niskich. Analogiczne rezultaty Mendel
otrzyma∏ podczas krzy˝owania roÊlin charakteryzujà-
cych si´ pozosta∏ymi wybranymi przez niego cechami.
Za wystàpienie ka˝dej cechy sà odpowiedzialne
geny. Danà cech´ warunkujà zawsze dwa allele, przy
czym ka˝dy organizm rodzicielski przekazuje swemu
potomstwu tylko jeden z nich. Wszystkie organizmy
majà dwa allele danego genu (np. groch ma dwa alle-
le genu warunkujàcego wysokoÊç roÊlin), jeden odzie-
dziczony po jednym rodzicu, a drugi po drugim. Takie
za∏o˝enia przyjà∏ Mendel (stosujàc innà terminologi´)
i okaza∏y si´ one trafne.
11
Zastanów si´
i odpowiedz
na pytanie
åw. 1. Czy w prowadzo-
nych przez Mendla
krzy˝ówkach przeniesienie
py∏ku z roÊlin o p´dach
wysokich na s∏upki
roÊlin niskich da∏o taki
sam efekt jak przeniesienie
py∏ku z roÊlin o p´dach
niskich na s∏upki roÊlin
wysokich?
åw. 2. Czy stràki
powstajàce na roÊlinie
przyjmujàcej py∏ek
stanowià ju˝ pokolenie F
1
?
?
Rys. 5.
Dziedziczenie pojedynczej cechy (wysokoÊci ∏odyg) w pierwszym i drugim
pokoleniu potomnym u grochu
gamety
gamety
P (rodzice)
roÊlina
wysoka
T T
T
T
roÊlina
wysoka
T T
T
T
roÊlina
wysoka
T t
T
t
roÊlina
wysoka
T t
T
t
pokolenie F
1
gamety
roÊlina
wysoka
T t
T
t
roÊlina
niska
t
t
t
t
roÊlina
niska
t
t
t
t
pokolenie F
2

Wszystkie roÊliny nale˝àce do pokolenia F
1
majà taki sam zestaw alleli warunku-
jàcych danà cech´. Ka˝da z nich posiada allel dominujàcy (A) pochodzàcy od jedne-
go z rodziców i allel recesywny (a) pochodzàcy od drugiego z rodziców. Poniewa˝
przejawia si´ jedynie dzia∏anie allelu dominujàcego (dzia∏anie allelu recesywnego
jest t∏umione), to wszystkie roÊliny sà jednolite pod wzgl´dem analizowanej cechy
– w podanym przyk∏adzie wszystkie majà wysokie p´dy.
Natomiast w drugiej krzy˝ówce prowadzàcej do powstania pokolenia F
2
rozszcze-
pienie cech zosta∏o spowodowane przypadkowà segregacjà alleli. W jej efekcie powsta-
∏y osobniki o ró˝nych kombinacjach alleli warunkujàcych okreÊlonà cech´. Przy za∏o˝e-
niu, ˝e allele sà przekazywane potomstwu na drodze losowej, stosunek liczby mo˝liwych
kombinacji, w których wyst´puje co najmniej jeden allel dominujàcy (uk∏ad AA lub Aa,
powodujàcy ujawnienie si´ w fenotypie cechy dominujàcej), do liczby kombinacji za-
wierajàcej wy∏àcznie allele recesywne (uk∏ad aa, wywo∏ujàcy ujawnienie si´ cechy
recesywnej) wynosi 3 : 1. Ten teoretyczny stosunek fenotypowy nie oznacza jednak,
˝e w pokoleniu F
2
otrzymuje si´ trzy roÊliny wysokie i jednà niskà. Informuje jedynie, ˝e
prawdopodobieƒstwo pojawienia roÊlin o p´dach wysokich i niskich wynosi 3 : 1.
W tego typu eksperymentach bardzo rzadko teoretycznie obliczone prawdopo-
dobieƒstwo idealnie odpowiada rzeczywistym wynikom. Cz´sto wyst´pujà mniej
lub bardziej znaczne odchylenia od stosunku 3 : 1. Zale˝y to od korzystniejszych
lub mniej korzystnych warunków, w jakich znajdujà si´ poszczególne osobniki, lub
te˝ od wi´kszej lub mniejszej ˝ywotnoÊci danej klasy organizmów. OczywiÊcie im
wi´ksza próba b´dzie badana, tym wi´ksza b´dzie zgodnoÊç wyników z obliczo-
nymi. Na przyk∏ad Mendel, analizujàc wysokoÊç p´dów, otrzyma∏ w pokoleniu F
2
787 roÊlin o p´dach wysokich i 277 roÊlin o p´dach niskich, co odpowiada stosun-
kowi liczbowemu 2,84 : 1. W innym eksperymencie analizowa∏ barw´ nasion
i otrzyma∏ w pokoleniu F
2
6022 nasiona o barwie ˝ó∏tej i 2001 nasion zielonych,
co w tym przypadku odpowiada stosunkowi liczbowemu 3,01 : 1.
12
INFORMACJE DODATKOWE
Zastanów si´
i odpowiedz
na pytanie
Jakà barw´ mogà mieç
nasiona w jednym stràku
roÊlin pokolenia F
2
u grochu zwyczajnego?
?
