
Fizjologia roślin
Wykłady
Created by Katilla
Zagadnienia:
1. Gospodarka wodna
2. Gospodarka mineralna
3. Fotosynteza
4. Faza jasna fotosyntezy
5. Faza „ciemniowa”
6. Ekologia fotosyntezy
7. Oddychanie
8. Wzrost – regulatory wzrostu
9. Rozwój roślin
10. Ruchy roślin i allelopatia
11. Reakcja roślin na czynniki stresowe

Fizjologia roślin
Wykłady
Created by Katilla
1
Gospodarka wodna
Fizjologia roślin – zajmuje się poznaniem zjawisk zachodzących organizmach roślin. Zajmuje się wpływem
czynników środowiska. Traktuje rośliny jako integralną całość.
Funkcje wody w roślinie:
Rozpuszczalnik wszystkich związków organicznych
Woda wchodzi w reakcje (np. hydroliza, fotosynteza) – substrat
Produkt reakcji
Woda tworzy otoczkę hydratacyjną, szczególnie bardzo ważna w błonach lipidowo – białkowych
Decyduje o turgorze komórki
Czynnik wzrostu
Udział w transporcie soli mineralnych i związków organicznych
Ochładzanie rośliny przez transmitancję
Woda jest dipolem. Powoduje to że woda może być przyciągana i odpychana przez różne cząsteczki
polarne. Tworzą się wiązania wodorowe.
Wiązania wodorowe pomiędzy cząsteczkami umożliwiają tworzenie się agregatów o uporządkowanej
strukturze pseudokrystalicznej, ale trwają one krótko.
Woda występująca w komórce tworzy grona – struktury powiązane wiązaniami wodorowymi – ulegają one
rozbiciu lub połączeniu.
Polarne właściwości wody powodują to że jest doskonałym rozpuszczalnikiem dla związków
zjonizowanych. Tworzą otoczki wokół jonów i cząsteczek hydrofilowych
Właściwości fizyko – chemiczne i fizjologiczne wody:
1. Woda jest cieczą o szerokim zakresie temperatur
2. Ma wysokie ciepło parowania, chroni roślinę zatem przed gwałtownymi zmianami temperatury.
Dzięki dwubiegunowemu charakterowi cząsteczki mogą się łączyć wiązaniami wodorowymi co
powoduje, że woda ma wysokie ciepło parowania. Zmiana 1g wody o temp. 20
o
C w parę wodną
wymaga nakładu energii cieplnej w ilości ok. 2256 J
3. Ma wysokie ciepło topnienia, co wpływa istotnie na tworzenie wiązań wodorowych między jej
cząsteczkami
4. Posiada wysoką stała dielektryczną, co umożliwia jej zobojętnienie ładunków
5. Zjawiska adhezji i kohezji
Dyfuzja – to proces samorzutnego rozprzestrzeniania się cząsteczek lub energii w danym ośrodku (np. w
gazie, cieczy lub ciele stałym) będący konsekwencją chaotycznych zderzeń cząsteczek dyfuncującej
substancji między sobą lub z cząsteczkami otaczającego ją ośrodka.
Selektywność błon
Błona półprzepuszczalna pozwala na przenikanie cząsteczek rozpuszczalnika, a zatrzymuje cząsteczki
substancji rozpuszczonej.
Fizjologia roślin
Wykłady
Created by Katilla
2
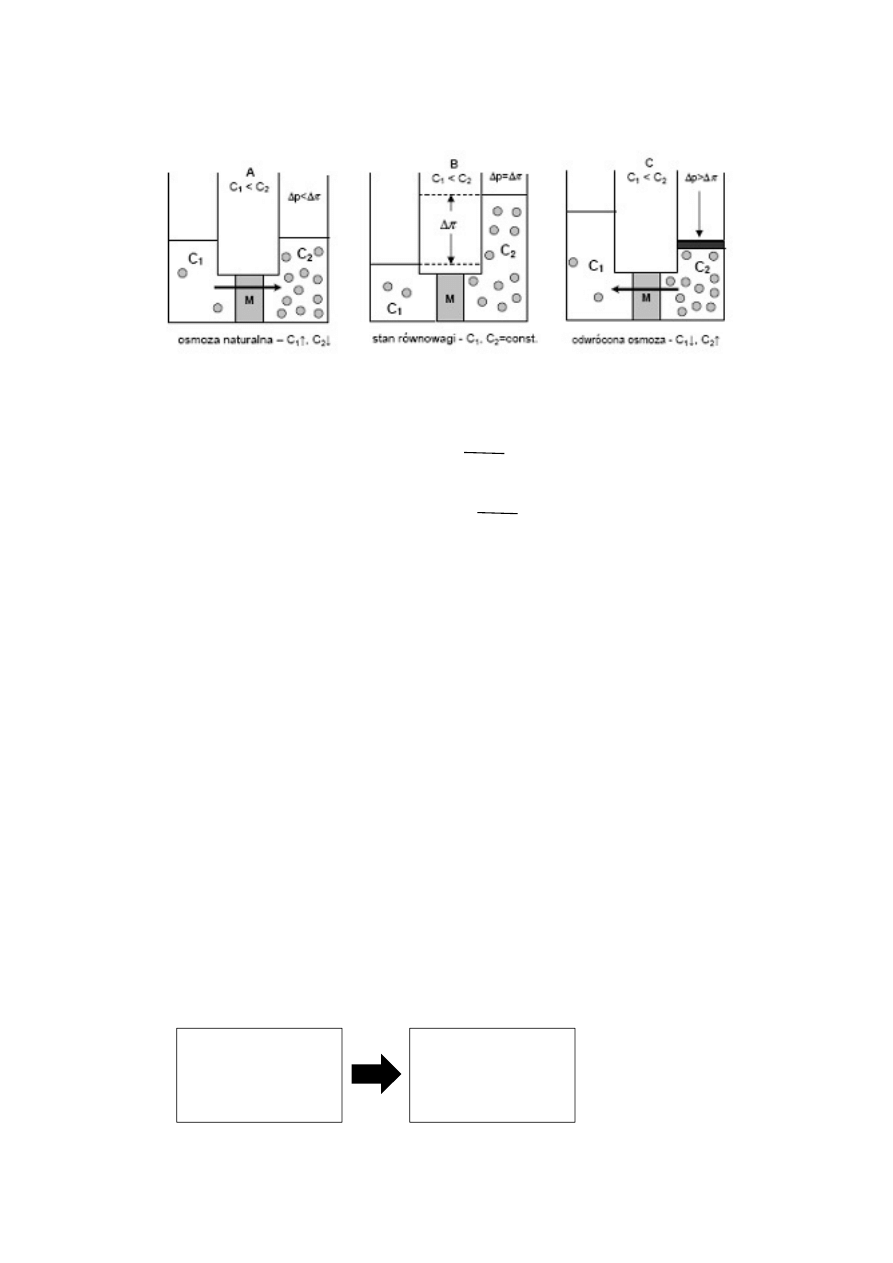
Osmoza – specyficzny rodzaj dyfuzji przez błonę półprzepuszczalną.
Wzory na ciśnienie osmotyczne
Nie elektrolitów:
П =
Elektrolitów:
П =
n – liczba moli
R – stała gazowa = 8,3143 Jmol
-1
T – temperatura bezwzględna
V – objętość
Woda przechodzi w roślinie za pomocą specyficznych kanałów wodnych – są to podwójne łańcuchy
polipeptydowe, które przechodzą w poprzek błony. Transport błony jest regulowany przez fosforylację i
defosforylację reszt fosforowych.
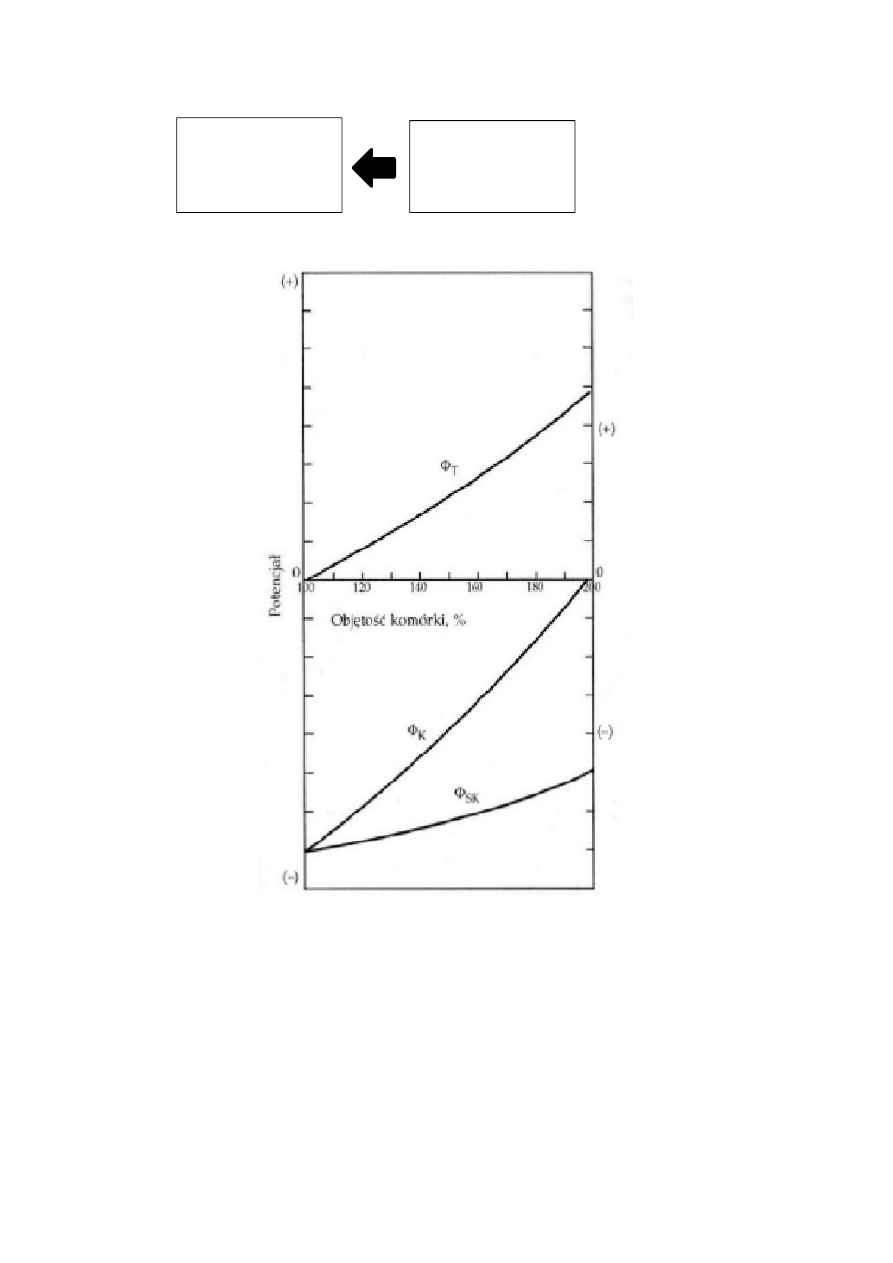
Parametry osmotyczne komórki roślinnej decydujące o pobieraniu wody:
1. Ψ
ok
– potencjał osmotyczny wyrażony w MPa zawsze ze znakiem „ – ”
2. Ψ
p
– potencjał ciśnieniowy (ciśnienie turgorowe) wyrażone również w MPa, ale zawsze ze znakiem
„ + ”
3. Ψ
m
– potencjał matrycowy, nazywany potencjałem inhibicyjnym. Ma on znak „ – ”, ale w zjawiskach
osmotycznych nie odgrywa większej roli i jego wartość można pominąć w wyliczaniu potencjału
chemicznego wody w komórce roślinnej (Ψ
wk
)
Zatem potencjał wody komórki roślinnej można zapisać przy pomocy wzoru:
Ψ
wk
= (- Ψ
ok
) + Ψ
p
Przepływ wody pomiędzy komórkami:
nRT
V
nRT
V
k
Komórka A
Ψ
ok
= - 1 MPa
Ψ
p
= 0,5 MPa
Ψ
wk
= - 0,5 MPa
Komórka B
Ψ
ok
= - 1,2 MPa
Ψ
p
= 0,5 MPa
Ψ
wk
= - 0,7 MPa
Fizjologia roślin
Wykłady
Created by Katilla
3
Aby był w roślinie stały ciąg wodny musi być duża różnica potencjału wodnego.
Komórka
Ψ
ok
= - 1 MPa
Ψ
p
= 0,5 MPa
Ψ
wk
= - 0,5 MPa
Roztwór glebowy
Ψ
o
= - 0,2 MPa = Ψ
w
ponieważ:
Ψ
p
= 0
Fizjologia roślin
Wykłady
Created by Katilla
4
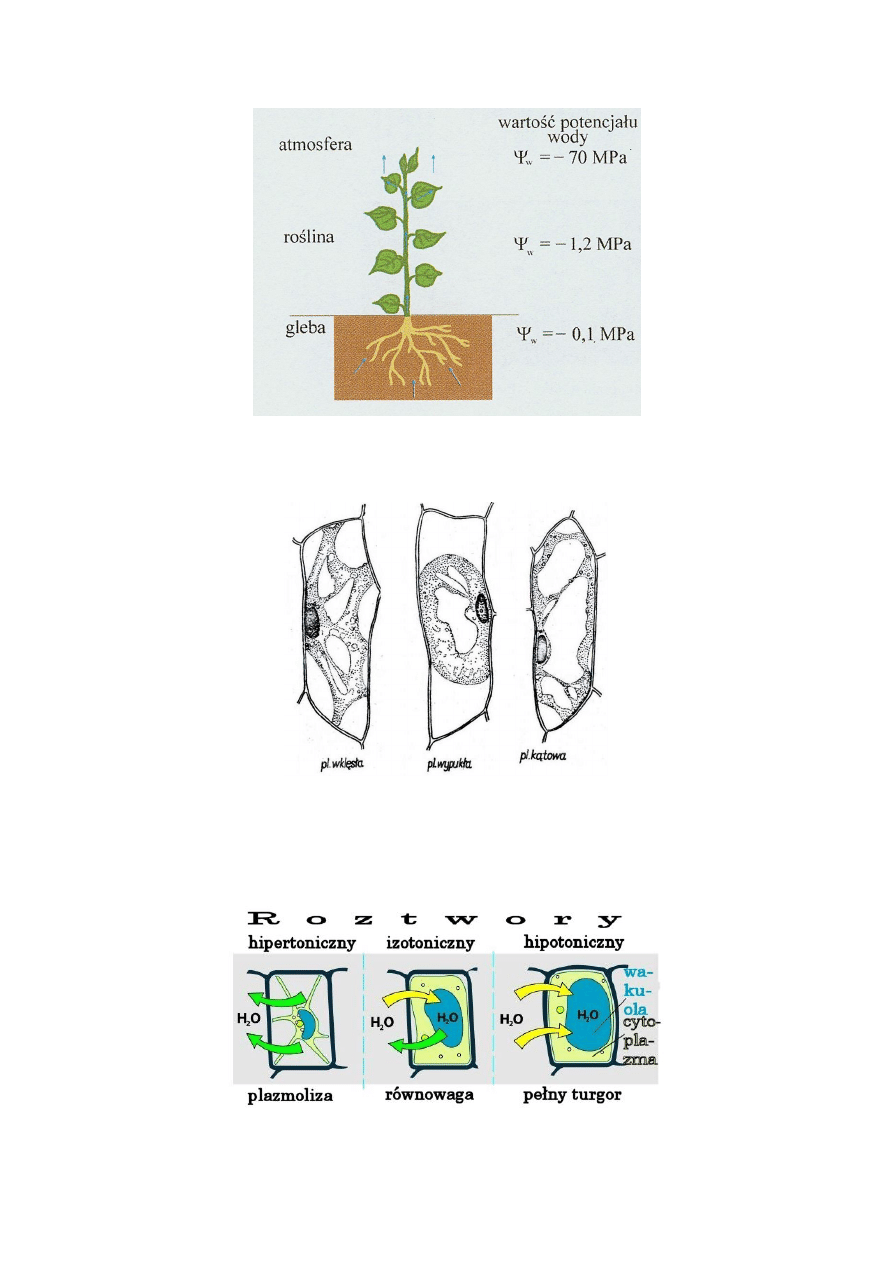
Plazmoliza - proces tracenia wody w komórce w roztworze hipertonicznym. W wyniku tego następuje
obkurczenie cytoplazmy od ścian komórki. Dotyczy ona wyłącznie komórek roślinnych.
Rodzaje roztworów
Roztwór izotoniczny
Roztwór hipertoniczny
Roztwór hipotoniczny
Fizjologia roślin
Wykłady
Created by Katilla
5
Deplazmoliza – komórka splazmolizowana ulega deplazmolizie tylko wtedy, gdy protoplast nie traci
kontaktu ze ściana komórkową.
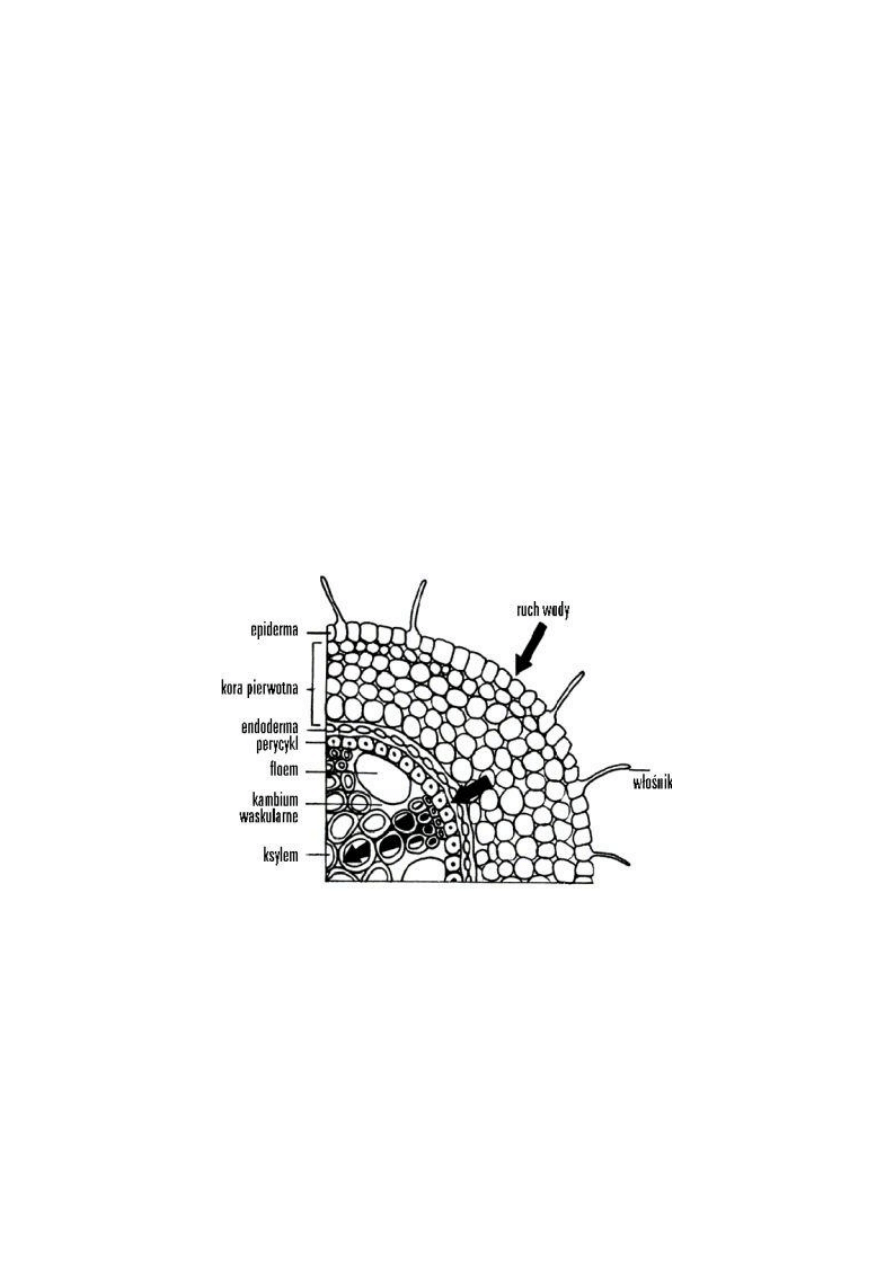
Transport wody bliski w korzeniu
Drogi przemieszczania H
2
O w roślinie do naczyń i cewek w wiązkach przewodzących:
1. Droga osmotyczna – zgodnie z gradientem potencjału wody soku komórkowego
2. Apoplastem – w obrębie ścian i przestrzeni międzykomórkowych
3. Symplastem – przez cytoplazmę komórek połączonych plazmodesmami (w obrębie tkanek). Przez
kanały wodne w błonach (akwaporyny)
Akwaporyny – integralne białka błonowe umożliwiające transport wody
przez półprzepuszczalne błony
komórek organizmów żywych.
Natomiast transport daleki dotyczy tylko transportu naczyniami lub cewkami czyli w drewnie od korzenia
do organów rośliny, czyli tam gdzie jest potrzebna adhezja, kohezja i siła ssąca.
kanał symplastyczny – przenikanie wody z cytoplazmy jednej komórki do cytoplazmy drugiej
komórki przez plasmodesmy (pasma cytoplazmy przechodzące przez jamki w ścianie komórkowej)
kanał apoplastyczny – transport wody w obrębie ścian komórkowych i kapilar w ścianach
komórkowych
Komórki budujące warstwę endodermy mają pośrodku ścian pasemkowate zgrubienia zwane
pasemkami Caspary’ego
Zawierają one nie przepuszczające wodę substancję tłuszczową zwaną suberyną. Endoderma,
dzięki takiej budowie ścian komórkowych, reguluje przemieszczanie się wody w poprzek korzenia,
od włośników do wiązek przewodzących. Woda dociera do śródskórni przepływając „kanałami”
zbudowanymi przez ściany komórkowe i przestrzenie międzykomórkowe skórki i kory pierwotnej,
nie wnikając do wnętrza mijanych komórek. Nieprzepuszczalne, suberynowe wyściółki ścian
endodermy stanowią dla niej nieprzeniknioną barierę.
Transport ksylenowy napędza transpiracja.
Gdy nie ma liści to działa parcie korzeniowe.