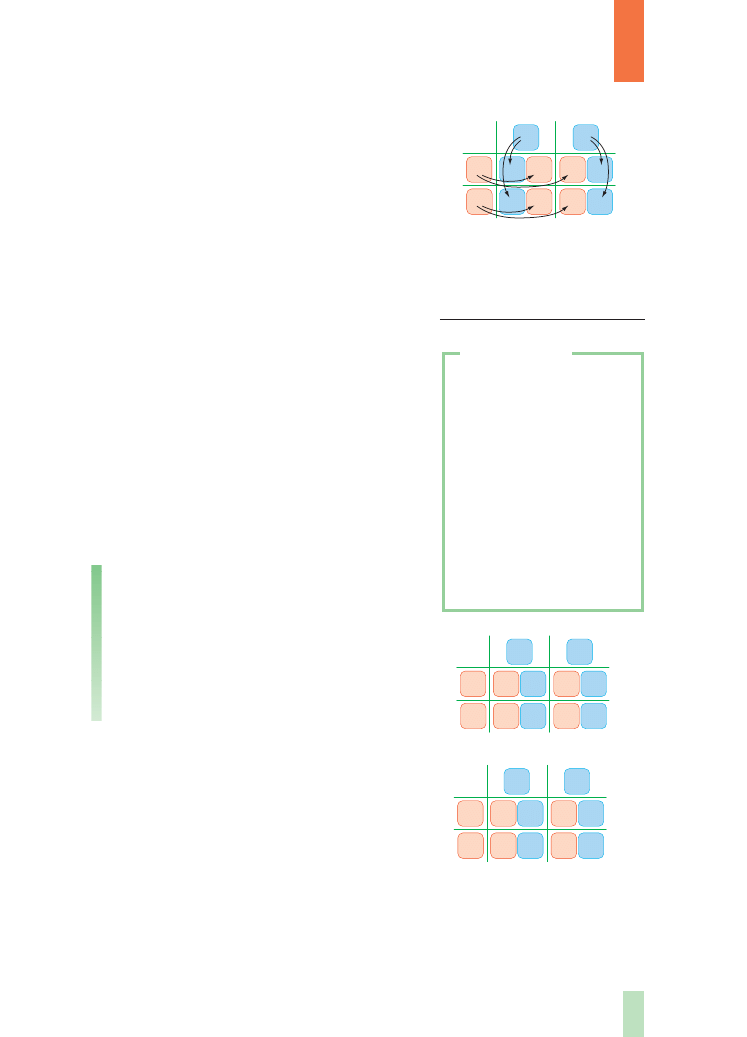
Wyniki ka˝dej krzy˝ówki mo˝na przewidzieç,
konstruujàc odpowiednià tabel´, tzw.
szachownic´
genetycznà (rys. 6.). Jest to prosty sposób otrzyma-
nia wszystkich teoretycznych kombinacji alleli. Na-
tomiast przez interpretacj´ ka˝dego z genotypów
(tworzonych przez allele genów) mo˝na okreÊliç
równie˝ fenotypy. Zaletà szachownicy genetycznej
jest przejrzystoÊç wyników i ∏atwoÊç jej konstru-
owania. Warunkiem w∏aÊciwego jej przygotowa-
nia jest poprawne wypisanie wszystkich mo˝liwych
kombinacji alleli w gametach. Zadanie to nie jest
trudne, wymaga jednak nieco wprawy. Szachownic´ genetycznà uzyskuje si´, umiesz-
czajàc z lewej strony w pierwszej kolumnie wszystkie typy gamet ˝eƒskich, nato-
miast na górze (górny rzàd) wszystkie typy gamet m´skich (lub odwrotnie). Wystarczy
ju˝ tylko wype∏niç kolejne pola szachownicy, wpisujàc symbole alleli wnoszonych
przez ∏àczàce si´ ze sobà gamety. W ten sposób otrzymuje si´ wszystkie mo˝liwe
kombinacje alleli u potomstwa. W∏aÊciwy zapis powinien zawieraç na pierwszym
miejscu allel dominujàcy, oczywiÊcie w przypadku gdy jest on reprezentowany w da-
nym genotypie, czyli Aa, a nie aA. Z szachownicy wynika, ˝e rozszczepienie fenoty-
pów w pokoleniu F
2
krzy˝ówki jednogenowej (dotyczàcej dziedziczenia pojedynczej
13
I prawo Mendla:
Organizmy rozmna˝a-
jàce si´ p∏ciowo po-
siadajà dwa allele
ka˝dego genu, które
ulegajà rozdzieleniu
(segregacji) podczas
formowania si´ ga-
met. W ten sposób
ka˝da z komórek roz-
rodczych zawiera tyl-
ko jeden allel danego
genu.
DEFINICJA
A
a
A
a
a
A
a
a
A
A
A
a
a
a
A
a
a
A
a
a
A
a
a
a
a
a
A
A
a
A
a
A
A
a
A
a
Rys. 7.
Przyk∏ad krzy˝ówki
testowej, umo˝liwiajàcej
okreÊlenie genotypu osobnika
A – homozygoty
B – heterozygoty
cechy) nast´puje w stosunku 3: 1, natomiast rozk∏ad
genotypów mo˝na wyraziç stosunkiem liczbowym
1AA : 2Aa : 1aa.
Analiza wyników badaƒ doprowadzi∏a Mendla
do sformu∏owania ogólnego prawa genetycznego.
Jest ono znane pod nazwà
I prawa Mendla (okre-
Êlanego równie˝ jako prawo segregacji lub prawo
czystoÊci gamet).
Wiedzàc, ˝e homozygoty dominujàce majà ten
sam fenotyp co heterozygoty, zasadne staje si´ pyta-
nie o sposób odró˝niania tych organizmów. W tym
celu stosuje si´ tzw.
krzy˝ówk´ testowà (rys. 7.),
w której krzy˝uje si´ testowanego osobnika (tego,
którego genotyp nie jest znany) zawsze z homozygo-
tà recesywnà. Na podstawie wyników takiej krzy-
˝ówki mo˝na okreÊliç genotyp testowanego osobni-
ka. Interpretacja wyników tej krzy˝ówki przypomina
rozwiàzywanie równania z jednà niewiadomà.