Fizjologia roślin
Wykłady
Created by Katilla
6
Parcie korzeniowe – to transport aktywny jonów do ksylemu, podczas którego potrzebna jest energia w
postaci ATP z oddychania. Poprzez transport jonów do ksylemu obniża się potencjał wodny w ksylemie i
napływa do niego woda. Jest to fizjologiczne zjawisko u roślin polegające na wypieraniu wody przez
korzenie do łodyg i liści. Przyczynia się ono do krążenia soków roślinnych, zwłaszcza przed pojawieniem się
liści, kiedy niemożliwa jest transpiracja. Zwykle osiąga wartość 0,2 – 0,3 MPa, wyjątkowo osiąga 0,9 MPa.
Parcie korzeniowe można zaobserwować uszkadzając roślinę – przez pewien czas z miejsca nacięcia
wycieka roztwór. Zjawisko to nazywa się płaczem roślin. Przykładem działania parcia korzeniowego u roślin
nieuszkodzonych jest gutacja.
Gutacja – zjawisko wydzielania kropel wodnych roztworów związków
organicznych i soli mineralnych przez rośliny i grzyby. U roślin
naczyniowych krople wydzielane są przez specjalne struktury
umieszczone na krańcach blaszki liściowej, zwane hydatodami. Gutacja
następuje, gdy roślina ma dostęp do dużych ilości wody w glebie oraz w
warunkach środowiskowych niesprzyjających transpiracji (wysoka
wilgotność powietrza) wskutek parcia korzeniowego. Pomaga pozbyć
się nadmiaru turgoru i niektórych soli. Występuje zwłaszcza u młodych
roślin, nasturcji, zbóż, kapusty, pokrzywy.
W naczyniach ksylenu nieprzerwany słup wody, gdy naczynia zablokowane to woda transportowana
ścianami.
Szybkość przepływu wody w ksylemie zależy od czynników wpływających na transpirację.
Pompa protonowa – integralne białko błonowe, zdolne do transportu protonów (jonów wodorowych H+)
przez błony biologiczne przeciwnie do ich gradientu stężenia. Proces transportu jest procesem
wymagającym energii.
Miłość po grób – gdzie potas tam i woda
K
+
+ H
2
O =
Czynniki wpływające na pobieranie wody przez rośliny:
1. Czynniki glebowe:
Dostępność wody fizjologicznie użytecznej
Temperatura gleby
Natlenienie gleby
Zasolenie gleby
2. Czynniki transpiracyjne (wpływające na intensywność transpiracji):
Opór dyfuzyjny warstwy granicznej – zależy od powierzchni liścia (r
g
)
Opór dyfuzyjny kutikuli (r
k
)
Opór dyfuzyjny szparek (r
sz
) – zależy od liczby i stopnia rozwartości aparatów
szparkowych
T
r
=
blablabla
Światło
ΔΨ
w
r
sz
+ r
k
+ r
g
Fizjologia roślin
Wykłady
Created by Katilla
7
Temperatura
Różnica potencjałów wody w liściu i powietrza – Δ Ψ
w
, nazywana często niedosytem
wilgotności
Ruchy powietrza
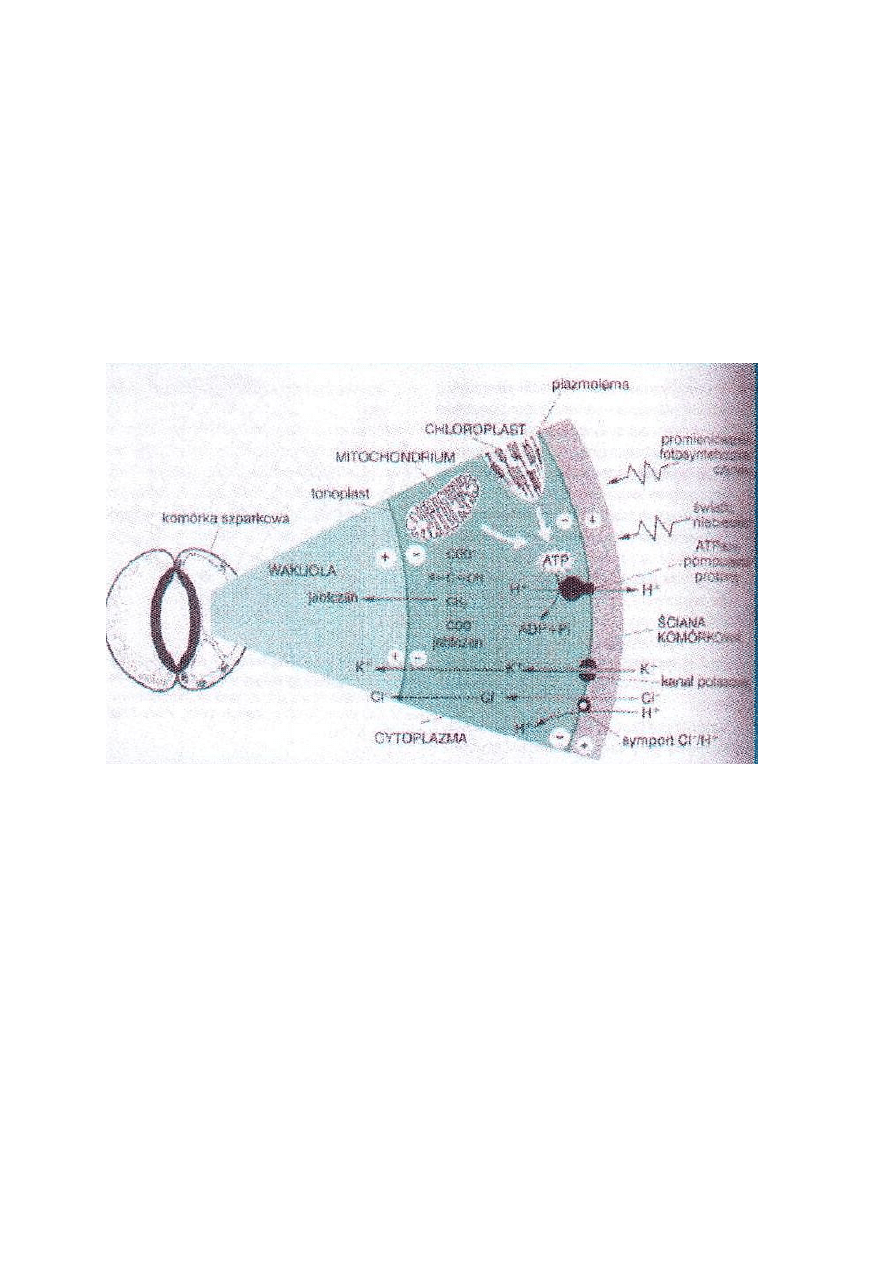
Aparaty szparkowe są w epidermie, mają chloroplasty, głównie po dolnej stronie liścia, najlepiej, aby było
dużo małych, regulacja rozwarcia aparatu szparkowego przez światło ok. 500 nm (niebieskie), które
pochłaniane przez fototropinę lub zeaksantynę, następuje pompowanie wodoru w poprzek błony, jony
potasu wchodzą do komórek wakuoli komórek szparkowych, może też przechodzić jabłczan do wakuoli,
obniża się potencjał wodny komórek szparkowych i napływa woda, komórka powiększa się i rozwiera się
aparat szparkowy.
Na aktywne zamykanie aparatów szparkowych wpływa kwas abscysynowy (ABA), który jest hormonem
stresowym, reaguje on ze specyficznym receptorem i ciąg transdukcyjny i zostaje uwolniony wapń, który
reaguje z kalmoduliną, otwiera wylotowe kanały potasowe i wapń wychodzi.
Zamykanie hydropasywne występuje w czasie południa, gdy duża transpiracja i brakuje wody.
Efekt brzeżny.
Intensywność dyfuzji gazu przez otwory jest proporcjonalna do ich średnicy a nie do ich powierzchni
(prawo Stephena 1881).
Rodzaje transpiracji:
Transpiracja szparkowa – transpiracja zachodząca poprzez aparaty szparkowe. Para wodna
początkowo znajdująca się w przestrzeniach międzykomórkowych przedostaje się do komory
powietrznej i na drodze dyfuzji, przez szparki wydostaje się na zewnątrz. Udział tego rodzaju
transpiracji jest zwykle wysoki – 80% transpiracji ogółem.
Transpiracja kutikularna – transpiracja, która zachodzi poprzez zewnętrzną powierzchnię liścia,
dodatkowo pokrytego tłuszczową warstwą ochronną kutykulą. W większości przypadków ma

Fizjologia roślin
Wykłady
Created by Katilla
8
niewielki udział w ogólnej transpiracji. Kutykula nie jest szczelna lecz umożliwia parowanie wody za
sprawą imbibicji. Ilość przepuszczanej wody zależy od:
Jednostki transpiracji:
1. Intensywność transpiracji to ilość wytranspirowanej wody na jednostkę powierzchni lub masy
liścia w jednostce czasu (g H
2
O
*
dm
-1
*
h
-1
lub g H
2
O
*
g św m
-1
*
h
– 1
)
2. Współczynnik transpiracji to ilość wody wytranspirowanej w stosunku do przyrostu suchej masy
rośliny. Wartość waha się w granicach 100 – 1 000.
Współczynnik transpiracji nie jest wartością stałą dla danego gatunku rośliny, zależy od:
poziomu wilgotności gleby – zwiększa się, gdy suchsza,
temperatury,
zasobności gleby w składniki mineralne – zmniejsza się wraz ze wzrostem zasobności,
rodzaju gleby – zwiększa się, dla lżejszych gleb,
kwasowości gleby
3. Współczynnik produktywności transpiracji to ilość wytwarzanej wody (g suchej masy x 1 000 g
H
2
O). Wartość waha się w granicach 1 – 8.
4. Współczynnik transpiracji względnej to stosunek ilości wody wytranspirowanej przez jednostkę
powierzchni liścia do ilości wody wyparowanej w takiej samej wolnej powierzchni lustra wody.
Wartość waha się w granicach 0,01 – 0,3 a nawet 0,8.
Zawartość wody w roślinie:
W tkankach o dużej aktywności metabolicznej (80 – 90%)
Chloroplasty i mitochondria (ok. 50%)
Owoce soczyste (85 – 90%)
Liście (80 – 90%)
Korzenie (70 – 90%)
Pień drzewa (ok. 50%)
Nasiona dojrzałe (10 – 15%)
Nasiona roślin oleistych (5 – 7%)
Bilans wodny u roślin w ciągu dnia jest lekko ujemny.
Rośliny dzielone przez wzgląd na bilans wodny:
Hydrostabilne – izohydryczne, małe floktuacje bilansu wodnego, sprawne aparaty szparkowe,
dobry system korzeniowy
Hydrolabilne – anizohydryczne, duże zmiany zawartości wody, bilans wody przez długi czas
ujemny, słaboreagujące aparaty szparkowe, rośliny występujące na obszarach objętym dużym
promieniowaniem słonecznym
Woda dostępna dla roślin:
Kapilarna
Grawitacyjna
Gruntowa
Pojemność wodna – zdolność do zatrzymywania wody:
Maksymalna – wszystkie przestrzenie pomiędzy cząsteczkami gleby są wypełnione wodą
Połowa – wszystkie kapilary wypełnione wodą, ale brak wody gruntowej
Wilgotność trwałego więdnięcia określa ilość wody niedostępnej dla roślin

Fizjologia roślin
Wykłady
Created by Katilla
9
Woda niedostępna:
Krystalizacyjna – w siatce minerałów 600
o
C
Higroskopowa – zatrzymana na powierzchni koloidów 105
o
C
Błonkowata – tworzy kolejne warstewki wody wokół koloidów glebowych 105
o
C
Trwałe więdnięcie – stan rośliny spowodowany brakiem wody, przy którym nie odzyskuje ona turgoru
nawet po przeniesieniu jej do ciemności i atmosfery wysyconej parą wodną.
Współczynnik trwałego więdnięcia – ilość wody w glebie, czyli wilgotność gleby, przy której rośliny
zaczynają nieodwracalnie więdnąć. Wyróżnia się:
punkt (wilgotność) trwałego więdnięcia roślin – to wilgotność gleby przy której rozpoczyna się
więdnięcie rośliny, przy krótkotrwałej suszy więdnięcie jest przejściowe i po nawodnieniu wody
roślina może odzyskać turgor.
całkowite hamowanie wzrostu roślin
początek hamowania wzrostu roślin
Współczynnik więdnięcia występuje na poszczególnych typach gleb przy różnej zawartości wody. Zależy on
od siły ssącej kapilar, a ta z kolei uzależniona jest od składu mechanicznego, zawartości koloidów
organicznych i mineralnych, czy struktury gleby. Wzrasta on w miarę zwiększania się zawartości części
spławialnych w glebie.
Susza fizjologiczna – okres, w którym roślina nie może pobierać wody z otoczenia, mimo iż woda tam
występuje. Powodem tego może być zbyt niska temperatura powietrza, zbyt duże zasolenie gleby lub też
jej zbyt małe napowietrzenie (zbyt mało tlenu). Susza fizjologiczna występuje najczęściej późną jesienią,
zimą i wczesną wiosną. Niektóre rośliny są odpowiednio przystosowane do radzenia sobie w czasie suszy –
takie przystosowanie to kseromorfizm.
Zastosowanie zbyt dużej ilości nawozu powoduje hipertonię roztworu glebowego i suszę fizjologiczną.

Fizjologia roślin
Wykłady
Created by Katilla
10
Gospodarka mineralna
Pierwiastki niezbędne dla roślin:
1. Organogeny:
Węgiel
Wodór
Tlen
Pobierane w postaci CO
2
, wody i tlenu atmosferycznego
2. Makroelementy (makropierwiastki):
Fosfor
Azot
Potas
Siarka
Wapń
Magnez
3. Mikroelementy (pierwiastki śladowe):
Żelazo
Mangan
Miedź
Cynk
Bor
Molibden
Makroelementy – więcej niż 1% suchej masy.
Reutylizacja:
Wycofywanie pierwiastków z organów starszych, bądź starzejących się do liści młodych lub
organów zimujących
Ruchliwe pierwiastki – N, P, K, Mg, Zn
Ogólne objawy niedoboru:
Chlorozy
Nekrozy
Przebarwienia
Śluzowacenie
Brak kwitnienia lub osłabione kwitnienie
Zahamowany wzrost
AZOT
Rola azotu:
Składnik białek, kwasów nukleinowych, nukleotydów (ATP, NAD, NADP, FAD, FMN) przenoszących
energię, transportujących wodór i elektrony, CoA bierze udział w procesie oddychania i syntezie
tłuszczów, amidów
Chlorofile i cytochromy uczestniczą w oddychaniu i fotosyntezie.
Pozostałe związki organiczne pełniące wiele funkcji fizjologicznych.

Fizjologia roślin
Wykłady
Created by Katilla
11
Pobieranie azotu:
W postaci mocznika (CO(NH
2
)
2
, jonów NH
4
+
, NO
3
-
oraz azot atmosferyczny, dzięki symbiozie z
bateriami.
Rośliny wydzielają ureazę, enzym powodujący rozkład mocznika do amoniaku i dwutlenku węgla.
Transport przez ksylem i floem
Niedobór azotu – rośliny są słabe, wolno rosną, są dużo bledsze, słabo kwitną i zawiązują małe nasiona.
FOSFOR
Rola fosforu:
Strukturalna – fosfolipidy budują błony komórkowe, nukleotydy, kwasy nukleinowe
NADP i NAD – biorą udział w fotosyntezie i oddychaniu
ATP – nośnik energii
FAD, FMN – procesy oddychania
Ufosforylowane formy są aktywne i mogą wchodzić w reakcje
Pobieranie fosforu w postaci jonu ortofosforanowego. Transport przez ksylem i floem.
Niedobór fosforu:
Silnie zahamowany wzrost części nadziemnych
Odrzucanie liści
Niewiele pędów bocznych
Zamieranie pączków
Słabe kwitnienie
Niski plon
W przypadku deficytu azotu i fosforu dochodzi do: upośledzenia syntezy aminokwasów, a nadmiar cukrów
roślina przeznacza na syntezę antocyjanów, powodujących czerwone zabarwienie łodyg (brak fosforu) lub
liści (brak azotu).
POTAS
Rola potasu:
Reguluje gospodarkę wodną (parcie korzeniowe, turgor, otwieranie aparatów szparkowych)
Bierze udział w syntezie ATP
Aktywator ponad 50 enzymów
Wpływa na syntezę cukrów, aminokwasów
Poprawia zimotrwałość roślin i zwiększa odporność na choroby. Niedobór potasu – upośledza
syntezę białka, a akumulacja aminokwasów zwiększa ryzyko ataku ze strony patogenów
odżywiających się aminokwasami, jako dobrym źródłem azotu.
Pobór potasu – w postaci jonów K
+
, bardzo szybko jest pobierany i transportowany, wymaga do tego
nakładu energii, zatem brak węglowodanów obniża zawartość potasu w roślinie.
Objawy niedoboru potasu:
Zwiędły pokrój rośliny – brak turgoru
Liście matowe, niebieskozielone, z chlorozą między żyłkami, brązowienie wierzchołków liści,
zasychanie brzegów liści
Skrócone międzywęźla, słabo rozwinięty system korzeniowy, zmniejszony plon

Fizjologia roślin
Wykłady
Created by Katilla
12
WAPŃ
Rola wapnia:
Reguluje odczyn gleby, poprawia strukturę biokoloidów
Spalają blaszkę środkową (sole kwasu pektynowego)
Kofaktor enzymów
Składnik pektyn i ściany komórkowej – stabilizuje błony komórkowe
Tworzy kompleksy z różnymi białkami
Reguluje uwodnienie cytoplazmy (jest antagonistą potasu)
Pełni funkcję sygnałową w komórce – wtórny przenośnik informacji w regulacji metabolizmu (z
kalmoduliną)
Otwiera aparaty szparkowe
Fityna – związek zapasowy magnezowo – wapienno - fosforowy
Pobór wapnia – powoli w postaci jonów Ca
2+
, chelatów. Transportowany przez ksylem.
Niedobory wapnia:
Sucha zgnilizna pomidorów
Zahamowanie wzrostu
Śluzowacenie korzeni
Słabo rozwinięty system korzeniowy
Zasychanie wierzchołków liści i kwiatostanów
Brązowe plamy na liściach lub jasne smugi
MAGNEZ
Rola magnezu:
Buduje chlorofil
Spaja podjednostki rybosomów
Buduje blaszkę środkową
Wchodzi w skład fityny
Wchodzi w skład chelatów, jest kofaktorem wielu enzymów (pomaga przyłączyć się enzymowi do
substratu) głównie uczestniczących w przenoszeniu reszt fosforanowych
Aktywuje enzymy szlaku glikolizy, cyklu Krebsa, przemiany związków azotowych
Aktywuje główne enzymy fosforylacyjne
Rola magnezu
Chelaty:
Chlorofil
Układy hemowe – cytochromy, katalaza
Kobalamina – witamina B
12
Barwniki kwiatów
Enzymy
Pobór magnezu – w postaci jonu Mg
2+
. Transport przez floem i ksylem.
Objawy niedoboru magnezu:
Chlorozy międzyżyłkowe
Plamy nekrotyczne
Zahamowanie wzrostu szczególnie korzeni