W wypadku jednej cechy istniejà dwie mo˝liwoÊci:
albo potomstwo b´dzie jednolite fenotypowo (jedna
klasa osobników) (rys. 7A.), albo powstanà organi-
zmy o dwu ró˝nych fenotypach w stosunku liczbo-
wym 1: 1 (dwie klasy osobników) (rys. 7B.). W pierw-
szym przypadku testowany osobnik by∏ homozygotà
dominujàcà, natomiast w drugim – heterozygotà.
Mendel wykona∏ równie˝ krzy˝owanie te-
stowe. Krzy˝ówka testowa daje dodatkowo mo˝-
liwoÊç bezpoÊredniego sprawdzenia za∏o˝enia,
˝e segregacja alleli u heterozygotycznych rodzi-
ców nast´puje w stosunku 1: 1. Takie krzy˝ów-
ki sà wykorzystywane do celów u˝ytkowych, na
przyk∏ad gdy hodowcy starajà si´ uzyskaç od-
miany genetycznie czyste (homozygotyczne)
pod wzgl´dem okreÊlonych cech.
Przedstawione przyk∏ady mia∏y na celu analiz´
dziedziczenia jednej wybranej cechy. Takie krzy-
˝ówki noszà nazw´
krzy˝ówek jednogenowych.
Grzegorz Mendel bada∏ tak˝e jednoczesne dziedzi-
czenie dwóch ró˝nych cech przez
krzy˝ówki dwu-
genowe. Eksperymenty te podjà∏ w celu wyjaÊnie-
nia, czy ró˝ne cechy dziedziczà si´ niezale˝nie od
siebie, czy te˝ istnieje mi´dzy nimi jakieÊ sprz´˝e-
nie, powodujàce ∏àczne (zale˝ne) przekazywanie
danych cech. Przyk∏adem takiego doÊwiadczenia
jest analiza dziedziczenia barwy nasion grochu
oraz rodzaju ich powierzchni. Jedna para alleli wy-
znacza barw´ nasion grochu (A – ˝ó∏tà, a – zielonà),
Rys. 6.
Szachownica
genetyczna przedstawiajàca
kombinacj´ alleli w pokoleniu F
2
po skrzy˝owaniu osobników
heterozygotycznych
A
B
INFORMACJE DODATKOWE
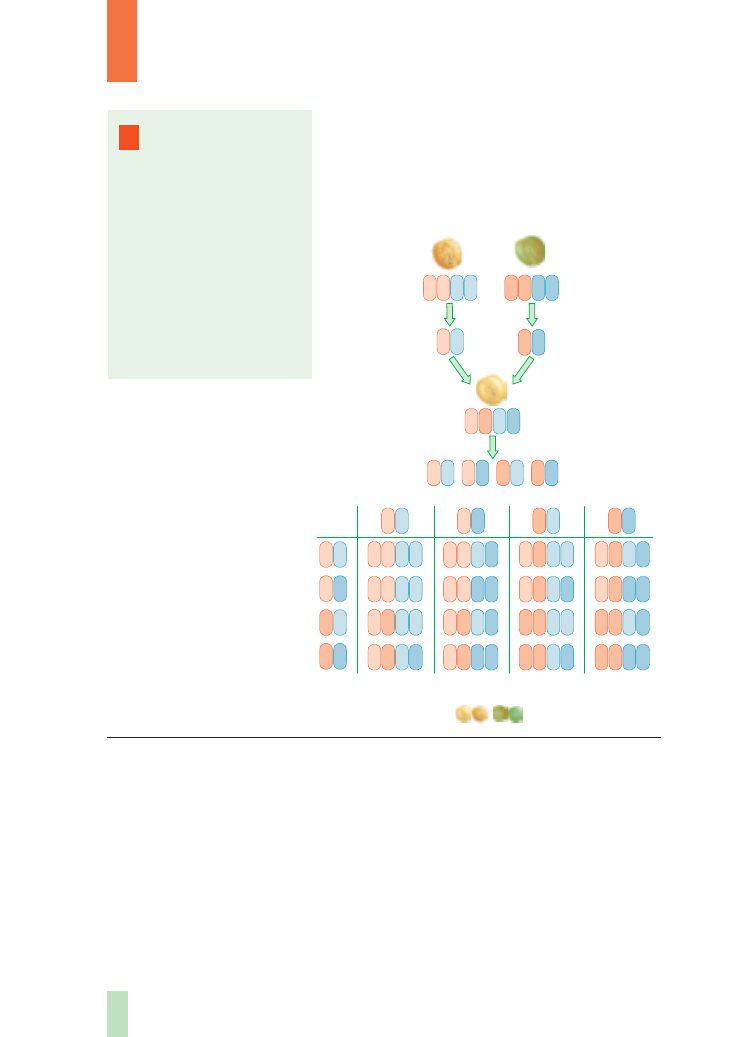
14
natomiast drugie wyros∏y z nasion o zielonej barwie i pomarszczonej powierzchni (ge-
notyp aabb – podwójna homozygota recesywna). Po skrzy˝owaniu takich roÊlin
pierwsze pokolenie potomne (F
1
) by∏o jednolite – wszystkie nasiona mia∏y ˝ó∏tà bar-
w´ i g∏adkà powierzchni´ (genotyp AaBb – podwójna heterozygota). U wszystkich
osobników w tym pokoleniu przejawia si´ dzia∏anie alleli dominujàcych obu genów
(A i B). Po samozapyleniu roÊlin pokolenia F
1
Mendel stwierdzi∏ wyst´powanie w po-
koleniu F
2
czterech fenotypowo ró˝nych klas nasion w nast´pujàcych stosunkach
liczbowych: 9/16 nasion ˝ó∏tych i g∏adkich, 3/16 nasion ˝ó∏tych i pomarszczonych,
3/16 nasion zielonych i g∏adkich oraz 1/16 nasion zielonych i pomarszczonych.
Wynik tego eksperymentu mo˝na wyjaÊniç za pomocà zasady czystoÊci gamet
i niezale˝nej segregacji. RoÊlina podwójnie heterozygotyczna (AaBb) b´dzie wytwarzaç
Rys. 8.