Fizjologia roślin
Wykłady
Created by Katilla
13
„tygrysowatość” lub „paciorkowatość” liści
SIARKA
Rola siarki:
Tworzy reaktywne grupy tiolowe – SH oraz mostki siarczkowe
Występuje w metioninie, cysteinie i cystynie, glutationie, w koenzymie A, w ferredoksynie
Wzmacnia strukturę białek
Jest w olejkach gorczycznych o charakterze bakteriobójczym
Pobór siarki – w postaci jonu SO
4
2+
. Transport przez floem i ksylem.
Objawy niedoboru:
Rzadko występują
Liście drobne, korzenie białe słabo rozwinięte
Chloroza liści
ŻELAZO
Rola żelaza:
Stymulator syntezy chlorofilu
Kofaktor wielu enzymów oksydo – redukcyjnych (katalazy, peroksydazy, dysmutazy anionorodnika
ponadtlenkowego, cytochromów, ferredoksyny, reduktazy azotanowej)
Bierze udział w transporcie elektronów, w procesach fotosyntezy, oddychania, asymilacji azotu
Ok. 80% żelaza znajduje się w chloroplastach
Pobieranie żelaza:
Jony żelaza Fe
2+
i Fe
3+
Trudno przyswajalne zwłaszcza w środowisku zasadowym, nadmiar wapnia ogranicza pobór żelaza
Podaje się roślinom głównie w postaci chelatów
Objawy niedoboru – chloroza, bielenie liści młodszych.
MANGAN
Rola manganu:
Kofaktor enzymów (łączy się z nimi chelatowo), enzymy cyklu Krebsa, enzymy redukcji azotanów
Aktywator dekarboksylaz, dehydrogenaz
Bierze udział w fotodysocjacji wody
Występuje w kompleksie PS II – białko, w kwaśnej fosfatazie
Pobór magnezu – jako jony dwu – , trój – lub czterowartościowe. W postaci chelatów. W wodzie się
rozpuszcza tylko jon dwuwartościowy, pozostałe jony tworzą nierozpuszczalne tlenki.
Niedobór manganu:
Chloroza miękiszu liściowego
Nekroza międzyżyłkowa
Chloroza mozaikowa. Najbardziej wrażliwy sposób, brak manganu wywołuje szarą plamistość

Fizjologia roślin
Wykłady
Created by Katilla
14
MIEDŹ
Rola miedzi:
Wchodzi w skład enzymów oksydaz przenoszących elektrony na tlen cząsteczkowy – oksydazy
cytochromowej w łańcuchu oddechowym, oksydazy polifenolowej utleniającej fenole do
chinonów, oksydazy askorbinowej
Wchodzi w skład plastocyjaniny transportującej elektrony w czasie fotosyntezy
Wchodzi w skład dysmutazy anionorodnika ponadtlenkowego
Występuje głównie w chloroplastach
Pobieranie miedzi:
Tylko w postaci jonu Cu
2+
związanego silnie z kompleksem sorpcyjnym, na glebach organicznych
bagiennych jest praktycznie niedostępna (choroba nowin)
Odczyn zasadowy zmniejsza dostępność miedzi.
Objawy niedoboru miedzi – wierzchołki młodych liści bieleją i zasychają, są skręcone i wąskie. Zboża nie
kłoszą się. Niebieskozielona barwa liści.
BOR
Rola boru:
W gospodarce węglowodanowej – transport cukrów oraz wbudowanie cukrów strukturalnych w
ścianę komórkową
Uczestniczy w tworzeniu ściany komórkowej
Kontroluje procesy kwitnienia i zapylania
Pobór boru:
Jako anion BO
3
+
, B
4
O
7
2-
, H
2
BO
3
Nadmierne wapniowanie obniża dostępność boru
Objawy niedoboru boru:
Zahamowanie wzrostu, obumieranie stożków wzrostu
Kędzierzawość liści, pękanie organów
Słabe kwitnienie, niezdolność do zapłodnienia ((łagiewka pyłkowa nie kiełkuje)
MOLIBDEN
Rola molibdenu:
W enzymach biorących udział w przemianie azotanów – nitrogeneza, reduktazy azotanowe.
Najwięcej molibdenu potrzebują rośliny motylkowe.
Pobór molibdenu:
W postaci jonów MoO
4
+
Odczyn zasadowy zwiększa jego dostępność
Objawy niedoboru:
Ślepnięcie kalafiorów
U motylkowatych liście bledną, brzegi się zwijają, chlorotyczna nekroza

Fizjologia roślin
Wykłady
Created by Katilla
15
NIKIEL
Pobierany w postaci jonów Ni
2+
Składnik ureazy i hydrogenaz
CHLOR
Pobierany w postaci Cl
-
Konieczny w reakcjach uwalniania tlenu w chloroplastach podczas rozkładu wody
Niedobór: chloroza i nekroza liści, redukcja wzrostu liści, w warunkach naturalnych nie występuje
CYNK
Pobierany w postaci Zn
2+
i chelatów
Składnik dehydrogenazy alkoholowej, anhydrazy węglanowej
Rośliny wskaźnikowe:
Kalafior i kapusta wrażliwe na niedobór N i Ca
Rzepak, rzepa i lucerna – wrażliwe na P
Ziemniak i lucerna – na K
Kalafior i ziemniak – na Mg
Owies – na Mn
Burak, pomidor i słonecznik – wrażliwe na brak B
Jony wymienne mogą pochodzić z kompleksu sorpcyjnego do roztworu glebowego, a na ich miejsce
wchodzą inne jony o takim samym ładunku.
CO
2
+ H
2
O → H
2
CO
3
→ H
+
+ HCO
3
-
Za kationy roślina „płaci” protonami, a za aniony jonami – OH i HCO
3
-
.
Proces bierny transportu jonów przez błonę odbywa się na drodze dyfuzji, która może być:
1. Prosta – zachodzi przy spełnionych warunkach umożliwiających przecieg tego fizycznego procesu
aż do osiągnięcia stanu równowagi dynamicznej
2. Złożona – gradient stężeniowy + gradient potencjału elektrycznego
3. Ułatwiona – zachodzi przy udziale przenośników – permeaz. Przenośnik podobnie jak enzym w
znaczący sposób obniża bariery energetyczne związane z dehydratacją i ponowną hydratacją
jonów transportowanych przez błonę. Przenośnikiem mogą być specyficzne jonofory.
Jony niewymienne
Większość substancji mineralnych, które występują w glebie, posiadają postać związków
nierozpuszczalnych w wodzie. Minerały te nie rozpuszczają się w niej, ponieważ posiadają jony
niewymienne, czyli jony, które nie ulegają wymianie. Związki te są dostępne dla roślin dzięki procesom
wietrzenia i rozpuszczania, które są przyspieszane poprzez substancje o charakterze kwaśnym (np. kwas
jabłkowy, szczawiowy i węglowy) wydzielane przez korzeń rośliny.

Fizjologia roślin
Wykłady
Created by Katilla
16
Pobór jonów przez roślinę:
wraz ze strumieniem transpiracyjnym
wypełniają apoplast i natrafiają na wpływ blaszki środkowej
Równowaga Donnana (równowaga membranowa) – równowaga ustalająca się między 2 roztworami
przedzielonymi membraną półprzepuszczalną.
Białka nie mogą przenikać przez błony półprzepuszczalne, ponieważ są wielkocząsteczkowymi koloidami,
jednak dzięki występowaniu w formie jonów (kationów lub anionów) wpływają na rozmieszczenie
elektrolitów dyfundujących przez błony komórkowe. W przestrzeni oddzielonej błonami
półprzepuszczalnymi, niepozwalającymi na przechodzenie np. anionów białczanowych, zasada
elektroobojętności zostaje zachowana w inny sposób. Ze strony, w której znajdują się jony koloidalne,
niezależnie od znaku, po tej stronie jest mniej jednoimiennych jonów elektrolitu, a więcej jonów o innym
znaku, w porównaniu do roztworu po drugiej stronie błony. Zjawisko to polega więc na nierównomiernym
rozmieszczeniu dyfundujących jonów elektrolitu, zależnym od stężeń jonów koloidalnych po obu stronach
błony biologicznej. Odgrywa istotną rolę w procesach zachodzących w organizmach żywych.
Transport bliski:
w obrębie kilku komórek
mogą wykorzystywać kanaliki retikulum endoplazmatycznego
Pasywny transport jonów przez błony:
kanały białkowe
nośniki białkowe
Aktywny transport
symport
antysport
Symport – to jeden z rodzajów transportu aktywnego zachodzących wewnątrz organizmów żywych,
podczas którego przez jedno białko transportowane są dwie cząsteczki jednocześnie (w tym samym
kierunku).
Antyport – forma transportu aktywnego wtórnego przez błony biologiczne. W antyporcie przemieszczenie
jednej substancji (jonu lub cząsteczki, np. metabolitu) do wnętrza określonego przedziału zachodzi
równocześnie z usuwaniem drugiej substancji z tego przedziału. Przykładem antyportu jest pompa sodowo-
potasowa lub wymiennik sodowo-wapniowy.
Uniport – kationy do środka, aniony na zewnątrz
Transport daleki jonów – odbywa się ksylemem.
Za transport jonów może być odpowiedzialnych kilka przenośników o różnym K
m
.
jon potasu – 2 przenośniki
transport N – NH
4
– 3 przenośniki
podobnie jest z transportem NO
3
-
i H
2
PO
4
-
W drzewach i roślinach rosnących na grubość występuje transport jonów w poprzek.
Następne przenośniki mają coraz mniejsze
powinowactwo do jonów

Fizjologia roślin
Wykłady
Created by Katilla
17
Wpływ podwyższonego poziomu składnika mineralnego w roztworze glebowym na pobieranie innych
składników:
Pierwiastek
Jony, których pobieranie jest hamowane
NH
4
+
NO
3
-
, Mg
2+
, Ca
2+
NO
3
-
SO
4
3-
, Cl
-
HPO
4
2-
Fe
2+
, Zn
2+
K
+
Ca
2+
, Mg
2+
, Na
+
i inne
Ca
2+
Fe
2+
, Mn
2+
, Zn
2+
Czynniki środowiska wpływające na odżywianie mineralne rośliny:
Stężenie roztworu glebowego
pH podłoża
temperatura – niska temperatura hamuje pobór składników mineralnych, zwłaszcza azotu i fosforu
dostęp tlenu jest konieczny do aktywnego poboru składników
światło
tlen
mikoryza
wzajemny wpływ kationów i anionów w roztworze glebowym
Sól fizjologicznie kwaśna – np. KCl – roślina pobiera K i wydziela H
+
i zakwasza środowisko.
Sól fizjologicznie zasadowa – np. NaNO
3
– roślina pobiera NO
3
-
i wydziela OH
-
, co powoduje alkalizację
środowiska.
Transport jonów zmienia się przy zmianie faz rozwojowych. Przy kiełkowaniu nie zachodzi pobieranie
składników mineralnych.
Mikoryza – rodzaj współżycia rośliny z grzybami. Występuje często u drzew, zbóż, tytoniu, pomidorów.
Strzępki grzyba oplatają korzeń, który nie tworzy włośników. Strzępki pobierają wodę wraz z różnymi
składnikami mineralnymi, zazwyczaj niedostępnymi dla roślin. Zwiększa się powierzchnia sorpcyjna.
Mikoryza:
ektodermalna
ektotroficzna
endotroficzna
Symbioza mikoryczna. Grzyby rozkładające celulozę pektyny umożliwiają storczykom kiełkowanie nasion i
wzrost na martwym podłożu organicznym. Ponadto, dostarczają asymilatów, dopóki roślina się nie
usamodzielni.
Fizjologia roślin
Wykłady
Created by Katilla
18
Symbioza z bakteriami brodawkowymi.
Co daje roślina bakteriom?
Proces redukcji NO
3
-
do NH
4
katalizuje reduktaza azotanowa i azotynowa.
Rhisobium, Azotobacter.
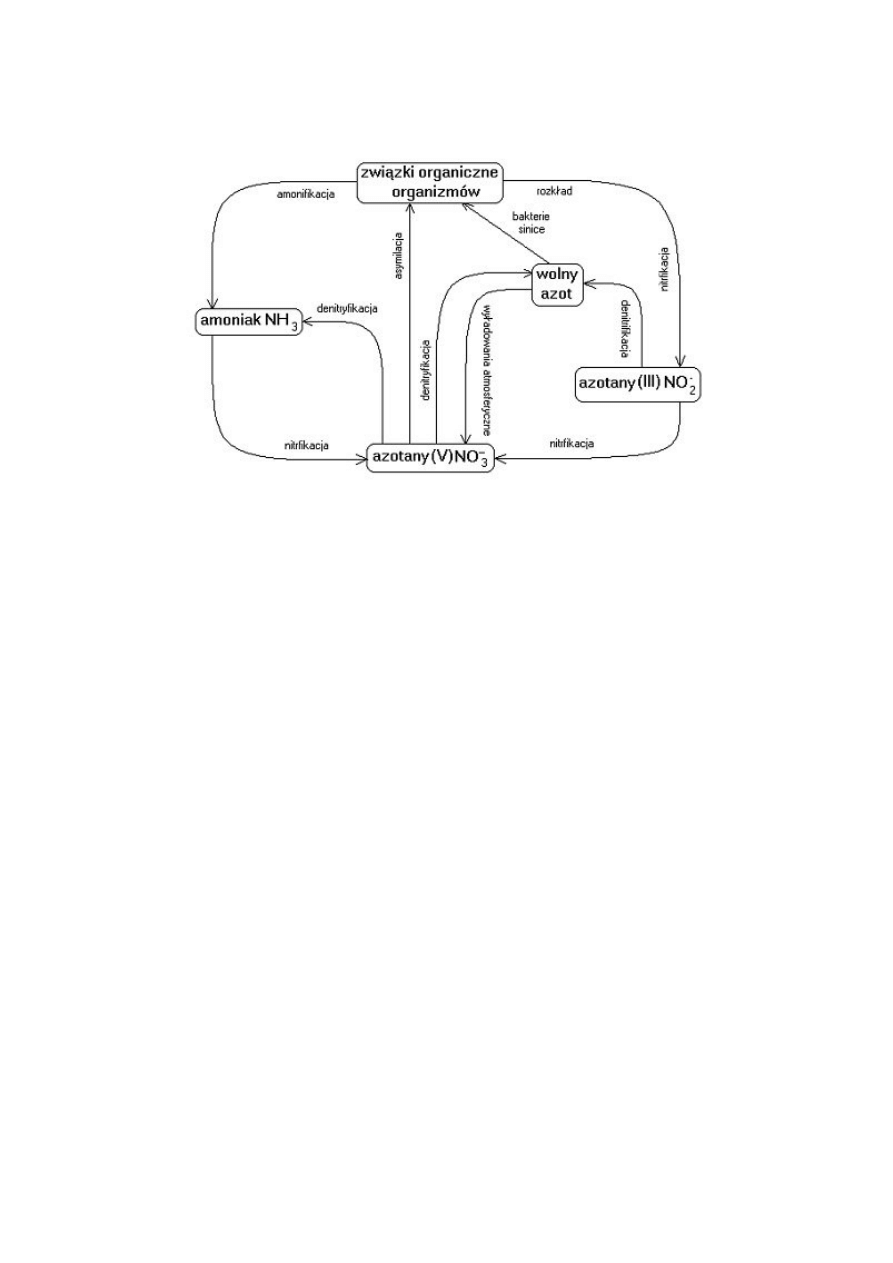
Udział bakterii w przemianach azotowych.
Amonifikacja – rozkład materii do amoniaku.
Nitryfikacja – przekształcenie amoniaku do azotanów.
Denitryfikacja – przekształcenie azotanów do amoniaku i wolnego azotu.
Antagonizm jonów:
Znoszenie toksycznego wpływu jednej soli przez drugą np. działanie KCl znoszone jest przez CaCl
2
K uwadnia cytoplazmę, Ca odwadnia, jony potasu zwiększają przepuszczalność błon
cytoplazmatycznych, wprowadzenie do cytoplazmy wapnia przywraca równowagę
Przykłady jonów antagonistycznych – Na
+
, Mg
2+
, NH
4
+
, Ca
2+
Plazmoliza wywołana wysokim stężeniem soli potasowej i wapniowej.
Obecność jednych jonów może hamować pobór innych jonów przez roślinę np. jony Al
3+
hamują pobieranie
jonów Cu
2+
, Ca
2+
hamuje pobór jonów Mn
2+
, a stymuluje absorpcję anionów. Duże stężenie NO
3
-
stymuluje
pobieranie K
+
. Duża aktywność Cl
-
zmniejsza pobór NO
3
-
.
Hamowanie pobierania jonów przez inne może być powodowana konkurencją o nośnik.
Prawa nawożenia:
Prawo minimum Libiga – pierwiastek, którego jest w podłożu najmniej, decyduje o plonie/
Prawo Mitscherlicha – każdy kg dostarczonego do gleby nawozu może dać różny efekt: -im mniej
danego składnika w glebie, a inne występują w ilości optymalnej, tym efekt nawożenia większy
(wyższy plon). -im tego składnika w glebie więcej, tym efekt nawożenia mniejszy.

Fizjologia roślin
Wykłady
Created by Katilla
19
Prawo maksimum– nadmiar składnika pokarmowego w glebie ogranicza skuteczność działania
innych i w następstwie powoduje obniżkę plonów.
Prawo pierwszeństwa wartości biologicznej – stosowanie nawozów musi mieć na celu nie tylko
wielkość plonu ale również wysoką jakość tego plonu.
Prawo zwrotu składników pokarmowych – aby utrzymać żyzność gleby należy jej zwracać
substancje przyswajalne, nie tylko pobrane przez rośliny i zebrane z plonem, ale również te które
znikają w glebie w formie przyswajalnej w następstwie stosowania dużych ilości nawozów
mineralnych.
Roztwór glebowy.
Wpływ metali ciężkich na rośliny
Metale ciężkie to pierwiastki o gęstości przekraczającej 6 g/ cm
3
. Obecnie silnie eksploatowane są do
środowiska, lecz nie ulegną biodegradacji.
Zn:
Niezbędny do produkcji auksyn, jest aktywatorem wielu enzymów, bierze udział w metabolizmie
azotowym i fosforowym
Nadmierne ilości powodują:
Zmniejszenie tempa wzrostu korzeni i liści
Hamują pobieranie przez rośliny fosforu i żelaza
Jego brak powoduje chlorozę liści (bielenie)
Cu:
Występuje w przenośnikach elektronów (oksydaza cytochromowa i askorbinowa, plastocyjanina)
Nadmierne ilości powodują:
Łączenie się z siarką – dezaktywacja białek – enzymów
Hamuje pobór żelaza
Zmienia strukturę białek
Co:
Aktywator enzymów
Inhibitor biosyntezy etylenu
Nadmiar powoduje chlorozę i nekrozę liści, wypiera z centrów aktywnych inne pierwiastki
Cd:
Ma duże powinowactwo do grup tiolowych enzymów (– SH) i białek zmieniając ich strukturę
Obniża zawartość chlorofilu
Hamuje wzrost korzeni
Wywołuje chlorozę i nekrozę liści, powodując przedwczesne ich opadanie
Pb:
Zmniejsza zawartość witaminy C i karotenu w roślinach
Wykazuje powinowactwo do błon cytoplazmatycznych
Wchodzi w interakcje z żelazem, miedzią i cynkiem
Wypiera z aktywnych centrów inne jony
Łączy się z grupami tiolowymi
Hamuje szereg procesów:

Fizjologia roślin
Wykłady
Created by Katilla
20
Transpirację
Fotosyntezę
Oddychanie
Metabolizm tłuszczów
Poprzez inaktywację szeregu enzymów
Hamuje powstawanie grupy hemowej, stąd też inhibituje syntezę chlorofilu
Al:
Powoduje zahamowanie wzrostu i grubienie korzeni
Zmniejsza liczbę korzeni
Czerwienienie łodyg
Zmniejszenie masy części nadziemnych
Przyłącza się do białek enzymatycznych, kalmoduliny, nukleotydów
Tworzy kompleksy z kwasami pektynowymi i białkami ściany komórkowej
Wypiera Cu
Nadmiar powoduje niedobór fosforu
Działa antagonistycznie do wapnia
Wpływa ujemnie na rozwój tkanek generatywnych
Gromadzi się głównie w tkankach merystematycznych i w strefie elongacyjnej

Fizjologia roślin
Wykłady
Created by Katilla
21
Fotosynteza
Fotosynteza – zmiana enzrgii świetlnej na energię wiązań chemicznych w chloroplastach.
Ogólne równanie fotosyntezy:
6 H
2
O + 6 CO
2
C
6
H
12
O
6
+ 6 O
2
Chloroplast:
Podwójna błona lipidowo – białkowa
grana – faza jasna
stroma – faza ciemna
swój genom – kolisty 120 – 160 tys. bp, koduje około 120 białek
błony tylakoidów mają dużo galaktolipidów
Chlorofil:
Atom Mg
2+
Układ porfirynowy
Ogon hydrofobowy – alkohol fitol
Barwniki karotenowe:
Barwniki pomocnicze
Np. β – karoten, ksantofile
Pochłaniają światło
Anteny
Funkcja ochronna
Chlorofil – barwnik, potrafi absorbować promieniowanie, a przez to zwiększać swoją energię wewnętrzną
→ chlorofil się wzbudza.
W błonach, albo na błonach znajdują się anteny i energia może być przekazana do centrum reakcji
fotochemicznej → produkty ATP lub zredukowanego NADP.
Chlorofil, który potrafi się wzbudzać, magazynuje energię i przekazuje ją do centrum reakcji
fotochemicznej. Najważniejsze chlorofile to – a i b.
Rośliny utleniając wodę produkują wodór, który jest paliwem – najważniejszy nośnik energii.
Rośliny wykorzystują CO
2
do zbudowania łańcucha węglowego, po to, aby do atomu węgla przyczepić
atom wodoru → magazynowanie wodoru.
Dalej w procesie oddychania tlenowego wodór łączy się z O
2
w sposób bezwybuchowy i tworzy się energia
(w reakcji utleniania).
Wodór jest najbardziej rozpowszechnionym paliwem we wszechświecie.

Fizjologia roślin
Wykłady
Created by Katilla
22
Gdy jest słońce → wodór gromadzony jest w łańcuchach węglowych
Gdy słońca brak → wodór łączy się z tlenem, po to, by uzyskać energię
Chlorofil występuje w błonach chloroplastowych lub trochę na zewnątrz. Jest on wiązany za pomocą białek
i w ten sposób powstaje kompleks LHC (kompleks zbierający światło).
Cząsteczki chlorofilu są wzbudzane i mogą dzięki temu transportować energię przez siebie. Proces ten jest
tym wydajniejszy im bardziej cząsteczki są do siebie zbliżone. Cząsteczki chlorofilu wiązane są z białkiem, bo
dzięki temu można je bardzo ciasno upakować.
Chlorofil:
Bardzo odporny na wysoką temperaturę
Nie rozpuszcza się w wodzie, ale rozpuszcza się w benzynie
Świeci na czerwono
Mniej energii w 2, aby była równowaga, bo długość fali jest większa.
ZASADA ZACHOWANIA ENERGII.
E
2
trwa dłużej niż E
1
E
1
– bardzo krótki
W ciemności można wygasić energetycznie chlorofil.
E
2
może być wykorzystywany przez rośliny.
Energia wzbudzenia jest przeznaczona na cele fotochemiczne
① transportowany do CR ⇨ reakcje fotochemiczne
→ ATP
→ NADP
② ciepło
③ fluorescencja – na ciemnoczerwono, bo fala jest dłuższa od jasnej czerwieni
W liściu zasób energii jest głównie pożytkowany na cel ①, w postaci fluorescencji ucieka 3 – 5% energii (jest
to za mało, by można było by zobaczyć czerwień).
Liść jest zielony bo przepuszcza zieleń, natomiast niebieski i czerwony pochłania.
Właściwości fizyczne chlorofilu, np. absorbancja, zależą od głowy chlorofilu.
Właściwości chemiczne chlorofilu, np. rozpuszczalność, zależą od ogona chlorofilu
Fizjologia roślin
Wykłady
Created by Katilla
23
Faza „ciemniowa
C3
C4
pochwa okołowiązkowa
słabo rozbudowana
silnie rozbudowana, zwykle
dwuwarstwowa
dimorfizm chloroplastów
brak
występuje
retikulum endoplazmatyczne
normalne
rozbudowane
peroksysomy
liczne
brak lub nieliczne
świetlny punkt wygasania
jest
brak
optimum temperaturowe
25
o
C
35
o
C
gradient stężenia CO
2
między
atmosferą i chloroplastami
średni
wysoki
gospodarka wodna
mało oszczędna
oszczędna
wtórny akceptor CO
2
brak
RuDP
zużycie ATP (liczba cząsteczek
na redukcję 1 cząsteczki O
2
)
3
4 – 5
akumulacja skrobi
normalna
wyłącznie w chloroplastach
komórek pochwy
okołowiązkowej
Wiązanie CO
2
u roślin typu CAM
Np. kaktusy, w dzień aparaty szparkowe są zamknięte, w nocy otwarte, w nocy wchodzi CO
2
podobnie jak
C4, tylko kwas jabłkowy gromadzi się w wakuoli, a w dzień do cytozolu i CO
2
włączone do cyklu Calvina w
chloroplastach.
Fotooddychanie
Przy aktualnym stężeniu O
2
i CO
2
w powietrzu aktywność karboksylazy Rubisco 2,5 razy przewyższa
aktywność oksygenazową. Gdyby stężenie oby tych gazów było podobne to aktywność oksygenazowa
Rubisco byłaby 80x niższa niż aktywność karboksylazowa i mamy rozkład O
2
.
Współdziałają chloroplasty, peroksysomy i mitochondrium.
Rozpad rybulozo – 1,5 – bisfosforan na kwas glikolowy do peroksysomu i kwas 3 – fosfoglicerynowy.
Ostatecznie do glicyny w peroksysomie i seryny w mitochondrium
Zabezpiecza przed nadmierną radiacją.
Wpływ czynników zewnętrznych i wewnętrznych na intensywność fotosyntezy:
1. Światło – rośliny wykorzystują do fotosyntezy do 3,5% C
3
i 7% C
4
światła padającego
Światło z zakresu PAR (400 – 700 nm)
Punkt kompensacyjny świetlny – takie natężenie światła, że zachodzi równowaga między
fotosyntezą a oddychaniem
Punkt wysycenia – takie natężenie światła, że maksymalna fotosynteza tylko u C3

Fizjologia roślin
Wykłady
Created by Katilla
24
Słabe światło – epistrofia – chloroplasty prostopadle do światła
Mocne światło – parastrofia – chloroplasty równolegle do światła
2. CO
2
0,0360 ppm – stężenie w atmosferze, za małe dla roślin bariery dla CO
2
:
Warstwa graniczna
Aparaty szparkowe
Dyfuzja w cieczy
Punkt kompensacyjny: C3 5 – 10 Pa, C4 0,2 – 0,5 Pa
Przy zbyt dużym stężeniu zamykają się aparaty szparkowe
3. Temperatura
6 – 50
o
C – zakres fotosyntezy
Wpływ temperatury ma charakter kompleksowy (r. enzymatyczne, r. fizyczne), bardziej zależna
faza ciemna, w fazie jasnej wpływa na płynność błony, gdy wzrośnie temperatura u C3 rośnie punkt
kompensacyjny CO
2
C4 mają optimum temperaturowe wyższe
Powyżej 40
o
C gwałtownie spada fotosynteza
Przy zbyt niskiej temperaturze ograniczona jest synteza cukrów, spadek płynności błony
4. Woda
Gdy roślina zabezpiecza się przed utratą wody to niższa fotosynteza.
5. Związki mineralne