Dziedziczenie
dwóch cech
(barwy i rodzaju
powierzchni nasion)
w pierwszym
i drugim pokoleniu
potomnym u grochu
gamety
gamety
P (rodzice)
A
A
A
A
pokolenie F
1
pokolenie F
2
fenotypy
B
B
B
B
a
a
a
a b
b
A B
B
a
a b
A b
A B
B
a
a b
A b
b
b
B
a
A
B B
a
b
A
B
a
B B
a
a
b
B
a
a
b
A A B
A b
b
b
A A
b
A
B
a
b
b
A a
A B
A A B B
A A B b
A
B B
a
b
A
B
a
a b
b
A
B
a
b
b
A a
B b
a
a
b b
a
a
9 : 3 : 3 : 1
a druga para alleli wyznacza rodzaj powierzchni
nasion (B – g∏adkà, b – pomarszczonà).
Do krzy˝owania Mendel wybra∏ roÊliny pocho-
dzàce z linii czystych. Pierwsze z nich wyros∏y
z nasion o ˝ó∏tej barwie i g∏adkiej powierzchni (ge-
notyp AABB – podwójna homozygota dominujàca),
Zastanów si´ i podaj
odpowiedê
Gen determinujàcy barw´
˝ó∏tà (A) i gen warunkujàcy
g∏adkoÊç nasion (B) sà
dominujàce w stosunku
do recesywnych genów
warunkujàcych: barw´ zie-
lonà (a) i pomarszczenie
nasion (b). Na podstawie
genotypu okreÊl fenotyp
nast´pujàcych osobników:
AaBB, AaBb, aaBb, aabb,
Aabb, aaBB, AABB.
?
to w tzw.
II prawo Mendla (prawo niezale˝nej se-
gregacji cech, prawo niezale˝nego dziedziczenia).
Wzajemne relacje pomi´dzy allelami mogà
byç znacznie bardziej z∏o˝one, ni˝ wynika∏oby
to z doÊwiadczeƒ Mendla. Analiza dziedziczenia
cech u ró˝nych organizmów wykaza∏a, ˝e nie za-
wsze w stanie heterozygotycznym jeden z tworzà-
cych par´ alleli jest w pe∏ni dominujàcy w stosunku
cztery rodzaje gamet (zarówno ˝eƒskich, jak i m´skich) o ró˝nych kombinacjach al-
leli – AB, Ab, aB, ab – w stosunku 1: 1: 1: 1, a wi´c w równych iloÊciach. Zap∏odnie-
nie 4 ró˝nych rodzajów gamet ˝eƒskich przez 4 typy gamet m´skich daje 16 kombi-
nacji po∏àczeƒ, tworzàcych w drugim pokoleniu potomnym (F
2
) 9 ró˝nych genotypów.
Wytworzà one nasiona o 4 ró˝nych fenotypach (4 klasy) w stosunku 9 : 3 : 3 : 1, co
jest zgodne z wynikami eksperymentów otrzymanymi przez Mendla.
DoÊwiadczenia Mendla pokaza∏y wi´c, ˝e allele ró˝nych genów nie sà po∏àczone
w zespo∏y, lecz dziedziczà si´ niezale˝nie od siebie. Wyniki tych eksperymentów uj´-
15
II prawo Mendla:
Allele dwóch ró˝nych
genów dziedziczà si´
niezale˝nie. Sà one
przekazywane do ga-
met, tworzàc wszyst-
kie mo˝liwe kombina-
cje z jednakowà cz´-
stoÊcià.
DEFINICJA
do drugiego. Czasami obserwuje si´, ˝e heterozy-
gota ma fenotyp poÊredni pomi´dzy fenotypami
rodziców. W takim przypadku ˝aden z alleli nie
dominuje nad drugim, a ten rodzaj zale˝noÊci
okreÊla si´ jako
dominowanie niepe∏ne. Jako przy-
k∏ad mo˝e pos∏u˝yç dziedziczenie barwy kwia-
tów u dziwaczka (fot. 2.). U tej roÊliny kwiaty sà
najcz´Êciej czerwone lub bia∏e. Po skrzy˝owaniu
czystej linii tej roÊliny o kwiatach czerwonych
z czystà linià wytwarzajàcà kwiaty bia∏e otrzyma-
no w pokoleniu F
1
wynik zaskakujàcy. Wszystkie
roÊliny mia∏y kwiaty o barwie poÊredniej mi´dzy
fenotypami rodziców (ró˝owej). RoÊliny dziwacz-
ka kwitnàce na ró˝owo to heterozygoty, u któ-
rych ˝aden z alleli nie dominuje w pe∏ni. W po-
koleniu F
2
otrzymano natomiast rozszczepienie
cech kwiatów (czerwone : ró˝owe : bia∏e) w sto-
sunku 1 : 2 : 1. W tym przypadku rozk∏ad feno-
typów odpowiada rozk∏adowi genotypów.
Fot. 2.
Kwitnàcy dziwaczek
P (rodzice)
A A
pokolenie F
1
a a
A
a
A
a
pokolenie F
2
A a
A a
A A
a
a
a
A
A a
A a
INFORMACJE DODATKOWE
Rys. 9.
Dominowanie niepe∏ne u dziwaczka

rzàce takà par´ to tzw.
chromosomy homologiczne.
Na przyk∏ad komórki budujàce cia∏o cz∏owieka
(tzw. komórki somatyczne) majà po 46 chromoso-
mów stanowiàcych 23 pary, natomiast gamety za-
wierajà po jednym chromosomie z ka˝dej pary,
czyli majà 23 chromosomy. Zespó∏ chromosomów
(komplet chromosomów, oznaczany jako n) tworzà
chromosomy zawarte w plemniku bàdê komórce ja-
jowej. Geny zlokalizowane w jednym chromosomie
16
Wykonaj zadanie
i podaj odpowiedê
Jakie b´dzie pokolenie F
2
w przypadku przekazy-
wania potomstwu dwóch
niezale˝nie dziedziczàcych
si´ cech wykazujàcych
niepe∏nà dominacj´?