Fizjologia roślin
Wykłady
Created by Katilla
25
Oddychanie
C
6
H
12
O
6
+ 6O
2
→ 6CO
2
+ 6H
2
O
Glikoliza
Dekarboksylacja pirogronianu
Cylk Krebsa
Łańcuch oddechowy
GLIKOLIZA
Substratami mogą być cukry złożone.
Cukier będący substratem glikolizy jest u roślin magazynowany najczęściej w postaci skrobi, zaś formę w
jakiej jest przewodzony i dostarczany do komórek, głównie stanowi sacharoza.
Z rozkładu skrobi powstaje glukoza lub glukozo – 1 – fosforan, w przypadku sacharozy – glukoza i fruktoza.
Wstępne przekształcenia prowadzą do powstania fruktozo – 1,6 – bisfosforanu, który następnie zostaje
rozłożony i utleniony do pirogronianu. Powstają przy tym z 1 cząsteczki heksozy 2 cząsteczki ATP i 2
cząsteczki NADH. Glikoliza odbywa się bez pobrania O
2
, jest więc to beztlenowa faza oddychania.
Jeśli za punkt wyjścia przyjmujemy glukozę, to ulega ona na początku fosforylacji z udziałem ATP do
glukozo – 6 – fosforanu, a następnie zostaje zamieniona na swój izomer fruktozo – 6 – fosforan, a ten
ponownie fosforylowany na fruktozo – 1,6 – bisfosforan. Reakcje są katalizowane przez odpowiednie
enzymy – heksokinazę, izomerazę glukozofosforanową i fosfofruktokinazę.
Rozerwanie wiązań pomiędzy 3 a 4 węglem fruktozo – 1,6 – bisfosforanu z udziałem aldolazy
fruktozobisfosforanowej prowadzi do powstania dwóch trioz: aldehydu 3 – fosfoglicerynowego i
fosfodihydroksyacetonu. Znajdują się one w dynamicznej równowadze, w której uczestniczy izomeraza
triozofosforanowa. W dalszych przemianach bierze udział aldehyd 3 – fosfoglicerynowy. Z udziałem
dehydrogenazy aldehydu 3 – fosfoglicerynowego i kinazy fosfoglicerynianowej zostaje utleniony do 3 –
fosfoglicerynianu (PGA). W trakcie tych przemian NAD zostaje zredukwowany do NADH oraz powstaje
ATP. Następnie fosfogliceromutaza i hydrataza fosfoglicerynianowa katalizują przekształcenie 3 –
fosfoglicerynianu w fosfoenolopirogronian (PEP). Ten zaś pod wpływem kinazy pirogronianowej
przechodzi w pirogronian – powstaje przy tym cząsteczka ATP.
Pirogronian przechodzi z cytosolu do mitochondrium i tam ulega dalszym przemianom i ostatecznie
utlenieniu do CO
2
i H
2
O – w warunkach tlenowych. W warunkach beztlenowych pirogronian może być
substratem dla procesów fermentacji.
FERMENTACJA
W warunkach niedotlenienia tkanek roślinnych (np. korzenie znajdujące się w glebie zalanej lub wysyconej
wodą) procesy oddychania komórkowego zostają zahamowane. Dotyczy to również utleniania NADH i
regeneracji NAD
+
. Gdy wyczerpie się zapas NAD
+
do utleniania aldehydu 3 – fosfoglicerynowego, może
nastąpić zahamowanie glikolizy. Metabolizm oddechowy zostaje wtedy skierowany na drogę fermentacji.
NAD
+
zostaje odtworzony dzięki użyciu NADH do redukcji pirogronianu do mleczanu lub aldehydu
octowego (powstałego po dekarboksylacji pirogronianu) do etanolu. W pierwszym przypadku reakcja jest
katabolizowana przez dehydrogenazę mleczanową, w drugim przez dekarboksylazę pirogronianową a
następnie dehydrogenazę alkoholową.

Fizjologia roślin
Wykłady
Created by Katilla
26
Tkanki roślinne mogą oddychać beztlenowo tylko przez ograniczony czas, gdyż produkty fermentacji, kwas
mlekowy i etanol, w większych stężeniach są szkodliwe dla tkanek. Zysk energetyczny w fermentacjach jest
niewielki i wynosi 2 cząsteczki ATP na 1 cząsteczkę rozłożonej glukozy.
OKSYDACYJNA DEKARBOKSYLACJA PIROGRONIANU
Dalsze procesy oddychania przebiegają w mitochondriach i w końcowej fazie związane są z pobraniem
tlenu.
Mitochondria, podobnie jak chloroplasty, otoczone są podwójną błoną. Błona zewnętrzna jest łatwo
przepuszczalna dla większości jonów i metabolitów komórkowych o masie poniżej 10 kDa. Natomiast błona
wewnętrzna, tworząca do środka głębokie wpuklenia, odznacza się brakiem przepuszczalności dla
większości jonów i organicznych cząsteczek. Istnieje w niej jednak specjalny nośnik, dzięki któremu
pirogronian może przeniknąć z cytosolu do wypełniającej wnętrze mitochondrium bezpostaciowej
macierzy, w której następuje jego oksydacyjna dekarboksylacja. Bierze w niej udział NAD
+
, koenzym A oraz
kompleks enzymów zwany oksydazą pirogronianową.
Dekarboksylacja pirogronianu uwalnia CO
2
, a powstały przy tym aldehyd octowy zostaje utleniony do
octanu (reszty acetylowej). Reszta acetylowa zostaje przyłączona do zawierającego grupę tiolową (-SH)
koenzymu A (HS – CoA) i powstaje acetylo – CoA (CH
3
CO – ScoA). Równocześnie NAD
+
zostaje
zredukowany do NADH.
CH
3
COCOO
-
+ HSCoA + NAD
+
→ CO
2
+ CH
3
CO – SCoA + NADH
Acetylo – CoA jest bardzo ważnym związkiem pośrednim przenoszącym grupy acetylowe. Wiązanie
tioestrowe w acetylo – CoA zawiera znaczną ilość energii, która zostaje wykorzystana podczas syntezy
cytrynianu. W ten sposób octan zostaje włączony do cyklu Krebsa.
CYKL KREBSA
1. Wytwarzanie cytrynianu ze szczawiooctanu i acetylo – CoA, które jest katalizowane przez
syntazęcytrynianową.
2. Izomeryzacja cytrynianu do izocytrnianu katalizowana przez akonitazę.
3. Utlenienie izocytrynianu do α – ketoglutaranu katalizowane przez dehydrogenazę
izocytrynianową, reakcja ta wymaga NAD
+
.
4. Utlenienie α – ketoglutaranu do bursztynylo – CoA, które jest katalizowane przez kompleks
dehydrogenazy α – ketoglutaranowej, reakcja ta wymaga NAD
+
.
5. Przekształcenie bursztynylo – CoA w bursztynian, które katalizowane jest przez syntetazę
bursztynylo – CoA, reakcja ta wymaga fosforanu nieorganicznego i GDP (lub ADP).
6. Utlenianie bursztynianu do fumaranu, które jest katalizowane przez dehydrogenazę
bursztynianową, w reakcji uczestniczy FAD.
7. Uwodnienie fumaranu do jabłczanu, które katalizowane jest przez fumarazę.
8. Utlenianie jabłczanu do szczawiooctanu, które katalizowane jest przez dehydrogenazę
jabłczanową, reakcja ta wymaga NAD
+
.
W procesach zachodzących w macierzy mitochondrialnej, tj. w wyniku oksydacyjnej dekarboksylacji
pirogronianu i cyklu Krebsa, z jednej cząsteczki pirogronianu powstają 3 cząsteczki CO
2
, 4 cząsteczki NADH,
1 cząsteczka FADH
2
i 1 cząsteczka ATP.

Fizjologia roślin
Wykłady
Created by Katilla
27
Niekoniecznie jednak cały węgiel zawarty w pierwotnym substracie oddechowym jest utleniany do CO
2
.
Organiczne szkielety produktów pośrednich glikolizy i cyklu Krebsa mogą być użyte do syntezy różnych
ważnych substancji jak np.:
wchodzący w skład związków tłuszczowych glicerol (z aldehydu fosfoglicerynowego),
związki fenolowe (z fosfoenolopirogronianu),
alanina (z pirogronianu),
kwasy tłuszczowe, flawonoidy (z acetylo – CoA)
glutaminian (z oksoglutaranu)
cytochromy, fotochrom (z sukcynylo – CoA)
asparaginian (z szczawiooctanu)
Gdy jednak pośrednie produkty cyklu Krebsa zostaną użyte do innych procesów, nie może zajśćregeneracja
szczawiooctanu. Cykl zostałby więc zahamowany. Zapobiega temu karboksylacja fosfoenolopirogronianu
dająca jako produkt szczawiooctan. Stanowi ona fragment procesu zwanego szlakiem jabłczanowym.
SZLAK JABŁKOWY
Reszta z Szymańskiej od strony 134.
Czynniki wpływające na procesy oddychania:
temperatura
stężenie O
2
stężenie CO
2
uwodnienie komórek
dostępność składników mineralnych
poziom substratów oddechowych – światło
uszkodzenia mechaniczne i czynniki chorobotwórcze
rodzaj i wiek rośliny
inhibitory i stymulatory wzrostu
Wpływ temperatury
Wykresy z ćwiczeń
Wpływ stężenia O
2
na oddychanie tlenowe i beztlenowe.
Wykres z ćwiczeń
Poniżej 10% O
2
– uruchomienie oddychania beztlenowego, spadek aktywności oddychania tlenowego
5% O
2
– przewaga oddychania beztlenowego nad tlenowym
2 – 3% O
2
– oddychanie beztlenowe – brak łańcucha oddechowego – bardzo mały zysk energetyczny
Wpływ stężenia CO
2
Nadmiar CO
2
hamuje oddychanie. Taki stan może wystąpić:
w glebie i wtedy zahamowane jest oddychanie w korzeniu
w nasionach z grubą łupiną i wtedy zahamowane jest ich kiełkowanie
Uwodnienie komórek a procesy oddechowe
Wykres z ćwiczeń

Fizjologia roślin
Wykłady
Created by Katilla
28
Woda jest potrzebna do procesów hydrolizy, do uwodnienia materiałów zapasowych.
Uszkodzenia mechaniczne i czynniki chorobotwórcze
Wzrost intensywności oddychania w uszkodzonych miejscach na skutek:
uruchomienia reakcji obronnych – zwiększenie zapotrzebowania na metabolity
intensywne podziały mitotyczne w celu zabliźnienia rany
ułatwionej wymiany gazowej
Wiek rośliny
Wykres z ćwiczeń
Podczas dojrzewania jabłek wydziela się etylen, który przyspiesza dojrzewanie poprzez stymulację
intensywności oddychania, w wyniku czego owoce miękną i nabierają smaku. Jest to zjawisko negatywne
dla roślin.
Współczynnik oddechowy (RQ) – stosunek objętościowy ilości wydzielonego CO
2
do ilości pobranego O
2
.
RQ = CO
2
/O
2
Na wartość RQ może mieć wpływ:
rodzaj badanego organu
faza rozwojowa
rytmika dobowa
gatunek rośliny
Gdy RQ < 1 – utlenienie glukozy jest niezupełne (np. w mięsistych owocach w wyniku gromadzenia się
kwasów organicznych)
Gdy RQ > 1 – występują warunki organiczonego dostępu tlenu
Fizjologia roślin
Wykłady
Created by Katilla
29
Wzrost – regulatory wzrostu
AUKSYNY
Występują w tkankach w niewielkich ilościach. Zależnie od stężenia mogą działać stymulująco lub
hamująco.
Biosynteza w stożkach wzrostu, pąkach, liściach, owocach ⇨ wszystkie merystemy nadziemne
Prekursor – tryptofan
Auksyny są transportowane polarnie do cząsteczki korzenia (auksyny mają grupę – COOH, która dysocjuje i
auksyna staje się anionem).
Auksyny:
Przyspieszają wzrost elongacyjny
Wspólnie z cytokinami uczestniczą w dominacji pąka szczytowego
Przyspieszają ukorzenianie się sadzonek pędów zielnych i zdrewniałych
Biorą udział w inicjacji kwitnienia – głównie w okresie post indukcyjnym poprzez przyspieszenie
organogenezy kwiatów
Wpływają na powstawanie warstewki odcinającej przy liściach i owocach w zależności od stężenia
Biorą udział w wywoływaniu tropizmu u roślin
U roślin pomidora, ogórka, truskawki wywołują zjawisko partenokarpii
Wspólnie z cytokininami uczestniczą w organogenezie w kulturach in vitro (cytokiny < auksyny –
wzrost korzenia, cytokiny > auksyny – rozwój pędu)
Zarówno namnażanie jak i wydłużanie komórek zależne od auksyn.
Działanie auksyn na wzrost wydłużeniowy:
Bezpośrednie
(wzrost szybki)
stymulacja pobierania K
+
i wydzielania H
+
(pompa protonowa)
↓
osmotyczne pobieranie H
2
O
↓
wzrost komórek
(reakcja natychmiastowa, max 1 h)
pośrednie
(wzrost powolny, poprzez aparat genetyczny)
aktywacja DNA
↓
synteza mRNA
↓
synteza białek enzymatycznych
↓
synteza składników ścian komórkowych
↓
wzrost komórek
(reakcja opóźniona, po 2 h)
IAA
IAA
receptor

Fizjologia roślin
Wykłady
Created by Katilla
30
GIBERELINY
Stymulują tylko wzrost wydłużeniowy komórek.
Przekursorem jest acetylo – CoA.
Biosynteza w dojrzewających nasionach, owocach, pręcikach kwiatów, częściach wierzchołkowych korzeni,
w najmłodszych liściach z pominięciem merystemów.
Gibereliny:
Przyspieszają wzrost całych roślin
Przyspieszają kiełkowanie nasion
Indukcja kwitnienia u roślin długiego dnia
Skracają cykl rozwojowy roślin dwuletnich do jednego roku
U niektórych roślin zastępują zabieg termoindukcji kwitnienia
Skracają okres naturalnego spoczynku roślin
Wspólnie z cytokininami hamują starzenie się roślin
Mają wpływ na spoczynek drzew
Partenokarpia u brzoskwini, jabłoni, winorośli i innych pestkowych
CYTOKININY
Wszystkie naturalne cytokininy są pochodnymi adeniny i pirofosforanu izopentynylu.
Biosynteza w wierzchołkach wzrostu korzeni (biosynteza w merystemach rozwijających się owoców,
pąków, kiełkujących nasion, kambium nie jest wykluczona).
Degradacja – oksydacyjne oderwanie reszty izopentenylowej i właczenie adeniny do metabolizmu.
Wpływ cytokinin na komórkę:
Przyspieszenie podziałów komórkowych i powiększanie się komórek
Wpływ na wzrost roślin (rola w organogenezie i dominacji wierzchołkowej – stymulacja rozwoju
pąków bocznych)
Hamowanie starzenia się roślin (hamowanie rozpadu chlorofilu)
Stymulowanie kiełkowania nasion niektórych gatunków roślin
Skracanie stanu spoczynku roślin
ETYLEN
Powstawanie z metioniny. Przekształcenia do kwasu l – aminocyklopropano – 1 – karboksylowego (ACC) –
bezpośredni prekursor i forma transportowa etylenu.
Zdolność do biosyntezy w każdej żywej komórce.
Etylen:
Hamuje wzrost wydłużeniowy komórek
Przyspiesza dojrzewanie owoców
Przyspiesza starzenie się roślin:
Przyspiesza rozkład chlorofilu
Przyspiesza oddychanie owoców klimakterycznych, a przez to skraca czas
przechowywania
Przyspiesza powstawanie warstewki odcinającej podobnie jak ABA
Obniża turgor płatków korony kwiatów, powodując ich więdnięcie i zamieranie
Stymuluje rozwój słupkowia podobnie jak auksyny
Przyspiesza ukorzenianie się sadzonek poprzez stymulację organogenezy korzeni przybyszowych
Stymuluje rozwój roślin cebulowych