?
Wspó∏zale˝noÊç pomi´dzy genem a cechà mo-
˝e byç jeszcze bardziej z∏o˝ona, ni˝ wynika∏oby
z przedstawionych przyk∏adów.
Chromosomowa teoria
dziedzicznoÊci
Badania z prze∏omu XIX i XX wieku doprowadzi∏y
do poznania procesów mitozy i mejozy, dajàc pod-
stawy do w∏aÊciwego rozumienia i interpretowania
praw przekazywania genów ustalonych przez
Grzegorza Mendla. Uderzajàce podobieƒstwo mi´-
dzy zachowaniem si´ chromosomów podczas me-
jozy a opisanym przez Mendla zachowaniem si´
genów w czasie przekazywania ich z pokolenia na
pokolenie doprowadzi∏o do sformu∏owania teorii
lokalizacji genów w chromosomach. Teoria ta zna-
laz∏a doÊwiadczalne potwierdzenie w serii prac
przeprowadzonych w latach 1910–1914 przez
Thomasa Morgana i jego wspó∏pracowników. Da-
∏y one poczàtek wspó∏czesnej genetyce i przynios∏y
Morganowi Nagrod´ Nobla.
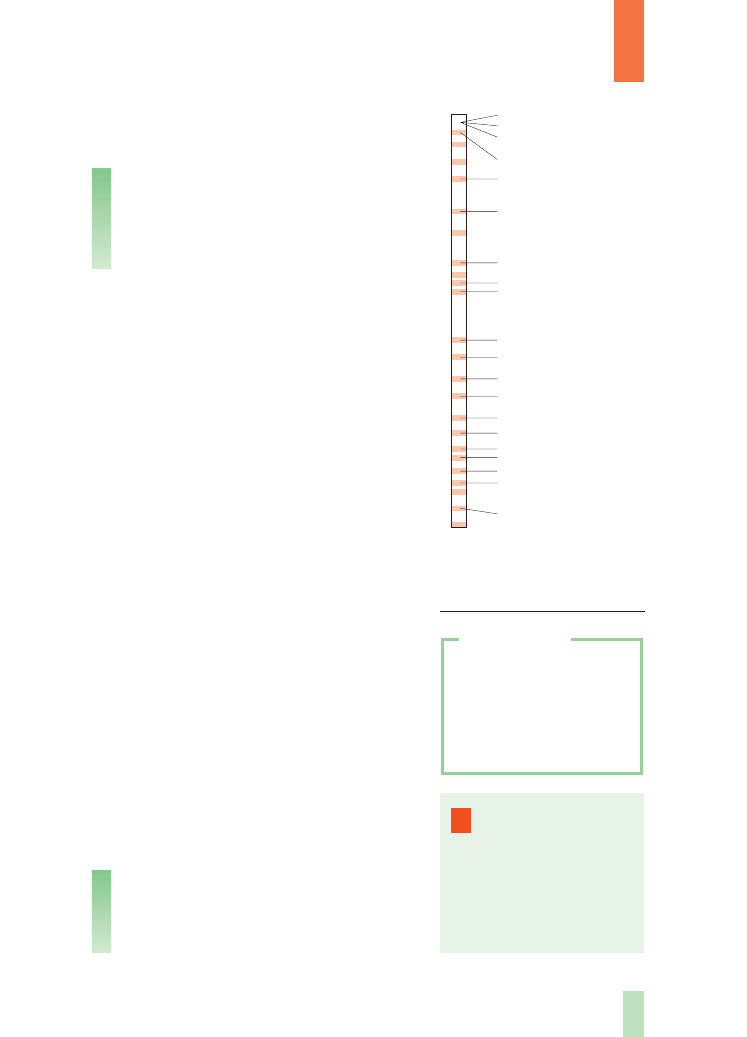
Podstawowym obiektem badaƒ Morgana i jego
nast´pców by∏a (i jest) muszka owocowa (Droso-
phila melanogaster, czytaj: drozofila melanogaster).
Liczba chromosomów jest charakterystyczna
i sta∏a dla ka˝dego gatunku. Zwykle nie przekra-
cza kilkudziesi´ciu, natomiast liczba cech charak-
terystycznych dla danego organizmu jest o wiele
wi´ksza. ¸atwo stàd wywnioskowaç, ˝e ka˝dy
z chromosomów musi zawieraç wiele genów. Na
przyk∏ad liczb´ genów u muszki owocowej szacu-
je si´ na kilkadziesiàt tysi´cy. Je˝eli weêmie si´ pod
uwag´, ˝e podstawowymi strukturami rozdzielany-
mi do komórek podczas podzia∏ów sà chromoso-
my, to wszystkie geny zlokalizowane w tym samym
chromosomie sà przekazywane ∏àcznie.
U wi´kszoÊci rozmna˝ajàcych si´ p∏ciowo organi-
zmów chromosomy tworzà pary. Chromosomy two-
Fot. 3.
Muszka owocowa
(obraz powi´kszony)
Chromosomy homo-
logiczne majà t´ samà
wielkoÊç, kszta∏t i ge-
ny okreÊlonych cech,
ró˝nià si´ zaÊ pocho-
dzeniem: jeden jest
dziedziczony po ojcu,
a drugi – po matce.
DEFINICJA
Fot. 4.
Komplet chromosomów
cz∏owieka (mikrofotografia
skaningowa)
17
stanowià tzw.
grup´ genów sprz´˝onych. Ca∏kowi-
cie niezale˝nie od siebie dziedziczà si´ jedynie te
geny, które le˝à w ró˝nych chromosomach.