Fizjologia roślin
Wykłady
Created by Katilla
31
Stymuluje kiełkowanie pyłku
Stymuluje organogenezę kwiatów żeńskich u dyniowatych
KWAS ABSCYSYNOWY (ABA)
Biosynteza z acetylo – CoA poprzez kwas mewalonowy.
Droga pośrenia z karotenoidów – prekursor wiolaksantyna.
Biosynteza w pąkach, dojrzałych liściach, łodygach, nasionach, dojrzałych owocach, korzeniach.
Wpływ ABA na rośliny:
Hamuje podziały komórkowe i wzrost wydłużeniowy
Przyspiesza powstawanie warstewki odcinającej, a zatem opadanie liści i owoców
Uczestniczy w zamykaniu się aparatów szparkowych poprzez przyspieszanie wypływu jonów K
+
w
komórkach przyszparkowych
Wpływa na spoczynek nasion i pąków
Hamuje aktywność polimerów DNA i RNA
Hamuje aktywność α – amylazy w endosporium ziarniaków zbóż – w opozycji do giberelin
Testy do wykrywania ABA:
Test odpadania ogonków liściowych bawełny
Test hamowania wzrostu koleoptyli pszenicy
Test aparatów szparkowych
KWAS JASMONOWY
Syntetyzowany z nienasyconych kwasów tłuszczowych
Reguluje wzrost, kiełkowanie, starzenie
Pojawia się przy działaniu stresów
Cząsteczka sygnałowa sygnalizująca patogenezę
Hormony podobne w budowie do hormonów płciowych zwierzęcych:
Budowa podobna do hormonów płci
Głównie działają na wzrost wydłużeniowy
Powodują rozluźnienie ściany komórkowej
Nazwa brasinolidy lub brasinosteroidy
KWAS SALICYLOWY
Odpowiedzialny za termoregulację
Może indukować kwitnienie niektórych roślin
Infekcja pojedynczego liścia:
1. Akumulacja kwasu salicylowego → przekazywanie sygnału przez floem → indukcja odporności
systemicznej
2. Synteza i uwalnianie estru metylenowego kwasu salicylowego → przekazywanie sygnału do innych
organów lub sąsiednich roślin → indukcja odporności
POLIAMINY
Główne to putrescyna, spermidyna i spermina
Regulatory wzrostu u roślin
Wtórne przekaźniki

Fizjologia roślin
Wykłady
Created by Katilla
32
Występują we wszystkich etapach rozwoju rośliny
Indukują wzrost
Wpływają na polimerazę RNA
Działają na podziały komórkowe
Indukują powstawanie pąków bocznych
TRIAKONTANOL
30 węglowodorowy alkohol
Stymuluje wzrost
Stymuluje kwitnienie u niektórych roślin
Wtórny regulator
Nietoksyczny
TRAUMATYNA
Hormon przyranny
Stymuluje procesy zaleczania
RETARDANTY
Zawierają azot często w pierścieniach
Wykorzystywane przy produkcji roślin o zmniejszonych rozmiarach
Efekty działania:
Związki hamujące wzrost wydłużeniowy łodygi
Pogrubienie łodygi i zwiększenie zawartości tkanek mechanicznych
Hamują starzenie
Wzrost zawartości białek, chlorofilu i składników mineralnych w częściach nadziemnych
Obniżanie pobierania wody
Zwiększenie transportu asymilatów
Wzrost odporności na stresy
Wzrost odporności na grzyby
Wzrost pobierania składników pokarmowych z gleby
Fizjologia roślin
Wykłady
Created by Katilla
33
Rozwój roślin
Rozwój – pojęcie jakościowe, wszystkie zmiany ukierunkowane genetycznie, nieodwracalnie, zachodzą
przez cały cykl życia organizmu, obejmuje wzrost, różnicowanie komórek, starzenie komórek, śmierć.
Komórki nie raz w czasie różnicowania się ulegają starzeniu.
Regulacja cyklu komórkowego:
W interfazie fazy G
1
, S, G
2
W G
1
następuje biosynteza białek i kwasów rybonukleinowych
W S biosynteza DNA (podwojenie chromatyny)
W G
2
synteza białek
I faza rozwoju rośliny jako całości jest embriogeneza, która zachodzi w nasionach, powstaje zawiązek
rośliny mający genom od obydwu rodziców, z zygoty powstaje organizm.
typ mutacji
delecje części
gurke
apikalnej
fackel
centralnej
monopteros
bazalnej
gnom
terminalnych
Mutacje zachodzące podczas wczesnych faz embriogenezy rzodkiewnika.
Wzrost – przyczyną pobierania wody, gromadzenie się w wakuolach i ciśnięcie wakuoli na ściany w wyniku
czego rozbudowują się systemicznie.
Głównym czynnikiem hormonalnym jest auksyna i trochę giberelina, geny odpowiedzialne to geny związane
z wapniem
Auksyna – białko w ścianie komórkowej, powoduje rozluźnienie ściany struktury ściany komórkowej, gen
kodujący biorący udział w regulacji
Kiełkowanie
W nasieniu – zarodek (materiał genetyczny od rodziców), materiał zapasowy w bielmie lub liścieniach,
łupina nasienna.
3 fazy:
1. Fizyczna (imbicji)
2. Biochemiczna (kataboliczna)
3. Fizjologiczna (anaboliczna)
Ad. 1. Gwałtowne pobieranie wody przez koloidy białkowe i polisacharydy
Ad. 2. Materiały zapasowe hydrolizują, pobieranie wody na stałym poziomie
Ad. 3. Pobieranie wody wzrasta, bo pojawiły się związki osmotycznie czynne
Czynniki zewnętrzne regulujące kiełkowanie:

Fizjologia roślin
Wykłady
Created by Katilla
34
Temperatura
Światło – nasiona fotoblastyczne – wymagają światła aby zacząć kiełkować
Tlen
Dwutlenek węgla – za wysokie stężenie hamuje kiełkowanie
Żywotność nasion – określa zdolność do kiełkowania
Długowieczność nasion – najstarsze nasiona łubinu z lodowców sprzed 10 000 lat
Faza siewki
Rozpoczyna się samodzielne życie rośliny, okres młodociany, roślina nie ma zdolności kwitnienia,
intensywny wzrost, silna reakcja na bodźce tropiczne, duża zdolność regeneracyjna, nabiera właściwości
spolaryzowania, rozwija się korzeń, formowanie blaszki liściowej i aparaty fotosyntetyczne, wzrasta
właściwość na regulację fitochormonalną
Regulacja:
hormonalna – głównie hormony stymulujące wzrost (auksyny, gibereliny, cytokininy)
genetyczna – TMM – zjawisko spowodowane mutacją genu regulującego liczbę aparatów
szparkowych,
czynniki środowiskowe
światło przez fitochromy formy P
R
i P
FR
, temperatura (minimalna, optymalna, maksymalna)
woda
skład chemiczny gleby
Korelacje inhibicyjne – wzrost jednych organów hamuje wzrost drugich. Inhibicja między liściami i
owocami.
Czas uwarunkowany genetycznie.
Faza generatywna
Hormonem kwitnienia są gibereliny u niektórych roślin.
Przejście w fazę generatywną warunkowaną czynnikami środowiskowymi
Światło
(długość trwania, jakość)
⌇
Fitochrom
↓
Łańcuch transdukcji sygnału
↓
Zmiany metaboliczne
↓
Ekspresja genów
↓
Stan indukcji
Induktor kwitnienia
↓
Transport induktora
↓

Fizjologia roślin
Wykłady
Created by Katilla
35
Wierzchołek wzrostu
Ewokacja
Schemat przebiegu fotoperiodycznej indukcji kwitnienia
SDP – rośliny dnia krótkiego
LDP – rośliny dnia długiego
DNP – rośliny nie zależne od długości dnia
Rośliny dnia krótkiego
Termoindukcja
Wernalizacja – u wielu roślin okresowe działanie niskiej temperatury jest konieczne, aby mogło dojść do
kwitnienia, lub przynajmniej niska temperatura przyspiesza kwitnienie.
Działanie chłodu ma charakter bodźcowy, dlatego nazywamy go termoindukcją.
Termoindukcja zachodzi pod wpływem dłuższego działania (3 – 8 tygodni) temperatur wynoszących od 0 –
10
o
C. Jeśli w tym czasie temperatura ulegnie wzrostowi (np. do ok. 35
o
C), efekt termoindukcji nie pojawi się
i wystąpi dewenalizacja.
Wrażliwość roślin na okresowe działanie chłodu jest bardzo różna. Pełną niewrażliwość wykazują rośliny
jednoroczne, u niektórych roślin wernalizacja powoduje przyspieszenie kwitnienia, lecz są i takie, u których
kwitnienie całkowicie od niej zależy.
Rośliny różnią się fazą rozwojową, w której są wrażliwe na wernalizację:
Rośliny ozime – mogą przejść wernalizację w stadium spęczniałego nasienia. W środowisku
naturalnym ich nasiona lub ziarniaki kiełkują wczesną jesienią i już jako siewki przechodzą zimą
wernalizację i kwitną w następnym sezonie wegetacyjnym
Rośliny dwuletnie – są wrażliwe na bodziec wernalizacyjny dopiero po osiągnięciu pełnego cyklu
wegetatywnego. W pierwszym roku wegetacji tworzą zwykle dobrze rozwinięty krótki pęd z gęsto
osadzonymi liśćmi (np. kapusta, burak) i w takiej postaci zimują przechodząc wernalizację. W
następnym roku zaś tworzą pęd kwiatonośny i kwiaty.
Wernalizacja
↓
Wierzchołek wzrostu
↓

Fizjologia roślin
Wykłady
Created by Katilla
36
Przemiany stanu błon komórkowych
↓
Przemiany metaboliczne i hormonalne
↓
Zmiany wrażliwości wierzchołka wzrostu
↓
Demetylacja organów
↓
Ekspresja genów kwiatowych
↓
Morfogeneza kwiatu
Nie jest dziedziczna, podczas wzrasta stężenie hormonów młodości, gibereliny mogą wystąpić u niektórych
roślin, α – tokoferol też może zastąpić
Rośliny monokarpiczne – faza generatywna tylko raz w życiu
Alikarpiczne – faza generatywna występuje wiele razy
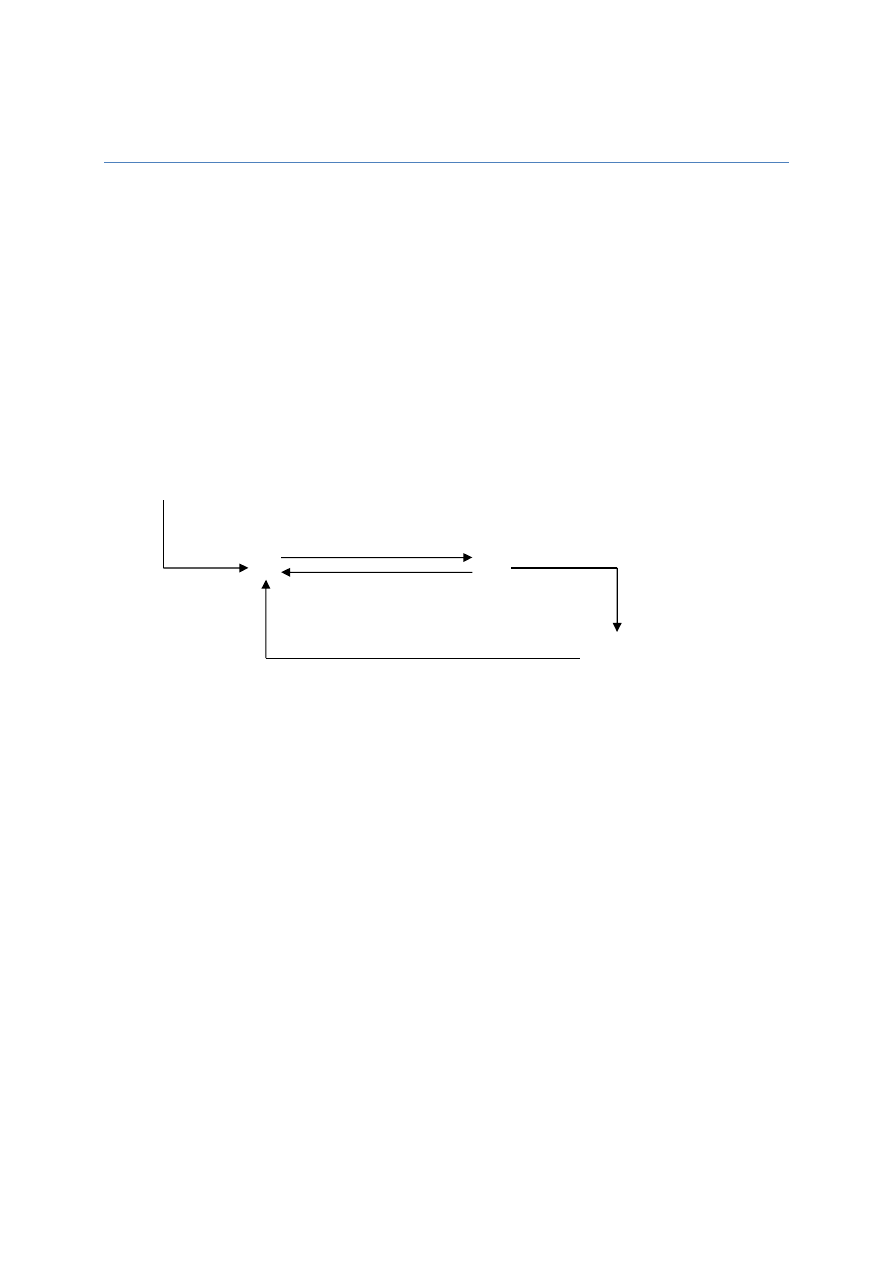
Fizjologia roślin
Wykłady
Created by Katilla
37
Ruchy roślin i allelopatia
Fotomorfogeneza – niezależny od fotosyntezy, bezpośredni wpływ światła na rozwój roślin (rośliny
odczuwają wpływ światła o długości 300-800nm)
Etiolacja – wzrost rośliny w ciemności, synteza chlorofilu zatrzymana na etapie bezbarwnego
protochlorofilidu A
Deetiolacja – po wystawieniu na światło następuje synteza chlorofilu
Fotoreceptory roślin:
Fitochromy
Kryptochromy
Fototropiny
FITOCHROM
Procesy regulowane przez fitochrom:
Indukcja kiełkowania nasion fotoblastycznych
Wzrost wydłużeniowy pędu
Rozwój blaszek liściowych
Indukcja kwitnienia
Tworzenie organów zapasowych
Synteza chlorofilów i barwników antocyjanowych
Sezonowość wzrostu
Rytmy dobowe
Kryptochromy:
długość światła 370-500nm
hamowanie wzrostu wydłużeniowego hipokotyli
akumulacja antocyjanów
indukcja kwitnienia
Fototropina:
Długość światła 340 – 525 nm
Fototropizm – ruch wzrostowy pod wpływem bodźca świetlnego
Hamowanie wzrostu pędu
Synteza w ciemności
P
r
P
Fr
Powolny rozpad
Światło białe lub czerwone 660nm
daleka czerwień 730nm
powolna przemiana w ciemności