Ka˝dy gen zajmuje w chromosomie okre-
Êlone miejsce (czyli locus, czytaj: lokus). Chro-
mosomy homologiczne zawierajà te same loci
(czytaj: loci) genowe u∏o˝one w identycznej ko-
lejnoÊci. Obydwa allele danego genu zajmujà
to samo miejsce w tych chromosomach.
W obr´bie ka˝dego chromosomu geny sà u∏o-
˝one liniowo. W zwiàzku z tym mo˝na wyznaczyç
pozycj´ ka˝dego genu w chromosomie, tworzàc
tzw.
mapy chromosomowe. Mapa chromosomu sta-
nowi ilustracj´ liniowego uk∏adu genów nale˝àcych
do danej grupy genów sprz´˝onych, ∏àcznie z poda-
niem kolejnoÊci i odleg∏oÊci mi´dzy nimi. Mapowa-
nie chromosomów nie jest sprawà prostà, a do usta-
lania po∏o˝enia genów stosuje si´ ró˝ne metody.
Wed∏ug II prawa Mendla allele dwóch ró˝nych
genów dziedziczà si´ niezale˝nie. Morgan udowod-
ni∏, ˝e tylko niektóre geny mogà dziedziczyç si´ nie-
zale˝nie od siebie, inne natomiast, stanowiàc grup´
genów sprz´˝onych, sà przekazywane ∏àcznie. W ja-
ki sposób zatem mogà byç dziedziczone dwa geny?
JeÊli geny le˝à na ró˝nych chromosomach (nie
sà sprz´˝one), to dziedziczà si´ niezale˝nie od sie-
bie, zgodnie z II prawem Mendla. Je˝eli natomiast
geny sà sprz´˝one (le˝à na jednym chromosomie),
to dziedziczà si´ ∏àcznie.
Dosyç cz´sto zdarza si´, ˝e w potomstwie
oprócz przewa˝ajàcej liczby obu typów rodziciel-
skich pojawiajà si´, choç rzadziej, tak˝e nowe
uk∏ady alleli, niewyst´pujàce w pokoleniu rodzi-
cielskim. Noszà one nazw´ zrekombinowanych.
Natomiast proces, w którego wyniku powstajà takie
nowe kombinacje alleli, nosi nazw´
rekombinacji.
Wyst´powanie wÊród genów sprz´˝onych pew-
nej liczby zrekombinowanych uk∏adów alleli Mor-
gan zinterpretowa∏ jako efekt wzajemnej wymiany
odcinków mi´dzy chromosomami homologiczny-
mi. Zjawisko to nazwa∏
crossing-over (czytaj:
krosing o∏wer).
Zrozumienie istoty procesu
crossing-over
wymaga znajomoÊci przebiegu mejozy. Kluczo-
wym momentem jest ten etap mejozy, kiedy
chromosomy homologiczne uk∏adajà si´ obok
siebie w pary. Wtedy sà one ju˝ podwojone,
cia∏o ˝ó∏te
skrzyd∏a ow∏osione
skrzyd∏a bez
poprzecznych ˝y∏ek
oczy karminowe
oczy romboidalne
oczy cynobrowe
oczy wst´gowate
oczy goêdzikowe
skrzyd∏a miniaturowe
skrzyd∏a faliste
skrzyd∏a pofa∏dowane
skrzyd∏a muszelkowate
mikroszczecinki
skrzyd∏a szczàtkowe
skrzyd∏a rozwidlone
cia∏o ciemne
w∏oski falisto skr´cone
bez szczecinek
oczy bia∏e
oczy rubinowe
Rys. 10.
Lokalizacja wybranych
genów na mapie chromosomu X
muszki owocowej
èród∏o: B. Rodkiewicz, G. Kerszman,
Zarys genetyki, Warszawa 1987
INFORMACJE
DODATKOWE
INFORMACJE
DODATKOWE
Zastanów si´
i odpowiedz
na pytanie
Czy niezale˝ne dziedzicze-
nie mo˝e byç równie˝ êró-
d∏em powstawania nowych
kombinacji genów?
?
Rekombinacja to wy-
st´powanie u potom-
stwa innych ni˝ u rodzi-
ców kombinacji genów,
a w efekcie tak˝e in-
nych zespo∏ów cech.