Fizjologia roślin
Wykłady
Created by Katilla
38
Spoczynek – zahamowanie wzrostu i silne ograniczenie aktywności życiowej przy pełnym zachowaniu
zdolności do życia
Spoczynek bezwzględny – przyczyna wewnątrz organizmu: wysoki stosunek ABA i niskie stężenie
giberelin(stratyfikacja); u nasion okrywa nasienna nieprzepuszczalna dla wody i gazów (skaryfikacja) oraz
niedojrzałość morfologiczna zarodka
Spoczynek względny – spoczynek narzucony, przyczyna na zewnątrz organizmu, np. brak wody, niska
temperatura, brak tlenu, brak światła, nadmiar CO
2
, obecność związków toksycznych (u nasion), u roślin
wieloletnich "hamowanie korelacyjne" gdy obecne są liście na drzewie
Formy spoczynku:
spoczynek całego organizmu
spoczynek organu
spoczynek zarodka w nasieniu
Skaryfikacja – zabieg mający na celu przerwanie spoczynku bezwzględnego nasion, wywołany
nieprzepuszczalną dla wody i gazów okrywą nasienną (uszkodzenie okrywy nasiennej; skaryfikacja
mechaniczna, chemiczna, termiczna)
Stratyfikacja – zabieg mający na celu przerwanie spoczynku bezwzględnego nasion, wywołany dużym
stężeniem inhibitorów lub małym stymulatorów kiełkowania (przetrzymywanie nasion w wilgotnym,
sterylnym podłożu w temperaturach chłodu)
Rośliny wyższe nie mają możliwości przemieszczania się
Występują ruchy organów, cytoplazmy, plastydów
Ruchy wywołane są bodźcami ukierunkowanymi i nieukierunkowanymi
Ruchy lokomotoryczne
Bodziec
Tropizm
Nastia
Światło
Fototropizm
Fotonastia
Ciążenie
Geotropizm
-
Bodziec chemiczny
Chemotropizm
Chemonastia
Woda
Hydrotropizm
-
Temperatura
Termotropizm
Termonastia
Dotyk
Tigmotropizm
Sejsmo-tigmonastia
Rytm dobowy
-
Nyktinastia
Zranienie
Traumatropizm
-
Ładunek elektryczny
Elektrotropizm
-
Ruchy higroskopowe:
Zależą od stopnia uwodnienia tkanek
Wysychanie pewnych owoców powoduje ich pękanie
Allelopatia – wzajemne oddziaływanie roślin wyższych na siebie. Może być ujemne lub dodatnie.
Jest to biochemiczne oddziaływanie w obrębie wszystkich klas roślin (Hans Molish 1937).
Wszystkie organy danej rośliny mogą mieć wpływ na inne rośliny.

Fizjologia roślin
Wykłady
Created by Katilla
39
Związki allelopatyczne = związki allelochemiczne = allelozwiązki = blastokoliny
Zależność pomiędzy stężeniem związku allelopatycznego, a reakcją wzrostową rośliny:
Wykres!!!
Związki allelopatyczne, tak jak inne związki biologicznie czynne, występujące w niskich stężeniach wykazują
działanie stymulujące, natomiast w wysokich stężeniach są inhibitorami.
Sposoby wydzielania związków z roślin:
Ulatnianie
Ługowanie
Eksudacja
Rozkład resztek roślin
Zmiany wywołane przez allelozwiązki:
Zmiana potencjału chemicznego
Modyfikacja kanałów jonowych i aktywności ATPaz
Peroksydacja lipidów
Obniżenie potencjału wody
Zaburzenia transportu elektronów w chloroplastach i mitochondriach
Hamowanie fosforylacji
Hamowanie mitozy i elongacji komórek
Zmiany aktywności enzymów
Generowani reaktywnych form tlenu
Hamowanie wzrostu
Spadek intensywności transpiracji
Hamowanie fotosyntezy i oddychania
Ograniczenie pobierania i transportu wody i składników mineralnych
Deformacje morfologiczno – anatomiczne w strefach wzrostu
Przykłady pozytywnego oddziaływania:
Wpływ kąkolu polnego czy chabra bławatka na rozwój pszenicy
Wpływ kompostu z roślin na kiełkowanie nasion
Przykład negatywnego oddziaływania:
Wpływ orzecha włoskiego na wzrost roślin w sąsiedztwie
Wpływ maku polnego czy ostrożnia polnego rozwój pszenicy
Juglon – pierwszy udokumentowany związek allelochemiczny charakterystyczny dla orzecha włoskiego .
Znajduje się we wszystkich zielonych częściach rośliny (liście, okrywy orzechów, kwiatostany). Juglon
słabiej działa na jeżynę.
Zastosowanie:
Allelozwiązki są składnikiem proekologicznych herbicydów, które łatwo degradują w glebie
Są składnikiem preparatu do stymulacji i rozwoju roślin
Uprawa współrzędowa (rośliny uprawne i korzystnie na nie działające)

Fizjologia roślin
Wykłady
Created by Katilla
40
Tworzenie kompostów i przyorywanie resztek pożniwnych może mieć zarówno korzystne jak i
negatywne działanie, zależnie od gatunku rośliny
Następstwo roślin po sobie (płodozmian) – należy dobierać gatunki tak, by nie miały na siebie
negatywnego wpływu
Pietruszka
Apiol
Cytryna
Limonen, cytral
Pomarańcza
Limonen, cytral
Cebula
Glikozydy siarkowe
Chrzan
Olejki gorczyczne

Fizjologia roślin
Wykłady
Created by Katilla
41
Reakcja roślin na czynniki stresowe
Stres to stan ogólnego „napięcia” w organizmie, w którym na skutek działania niesprzyjających czynników
środowiska, może dojść do upośledzenia albo ograniczenia metabolizmu lub rozwoju.
Czynnikiem stresowym może być:
Zbyt niska lub wysoka temperatura
Brak lub nadmiar wody
Nadmierne zasolenie lub brak soli mineralnych w podłożu
Nieodpowiednia ilość lub jakość promieniowania
Wiatr
Środki chwastobójcze lub owadobójcze
Atak patogenów, owadów, zwierząt
Istnieją w roślinach mechanizmy obronne przed działaniem czynników stresowych. Odporność na stres
może być:
Konstytutywna – polegająca na trwałym wyposażaniu rośliny w cechy chroniącymi przed
negatywnym wpływem środowiska
Indukowana - czyli cechy, które pojawiają się w roślinie w odpowiedzi na czynnik stresowy
Odporność konstytutywna, np.:
Kseromorficzna budowa chroniąca przed skutkami suszy
Obecność w tkankach substancji antybiotycznych (alkaloidów, toksycznych glikozydów, związków
fenolowych) chroniąca przed atakami patogenów
Wytwarzanie związków chemicznych, których nie znoszą zwierzęce szkodniki (owady, zwierzęta
roślinożerne)
Odpowiednia budowa morfologiczna, np. wytwarzanie kolców, cierni, która chroni przed
roślinożercami
Zakończenie cyklu rozwojowego przed nastaniem niekorzystnej pory roku:
Efemerydy – kończą cykl rozwojowy przed nastaniem lata
Rośliny jednoroczne – kończą cykl rozwojowy przed nastaniem zimy
W uruchamianiu systemu obronnego często biorą udział niskocząsteczkowe sygnały przekaźnikowe w
postaci np. hormonów – kwas abscysynowy, kwas jasmonowy, etylen. W licznych przypadkach stwierdzono
przy tym szybką zmianę ekspresji genów i pojawienie się w komórkach roślinnych nowych specjalnych
białek stresowych, których skład bywa częściowo podobny dla niektórych rodzajów stresów.
Gdy stres nie jest zbyt silny, czyli nie prowadzi do szybkiej śmierci rośliny, wzrasta w roślinie odporność na
działanie danego czynnika stresowego, czyli następuje „hartowanie” rośliny zwane również aklimatyzacją.
CHŁÓD
Chłód to temperatura w zakresie 0 – 10
o
C.

Fizjologia roślin
Wykłady
Created by Katilla
42
WOLNE RODNIKI
Wolny rodnik to atom lub cząsteczka posiadająca na orbitalu walencyjnym nie sparowany elektron.
Właściwość ta powoduje znaczną reaktywność i zdolność „atakowania” różnorodnych składników żywej
komórki.
Do reaktywnych form tlenu (RFT) należą:
Wolne rodniki:
Anionorodnik ponadtlenkowy O
2
⦁-
Rodnik wodorotlenkowy HO
2
⦁-
Rodnik hydroksylowy OH
⦁
Związki wykazujące znaczną reaktywność lub łatwo ulegające przekształceniom w wolne rodniki:
Tlen singletowy
1
O
2
Ozon O
3
Nadtlenek wodoru H
2
O
2
RFT powstają w żywej komórce podczas naturalnych biochemicznych procesów oksyredukcji (np.
oddychanie, fotosynteza) jako wynik kolejnych stopni redukcji (jedno -, dwu -, trójelektronowej) cząsteczki
tlenu lub mogą być generowane w wyniku oddziaływania różnorodnych czynników abiotycznych i
biotycznych.
Czynniki generujące RFT:
Stresy środowiskowe:
Stres wodny, zasolenia, niskiej temperatury
Światło
Oddziaływanie metali ciężkich
Uszkodzenia mechaniczne
Stosowanie pestycydów
Inwazja patogenów
Czynniki pozbiorcze:
Warunki zbioru
Warunki przechowywania
Warunki wstępnej obróbki technologicznej
RFT reagują ze składnikami organizmów żywych, co może wywołać niepożądane lub groźne skutki.
Szczególnie niebezpieczne są reakcje RFT z podstawowymi strukturami komórkowymi (białka, błony
lipidowo – białkowe, kwasy nukleinowe).
O
2
H
2
O
OH
⦁
O
2
⦁-
H
2
O
2
HO
2
⦁-
1
O
2
e
-
e
-
H
+
e
-
H
+
e
-
2H
+

Fizjologia roślin
Wykłady
Created by Katilla
43
Tkanki roślinne posiadają sprawnie funkcjonujący system obrony przed reaktywnymi formami tlenu, który
polega na:
Udziale enzymów neutralizujących RFT – dysmutaza ponadtlenkowa, katalaza, peroksydaza
Interakcji RFT z antyoksydantami:
antyoksydanty hydrofilowe, np. kwas askorbinowy, związki fenolowe, glutation
antyoksydanty hydrofobowe, np. karotenoidy, tokoferole
Wyszukiwarka
Podobne podstrony:
Fizjologia roślin wykłady, Biologia, fizjologia roślin
Fizjologia roslin Wyklady gotowe
Fizjologia roślin - wykład 4 - 19.03.2013, Ogrodnictwo, Semestr IV, Fizjologia, Fizjologia egzamin,
FIZJOLOGIA ROŚLIN wykład 5
fizjologia roślin - wykład (8.01.2014), Semestr III, Fizjologia Roślin, Wykłady
fizjologia roslin wyklad, Rok III, Rok II, Semestr III, Fizjologia roslin
Fizjologia roślin (18.12.2013), Semestr III, Fizjologia Roślin, Wykłady
Zagad.Biologia 2011, I rok, Fizjologia roślin - wykłady, Fizjologia roślin
Fizjologia roślin - wykład II - semestr IV - 05.02.2013, Ogrodnictwo 2011, Fizjologia roslin
Fizjologia roślin - wykład (27.11.2013), Semestr III, Fizjologia Roślin, Wykłady
Fizjologia roślin - wykład I - semestr IV - 26.02.2013, Ogrodnictwo 2011, Fizjologia roslin
Fizjologia roślin - wykład (04.12.2013), Semestr III, Fizjologia Roślin, Wykłady
FIZJOLOGIA ROŚLIN wykład 3
FIZJOLOGIA ROŚLIN wykład 4
FIZJOLOGIA ROŚLIN wykład 1
FIZJOLOGIA ROŚLIN wykład 2
Fizjologia roślin wykłady, Biologia, fizjologia roślin
wykład 6 fizj roślin, biologia, fizjologia roślin
wyklad 4 fizj roślin, biologia, fizjologia roślin
więcej podobnych podstron