DEFINICJA
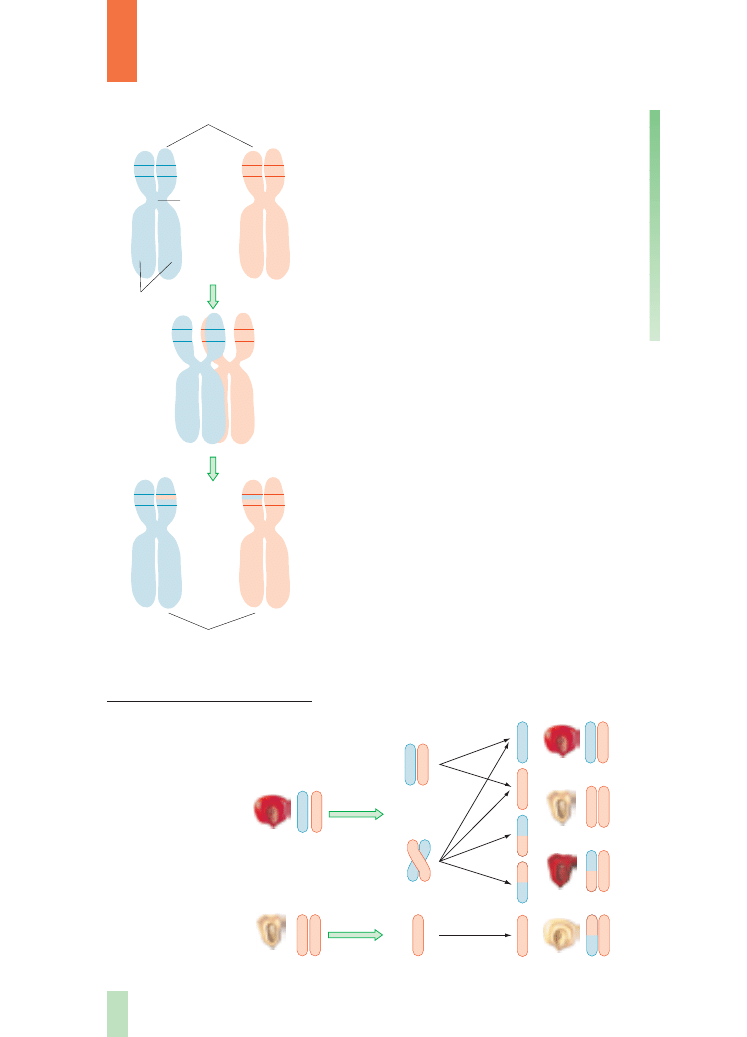
a wi´c ka˝dy chromosom jest zbudowany
z dwóch identycznych chromatyd (tzw. siostrza-
nych), po∏àczonych w rejonie zwanym centrome-
rem. Takà par´ chromosomów homologicznych
stanowià 4 chromatydy. Mo˝e wówczas docho-
dziç do p´kni´ç ramion chromosomów i wymiany
odcinków pomi´dzy chromosomami. Ten proces
wymiany odcinków, czyli crossing-over, zachodzi
mi´dzy dwiema chromatydami niesiostrzanymi
chromosomów homologicznych. W ten sposób
dochodzi do rozerwania istniejàcych sprz´˝eƒ
i powstania nowych kombinacji genów, wczeÊ-
niej nieistniejàcych w badanym materiale.
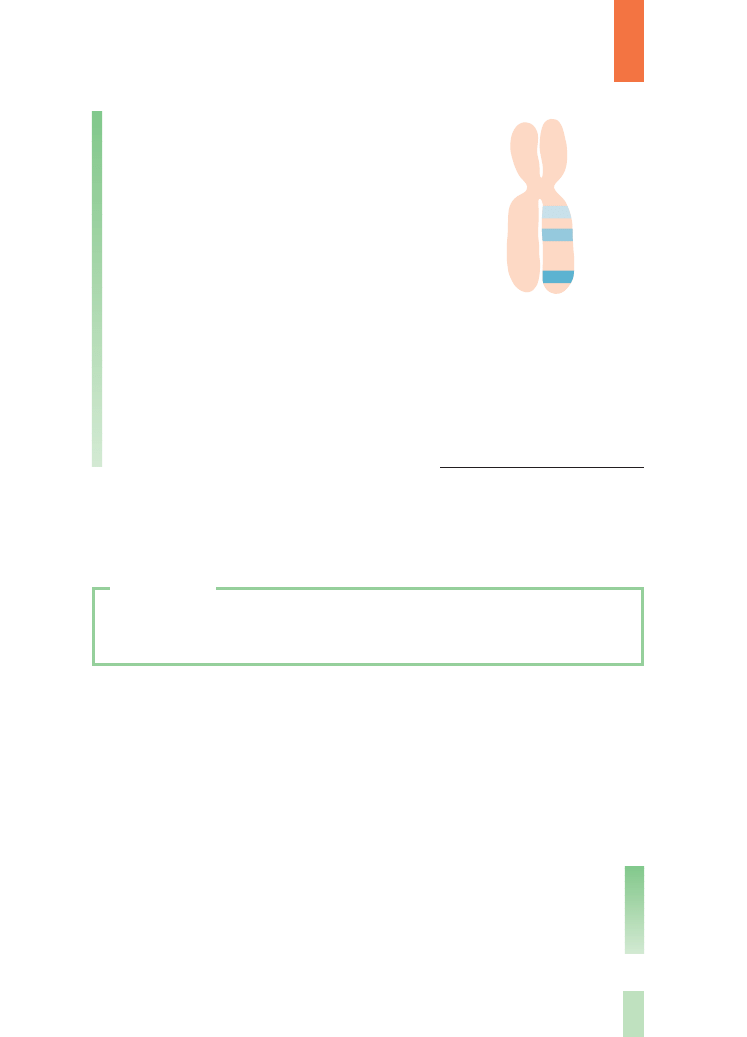
Przyk∏adem ilustrujàcym to zjawisko mogà byç
doÊwiadczenia nad przekazywaniem potomstwu
dwóch cech dziedziczàcych si´ zale˝nie (barwy
i kszta∏tu ziaren) u kukurydzy. Po skrzy˝owaniu po-
dwójnej heterozygoty o cechach dominujàcych AaBb
(ziarna czerwone, pe∏ne) z podwójnà homozygotà re-
cesywnà aabb (ziarna bezbarwne, sp∏aszczone) otrzy-
mano oko∏o 97% typów rodzicielskich. Oko∏o 3%
da∏o nowe uk∏ady alleli genów (ziarna czerwone
sp∏aszczone – Aabb i bezbarwne pe∏ne – aaBb). Gdy-
by zatem sprz´˝enie by∏o ca∏kowite, to w wyniku
krzy˝ówki testowej powinny powstaç 2 typy gamet
(AB i ab), a w konsekwencji ich zap∏odnienia przez
gamety wytwarzane przez podwójnà homozygot´
(ab) – tylko czerwone pe∏ne ziarna (Aabb) i bezbarw-
ne sp∏aszczone (aabb) w stosunku 1: 1. Pojawienie si´
nowych uk∏adów alleli w gametach (Ab i aB), czego
skutkiem jest powstanie nowych fenotypów, Êwiad-
czy o tym, ˝e sprz´˝enie analizowanych alleli w ob-
r´bie jednego chromosomu nie jest ca∏kowite.
18
para chromosomów homologicznych
centromer
wymiana
odcinków
chromatyd
chromosomy z nowymi
kombinacjami genów
A
B
A
B
a
b
a
b
B
a
a
b
A
B
A
b
A
B
a
B
A
b
a
b
chromatydy
Rys. 11.
Schemat procesu
crossing-over
Rys. 12.
Przekazywanie
potomstwu
dwóch cech
dziedziczàcych si´
zale˝nie (barwy
i kszta∏tu ziaren)
u kukurydzy
INFORMACJE DODATKOWE
typy rodzicielskie
nowe uk∏ady alleli
A
B
a
b
a
b
a
b
a
b
mejoza
mejoza
x
P
bez wymiany
F
1
z wymianà
gamety
A
B
a
b
A
B
a
b
a
b
a
b
A
B
a
b
a
b
a
b
A
B
lub
A
b
a
b
A
b
B
a
a
b
B
a
{
{
w Êwietle chromosomowej teorii dziedzicznoÊci Morgana? I prawo Mendla (czysto-
Êci gamet) w wi´kszoÊci przypadków jest s∏uszne. Geny stanowiàce grup´ genów
sprz´˝onych nie sà dziedziczone zgodnie z II prawem Mendla. II prawo Mendla jest
s∏uszne jedynie w odniesieniu do genów le˝àcych w ró˝nych chromosomach.
Morgan zak∏ada∏, ˝e zjawisko crossing-over po-
jawia si´ ca∏kowicie przypadkowo. Statystycznie
szansa na wystàpienie tego procesu pomi´dzy dwo-
ma genami zwi´ksza si´ wraz ze wzrostem odle-
g∏oÊci pomi´dzy nimi. Jest ona niewielka dla genów
le˝àcych blisko siebie na chromosomie, natomiast
zdecydowanie wi´ksza dla genów znacznie od sie-
bie oddalonych. Inaczej mówiàc, jeÊli geny le˝à bli-
sko siebie, to liczba komórek, w których nastàpi
crossing-over pomi´dzy nimi, b´dzie mniejsza, ni˝
w przypadku, gdy geny sà rozmieszczone dalej od
siebie. W zwiàzku z tym analiza liczebnoÊci zrekom-
binowanych genotypów w krzy˝ówkach testowych
pozwala na zmapowanie danego chromosomu, czy-
li wyznaczenie w nim po∏o˝enia wzgl´dem siebie
poszczególnych genów. Jest to jedna z metod mapo-
wania chromosomów. Trudno jednak bez zastrze-
˝eƒ przyjàç za∏o˝enia, które stanowià podstawy
takiej metody mapowania. Obecnie stosuje si´ wie-
le innych, bardziej nowoczesnych metod.
Jak zatem zinterpretowaç prawa Mendla
19
A
B
C
Geny A i C le˝à dalej od siebie
ni˝ geny A i B, w zwiàzku z tym
prawdopodobieƒstwo zajÊcia (cz´stoÊç)
crossing-over mi´dzy genami A i C
jest wi´ksze ni˝ mi´dzy A i B.
Rys. 13.
Szansa na zajÊcie
crossing-over pomi´dzy
dwoma genami zwi´ksza si´
wraz ze wzrostem odleg∏oÊci
pomi´dzy nimi
INFORMACJE DODATKOWE
POJ¢CIA
❙
LINIA CZYSTA
❙
PRAWA MENDLA
❙
SZACHOWNICA GENETYCZNA
❙
GRUPA GENÓW SPRZ¢˚ONYCH
❙
MAPA CHROMOSOMOWA
❙
REKOMBINACJA
❙
1.2.
STRUKTURA I FUNKCJE
KWASÓW NUKLEINOWYCH
R
ozwijajàca si´ od poczàtku XX wieku genetyka umo˝liwi∏a poznanie wielu zjawisk
zwiàzanych z dziedziczeniem cech. Nie uda∏o si´ wówczas wyjaÊniç ani moleku-
larnego pod∏o˝a informacji genetycznej, ani te˝ mechanizmów rzàdzàcych powiela-
niem si´ genów i fenotypowym ich przejawianiem si´, czyli
ekspresjà. Dok∏adniejsze
poznanie tych zagadnieƒ sta∏o si´ mo˝liwe dopiero pó∏ wieku póêniej, gdy opisano
struktur´ DNA oraz poznano funkcjonowanie kodu genetycznego.
Odkrywcà DNA jest niemiecki chemik Friedrich Miescher (czytaj: miszer),
pracujàcy w Tybindze. W 1869 roku stwierdzi∏ on, ˝e materia∏ wyizolowany z jà-
der komórek cz∏owieka nie jest bia∏kiem, poniewa˝ zawiera fosfor i jest odporny
na dzia∏anie enzymu rozk∏adajàcego bia∏ka. Odkrytà substancj´ nazwa∏ nukleinà,
a dzisiaj nosi ona nazw´ kwasu deoksyrybonukleinowego, czyli DNA.
INFORMACJE
DODATKOWE
Wyszukiwarka
Podobne podstrony:
java5 dziedziczenie id 226675 Nieznany
Abolicja podatkowa id 50334 Nieznany (2)
4 LIDER MENEDZER id 37733 Nieznany (2)
katechezy MB id 233498 Nieznany
metro sciaga id 296943 Nieznany
perf id 354744 Nieznany
interbase id 92028 Nieznany
Mbaku id 289860 Nieznany
Probiotyki antybiotyki id 66316 Nieznany
miedziowanie cz 2 id 113259 Nieznany
LTC1729 id 273494 Nieznany
D11B7AOver0400 id 130434 Nieznany
analiza ryzyka bio id 61320 Nieznany
pedagogika ogolna id 353595 Nieznany
Misc3 id 302777 Nieznany
cw med 5 id 122239 Nieznany
D20031152Lj id 130579 Nieznany
więcej podobnych podstron